Antioxidant, Anti-Inflammatory, and Microbial-Modulating Activities of Essential Oils: Implications in Colonic Pathophysiology


 ,
, 
Abstract
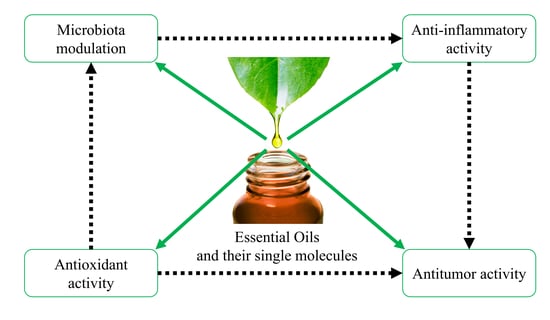
:
1. Introduction
2. EOs as Intestinal Anti-Inflammatory Agents
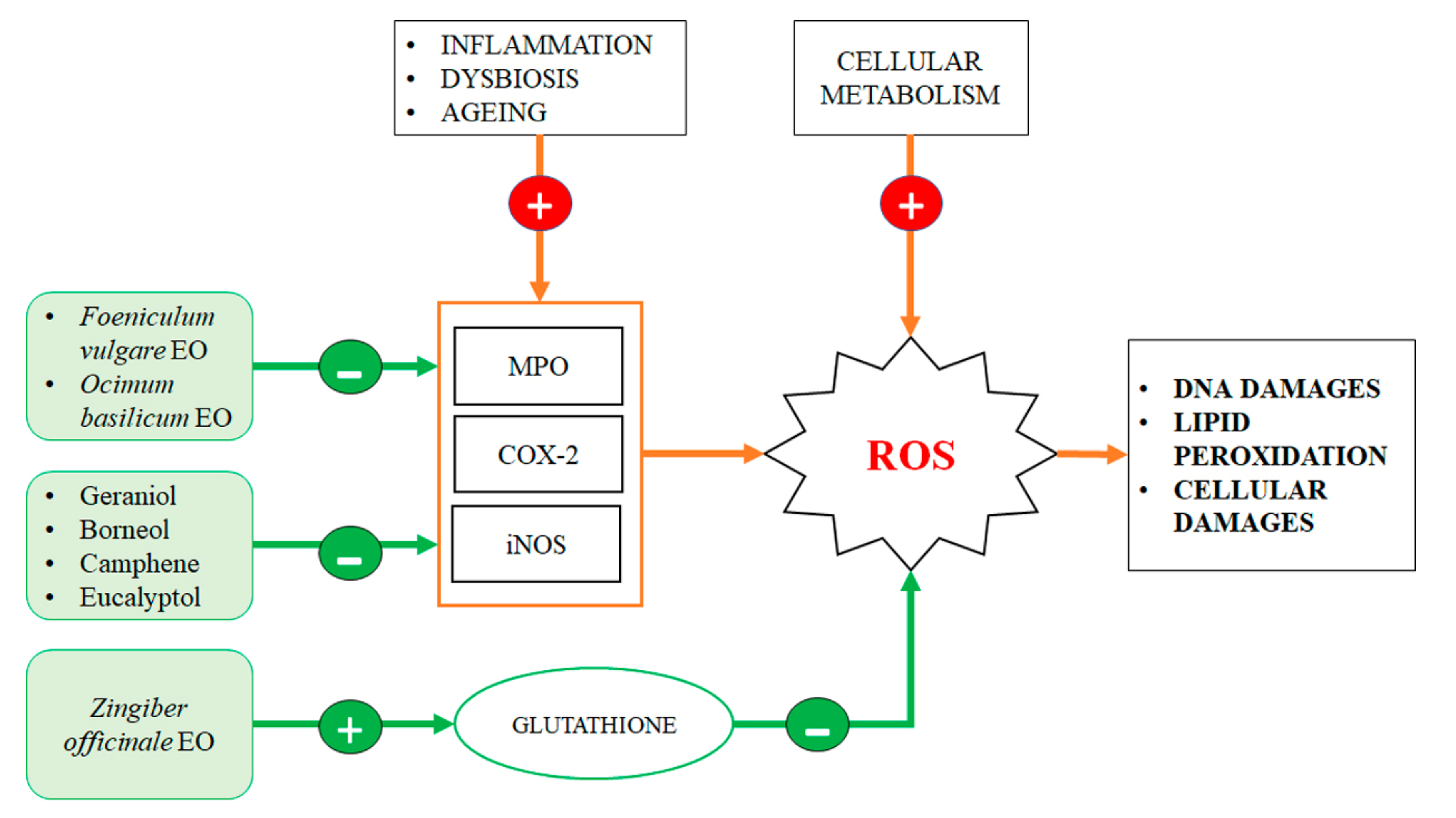
3. Antioxidant Effect of EO Components into the Gut
4. Intestinal Microbiota Modulation Exerted by EO Components
4.1. Antibacterial Proprieties of EOs and Bacterial Microbiota Modulation
4.2. EOs in the Modulation of Intestinal Mycobiome
4.3. EO Antiviral Activity and Their Possible Effects on the Intestinal Virome
5. Essential Oils Component with Antitumor Activity in Colorectal Carcinogenesis
6. Discussion and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ATP | Adenosine triphosphate |
| COX-2 | Cyclooxygenase-2 |
| CRC | Colorectal Cancer |
| DAI | Disease Activity Index |
| DSS | Dextran Sulfate Sodium |
| EO | Essential Oil |
| FAP | Familial Adenomatous Polyposis |
| FOXP3 | Forkhead box P3 |
| GALT | Gut-Associated Lymphoid Tissue |
| HFD | High Fat Diet |
| IBS | Irritable Bowel Syndrome |
| IBD | Inflammatory Bowel Disease |
| MIC | Minimum Inhibitory Concentration |
| MPO | Myeloperoxidase |
| NF-kβ | Nuclear Factor kappa-light-chain-enhancer of activated B cells |
| NOS | Nitric Oxide Synthase |
| ROS | Reactive Oxygen Species |
| TNF-α | Tumor Necrosis Factor-alpha |
References
- Sharma, M.; Koul, A.; Sharma, D.; Kaul, S.; Swamy, M.K.; Dhar, M.K. Metabolic engineering strategies for enhancing the production of bio-active compounds from medicinal plants. In Natural Bio-Active Compounds; Springer: Berlin/Heidelberg, Germany, 2019; pp. 287–316. [Google Scholar]
- Valdivieso-Ugarte, M.; Gomez-Llorente, C.; Plaza-Díaz, J.; Gil, Á. Antimicrobial, antioxidant, and immunomodulatory properties of essential oils: A systematic review. Nutrients 2019, 11, 2786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finkel, T. Oxidant signals and oxidative stress. Curr. Opin. Cell Biol. 2003, 15, 247–254. [Google Scholar] [CrossRef]
- Toni, M.; Massimino, M.L.; De Mario, A.; Angiulli, E.; Spisni, E. Metal dyshomeostasis and their pathological role in prion and prion-like diseases: The basis for a nutritional approach. Front Neurosci. 2017, 11, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavan, B.; Dalpiaz, A.; Marani, L.; Beggiato, S.; Ferraro, L.; Canistro, D.; Paolini, M.; Vivarelli, F.; Valerii, M.C.; Comparone, A.; et al. Geraniol pharmacokinetics, bioavailability and its multiple effects on the liver antioxidant and xenobiotic-metabolizing enzymes. Front. Pharm. 2018, 9, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzik, T.J.; Korbut, R.; Adamek-Guzik, T. Nitric oxide and superoxide in inflammation and immune regulation. J. Physiol. Pharm. 2003, 54, 469–487. [Google Scholar]
- De Fazio, L.; Spisni, E.; Cavazza, E.; Strillacci, A.; Candela, M.; Centanni, M.; Ricci, C.; Rizzello, F.; Campieri, M.; Valerii, M.C. Dietary geraniol by oral or enema administration strongly reduces dysbiosis and systemic inflammation in dextran sulfate sodium-treated mice. Front. Pharm. 2016, 7, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzello, F.; Ricci, C.; Scandella, M.; Cavazza, E.; Giovanardi, E.; Valerii, M.C.; Campieri, M.; Comparone, A.; De Fazio, L.; Candela, M.; et al. Dietary geraniol ameliorates intestinal dysbiosis and relieves symptoms in irritable bowel syndrome patients: A pilot study. BMC Complement Altern. Med. 2018, 18, 338. [Google Scholar] [CrossRef] [PubMed]
- De Fazio, L.; Cavazza, E.; Spisni, E.; Strillacci, A.; Centanni, M.; Candela, M.; Praticò, C.; Campieri, M.; Ricci, C.; Valerii, M.C. Longitudinal analysis of inflammation and microbiota dynamics in a model of mild chronic dextran sulfate sodium-induced colitis in mice. World J. Gastroenterol. 2014, 20, 2051–2061. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Shen, P.; Liu, J.; Gu, C.; Lu, X.; Li, Y.; Cao, Y.; Liu, B.; Fu, Y.; Zhang, N. In Vivo Study of the Efficacy of the Essential Oil of Zanthoxylum bungeanum Pericarp in Dextran Sulfate Sodium-Induced Murine Experimental Colitis. J. Agric. Food Chem. 2017, 65, 3311–3319. [Google Scholar] [CrossRef] [PubMed]
- Ji, K.L.; Gan, X.Q.; Xu, Y.K.; Li, X.F.; Guo, J.; Dahab, M.M.; Zhang, P. Protective effect of the essential oil of Zanthoxylum myriacanthum var. pubescens against dextran sulfate sodium-induced intestinal inflammation in mice. Phytomedicine 2016, 23, 883–890. [Google Scholar] [CrossRef] [PubMed]
- Rezayat, S.M.; Dehpour, A.R.; Motamed, S.M.; Yazdanparast, M.; Chamanara, M.; Sahebgharani, M.; Rashidian, A. Foeniculum vulgare essential oil ameliorates acetic acid-induced colitis in rats through the inhibition of NF-kB pathway. Inflammopharmacology 2018, 26, 851–859. [Google Scholar] [CrossRef] [PubMed]
- Ameh, S.J.; Ibekwe, N.N.; Ebeshi, B.U. Essential oils in ginger, hops, cloves, and pepper flavored beverages-A review. J. Diet Suppl. 2015, 12, 241–260. [Google Scholar] [CrossRef] [PubMed]
- Rashidian, A.; Mehrzadi, S.; Ghannadi, A.R.; Mahzooni, P.; Sadr, S.; Minaiyan, M. Protective effect of ginger volatile oil against acetic acid-induced colitis in rats: A light microscopic evaluation. J. Integr. Med. 2014, 12, 115–120. [Google Scholar] [CrossRef]
- Rashidian, A.; Roohi, P.; Mehrzadi, S.; Ghannadi, A.R.; Minaiyan, M. Protective effect of Ocimum basilicum essential oil against acetic acid-induced colitis in rats. J. Evid. Based Complementary Altern. Med. 2016, 21, NP36–NP42. [Google Scholar] [CrossRef] [PubMed]
- Mueller, K.; Blum, N.M.; Mueller, A.S. Examination of the anti-inflammatory, antioxidant, and xenobiotic-inducing potential of broccoli extract and various essential oils during a mild DSS-induced colitis in rats. Isrn. Gastroenterol. 2013, 2013, 710856. [Google Scholar] [CrossRef] [PubMed]
- Toden, S.; Theiss, A.L.; Wang, X.; Goel, A. Essential turmeric oils enhance anti-inflammatory efficacy of Curcumin in dextran sulfate sodium-induced colitis. Sci. Rep. 2017, 7, 814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuzarte, M.; Alves-Silva, J.M.; Alves, M.; Cavaleiro, C.; Salgueiro, L.; Cruz, M.T. New insights on the anti-inflammatory potential and safety profile of Thymus carnosus and Thymus camphoratus essential oils and their main compounds. J. Ethnopharmacol. 2018, 225, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Borges, R.S.; Ortiz, B.L.S.; Pereira, A.C.M.; Keita, H.; Carvalho, J.C.T. Rosmarinus officinalis essential oil: A review of its phytochemistry, anti-inflammatory activity, and mechanisms of action involved. J. Ethnopharmacol. 2019, 229, 29–45. [Google Scholar] [CrossRef] [PubMed]
- Sinha, S.; Biswas, D.; Mukherjee, A. Antigenotoxic and antioxidant activities of palmarosa and citronella essential oils. J. Ethnopharmacol. 2011, 137, 1521–1527. [Google Scholar] [CrossRef] [PubMed]
- Thapa, D.; Richardson, A.J.; Zweifel, B.; Wallace, R.J.; Gratz, S.W. Genoprotective effects of essential oil compounds against oxidative and methylated DNA damage in human colon cancer cells. J. Food Sci. 2019, 84, 1979–1985. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Peng, F.; Xie, J.; Peng, C. Pharmacological properties of geraniol. Planta. Med. 2019, 85, 48–55. [Google Scholar]
- Lee, S.J.; Umano, K.; Shibamoto, T.; Lee, K.G. Identification of volatile components in basil (Ocimum basilicum L.) and thyme leaves (Thymus vulgaris L.) and their antioxidant properties. Food Chem. 2005, 91, 131–137. [Google Scholar] [CrossRef]
- Kamada, N.; Seo, S.U.; Chen, G.Y.; Núñez, G. Role of the gut microbiota in immunity and inflammatory disease. Nat. Rev. Immunol. 2013, 13, 321–335. [Google Scholar] [CrossRef] [PubMed]
- Tap, J.; Mondot, S.; Levenez, F.; Pelletier, E.; Caron, C.; Furet, J.P.; Ugarte, E.; Muñoz-Tamayo, R.; Paslier, D.L.E.; Nalin, R.; et al. Towards the human intestinal microbiota phylogenetic core. Environ. Microbiol. 2009, 11, 2574–2584. [Google Scholar] [CrossRef] [PubMed]
- Swamy, M.K.; Akhtar, M.S.; Sinniah, U.R. Antimicrobial properties of plant essential oils against human pathogens and their mode of action: An updated review. Evid. Based Complement. Altern. Med. 2016, 2016, 3012462. [Google Scholar] [CrossRef] [PubMed]
- Tariq, S.; Wani, S.; Rasool, W.; Bhat, M.A.; Prabhakar, A.; Shalla, A.H.; Rather, M.A. A comprehensive review of the antibacterial, antifungal and antiviral potential of essential oils and their chemical constituents against drug-resistant microbial pathogens. Microb. Pathog. 2019, 134, 103580. [Google Scholar] [CrossRef] [PubMed]
- Solórzano-Santos, F.; Miranda-Novales, M.G. Essential oils from aromatic herbs as antimicrobial agents. Curr. Opin. Biotechnol. 2012, 23, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Lorenzi, V.; Muselli, A.; Bernardini, A.F.; Berti, L.; Pagès, J.M.; Amaral, L.; Bolla, J.M. Geraniol restores antibiotic activities against multidrug-resistant isolates from Gram-negative species. Antimicrob. Agents Chemother. 2009, 53, 2209–2211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thapa, D.; Losa, R.; Zweifel, B.; Wallace, R.J. Sensitivity of pathogenic and commensal bacteria from the human colon to essential oils. Microbiology 2012, 158, 2870–2877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, A.O.; Holley, R.A. Disruption of Escherichia coli, Listeria monocytogenes and Lactobacillus sakei cellular membranes by plant oil aromatics. Int. J. Food Microbiol. 2006, 108, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Zhang, C.; Li, C.; Lin, L. Antimicrobial mechanism of clove oil on Listeria monocytogenes. Food Control. 2018, 94, 140–146. [Google Scholar] [CrossRef]
- Wlodarska, M.; Willing, B.; Bravo, D.; Finlay, B.B. Phytonutrient diet supplementation promotes beneficial Clostridia species and intestinal mucus secretion resulting in protection against enteric infection. Sci. Rep. 2015, 5, 9253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Zheng, H.; Tang, Y.; Yu, W.; Gong, Q. Eugenol inhibits quorum sensing at sub-inhibitory concentrations. Biotechnol. Lett. 2013, 35, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Bayoub, K.; Baibai, T.; Mountassif, D.; Retmane, A.; Soukri, A. Antibacterial activities of the crude ethanol extracts of medicinal plants against listeria monocytogenes and some other pathogenic strains. Afr. J. Biotechnol. 2010, 9, 4251–4258. [Google Scholar]
- Senhaji, O.; Faid, M.; Kalalou, I. Inactivation of Escherichia coli O157:H7 by essential oil from cinnamomum zeylanicum. Braz. J. Infect. Dis. 2007, 11, 234–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meades, G., Jr.; Henken, R.L.; Waldrop, G.L.; Rahman, M.M.; Gilman, S.D.; Kamatou, G.P.; Viljoen, A.M.; Gibbons, S. Constituents of cinnamon inhibit bacterial acetyl CoA carboxylase. Planta Med. 2010, 76, 1570–1575. [Google Scholar] [CrossRef] [PubMed]
- Unlu, M.; Ergene, E.; Unlu, G.V.; Zeytinoglu, H.S.; Vural, N. Composition, antimicrobial activity and in vitro cytotoxicity of essential oil from cinnamomum zeylanicum blume (lauraceae). Food Chem. Toxicol. 2010, 48, 3274–3280. [Google Scholar] [CrossRef] [PubMed]
- Shahverdi, A.R.; Monsef-Esfahani, H.R.; Tavasoli, F.; Zaheri, A.; Mirjani, R. Trans-cinnamaldehyde from cinnamomum zeylanicum bark essential oil reduces the clindamycin resistance of clostridium difficile in vitro. J. Food Sci. 2007, 72, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Li, A.L.; Ni, W.W.; Zhang, Q.M.; Li, Y.; Zhang, X.; Wu, H.Y.; Du, P.; Hou, J.C.; Zhang, Y. Effect of cinnamon essential oil on gut microbiota in the mouse model of dextran sodium sulfate-induced colitis. Microbiol. Immunol. 2020, 64, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Abouelezz, K.; Abou-Hadied, M.; Yuan, J.; Elokil, A.; Wang, G.; Wang, S.; Wang, J.; Bian, G. Nutritional impacts of dietary oregano and Enviva essential oils on the performance, gut microbiota and blood biochemicals of growing ducks. Animal 2019, 13, 2216–2222. [Google Scholar] [CrossRef] [PubMed]
- Tiihonen, K.; Kettunen, H.; Bento, M.H.L.; Saarinen, M.; Lahtinen, S.; Ouwehand, A.C.; Schulze, H.; Rautonen, N. The effect of feeding essential oils on broiler performance and gut microbiota. Br. Poul. Sci. 2010, 51, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Maheshwari, M.; Abul Qais, F.; Althubiani, A.S.; Abulreesh, H.H.; Ahmad, I. Bioactive extracts of Carum copticum and thymol inhibit biofilm development by multidrug-resistant extended spectrum β-lactamase producing enteric bacteria. Biofouling 2019, 35, 1026–1039. [Google Scholar] [CrossRef] [PubMed]
- Thumann, T.A.; Pferschy-Wenzig, E.M.; Moissl-Eichinger, C.; Bauer, R. The role of gut microbiota for the activity of medicinal plants traditionally used in the European Union for gastrointestinal disorders. J. Ethnopharmacol. 2019, 245, 112153. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Xue, H.; Zhou, Z.; Peng, J. A carvacrol–thymol blend decreased intestinal oxidative stress and influenced selected microbes without changing the messenger RNA levels of tight junction proteins in jejunal mucosa of weaning piglets. Animal 2017, 11, 193–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Song, M.; Yun, W.; Lee, J.; Lee, C.; Kwak, W.; Han, N.; Kim, H.; Cho, J. Effects of oral administration of different dosages of carvacrol essential oils on intestinal barrier function in broilers. J. Anim. Physiol. Anim. Nutr. (Berl.) 2018, 102, 1257–1265. [Google Scholar] [CrossRef] [PubMed]
- Kelly, C.; Gundogdu, O.; Pircalabioru, G.; Cean, A.; Scates, P.; Linton, M.; Pinkerton, L.; Magowan, E.; Stef, L.; Simiz, E.; et al. The in vitro and in vivo effect of carvacrol in preventing Campylobacter infection, colonization and in improving productivity of chicken broilers. Foodborne Pathog. Dis. 2017, 14, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Castañeda-Correa, A.; Corral-Luna, A.; Hume, M.E.; Anderson, R.C.; Ruiz-Barrera, O.; Castillo-Castillo, Y.; Rodriguez-Almeida, F.; Salinas-Chavira, J.; Arzola-Alvarez, C. Effects of thymol and carvacrol, alone or in combination, on fermentation and microbial diversity during in vitro culture of bovine rumen microbes. J. Environ. Sci. Health B 2019, 54, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Betancourt, L.; Rodriguez, F.; Phandanouvong, V.; Ariza-Nieto, C.; Hume, M.; Nisbet, D.; Afanador-Téllez, G.; Van Kley, A.M.; Nalian, A. Effect of Origanum chemotypes on broiler intestinal bacteria. Poul. Sci. 2014, 93, 2526–2535. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, Y.; Fan, G.; Ren, J.N.; Zhang, L.L.; Pan, S.Y. Effects of orange essential oil on intestinal microflora in mice. J. Sci. Food Agric. 2019, 99, 4019–4028. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wu, h.; Dou, H.; Guo, L.; Huang, W. Microcapsule of sweet orange essential oil changes gut microbiota in diet-induced obese rats. Biochem. Biophys. Res. Commun. 2018, 505, 991–995. [Google Scholar] [CrossRef] [PubMed]
- Dhakad, A.K.; Pandey, V.V.; Beg, S.; Rawat, J.M.; Singh, A. Biological, medicinal and toxicological significance of Eucalyptus leaf essential oil: A review. J. Sci. Food. Agric. 2018, 98, 833–848. [Google Scholar] [CrossRef] [PubMed]
- Kamatou, G.P.P.; Vermaak, I.; Viljoen, A.M.; Lawrence, B.M. Menthol: A simple monoterpene with remarkable biological properties. Phytochemistry 2013, 96, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Roshan, N.; Riley, T.V.; Knight, D.R.; Steer, J.H.; Hammer, K.A. Natural products show diverse mechanisms of action against Clostridium difficile. J. Appl. Microbiol. 2019, 126, 468–479. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. Human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sam, Q.H.; Chang, M.W.; Chai, L.Y.A. The fungal mycobiome and its interaction with gut bacteria in the host. Int. J. Mol. Sci. 2017, 18, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iliev, I.D.; Funari, V.A.; Taylor, K.D.; Nguyen, Q.; Reyes, C.N.; Strom, S.P.; Brown, J.; Becker, C.A.; Fleshner, P.R.; Dubinsky, M.; et al. Interactions between commensal fungi and the C-type lectin receptor Dectin-1 influence colitis. Science 2012, 336, 1314–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, Y.; Tang, J. Candida albicans infection and intestinal immunity. Microbiol. Res. 2017, 198, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Botschuijver, S.; Roeselers, G.; Levin, E.; Jonkers, D.M.; Welting, O.; Heinsbroek, S.E.M.; de Weerd, H.H.; Boekhout, T.; Fornai, M.; Masclee, A.A.; et al. Intestinal fungal dysbiosis associates with visceral hypersensitivity in patients with irritable bowel syndrome and rats. Gastroenterology 2017, 153, 1026–1039. [Google Scholar] [CrossRef] [PubMed]
- Sokol, H.; Leducq, V.; Aschard, H.; Pham, H.; Jegou, S.; Landman, C.; Cohen, D.; Liguori, G.; Bourrier, A.; Nion-Larmurier, I.; et al. Fungal microbiota dysbiosis in IBD. Gut 2017, 66, 1039–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iliev, I.D.; Leonardi, I. Fungal dysbiosis: Immunity and interactions at mucosal barriers. Nat. Rev. Immunol. 2017, 17, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Limon, J.J.; Skalski, J.H.; Underhill, D.M. Commensal fungi in health and disease. Cell Host Microbe. 2017, 22, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Bona, E.; Cantamessa, S.; Pavan, M.; Novello, G.; Massa, N.; Rocchetti, A.; Berta, G.; Gamalero, E. Sensitivity of Candida albicans to essential oils: Are they an alternative to antifungal agents? J. Appl. Microbiol. 2016, 121, 1530–1545. [Google Scholar] [CrossRef] [PubMed]
- Gucwa, K.; Milewski, S.; Dymerski, T.; Szweda, P. Investigation of the antifungal activity and mode of action of thymus vulgaris, citrus limonum, pelargonium graveolens, cinnamomum cassia, ocimum basilicum, and eugenia caryophyllus essential oils. Molecules 2018, 23, 1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shreaz, S.; Bhatia, R.; Khan, N.; Maurya, I.K.; Ahmad, S.I.; Muralidhar, S.; Manzoor, N.; Khan, L.A. Cinnamic aldehydes affect hydrolytic enzyme secretion and morphogenesis in oral Candida isolates. Microb. Pathog. 2012, 52, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Espina, L.; Gelaw, T.K.; de Lamo-Castellví, S.; Pagán, R.; García-Gonzalo, D. Mechanism of bacterial inactivation by (+)-limonene and its potential use in food preservation combined processes. PLoS ONE 2013, 8, e056769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakre, A.; Zore, G.; Kodgire, S.; Kazi, R.; Mulange, S.; Patil, R.; Shelar, A.; Santhakumari, B.; Kulkarni, M.; Kharat, K.; et al. Limonene inhibits Candida albicans growth by inducing apoptosis. Med. Mycol. 2018, 56, 565–578. [Google Scholar] [PubMed]
- Stringaro, A.; Colone, M.; Angiolella, L. Antioxidant, antifungal, antibiofilm, and cytotoxic activities of Mentha spp. essential oils. Medicines 2018, 5, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botschuijver, S.; Welting, O.; Levin, E.; Maria-Ferreira, D.; Koch, E.; Montijin, R.C.; Seppen, J.; Hakvoort, T.B.M.; Schuren, F.H.J.; de Jonge, W.; et al. Reversal of visceral hypersensitivity in rat by Menthacarin®, a proprietary combination of essential oils from peppermint and caraway, coincides with mycobiome modulation. Neurogastroenterol. Motil. 2018, 30, e13299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, E.; Pina-Vaz, C.; Salgueiro, L.; Gonçalves, M.J.; Costa-de-Oliveira, S.; Cavaleiro, C.; Palmeira, A.; Rodrigues, A.; Martinez-de-Oliveira, J. Antifungal activity of the essential oil of Thymus pulegioides on Candida, Aspergillus and dermatophyte species. J. Med. Microbiol. 2006, 55, 1367–1373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vasconcelos, L.C.; Sampaio, F.C.; Albuquerque Ade, J.; Vasconcelos, L.C. Cell viability of Candida albicans against the antifungal activity of thymol. Braz. Dent. J. 2014, 25, 277–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatrath, A.; Gangwar, R.; Kumari, P.; Prasad, R. In vitro anti-biofilm activities of citral and thymol against Candida tropicalis. J. Fungi 2019, 5, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchese, A.; Barbieri, R.; Coppo, E.; Orhan, I.E.; Daglia, M.; Nabavi, S.F.; Izadi, M.; Abdollahi, M.; Nabavi, S.M.; Ajami, M. Antimicrobial activity of eugenol and essential oils containing eugenol: A mechanistic viewpoint. Crit. Rev. Microbiol. 2017, 43, 668–689. [Google Scholar] [CrossRef] [PubMed]
- Rajkowska, K.; Nowicka-Krawczyk, P.; Kunicka-Styczynska, A. Effect of clove and thyme essential oils on Candida biofilm formation and the oil distribution in yeast cells. Molecules 2019, 24, 1954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braga, P.; Dal Sasso, M.; Culici, M.; Alfieri, M. Eugenol and thymol, alone or in combination, induce morphological alterations in the envelope of Candida albicans. Fitoterapia 2007, 78, 396–400. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Khan, A.; Yousuf, S.; Khan, L.A.; Manzoor, N. Proton translocating ATPase mediated fungicidal activity of eugenol and thymol. Fitoterapia 2010, 81, 1157–1162. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Ahmad, A.; Akhtar, F.; Yousuf, S.; Xess, I.; Ahmad, L. Induction of oxidative stress as a possible mechanism of the antifungal action of three phenylpropanoids. Fems. Yeast Res. 2011, 11, 114–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnitzler, P. Essential oils for the treatment of Herpes simplex virus infections. Chemotherapy 2019, 64, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Loizzo, M.R.; Saab, A.M.; Tundis, R.; Statti, G.A.; Menichini, F.; Lampronti, I.; Doerr, H.W. Phytochemical analysis and in vitro antiviral activities of the essential oils of seven lebanon species. Chem. Biodivers. 2008, 5, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Astani, A.; Schnitzler, P. Antiviral activity of monoterpenes beta-pinene and limonene against herpes simplex virus in vitro. Iran. J. Microbiol. 2014, 6, 149–155. [Google Scholar] [PubMed]
- Wu, Q.F.; Wang, W.; Dai, X.Y.; Wang, Z.Y.; Shen, Z.H.; Ying, H.Z.; Yu, C.H. Chemical compositions and anti-influenza activities of essential oils from Mosla dianthera. J. Ethnopharmacol. 2012, 139, 668–671. [Google Scholar] [CrossRef] [PubMed]
- Kubiça, T.F.; Alves, S.H.; Weiblen, R.; Lovato, L.T. In vitro inhibition of the bovine viral diarrhoea virus by the essential oil of Ocimum basilicum (basil) and monoterpenes. Braz. J. Microbiol. 2014, 45, 209–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilling, D.H.; Kitajima, M.; Torrey, J.R.; Bright, K.R. Antiviral efficacy and mechanisms of action of oregano essential oil and its primary component carvacrol against murine norovirus. J. Appl. Microbiol. 2014, 116, 1149–1163. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.U.; Kweon, M.N. Virome-host interactions in intestinal health and disease. Curr. Opin. Virol. 2019, 37, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Reyes, A.; Haynes, M.; Hanson, N.; Angly, F.E.; Heath, A.C.; Rohwer, F.; Gordon, J.I. Viruses in the faecal microbiota of monozygotic twins and their mothers. Nature 2010, 466, 334–338. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Park, E.J.; Roh, S.W.; Bae, J.W. Diversity and abundance of single-stranded DNA viruses in human feces. Appl. Environ. Microbiol. 2011, 77, 8062–8070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, M.K.; Watanabe, M.; Zhu, S.; Graves, C.L.; Keyes, L.R.; Grau, K.R.; Gonzalez-Hernandez, M.B.; Iovine, N.M.; Wobus, C.E.; Vinjé, J.; et al. Enteric bacteria promote human and mouse norovirus infection of B cells. Science 2014, 346, 755–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldridge, M.T.; Nice, T.J.; McCune, B.T.; Yokoyama, C.C.; Kambal, A.; Wheadon, M.; Diamond, M.S.; Ivanova, Y.; Artyomov, M.; Virgin, H.W. Commensal microbes and interferon-λ determine persistence of enteric murine norovirus infection. Science 2015, 347, 266–269. [Google Scholar] [CrossRef] [Green Version]
- Malvezzi, M.; Carioli, G.; Bertuccio, P.; Boffetta, P.; Levi, F.; La Vecchia, C.; Negri, E. European cancer mortality predictions for the year 2018 with focus on colorectal cancer. Ann. Oncol. 2018, 29, 1016–1022. [Google Scholar] [CrossRef]
- Witold, K.; Anna, K.; Maciej, T.; Jakub, J. Adenomas-genetic factors in colorectal cancer prevention. Rep. Pr. Oncol. Radiother. 2018, 23, 75–83. [Google Scholar] [CrossRef]
- Shen, Y.; Wang, J.; Han, X.; Yang, H.; Wang, S.; Lin, D.; Shi, Y. Effectors of epidermal growth factor receptor pathway: The genetic profiling of KRAS, BRAF, PIK3CA, NRAS mutations in colorectal cancer characteristics and personalized medicine. PLoS ONE 2013, 8, e81628. [Google Scholar] [CrossRef] [PubMed]
- Roth, A.D.; Delorenzi, M.; Tejpar, S.; Yan, P.; Klingbiel, D.; Fiocca, R.; d’Ario, G.; Cisar, L.; Labianca, R.; Cunningham, D. Integrated analysis of molecular and clinical prognostic factors in stage II/III colon cancer. J. Natl. Cancer Inst. 2012, 104, 1635–1646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, U.; Jiao, S.; Schumacher, F.R.; Hutter, C.M.; Aragaki, A.K.; Baron, J.A.; Berndt, S.I.; Bezieau, S.; Brenner, H.; Butterbach, K.; et al. Colon Cancer Family Registry and the Genetics and Epidemiology of Colorectal Cancer Consortium. Identification of genetic susceptibility loci for colorectal tumors in a genome-wide meta-analysis. Gastroenterology 2013, 144, 799–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruger, C.; Zhou, Y. Red meat and colon cancer: A review of mechanistic evidence for heme in the context of risk assessment methodology. Food Chem. Toxicol. 2018, 118, 131–153. [Google Scholar] [CrossRef] [PubMed]
- Fagunwa, I.O.; Loughrey, M.B.; Coleman, H.G. Alcohol, smoking and the risk of premalignant and malignant colorectal neoplasms. Best Pr. Res. Clin. Gastroenterol. 2017, 31, 561–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louis, P.; Hold, G.L.; Flint, H.J. The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Microbiol. 2014, 12, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Gagniere, J.; Raisch, J.; Veziant, J.; Barnich, N.; Bonnet, R.; Buc, E.; Bringer, M.A.; Pezet, D.; Bonnet, M. Gut microbiota imbalance and colorectal cancer. World J. Gastroenterol. 2016, 22, 501–518. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.R.; Chang, D.K. Colorectal cancer in inflammatory bowel disease: The risk, pathogenesis, prevention and diagnosis. World J. Gastroenterol. 2014, 20, 9872–9881. [Google Scholar] [CrossRef] [PubMed]
- Bogaert, J.; Prenen, H. Molecular genetics of colorectal cancer. Ann. Gastroenterol. 2014, 27, 9–14. [Google Scholar] [PubMed]
- Vasen, H.F.A.; Tomlinson, I.; Castells, A. Clinical management of hereditary colorectal cancer syndromes. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Dekker, E.; Tanis, P.J.; Vleugels, J.L.A.; Kasi, P.M.; Wallace, M.B. Colorectal cancer. Lancet 2019, 394, 1467–1487. [Google Scholar] [CrossRef]
- Braun, M.S.; Seymour, M.T. Balancing the efficacy and toxicity of chemotherapy in colorectal cancer. Adv. Med. Oncol. 2011, 3, 43–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammond, W.A.; Swaika, A.; Mody, K. Pharmacologic resistance in colorectal cancer: A review. Adv. Med. Oncol. 2016, 8, 57–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhillon, S. Regorafenib: A Review in Metastatic Colorectal Cancer. Drugs 2018, 78, 1133–1144. [Google Scholar] [CrossRef] [PubMed]
- Fan, K.; Li, X.; Cao, Y.; Qi, H.; Li, L.; Zhang, Q.; Sun, H. Carvacrol inhibits proliferation and induces apoptosis in human colon cancer cells. Anti-Cancer Drugs 2015, 26, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Qi, F.; Yan, Q.; Zheng, Z.; Liu, J.; Chen, Y.; Zhang, G. Geraniol and geranyl acetate induce potent anticancer effects in colon cancer Colo-205 cells by inducing apoptosis, DNA damage and cell cycle arrest. J. BUON 2018, 23, 346–352. [Google Scholar] [PubMed]
- Carnesecchi, S.; Schneider, Y.; Ceraline, J.; Duranton, B.; Gosse, F.; Seiler, N.; Raul, F. Geraniol, a component of plant essential oils, inhibits growth and polyamine biosynthesis in human colon cancer cells. J. Pharm. Exp. 2001, 298, 197–200. [Google Scholar]
- Soda, K. Polyamine metabolism and gene methylation in conjunction with one-carbon metabolism. Int. J. Mol. Sci. 2018, 19, 3106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chauhan, A.K.; Bahuguna, A.; Paul, S.; Kang, S.C. Thymol elicits HCT-116 colorectal carcinoma cell death through induction of oxidative stress. Anticancer Agents Med. Chem. 2018, 17, 1942–1950. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Teng, Y.; Liu, S.; Wang, Z.; Chen, Y.; Zhang, Y.; Xi, S.; Xu, S.; Wang, R.; Zou, X. Cinnamaldehyde affects the biological behaviour of human colorectal cancer cells and induces apoptosis via inhibition of the PI3K/Akt signalling pathway. Oncol. Rep. 2016, 35, 1501–1510. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Obianom, O.N.; Ai, Y. Novel cinnamaldehyde-based aspirin derivatives for the treatment of colorectal cancer. Bioorg. Med. Chem. Lett. 2018, 28, 2869–2874. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Lee, E.; Kim, Y.; Park, S.; Khang, G.; Lee, D. Dual acid-responsive micelle-forming anticancer polymers as new anticancer therapeutics. Adv. Funct. Mater. 2013, 23, 5091–5097. [Google Scholar] [CrossRef]
- Carnesecchi, S.; Bradaia, A.; Fischer, B.; Coelho, D.; Schöller-Guinard, M.; Gosse, F.; Raul, F. Perturbation by geraniol of cell membrane permeability and signal transduction pathways in human colon cancer cells. J. Pharm. Exp. 2002, 303, 711–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carnesecchi, S.; Langley, K.; Exinger, F.; Gosse, F.; Raul, F. Geraniol, a component of plant essential oils, sensitizes human colonic cancer cells to 5-fluorouracil treatment. J. Pharm. Exp. 2002, 301, 625–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carnesecchi, S.; Bras-Gonçalves, R.; Bradaia, A.; Zeisel, M.; Gossé, F.; Poupon, M.F.; Raul, F. Geraniol, a component of plant essential oils, modulates DNA synthesis and potentiates 5-fluorouracil efficacy on human colon tumor xenografts. Cancer Lett. 2004, 215, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Effenberger-Neidnicht, K.; Schobert, R. Combinatorial effects of thymoquinone on the anti-cancer activity of doxorubicin. Cancer Chemother. Pharm. 2011, 67, 867–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Bai, Y.; Yang, Y. Thymoquinone chemosensitizes colon cancer cells through inhibition of NF-κB. Oncol. Lett. 2016, 12, 2840–2845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.C.; Lee, N.H.; Hsu, H.H.; Ho, T.J.; Tu, C.C.; Hsieh, D.J.; Lin, Y.M.; Chen, L.M.; Kuo, W.W.; Huang, C.Y. Thymoquinone induces caspase-independent, autophagic cell death in CPT-11-resistant LoVo colon cancer via mitochondrial dysfunction and activation of JNK and p38. J. Agric. Food Chem. 2015, 63, 1540–1546. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.C.; Lee, N.H.; Hsu, H.H.; Ho, T.J.; Tu, C.C.; Chen, R.J.; Lin, Y.M.; Viswanadha, V.P.; Kuo, W.W.; Huang, C.Y. Inhibition of NF-κB and metastasis in irinotecan (CPT-11)-resistant LoVo colon cancer cells by thymoquinone via JNK and p38. Environ. Toxicol. 2017, 32, 669–678. [Google Scholar] [CrossRef] [PubMed]
- Legault, J.; Pichette, A. Potentiating effect of β-caryophyllene on anticancer activity of α-humulene, isocaryophyllene and paclitaxel. J. Pharm. Pharm. 2007, 59, 1643–1647. [Google Scholar] [CrossRef] [PubMed]
- Ambrož, M.; Boušová, I.; Skarka, A.; Hanušová, V.; Králová, V.; Matoušková, P.; Szotáková, B.; Skálová, L. The influence of sesquiterpenes from Myrica rubra on the antiproliferative and pro-oxidative effects of doxorubicin and its accumulation in cancer cells. Molecules 2015, 20, 15343–15358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Zhan, M.L.; Tang, Y.; Xiao, M.; Li, M.; Li, Q.S.; Yang, L.; Li, X.; Chen, W.W.; Wang, Y.L. Effects of beta-caryophyllene on arginine ADP-ribosyltransferase 1-mediated regulation of glycolysis in colorectal cancer under high-glucose conditions. Int. J. Oncol. 2018, 53, 1613–1624. [Google Scholar] [PubMed] [Green Version]
- Guénette, S.A.; Ross, A.; Marier, J.F.; Beaudry, F.; Vachon, P. Pharmacokinetics of eugenol and its effects on thermal hypersensitivity in rats. Eur. J. Pharm. 2007, 562, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Xie, Y.; Yang, Q.; Cao, Y.; Tu, H.; Cao, W.; Wang, S. Pharmacokinetic study of cinnamaldehyde in rats by GC–MS after oral and intravenous administration. J. Pharm. Biomed. 2014, 89, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Allaoua, M.; Etienne, P.; Noirot, V.; Carayon, J.L.; Téné, N.; Bonnafé, E.; Treilhou, M. Pharmacokinetic and antimicrobial activity of a new carvacrol-based product against a human pathogen, Campylobacter jejuni. J. Appl. Microbiol. 2018, 125, 1162–1174. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Sheng, Y.; Hu, Y.; Sun, J.; Li, W.; Feng, H.; Tang, L. Determination of d-limonene in mice plasma and tissues by a new GC-MS/MS method: Comparison of the pharmacokinetics and tissue distribution by oral and inhalation administration in mice. Biomed. Chromatogr. 2019, 33, e4530. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Xu, W.; Zhang, J.; Liao, Y.; Firempong, C.K.; Adu-Frimpong, M.; Deng, W.; Zhang, H.; Yu, J.; Xu, X. Self-microemulsifying Drug Delivery System for Improved Oral Delivery of Limonene: Preparation, Characterization, in vitro and in vivo Evaluation. Aaps Pharm. Sci. Tech. 2019, 20, 153. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Yang, G.; Tang, Y.; Cao, D.; Qi, T.; Qi, Y.; Fan, G. Physicochemical characterization and pharmacokinetics evaluation of β-caryophyllene/β-cyclodextrin inclusion complex. Int. J. Pharma 2013, 450, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Kohlert, C.; Schindler, G.; März, R.W.; Abel, G.; Brinkhaus, B.; Derendorf, H.; Gräfe, E.U.; Veit, M. Systemic availability and pharmacokinetics of thymol in humans. J. Clin. Pharm. 2002, 42, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Sivaranjani, A.; Sivagami, G.; Nalini, N. Chemopreventive effect of carvacrol on 1,2-dimethylhydrazine induced experimental colon carcinogenesis. J. Cancer Res. Ther. 2016, 12, 755–762. [Google Scholar] [PubMed]
- Arigesavan, K.; Sudhandiran, G. Carvacrol exhibits anti-oxidant and anti-inflammatory effects against 1, 2 dimethyl hydrazine plus dextran sodium sulfate induced inflammation associated carcinogenicity in the colon of Fischer 344 rats. Biochem. Biophys. Res. Commun. 2015, 461, 314–320. [Google Scholar] [CrossRef] [PubMed]




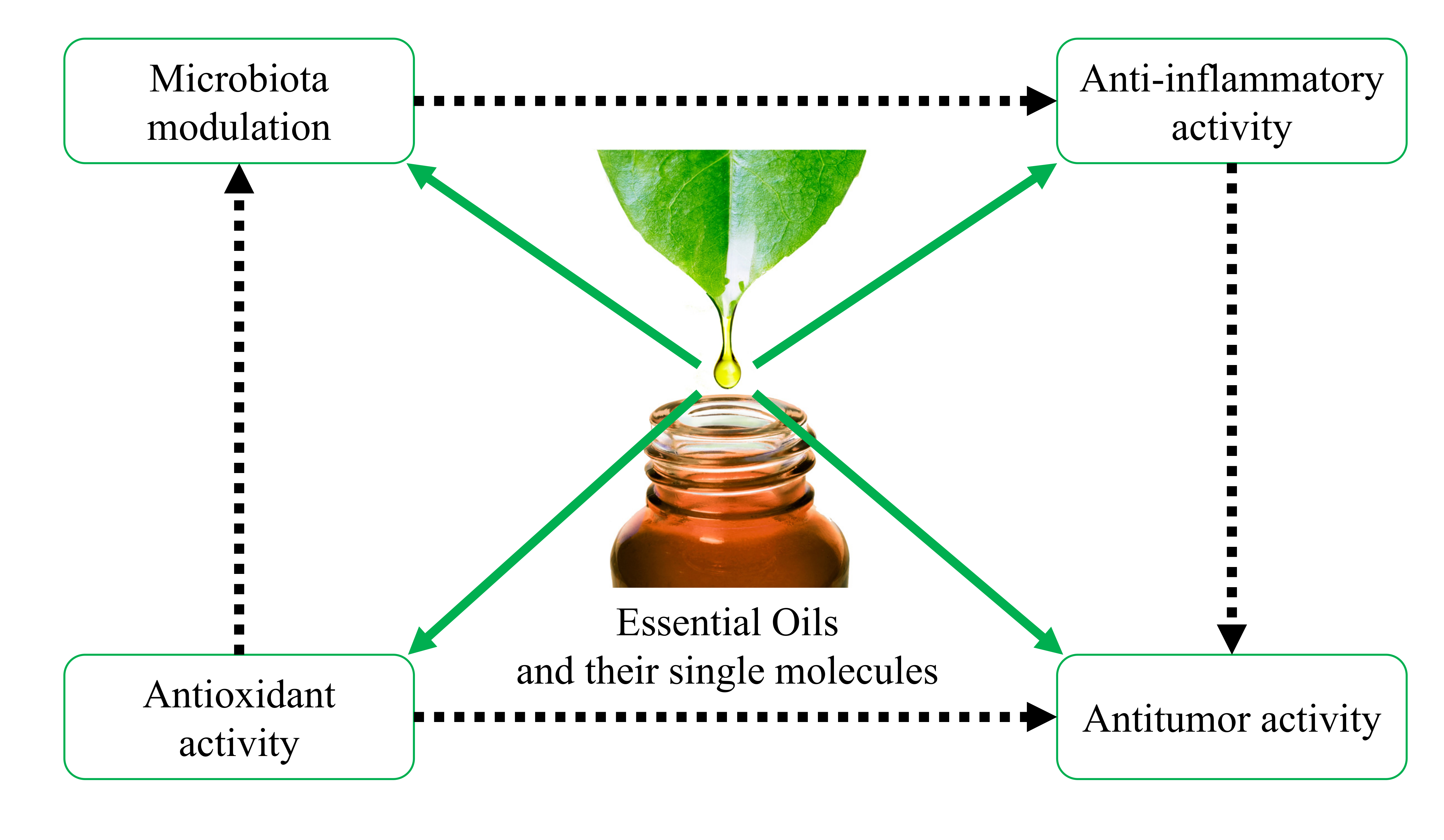{kind=link}
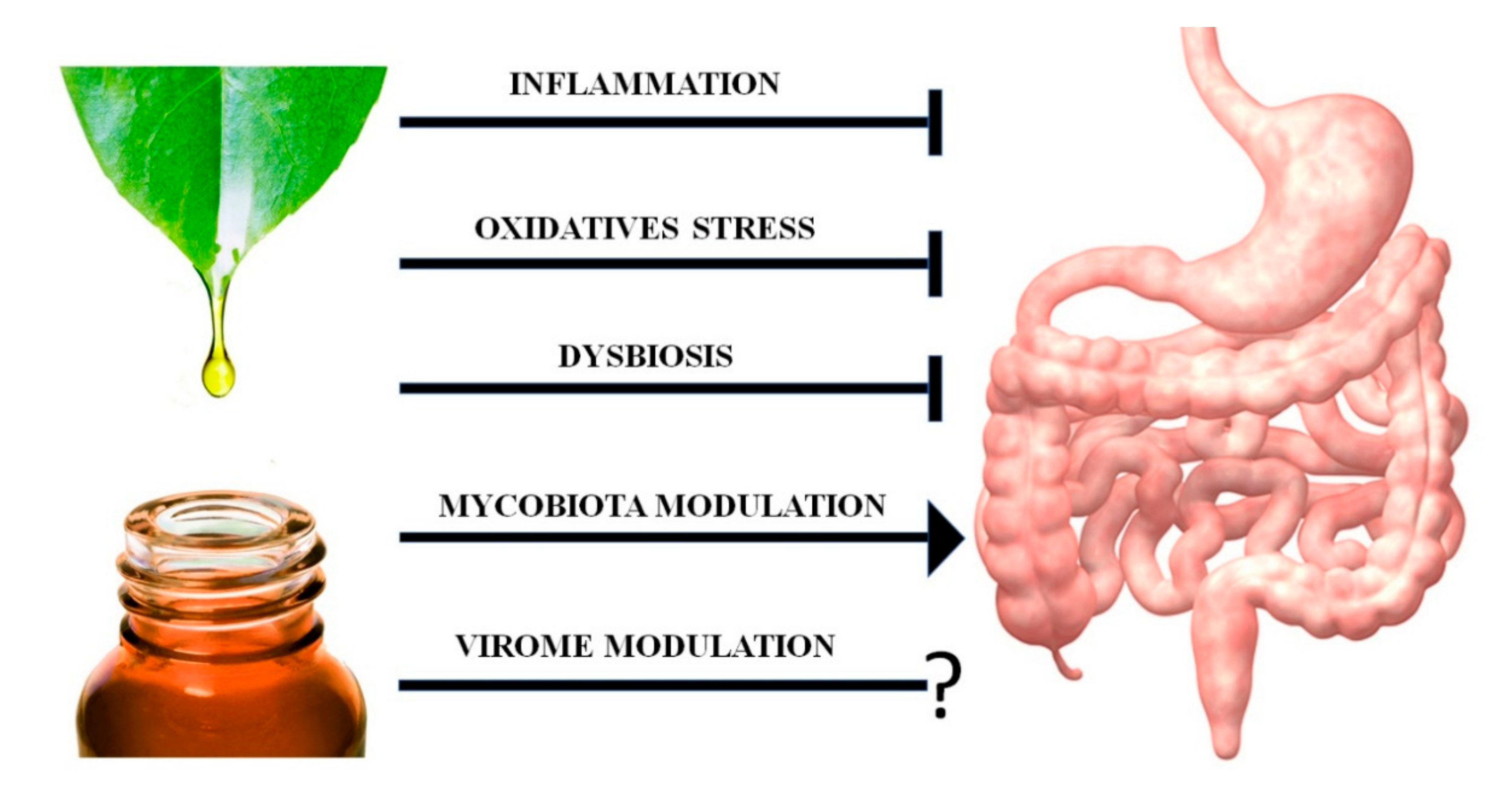{kind=link}
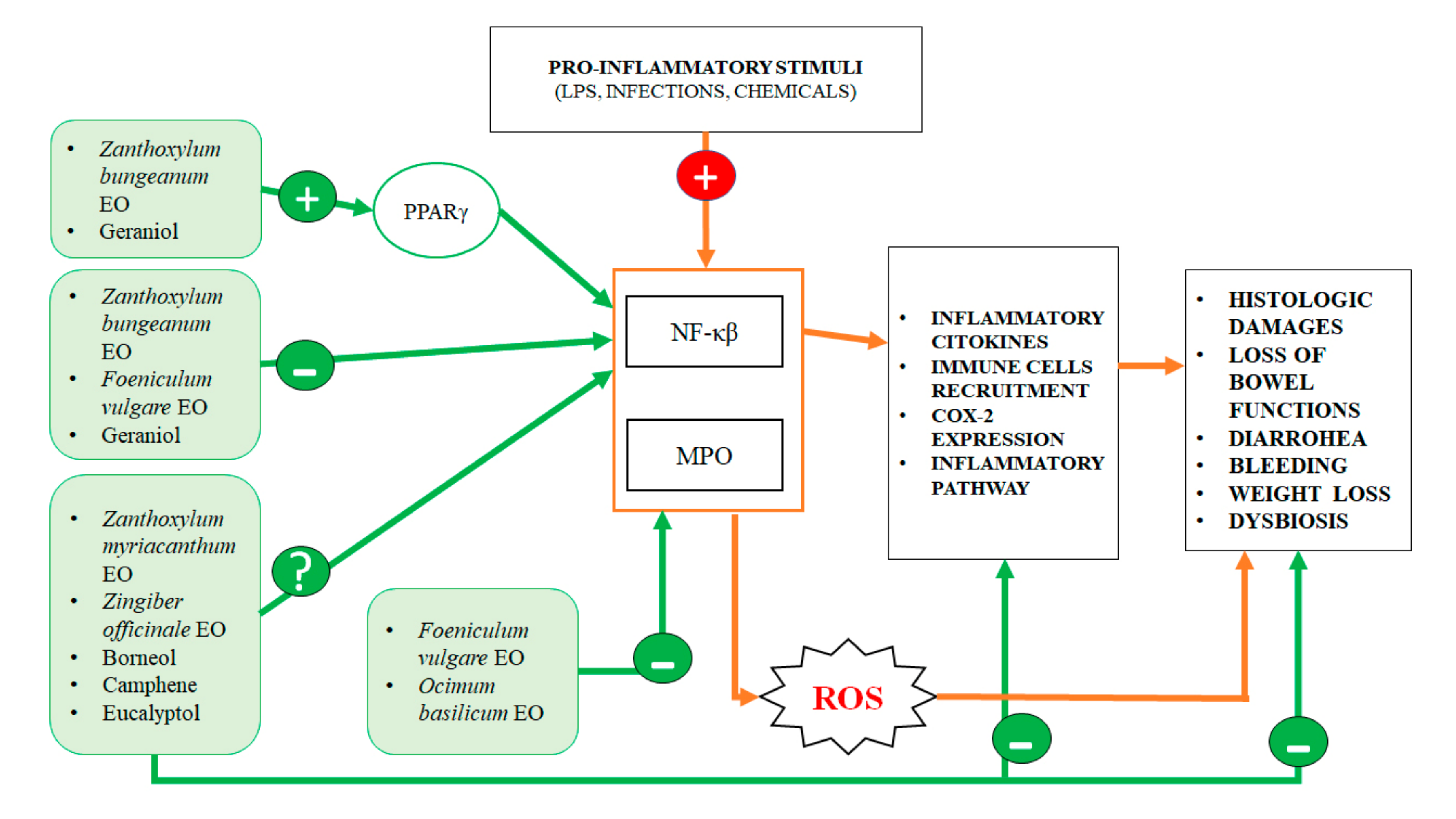{kind=link}
{kind=link}
| EOs Major Single Components | DNEL | NOAEL |
|---|---|---|
| for Oral Administartion (mg/kg bw/day) | ||
| Geraniol | 13.75 | 550 |
| Eugenol | 3 | 300 |
| Carvacrol | 0.0444 | 40 |
| d-limonene | 4.76 | 1000 |
| Cinnamaldehyde | 0.417 | 250 |
| Thymol | 8.3 | 667 |
| Eucalyptol | 600 | 600 |
| Menthol | 4.7 | 188 |
| Essential Oil | Major Compounds and Structures | Major Activity |
|---|---|---|
| Zanthoxylum bungeanum pericarp EO | terpinen-4-ol  | Anti-inflammatory |
eucalyptol  | ||
xanthoxylin  | ||
| Zanthoxylum myriacanthum EO | limonene  | Anti-inflammatory; Microbiota modulation; Fungistatic; |
β-phellandrene  | ||
α-phellandrene  | ||
| Citrus sinensis EO | limonene  | Anti-inflammatory; Microbiota modulation; Fungistatic |
| Foeniculum vulgare EO | trans-anethole  | Anti-inflammatory; |
| Zingiber officinale EO | zingiberene  | Anti-oxidant; Microbiota modulation; |
 α-curcumene α-curcumene | ||
 β-Bisabolene β-Bisabolene | ||
 α-sesquiphellandrene α-sesquiphellandrene | ||
| Ocimum basilicum EO | linalool  | Anti-oxidant; Antiviral; |
β-pinene  | ||
trans-verbenol  | ||
α-terpinolene  | ||
| Oenothera biennis, Rosa chinensis, Helichrysum italicum and Cymbopogon citratus EOs | Geraniol  | Anti-oxidant; Bactericide; |
| Thymus vulgaris EO | thymol  | Anti-oxidant, Bactericide, Bacteriostatic, Microbiota modulation; Fungicidal; Anti-viral; |
| Curcuma longa EO |  curcumin curcumin | Anti-inflammatory |
 α-turmerone α-turmerone | ||
| Rosmarinus officinalis and Eucalyptus spp. EO | eucalyptol  | Bactericide; Antiviral; |
| Origanum vulgare EO | carvacrol (c)  | Anti-oxidant; Bactéricide, Microbiota modulation Antiviral |
| Syzygium aromaticum EO | eugenol  | Bactericide; Biofilm inhibition; Fungicidal, |
| Cinnamomum spp. EO |  cinnamaldehyde cinnamaldehyde | Bacteriostatic, Microbiota modulation; Fungicidal; |
| Mentha spp. EO | menthol  | Bactericide Fungistatic |
| Essential Oil | Main Active Components | Tested Doses | Effect |
|---|---|---|---|
| Zanthoxylum bungeanum | Terpinen-4-ol, eucalyptol, xanthoxylin | 20, 40 and 80 mg/kg body weight/day Effective dose required: all, but 80 mg/kg is most effective | DSS model. Reduced weight loss, DAI and histological damages, proinflammatory cytokines expression (TNF-α, IL-1 β and IL-12) induced by colitis. Decreased NF-kβ levels and increased expression of PPARγ in the colon wall (in vivo) [10] |
| Zanthoxylum myriacanthum | Limonene, β-phellandrene, α-phellandrene, α-pinene and o-cymene | 35 and 70 mg/kg body weight/day Effective dose required: both doses are effective | DSS model. Similar then Z. bungeanum [11] |
| Foeniculum vulgare | Trans-anethole, fenchone, methyl chavicole and limonene | 100, 200 and 400 mg/kg body weight/day Effective dose required: 200 and 400 mg/kg | Acetic Acid model. Reduced histological lesions induced by colitis and expression in mucosal mRNA levels of MPO, TNF-α and NF-kβ (in vivo) [12] |
| Zingiber officinale | Zingiberene, α-curcumene, β-Bisabolene and α-sesquiphellandrene | 100, 200 and 400 mg/kg body weight/day Effective dose required: dose-dependent manner | Acetic Acid model. Reduced the extent of ulcerated areas, immune cell infiltrations and crypt damages induced by colitis (in vivo) [13,14] |
| Ocimum basilicum | Linalool, β-pinene, trans-verbenol and α-terpinolene | 160 and 320 mg/kg body weight/day Effective dose required: both doses are effective | Acetic Acid model. Decreased of histological damages induced by colitis and MPO activity (in vivo) [15] |
| Cymbopogon martini | Geraniol | 30 and 120 mg/kg body weight/day Effective dose required: 120 mg/kg but some effects are detected at lower dose | DSS model. Decreased DAI score, circulating TNF-α, IL-1 β, Il-17, IFNγ and COX-2 mRNA expression. Prevent weight loss, histological damages and dysbiosis induced by colitis (in vivo) [7] |
| Curcuma longa | Curcumin | 25–50 mg /kg body weight/day Effective dose required: both doses are effective | DSS model. Anti-inflammatory cytokines including IL-10 and IL-11 as well as FOXP3 were upregulated [17] |
| Antinflammatory Activity | ||||
| Essential Oil | Main Components | Effects | Mechanism of Action | References |
| Zanthoxylum bungeanum (pericarp) | Terpinen-4-ol, eucalyptol, xanthoxylin | Reduction of: weight loss, DAI, histological damages, tissue TNF-α, IL-1 β and IL-12 raise induced by colitis (in vivo) | Decrease of NF-kβ and increase PPARγ expression | [10] |
| Zanthoxylum myriacanthum | Limonene, β-phellandrene αphellandrene | Reduction of: weight loss, DAI, histological damages, tissue TNF-α, IL-1 β, IL-6 and IL-12 p35 raise induced by colitis (in vivo) | Inhibition of phosphorylation of IKK and IκB | [11] |
| Foeniculum vulgare | Trans-anethole, fenchone, methyl chavicole, limonene | Reduction of the histological lesions induced by colitis (in vivo) | Decrease of MPO, TNF-α and NF-kβ expression | [12] |
| Zingiber officinale (Ginger) | Zingiberene, α-curcumene, β-Bisabolene, α-sesquiphellandrene | prevent colonic tissue damages Induced by colitis (in vivo) | Not specified | [14] |
| Ocimum basilicum (Basil) | Linalool, β-pinene, trans-verbenol, α-terpinolene | Decrease of histological damages Induced by colitis (in vivo) | decrease of the MPO activity | [15] |
| N.A. | Geraniol | decrease of DAI score, preventing weight loss and histological damages and dysbiosis induced by colitis (in vivo) | Regulation of Wnt/β-catenin, p38MAPK, NFκB, PPARγ and COX-2 signaling pathways | [7,19] |
| N.A. | Borneol, camphene and eucalyptol | N.A. | iNOS and COX-2 regulation | [18] |
| Antioxidant Activity | ||||
| Essential Oil | Main Components | Effects | Mechanism of Action | References |
| N.A. | Geraniol | protection from DNA methylation damages (in vitro), reduction of colon inflammation and lipid peroxidation (in vivo) | Decrease of iNOS activity, increase of antioxidant enzymes | [7,20,21,22] |
| Zingiber officinale (Ginger) | Not specified | reduction of intestinal lipid peroxidation, in a model of induce free radicals neutralization (in vivo) | increase of antioxidant enzymes and serum glutathione levels | [130] |
| N.A. | Carvacrol | Reduced colonocyte damages caused by ROS (in vivo) | Not specified | [131] |
| N.A. | Thymol | protection against oxidative DNA damage (in vitro) | Not specified | [121] |
| Ocimum basilicum (Basil) | Not specified | Anti-inflammatory (in vivo) | Reduction of MPO activity | [15] |
| Antibacterial Activity and Microbiota Modulation | ||||
| Essential Oil | Main Components | Effects | Mechanism of Action | References |
| N.A. | Geraniol | Antibacterial against E. aerogenes, E. coli, P. aeruginosa and A. baumannii (in vitro), microbiota modulation (in vivo), boosting of beneficial bacteria (Collinsella and Faecalibacterium) | destabilization of phatogen bacterial cell wall and damage of transmembrane efflux pumps | [7,8,29] |
| N.A. | Eugenol | Antibacterial against E. coli, L. monocytogenes and L. sakei (in vitro), C. rodentium, P. aeruginosa and E. coli (in vivo) | Permeabilization of the bacterial membrane, depletion of ATP, DNA damage, inhibition of the intestinal bacterial adherence, inhibition of quorum sensing | [31,32,33,34] |
| N.A. | Cinnamaldehyde | Bacteriostatic against S. aureus, E. cloacae, A. baumannii and L. monocytogenes, bactericidal against E. coli, E. faecalis, E. faecium, E. aerogenes S. enterica and C. perfringens, C. difficile (in vitro), improvement of the ecological biodiversity (in vivo) | inactivation of its acetyl-CoA carboxylase enzyme | [35,36,37,38,39] |
| N.A. | Thymol | Bacteriostatic against C. difficile, C. perfringens, P. shermanii, P. freudenreichii and B. thetaiotaomicron (in vitro) Bactericide against L. monocytogenes, E. coli, S. enterica, S. aureus, C. botulinum, C. perfringens, S. sonnei, S. lutea, M. flavus, B. hermosphacta, L. innocua, L. monocytogenes, P. putida and S. putrefaciens (in vitro), Modulation of intestinal microbiota (in vivo) | Disruption of bacterial biofilm | [26,30,41,42,44,45] |
| N.A. | Carvacrol | Bactericide against E. coli, Salmonella, Campylobacter spp. (in vivo) Microbiota modulation | Inhibition of bacterial adhesion, invasion and biofilm development | [46,47] |
| N.A. | Limonene | Microbiota modulation | Not specified | [50] |
| Eucalyptus | Eucalyptol | Bactericide against S. aureus, E. coli, B. subtilis, K. pneumonia, S. enteritidis and P. aeruginosa. (in vitro) | Destabilizing and disrupting bacteria membrane | [52] |
| N.A. | Menthol | C.difficile decrease (in vitro) | Barrier disruption and significant leakage of cellular ATP | [54] |
| Antimycotic Activity | ||||
| Essential Oil | Main Components | Effects | Mechanism of Action | References |
| Thymus vulgaris, Citrus limonum, Pelargonium graveolens, Cinnamomum cassia, Ocimum basilicum, and Eugenia caryophyllus | Not specified | Fungistatic and Fungicidal against C. albicans and C. glabrata (in vitro) | Not specified | [64] |
| N.A. | cinnamaldehyde | Inhibition of C.albicans growth (in vitro) | Damage of cell membranes, modulation of potassium ion efflux | [65] |
| N.A. | Limonene | Fungistatic on C.albicans (in vitro) | Inhibition of adhesion, development and maturation of biofilm | [66,67] |
| Mentha | Menthol and (+)-carvone | Fungistatic on C.albicans (in vitro) | Not specified | [68] |
| Caraway | Not specified | Fungistatic on C.albicans (in vitro) | Not specified | [69] |
| Thymus vulgaris | Thymol, carvacrol | antifungal activity against C.albicans, Aspergillus, C.tropicalis and dermatophyte species (in vitro) | Damage of cell membranes | [27,70,71,72] |
| Clove oil | Eugenol | biofilm inhibition of C. albicans (in vitro) | Damage to cell membranes, ATP depletion | [27,73,74,75,76,77] |
| Antiviral Activity | ||||
| Essential Oil | Main Components | Effects | Mechanism of Action | References |
| N.A. | camphor, thymol and eucalyptol | Decreased infectivity of Bovine viral diarrhea virus | Damage of viral particle | [82] |
| N.A. | carvacrol | Reduced propagation of nonenveloped murine norovirus | Damage of viral capsid and RNA | [83] |
| Anticancer Activity | ||||
| Essential Oil | Main Components | Effects | Mechanism of Action | References |
| N.A. | Carvacrol | Antiproliferative effect (in vitro) | Block of cell cycle in G2/M phase, reduction of invasion and migration (HTC-116 and LoVo) | [105] |
| N.A. | Geraniol | Cytotoxic effect (in vitro) Cytostatic effect (in vitro) Regualtion polyamine metabolism (in vitro) Genoprotective effect (in vitro) Antitumoral effect in association with chemotherapeutic agents (in vitro and in vivo) | Induction of apoptosis (Colo-205); Block of cell cycle in S phase, downregulation of ODC and AdoMetDC, Reduction of methylation and ROS DNA damage, sensitization of cells to 5-FU; Inibithion of cell growth by downregulating TS and TK (SW-620) | [106,107,108,113,114] |
| N.A. | Thymol | Cytotoxic effect (in vitro) Genoprotective effect (in vitro) | Induction of ROS production and DNA damages, induction of cell death by mitochondrial pathways (HTC-116); Reduction of methylation and ROS DNA damage | [21,109] |
| N.A. | Cinnamaldehyde | Antiproliferative effect (in vitro) Antitumoral effect in association with chemotherapeutic agents (in vitro and in vivo) | Induction of apoptosis by an increase of Bax/Bcl-2 ratio, inhibition of proliferation by PI3K/Akt pathways, reduction of invasion and migration by increasing E-cadherin levels and downregulation of MMP-2 and MMP-9 (SW-480). Induction of apoptosis in association with camptothecin | [110,111,112] |
| N.A. | Thymoquinone | Antitumoral effect in association with chemotherapeutic agents (in vitro) | Increasing antineoplastic effect of doxorubicin, Increasing cell death by suppressing NF-kβ in association with cisplatin, Induction of autophagy in association with irinotecan | [116,117,118,119] |
| N.A. | β-caryophyllene | Antitumoral effect in association with chemotherapeutic agents, regulation of glucose homeostatis (in vitro) | Increase of anticancer activity of paclitaxel and doxorubicin Regulation of genes involved in glycolysis and cell growth, induction of apoptosis | [119,120,121] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spisni, E.; Petrocelli, G.; Imbesi, V.; Spigarelli, R.; Azzinnari, D.; Donati Sarti, M.; Campieri, M.; Valerii, M.C. Antioxidant, Anti-Inflammatory, and Microbial-Modulating Activities of Essential Oils: Implications in Colonic Pathophysiology. Int. J. Mol. Sci. 2020, 21, 4152. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21114152
Spisni E, Petrocelli G, Imbesi V, Spigarelli R, Azzinnari D, Donati Sarti M, Campieri M, Valerii MC. Antioxidant, Anti-Inflammatory, and Microbial-Modulating Activities of Essential Oils: Implications in Colonic Pathophysiology. International Journal of Molecular Sciences. 2020; 21(11):4152. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21114152
Chicago/Turabian StyleSpisni, Enzo, Giovannamaria Petrocelli, Veronica Imbesi, Renato Spigarelli, Demetrio Azzinnari, Marco Donati Sarti, Massimo Campieri, and Maria Chiara Valerii. 2020. "Antioxidant, Anti-Inflammatory, and Microbial-Modulating Activities of Essential Oils: Implications in Colonic Pathophysiology" International Journal of Molecular Sciences 21, no. 11: 4152. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21114152