Evolutionary Aspects of TRPMLs and TPCs



Walther Straub Institute of Pharmacology and Toxicology, Faculty of Medicine, Ludwig-Maximilians-Universität, 80336 Munich, Germany
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2020, 21(11), 4181; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21114181
Submission received: 14 May 2020
/
Revised: 5 June 2020
/
Accepted: 10 June 2020
/
Published: 11 June 2020
(This article belongs to the Special Issue TRP Channels)
{kind=link}
{kind=link}
Abstract
:Transient receptor potential (TRP) or transient receptor potential channels are a highly diverse family of mostly non-selective cation channels. In the mammalian genome, 28 members can be identified, most of them being expressed predominantly in the plasma membrane with the exception of the mucolipins or TRPMLs which are expressed in the endo-lysosomal system. In mammalian organisms, TRPMLs have been associated with a number of critical endo-lysosomal functions such as autophagy, endo-lysosomal fusion/fission and trafficking, lysosomal exocytosis, pH regulation, or lysosomal motility and positioning. The related non-selective two-pore cation channels (TPCs), likewise expressed in endosomes and lysosomes, have also been found to be associated with endo-lysosomal trafficking, autophagy, pH regulation, or lysosomal exocytosis, raising the question why these two channel families have evolved independently. We followed TRP/TRPML channels and TPCs through evolution and describe here in which species TRP/TRPMLs and/or TPCs are found, which functions they have in different species, and how this compares to the functions of mammalian orthologs.
1. Introduction
The earliest evidence for life on Earth dates back some 3.5 billion years [1]. First, prokaryotes (bacteria and archaea) and certain eukaryotes (protozoa, algae, and fungi) evolved, then gradually plants and animals started developing (their evolution started about 5–25 million years ago). The diversity and differential ability to adapt to distinct environments enabled different species to eventually inhabit even the most extreme areas of planet Earth [2]. The development of a range of sensory modalities was particularly essential for them to navigate through an ever-changing environment [2]. Transient receptor potential (TRP) channels, which mediate cation flow down their electrochemical gradients in response to various environmental stimuli, play fundamental roles for perception in most living organisms. As “cellular sensors”, they are critical for, e.g., vision, olfaction, taste, and temperature-, mechano-, and osmosensation [3,4,5].
The founding member of the TRP family was initially discovered as a receptor-operated sensory cation channel in a blind strain of Drosophila melanogaster [6,7,8], which paved the way for the discovery of the canonical (TRPC1–TRPC7) subfamily of mammalian TRPs [9,10]. Beyond the TRPC channels, the other 21 members of the mammalian TRP family can be grouped in five branches, based on sequence homology: vanilloid (TRPV1-TRPV6), melastatin (TRPM1-8), ankyrin (TRPA1), mucolipin (TRPML1-3), and polycystin (TRPP1-3). Additionally, the TRP channels can be split into two major groups: Group I (TRPC, TRPV, TRPM, and TRPA), and Group II (TRPML and TRPP). The Group II channels appear highly homologous in their transmembrane domains, and express a unique long loop between transmembrane segments S1 and S2 [11]. The existence of subfamilies which are not found in mammals, TRPN (NOMPC-like) expressed in some vertebrates and TRPVL (VL = voltage-like; Cnidaria and Capitella teleta), indicates that TRP channels have been somewhat plastic during metazoan evolution [12].
A common feature of all TRPs is their homotetrameric assembly of six-transmembrane subunits or domains (S1–S6). Unlike other described ion channel families, activation and regulatory mechanisms of TRP channels cannot be reliably predicted based on their subfamily memberships. Often TRPs are activated by multiple stimuli, suggesting that the physiologically relevant stimulus for any given TRP will be governed by the specific cellular context [13,14,15,16,17,18]. Therefore, a functional classification of TRPs may seem more informative. An alternative classification of TRP channels proposed by Zang et al. (2018) combines its physiological function with endogenous activation mechanisms, leading to three subgroups: metabotropic, sensory, and organellar TRPs. Thereby, a functional subgroup can contain members from different subfamilies, important when comparing TRP channels from different species. Fundamental physiological and cellular functions of TRP channels may be the reason for them being highly conserved in yeast and mammals. Nevertheless, TRP channel genes seem to be completely absent from land-plant genomes [12]. In this review, we discuss evolutionary aspects of organellar TRP channels, in particular the TRPML/MCOLN channels, and the functionally related two-pore channels (TPCs).
2. TRPMLs—More and More over Time
Accidental discoveries are often the most significant ones. Such a fortunate accident occurred in 1955, when Christian de Duve discovered the lysosome (Greek: “digestive body”) while attempting to localize the enzyme glucose-6-phosphatase [19]. The lysosome is traditionally understood as an intracellular macromolecule degradation center with a very low pH (approximately 4.5), maintained by lysosomal membrane proton pumps (v-ATPase). Furthermore, the lysosomal digestive capacity is maintained by more than 60 types of acidic hydrolases residing in its lumen [20]. Traditionally, lysosomes are considered to maintain cellular health upon removing toxic cellular components, recycling damaged organelles, terminating signal transduction cascades, and maintaining metabolic homeostasis. Recent studies have added to this view, highlighting their fundamental role as signaling hubs and as hosts of the major nutrient sensors in the cell [21].
Regulated ion fluxes across the lysosomal membrane are a crucial driving force for a number of processes, providing the ionic environment necessary for nutrient homeostasis, osmotic adaptation, or lysosomal trafficking and fusion [22]. Several ion channels controlling the flux of Cl−, K+, Na+, and Ca2+ ions are embedded within lysosomal membranes. Activation of, e.g., the TRPMLs facilitates lysosomal cation efflux, decreasing the lysosomal membrane potential and facilitating v-ATPase proton pumping. Resultantly, the lysosomal resting membrane potential lies between −40 and −20 mV [23,24,25]. These estimates are however a contentious topic, with positive membrane potentials also having been reported [26].
The endo-lysosomal channels have recently gained increasing attention as drug targets implicated in various pathologies [23,27,28,29,30,31,32,33,34]. TRPMLs which are highly critical for endosomal/lysosomal function and autophagy [35,36,37,38] form a separate branch within the superfamily of TRP channels. The TRPML channel family is comprised of three members in mammalian genomes, which share about 75% amino acid sequence similarity [38]. TRPML1, TRPML2, and TRPML3 expression can be detected in early, late, and recycling endosomes as well as in lysosomes to various degrees, allowing flux of cations such as Ca2+ into the cytosol from the endo-lysosomal lumen [11]. All three TRPMLs are activated by the phosphoinositide PI(3,5)P2, a major constituent of endo-lysosomal membranes, and by the luminal pH/proton concentration which differs between various endosomal organelles and lysosomes [27,32,39]. TRPML1 is expressed and present in all tissues, while expression of TRPML2 and TRPML3 appears cell-type specific, indicative of tissue-specific functions of these isoforms [40,41]. Genetic mutations resulting in loss of TRPML1 function by channel inactivation or mislocalization cause a rare genetic disorder called Mucolipidosis Type IV (MLIV), after which the channel initially was named (mucolipin-1, MCOLN1). Loss of MCOLN1 channel function results in a detrimental lysosomal storage disease marked by infantile mental retardation, corneal opacities, strabismus, and delayed motor development [42]. On the other hand, murine gain-of-function TRPML3 mutants result in the varitint-waddler phenotype, marked by hearing and pigmentation defects [43,44,45,46]. Diseases related to mutations in TRPML2 have not yet been described, although the TRPML2 knock-out mouse shows impaired macrophage mobilization attributing to impaired chemoattractant (CCL2) release [32,47].
The number of TRPML channels expressed varies between different species. Thus, studying their evolution may help to better understand their physiological roles and functions. The TRPML1 gene (MCOLN1) is highly conserved in the animal kingdom. Furthermore, many species have more than one TRPML gene. All known genomes of tetrapods have three copies of TRPML channels, with the exception of the domesticated pig (Sus scrofa) where Mcoln3 was found to be a severely truncated pseudogene, lacking five of the six transmembrane domains and the pore-coding region [48]. The human MCOLN1 gene is localized on chromosome 19, distinct from MCOLN2 and MCOLN3, which are closely located on chromosome 1 [49,50]. The MCOLN2 gene is located downstream in the same orientation as MCOLN3, in the genomes of most jawed vertebrates (Gnathostomes), but not pigs, Xenopus tropicalis, and zebrafish [50].
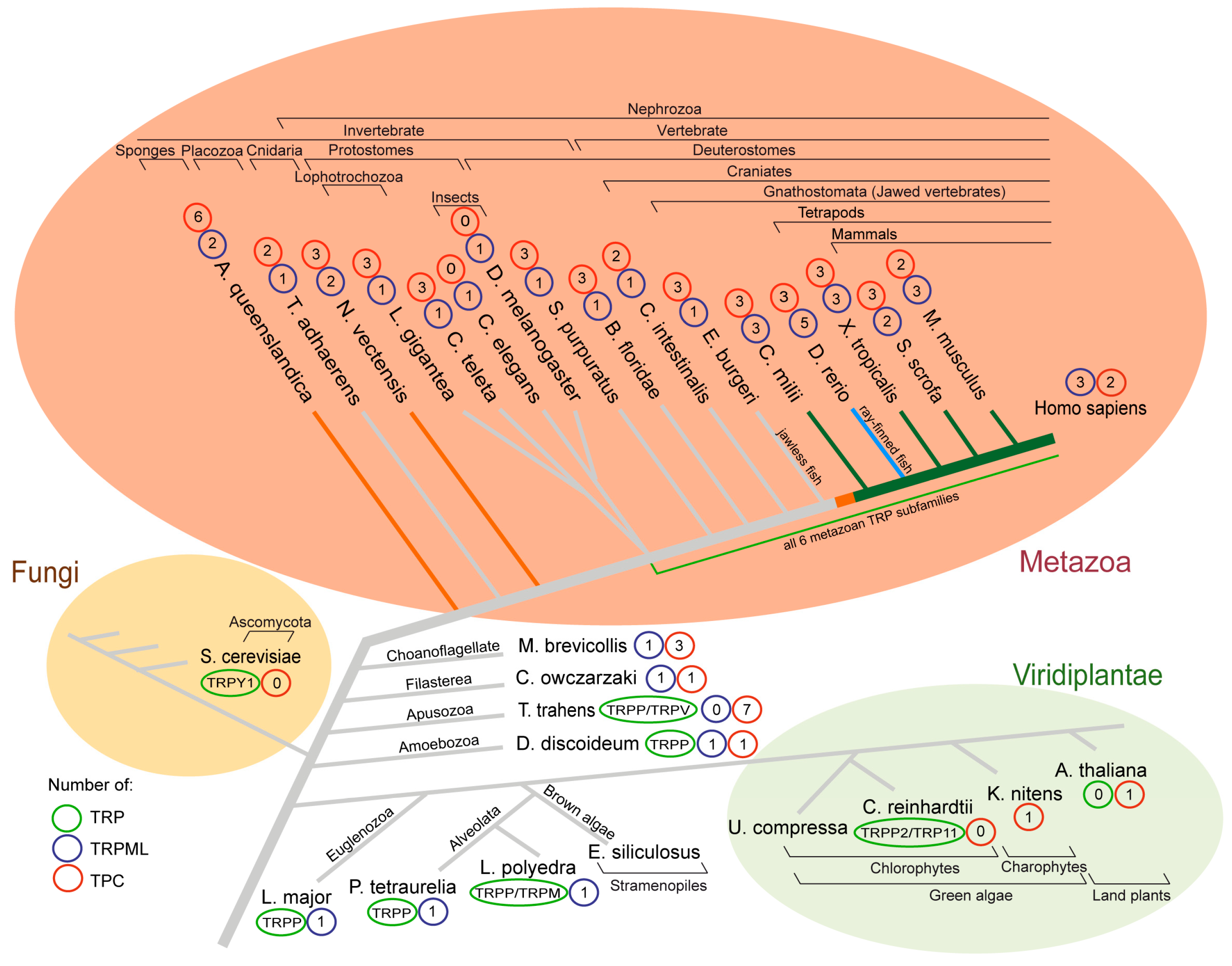
Some deuterostomes as well as protostomes (e.g., Strongylocentrotus purpuratus, Branchiostoma floridae, Ciona intestinalis, Capitella teleta, and Drosophila melanogaster) contain one Mcoln gene ortholog each (Figure 1). García Añoveros et al. (2014) described three episodes of Mcoln gene multiplications in the Nephrozoa clade. In 2017, the inshore hagfish (Eptatretus burgeri) genome was sequenced [51], revealing that the jawless fish genome encodes only one Mcoln homolog (Figure 1—phylogenetic tree; orange branches). It resultantly becomes evident that duplications of the ancestral Mcoln gene, resulting in three paralogs, occurred between the onset of craniates and jawed vertebrates (Gnathostomes) (Figure 1—phylogenetic tree; orange and green branches).
It is interesting to note that homology and localization of the Mcoln2 and Mcoln3 genes suggest their origin resulting from unequal crossing-over in a common ancestor, placing both gene copies in tandem [50]. Similar evolutionary events occurred in the case of closely linked Tmem138 and Tmem216 in tetrapod genomes but not in Xenopus tropicalis and zebrafish [52]. The most recent evolutionary Mcoln multiplication took place in zebrafish (Danio rerio), where a total of five Mcoln homologs can be found: mcoln2 and two mcoln1- and mcoln3-like genes each (Figure 1—phylogenetic tree; navy blue branch) [50].
3. TRPMLs—Evolutionary Beginnings
Sequence comparison of the human TRPML1 protein reveals high similarity to TRPML family members in insects [49]. Drosophila melanogaster encodes a single TRPML family member that shares 44% amino acid identity with human TRPML1 [12,62].
The functional overlap between Drosophila melanogaster Trpml and human TRPML1 becomes particularly evident when comparing associated loss-of-function phenotypes. As previously discussed, loss of TRPML1 in humans causes a lysosomal storage disease marked by prominent mental retardation and impaired motor neuron development [42]. Similarly, disruption of Drosophila melanogaster Trpml (trpml1 mutant) results in neurodegeneration and motor defects, resulting from impaired clearance of apoptotic cells from the fly brain. Mirroring MLIV, trpml1 flies exhibited impaired autophagy marked by accumulating macromolecules [63]. It was later revealed that trpml1 pupae exhibit impaired amphisome/lysosomal fusion and impaired mTORC1 activity, attributable to decreased fly viability [64]. The decreased mTORC1 activity was further linked to impaired development of the neuromuscular junction: Loss of Trpml function, and its associated decrease in Rag GTPases and mTORC1 activity, resulted in decreased JNK phosphorylation and, resultantly, impaired synapse development [65]. Excitingly, a high-protein diet (with the co-administration of an ALK inhibitor, permitting neuronal amino acid uptake) was found to suppress LSD-associated neuromuscular junction defects, appearing a potential therapeutic strategy for lysosomal storage diseases [65]. Although the overlap between trpml1 and MLIV is exciting, it should be noted that primary mouse Mcoln1−/− neurons isolated do not exhibit abnormal mTORC1 activity, underscoring potential differences between species or neuronal subpopulations [66]. It would certainly be intriguing to compare these findings with the human disease.
Insects in most cases express a single TRPML channel; however, Ixodes scapularis and Tetranychus urticae express two and three TRPML channels, respectively, implying potential functional divergence [12]. The Caenorhabditis elegans Mcoln-homolog is termed cup-5 and appears ubiquitously expressed in lysosomes. The null mutation of cup-5 causes maternal effect lethality and lamellar structures similar to those observed in human MLIV cells. Transfection of cup-5 mutant nematodes with human TRPML1 partially rescues their decreased viability, albeit not as well as overexpression of cup-5 itself [67]. Lophotrochozoans (Lottia gigantea and Capitella teleta) appear to be the first species to possess all six metazoan TRP channel subfamilies [12]. Primitive animals, such as sponges (Amphimedon queenslandica), Placozoa (Trichoplax adhaerens), and Cnidaria (Nematostella vectensis), contain only a few cell types, but encode at least one Mcoln gene, suggesting the Mcoln family already started growing around 600 million years ago [12,68,69,70].
Two choanoflagellates (Monosiga brevicollis and Salpingoeca rosetta), free-living unicellular and colonial flagellate eukaryotes considered the closest extant relatives of animals, express five TRP subfamilies, among them TRPML [53,71,72] (Figure 2). Furthermore, a TRPML ortholog has been found in Capsaspora owczarzaki, demonstrating that many metazoan-type TRP channels emerged already in the unicellular common ancestors of Metazoa. In 2012, a study showed that the apusozoan protist Thecamonas trahens expresses TRPP and TRPV channels, suggesting these might represent some of the most ancient metazoan-type TRP channels [53]. TRPMLs and TRPPs have high sequence similarity which may explain why Amoebozoan Dictyostelium discoideum located in the evolutionary tree close to Thecamonas trahens has both of them. Of note, Dictyostelium TRPML is, in contrast to mammalian TRPMLs, required for lysosomal Ca2+ uptake [73]. Only TRPP and TRPML have been reported for euglenozoans, Leishmania (e.g., Leishmania major) and Trypanosoma and the alveolate protist Paramecium tetraurelia, which evolved around 700 million years ago. In another unicellular eukaryotic protist Lingulodinium polyedra TRPMs, TPPMLs and TRPPs have been identified [2,12,54]. In conclusion, TRPML, alongside TRPP, TRPV, and TRPM, seems to have been present in a common ancestor of Unikonts and Bikonts.
4. TRPs in Fungi and Plants—Barely Present
Lysosomes and vacuoles share many similarities in molecular composition and function, e.g., degradation of cellular substances, ion accumulation and storage, and residency of luminal hydrolases, membrane transporters, the vacuolar proton pump and ion channels [74,75,76]. Fungi possess one vacuolar TRP channel, Trpy1p, which shows sequence similarity to other invertebrate/vertebrate TRPML channels of around 16% and 40%, respectively [77]. However, Trpy1p does phylogenetically not cluster with any of the metazoan TRP channels [53]. Trpy1p emerged in fungi upon diverging from metazoan ancestors (Figure 1), appearing structurally distinct from the other lysosomal Group 2 TRP channels, lacking the extensive S1–S2 luminal domain (Figure 2). Saccharomyces cerevisiae Trpy1p, together with green alga Chlamydomonas reinhardtii TRP1, represent the only functionally characterized TRP channels in unicellular organisms [2,55]. Trpy1p acts as a mechanosensor, regulating vacuolar osmotic pressure [55,78]. The well-known mammalian TRPML activator PI(3,5)P2 is key to adaptation of yeast cells to osmotic stress. Upon osmotic stress, yeasts activate several channels and permeases via a transient, 20-fold increase of PI(3,5)P2, resulting in the release of ions and solutes from the lumen of the vacuole. However, there is no proof for direct activation of Trpy1p via PI(3,5)P2 [74,77]. Trpy1p is also linked to increased levels of cytosolic Mn2+, which triggers its activation and Ca2+ efflux in response to oxidative stress [79].
TRPs are found in mammals, worms, insects, and yeast, but appear mysteriously absent from land plants (Figure 1). In green (Ulva compressa) and brown (Ectocarpus siliculosus) algae, application of mammalian TRP channel agonists and antagonists leads to a positive and negative regulation of cellular Ca2+ uptake, respectively [80,81], suggesting existence of TRP channels in 750 million to 1 billion year old phylae [82]. Furthermore, the single-cell alga Chlamydomonas reinhardtii, commonly found in soil and fresh water, has a fine-tuned navigation system based on Ca2+ conductance [83,84]. In 2007, Merchant et al. published the genome of Chlamydomonas reinhardtii [85] and multiple TRP channels have since been identified [2,56]. Arias-Darraz et al. suggested that sequence similarity as well as phylogenetic reconstruction indicate that CrTRP1 could be considered part of a novel family, structurally appearing a homolog to TRPC channels [86]. Interestingly, more than 80% of the putative TRP channels in algae and unicellular organisms appear to originate in TRPP/TRPML clusters, in agreement with TRPP and TRPV channels found in the protist Thecamonas trahens [2,53]. The TRPP/TRPML cluster includes CrTRPP2 and CrTRP11, the latter a distant relative of the TRPV family [56]. CrTRPP2 is structurally more similar to mammalian TRP group 2 channels than Saccharomyces cerevisiae Trpy1p (Figure 2). Two independent studies report behavioral changes after knocking down TRP channel transcripts in Chlamydomonas. Both CrTRPP2 and CrTRP11 are expressed in the flagella, affecting algae mating behavior as well as mechanosensory responses [56,87]. TRP channels in algae exhibit properties of TRP channels expressed by multicellular organisms, such as weak voltage dependence, activation by temperature, regulation by PI(4,5)P2 (phosphatidylinositol 4,5-bisphosphate), and pharmacological block by BCTC or gadolinium [56,86,87,88]. Thus, core functional features of metazoan TRP channels appear present in plant ancestors, suggesting that basic TRP characteristics evolved early in the history of eukaryotes [2].
5. TPCs in Plants (Calcium Induced Calcium Release Theory)
In contrast to TRPML channels, TPCs (two-pore channels), the other group of non-selective endo-lysosomal cation channels in mammalian genomes, are widely found in terrestrial (e.g., Arabidopsis thaliana) and marine plants (e.g., Klebsormidium nitens) [89]. All plants harbor at least one TPC gene, already present in the genome of charophytic algae around 793 million years ago [82,90,91]. TPCs belong to the superfamily of voltage-gated ion channels (VIC), and consist of 12 transmembrane domains (S1–S12) subdivided into two shaker-like domains, each of them containing six transmembrane domains (S1–S6) including the voltage-sensing S4 domain and the ion-conducting pore domain between S5 and S6. TPCs probably originate from a gene-duplication event of single-domain NaV channels [91].
TPC1 activity was first shown by Hedrich and Neher in barley mesophyll vacuoles [92]. Upon activation, plant TPC1 provides an ion-conducting pathway for various cations, mainly K+ and Na+ [93]. Plant TPC1 (or SV, slow vacuolar channel, its original name) is modulated by several factors, underpinning its complex regulation. Beside voltage, TPC1 is regulated by calmodulin [94], 14-3-3 proteins [95], kinases and phosphatases [96,97], pH [93,98], redox state [99], and Mg2+ and Ca2+ [92,100]. In addition, natural polyamines (e.g., spermidine [101,102]) and heavy metals [103] have been reported to inhibit ion passage through open TPC1 channels in red beet and radish.
Since loss of TPC1 function does not drastically impair plant growth [104], its physiological role is a matter of debate. However, roots of seedlings exposed to salt treatment show reduced growth in the TPC1 knockout tpc1-2 mutant compared to WT plants [105]. In contrast, TPC1 overexpression increases salt tolerance [105]. Interestingly, salt-triggered propagating Ca2+ signals in the root were attenuated in tpc1-2 mutants, but increased in TPC1 overexpression lines [105]. Furthermore, it was shown that systemic Ca2+ signals, generated upon wounding, were gone upon loss of TPC1 function. This observation pointed to a role of TPC1 in systemic Ca2+ signaling [106]. The Arabidopsis thaliana TPC1 fou2 variant (fatty acid oxygenation upregulated 2) point mutation D454N leads to an increased production of the stress hormone jasmonate, even under non-stressed conditions. The fou2 plants exhibit a strong growth retardation phenotype [107,108], probably originating from the increased vacuolar K+ release due to TPC1 hyperactivity [108] It is important to note that a TPC1-independent pathway of jasmonate signaling has also been postulated [109]. Since TPC1 participates likely indirectly in the generation/modulation of the Ca2+ wave, it seems to be reasonable to suggest a supreme trigger, regulating Ca2+ and K+ fluxes [109]. Vacuolar membrane depolarization may be one of the missing early triggers for jasmonate production. Furthermore, TPC1 is a prerequisite for vacuole membrane excitability [110], thus triggering of vacuolar membrane depolarization in local spots may be an elaborate way to encode more complicated information in long- and short-distance signaling pathways in plants.
The concept of Ca2+-induced Ca2+ release (CICR, calcium induced calcium release), initially proposed by Fabiato et al. (1985) in the animal field, was adapted by Ward and Schroeder (1994) to plant research [111,112,113]. Based on patch-clamp measurements, they postulated cytosolic Ca2+ signals generated by TPC1 in Vicia faba guard cell vacuoles. However, the ionic condition used in this study was far away from the physiological concentration for Ca2+ and K+. By applying non-physiological ionic conditions, TPC1 channel-mediated Ca2+ currents were also recorded in other species, but only in the opposite direction, from cytosol to vacuole [114,115,116]. Furthermore, an inhibitory effect of vacuolar Ca2+ was postulated [117], likely attributable to the highly conserved vacuolar Ca2+-binding motifs of TPC1 [91,118]. To solve the above long-lasting debate, structural models for the different species will be helpful. Of note, the gain-of-function Arabidopsis thaliana TPC1 channel variant (fou2) shows increased vacuolar Ca2+ and slightly lower resting cytosolic Ca2+ levels compared to WT, which would be difficult to reconcile with TPC1 releasing Ca2+ under physiological conditions [98,109]. In sum, the contribution of plant TPC1 to global as well as local Ca2+ signals remains debated and needs to be further evaluated. A similar complex debate exists in the field of mammalian TPC research, where the role of TPCs in endo-lysosomal Ca2+ release remains controversially discussed.
6. TPCs in Metazoa
The general structure of metazoan TPCs is very similar to their green orthologs. The unique TPC1 gene structure (2 × 6 TMDs) contrasts the 1 × 6 TMD channels (e.g., voltage-gated potassium channels or TRP channels) and the 4 × 6 TMD voltage-gated calcium or sodium channels [119,120]. This TPC-like architecture has been found also in unicellular eukaryotes such as diatoms or amoebae, but not in prokaryotes [91]. Remarkably, TPC genes in fungi seem to be lost immediately after separation from the animal lineage (Figure 1) [53]. However, a variable amount of TPC orthologs was found in choanoflagellates (e.g., Monosiga brevicollis [57]) and even in the evolutionarily older Filaserea and Apusozoa—Capsaspora owczarzaki and Thecamonas trahens, respectively [53]. The oldest living animals, sponges, have six homologs of the TPC gene, a higher number compared to other Metazoa (Figure 1). Despite the absence of the TPC1 gene in Drosophila melanogaster and Caenorhabditis elegans, other Protostomes such as Lophotrochozoans (Lottia gigantea and Capitella teleta) encode three orthologs of human TPCs. Deuterostomes such as sea urchin (Strongylocentrotus purpuratus) also possess three TPC isoforms (Figure 1; [121]). Intriguingly, only TPC1 and TPC2 channels are functional in humans and rodents, while TPC3 is a pseudogene in humans and some primates, and it is completely missing in rodents [25]. Two-pore channels 1–3 are localized in endo-lysosomes, but Danio rerio TPC3 can also function in the plasma membrane [122,123]. TPCs form a subfamily of eukaryotic voltage- and ligand-gated cation channels, however the proposed main ligands in plants and animals differ: calcium in plants, compared to phosphatidylinositol (3,5)-bisphosphate (PI(3,5)P2) and nicotinic acid adenine dinucleotide phosphate (NAADP) in animals [92,124,125]. Originally, TPCs were proposed as ion channels involved in NAADP-mediated Ca2+ release from intracellular acidic stores in animals [124]. It has been suggested that TPC-induced Ca2+ release from acidic stores can induce downstream Ca2+ release from the ER through Ca2+ induced Ca2+ release (CICR) [126]. It has also been postulated that mammalian TPCs are not Ca2+ release channels activated by NAADP, but rather Na+ release channels activated by PI(3,5)P2 [23,31]. The discovery of novel lipophilic small molecule agonists for TPCs in two independent high-throughput calcium imaging campaigns [127,128] partially reconciled this debate. Both campaigns revealed that TPC agonists can be identified in calcium imaging experiments, suggesting that TPCs are indeed permeable to Ca2+. Furthermore, agonists were found to differentially affect the Ca2+/Na+ permeability ratio, suggesting that the cation permeability of the channel can be modulated in a ligand-dependent manner, in turn resulting in distinct effects on key lysosomal functions such as lysosomal pH and exocytosis [127].
Loss of TPCs seems to not affect viability, as double TPC1/2 KO mice age normally [129]. There are also no disease-related (point) mutations known for human or rodent TPCs [31]. However, channel function becomes essential under stress conditions such as decreased availability of ATP and nutrients or other challenging conditions such as infection with viruses or bacterial toxins, or certain diets (e.g., high cholesterol diet) [23,25,130,131].
Evidence for a role of TPCs in Ca2+ signaling is also found in other species. Kelu et al. (2018) demonstrated that in zebrafish Ca2+ release via TPC2 from acidic stores/endo-lysosomes is required for the establishment of synchronized activity in primary motor neurons (PMNs), and postulated a role of NAADP/TPC/Ca2+ signaling in skeletal muscle differentiation [132]. Furthermore, Andrew Miller’s group discovered a putative link between ARC1-like (zebra fish ADP ribosyl cyclase (ARC) homolog) and NAADP generation, TPC2, and Ca2+ signaling during zebrafish myogenesis. Knock down or pharmacological inhibition of ARC1-like leads to an attenuation of Ca2+ signaling and disruption of slow muscle cell development [133]. In single- or multi-celled phagotrophic bacterivores, Dictyostelium discoideum TPC2 disruption leads to delayed development and prolonged growth in culture, delaying expression of early developmental genes. Extracellular cAMP-induced Ca2+ signals are delayed in tpc2-disrupted cells, and sensitivity to weak bases is increased, consistent with an increase in vesicular pH [134].
Despite extensive support for the physiological relevance of TPCs in endo-lysosomal Ca2+ signaling, several questions remain surrounding TPC function. It remains to be further elucidated how NAADP activates TPCs, directly or indirectly, and what the physiological consequences of ligand-dependent cation permeability changes in mammalian TPCs are. Furthermore, the physiological role of Na+ release from mammalian TPCs remains unclear, and the up- and downstream signaling pathways of TPCs need to be further elucidated.
7. Summary
Both TRPMLs and TPCs are comparatively old proteins. They coexist in many species with some remarkable exceptions, e.g., TPCs are missing from Caenorhabditis elegans and Drosophila melanogaster, while they are in contrast to TRPMLs present in most plants. Nevertheless, coexistence is highly conserved throughout the Metazoa kingdom and beyond. Unlike mammalian TPCs, plant TPC1 is a Ca2+-regulated, nonselective cation channel, insensitive to PI(3,5)P2 and NAADP [135]. The reasons for such divergent evolutionary developments are not known. In plants, this may be correlated with the complete loss of TRP channels already at early stages of plant evolution. While during evolution, the number of TPC isoforms has steadily decreased from seven in Thecamonas trahens and six in Amphimedon queenslandica to zero in insects and only two in humans, the number of Mcoln genes seems to increase, reaching three and even five isoforms late in evolution (Callorhinchus milii and Danio rerio), suggestive of a possible increasing physiological relevance of TRPML channels during evolution.
Author Contributions
D.J. wrote the manuscript and C.G. edited it. J.B. and E.K. designed the figures. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported, in part, by funding from the German Research Foundation, DFG (project number 239283807, SFB/TRR152 project P04 to C.G, DFG project GR-4315/2-1 to C.G., and DFG project GR-4315/4-1 to C.G.).
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| NAADP | Nicotinic acid adenine dinucleotide phosphate |
| BCTC | N-(4-tert-butyl-phenyl)-4-(3-chloropyridin-2-yl)tetrahydropyrazine-1(2H)-carboxamide |
References
- Schopf, J.W. Fossil evidence of Archaean life. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006, 361, 869–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arias-Darraz, L.; Cabezas, D.; Colenso, C.K.; Alegría-Arcos, M.; Bravo-Moraga, F.; Varas-Concha, I.; Almonacid, D.E.; Madrid, R.; Brauchi, S. A Transient Receptor Potential Ion Channel in Chlamydomonas Shares Key Features with Sensory Transduction-Associated TRP Channels in Mammals. Plant Cell 2015, 27, 177–188. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J. Molecular mechanism of TRP channels. Compr. Physiol. 2013, 3, 221–242. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Hu, M.; Yang, Y.; Xu, H. Organellar TRP channels. Nat. Struct. Mol. Biol. 2018, 25, 1009–1018. [Google Scholar] [CrossRef]
- Jordt, S.-E.; McKemy, D.D.; Julius, D. Lessons from peppers and peppermint: The molecular logic of thermosensation. Curr. Opin. Neurobiol. 2003, 13, 487–492. [Google Scholar] [CrossRef]
- Minke, B.; Wu, C.; Pak, W.L. Induction of photoreceptor voltage noise in the dark in Drosophila mutant. Nature 1975, 258, 84–87. [Google Scholar] [CrossRef]
- Montell, C.; Rubin, G.M. Molecular characterization of the Drosophila trp locus: A putative integral membrane protein required for phototransduction. Neuron 1989, 2, 1313–1323. [Google Scholar] [CrossRef]
- Hardie, R.C.; Minke, B. The trp gene is essential for a light-activated Ca2+ channel in Drosophila photoreceptors. Neuron 1992, 8, 643–651. [Google Scholar] [CrossRef]
- Wes, P.D.; Chevesich, J.; Jeromin, A.; Rosenberg, C.; Stetten, G.; Montell, C. TRPC1, a human homolog of a Drosophila store-operated channel. Proc. Natl. Acad. Sci. USA 1995, 92, 9652–9656. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Chu, P.B.; Peyton, M.; Birnbaumer, L. Molecular cloning of a widely expressed human homologue for the Drosophila trp gene. FEBS Lett. 1995, 373, 193–198. [Google Scholar] [CrossRef] [Green Version]
- Venkatachalam, K.; Montell, C. TRP channels. Annu. Rev. Biochem. 2007, 76, 387–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, G.; Shi, X.; Kadowaki, T. Evolution of TRP channels inferred by their classification in diverse animal species. Mol. Phylogenet. Evol. 2015, 84, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Hoenderop, J.G.J.; Voets, T.; Hoefs, S.; Weidema, F.; Prenen, J.; Nilius, B.; Bindels, R.J.M. Homo- and heterotetrameric architecture of the epithelial Ca2+ channels TRPV5 and TRPV6. EMBO J. 2003, 22, 776–785. [Google Scholar] [CrossRef] [Green Version]
- Cao, E.; Liao, M.; Cheng, Y.; Julius, D. TRPV1 structures in distinct conformations reveal activation mechanisms. Nature 2013, 504, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, C.E.; Armache, J.-P.; Gao, Y.; Cheng, Y.; Julius, D. Structure of the TRPA1 ion channel suggests regulatory mechanisms. Nature 2015, 520, 511–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; She, J.; Zeng, W.; Guo, J.; Xu, H.; Bai, X.-C.; Jiang, Y. Structure of mammalian endolysosomal TRPML1 channel in nanodiscs. Nature 2017, 550, 415–418. [Google Scholar] [CrossRef] [Green Version]
- Tang, Q.; Guo, W.; Zheng, L.; Wu, J.-X.; Liu, M.; Zhou, X.; Zhang, X.; Chen, L. Structure of the receptor-activated human TRPC6 and TRPC3 ion channels. Cell Res. 2018, 28, 746–755. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Wu, M.; Zubcevic, L.; Borschel, W.F.; Lander, G.C.; Lee, S.-Y. Structure of the cold- and menthol-sensing ion channel TRPM8. Science 2018, 359, 237–241. [Google Scholar] [CrossRef] [Green Version]
- de Duve, C.; Pressman, B.C.; Gianetto, R.; Wattiaux, R.; Appelmans, F. Tissue fractionation studies. 6. Intracellular distribution patterns of enzymes in rat-liver tissue. Biochem. J. 1955, 60, 604–617. [Google Scholar] [CrossRef] [Green Version]
- Kolter, T.; Sandhoff, K. Principles of lysosomal membrane digestion: Stimulation of sphingolipid degradation by sphingolipid activator proteins and anionic lysosomal lipids. Annu. Rev. Cell Dev. Biol. 2005, 21, 81–103. [Google Scholar] [CrossRef] [Green Version]
- Ballabio, A.; Bonifacino, J.S. Lysosomes as dynamic regulators of cell and organismal homeostasis. Nat. Rev. Mol. Cell Biol. 2020, 21, 101–118. [Google Scholar] [CrossRef] [PubMed]
- Perera, R.M.; Zoncu, R. The Lysosome as a Regulatory Hub. Annu. Rev. Cell Dev. Biol. 2016, 32, 223–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cang, C.; Zhou, Y.; Navarro, B.; Seo, Y.-J.; Aranda, K.; Shi, L.; Battaglia-Hsu, S.; Nissim, I.; Clapham, D.E.; Ren, D. mTOR regulates lysosomal ATP-sensitive two-pore Na(+) channels to adapt to metabolic state. Cell 2013, 152, 778–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinberg, B.E.; Huynh, K.K.; Brodovitch, A.; Jabs, S.; Stauber, T.; Jentsch, T.J.; Grinstein, S. A cation counterflux supports lysosomal acidification. J. Cell Biol. 2010, 189, 1171–1186. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Ren, D. Lysosomal physiology. Annu. Rev. Physiol. 2015, 77, 57–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Zhang, X.; Gao, Q.; Lawas, M.; Yu, L.; Cheng, X.; Gu, M.; Sahoo, N.; Li, X.; Li, P.; et al. A voltage-dependent K+ channel in the lysosome is required for refilling lysosomal Ca2+ stores. J. Cell Biol. 2017, 216, 1715–1730. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.-P.; Cheng, X.; Mills, E.; Delling, M.; Wang, F.; Kurz, T.; Xu, H. The type IV mucolipidosis-associated protein TRPML1 is an endolysosomal iron release channel. Nature 2008, 455, 992–996. [Google Scholar] [CrossRef] [Green Version]
- Cang, C.; Bekele, B.; Ren, D. The voltage-gated sodium channel TPC1 confers endolysosomal excitability. Nat. Chem. Biol. 2014, 10, 463–469. [Google Scholar] [CrossRef]
- Cang, C.; Aranda, K.; Seo, Y.; Gasnier, B.; Ren, D. TMEM175 Is an Organelle K(+) Channel Regulating Lysosomal Function. Cell 2015, 162, 1101–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimm, C.; Hassan, S.; Wahl-Schott, C.; Biel, M. Role of TRPML and Two-Pore Channels in Endolysosomal Cation Homeostasis. J. Pharmacol. Exp. Ther. 2012, 342, 236–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Zhang, X.; Dong, X.-P.; Samie, M.; Li, X.; Cheng, X.; Goschka, A.; Shen, D.; Zhou, Y.; Harlow, J.; et al. TPC proteins are phosphoinositide- activated sodium-selective ion channels in endosomes and lysosomes. Cell 2012, 151, 372–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plesch, E.; Chen, C.-C.; Butz, E.; Scotto Rosato, A.; Krogsaeter, E.K.; Yinan, H.; Bartel, K.; Keller, M.; Robaa, D.; Teupser, D.; et al. Selective agonist of TRPML2 reveals direct role in chemokine release from innate immune cells. eLife 2018, 7, e39720. [Google Scholar] [CrossRef] [PubMed]
- Moehring, F.; Cowie, A.M.; Menzel, A.D.; Weyer, A.D.; Grzybowski, M.; Arzua, T.; Geurts, A.M.; Palygin, O.; Stucky, C.L. Keratinocytes mediate innocuous and noxious touch via ATP-P2X4 signaling. eLife 2018, 7, e31684. [Google Scholar] [CrossRef]
- Kim, S.W.; Kim, D.H.; Park, K.S.; Kim, M.K.; Park, Y.M.; Muallem, S.; So, I.; Kim, H.J. Palmitoylation controls trafficking of the intracellular Ca2+ channel MCOLN3/TRPML3 to regulate autophagy. Autophagy 2018, 15, 327–340. [Google Scholar] [CrossRef] [Green Version]
- Zeevi, D.A.; Frumkin, A.; Bach, G. TRPML and lysosomal function. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2007, 1772, 851–858. [Google Scholar] [CrossRef] [Green Version]
- Vergarajauregui, S.; Connelly, P.S.; Daniels, M.P.; Puertollano, R. Autophagic dysfunction in mucolipidosis type IV patients. Hum. Mol. Genet. 2008, 17, 2723–2737. [Google Scholar] [CrossRef]
- Medina, D.L.; Di Paola, S.; Peluso, I.; Armani, A.; De Stefani, D.; Venditti, R.; Montefusco, S.; Scotto-Rosato, A.; Prezioso, C.; Forrester, A.; et al. Lysosomal calcium signalling regulates autophagy through calcineurin and TFEB. Nat. Cell Biol. 2015, 17, 288–299. [Google Scholar] [CrossRef] [Green Version]
- Di Paola, S.; Scotto-Rosato, A.; Medina, D.L. TRPML1: The Ca2+ retaker of the lysosome. Cell Calcium 2018, 69, 112–121. [Google Scholar] [CrossRef]
- Chen, C.-C.; Keller, M.; Hess, M.; Schiffmann, R.; Urban, N.; Wolfgardt, A.; Schaefer, M.; Bracher, F.; Biel, M.; Wahl-Schott, C.; et al. A small molecule restores function to TRPML1 mutant isoforms responsible for mucolipidosis type IV. Nat. Commun. 2014, 5, 4681. [Google Scholar] [CrossRef] [Green Version]
- Samie, M.A.; Grimm, C.; Evans, J.A.; Curcio-Morelli, C.; Heller, S.; Slaugenhaupt, S.A.; Cuajungco, M.P. The tissue-specific expression of TRPML2 (MCOLN-2) gene is influenced by the presence of TRPML1. Pflug. Arch. 2009, 459, 79–91. [Google Scholar] [CrossRef] [Green Version]
- Grimm, C.; Butz, E.; Chen, C.-C.; Wahl-Schott, C.; Biel, M. From mucolipidosis type IV to Ebola: TRPML and two-pore channels at the crossroads of endo-lysosomal trafficking and disease. Cell Calcium 2017, 67, 148–155. [Google Scholar] [CrossRef]
- Slaugenhaupt, S.A. The molecular basis of mucolipidosis type IV. Curr. Mol. Med. 2002, 2, 445–450. [Google Scholar] [CrossRef]
- Kim, H.J.; Li, Q.; Tjon-Kon-Sang, S.; So, I.; Kiselyov, K.; Muallem, S. Gain-of-function mutation in TRPML3 causes the mouse Varitint-Waddler phenotype. J. Biol. Chem. 2007, 282, 36138–36142. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Delling, M.; Li, L.; Dong, X.; Clapham, D.E. Activating mutation in a mucolipin transient receptor potential channel leads to melanocyte loss in varitint–waddler mice. Proc. Natl. Acad. Sci. USA 2007, 104, 18321–18326. [Google Scholar] [CrossRef] [Green Version]
- Grimm, C.; Cuajungco, M.P.; Aken, A.F.J.; van Schnee, M.; Jörs, S.; Kros, C.J.; Ricci, A.J.; Heller, S. A helix-breaking mutation in TRPML3 leads to constitutive activity underlying deafness in the varitint-waddler mouse. Proc. Natl. Acad. Sci. USA 2007, 104, 19583–19588. [Google Scholar] [CrossRef] [Green Version]
- Nagata, K.; Zheng, L.; Madathany, T.; Castiglioni, A.J.; Bartles, J.R.; García-Añoveros, J. The varitint-waddler (Va) deafness mutation in TRPML3 generates constitutive, inward rectifying currents and causes cell degeneration. Proc. Natl. Acad. Sci. USA 2008, 105, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Hua, Y.; Vergarajauregui, S.; Diab, H.I.; Puertollano, R. Novel Role of TRPML2 in the Regulation of the Innate Immune Response. J. Immunol. 2015. [Google Scholar] [CrossRef] [Green Version]
- Flores, E.N.; García-Añoveros, J. TRPML2 and the evolution of mucolipins. Adv. Exp. Med. Biol. 2011, 704, 221–228. [Google Scholar] [CrossRef]
- Bargal, R.; Avidan, N.; Ben-Asher, E.; Olender, Z.; Zeigler, M.; Frumkin, A.; Raas-Rothschild, A.; Glusman, G.; Lancet, D.; Bach, G. Identification of the gene causing mucolipidosis type IV. Nat. Genet. 2000, 26, 118–122. [Google Scholar] [CrossRef] [PubMed]
- García-Añoveros, J.; Wiwatpanit, T. TRPML2 and mucolipin evolution. Handb. Exp. Pharmacol. 2014, 222, 647–658. [Google Scholar] [CrossRef]
- Yates, A.D.; Achuthan, P.; Akanni, W.; Allen, J.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Azov, A.G.; Bennett, R.; et al. Ensembl 2020. Nucleic Acids Res. 2020, 48, D682–D688. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, B.; Lee, A.P.; Ravi, V.; Maurya, A.K.; Lian, M.M.; Swann, J.B.; Ohta, Y.; Flajnik, M.F.; Sutoh, Y.; Kasahara, M.; et al. Elephant shark genome provides unique insights into gnathostome evolution. Nature 2014, 505, 174–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, X.; Clapham, D.E. Ancestral Ca2+ signaling machinery in early animal and fungal evolution. Mol. Biol. Evol. 2012, 29, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Lindström, J.B.; Pierce, N.T.; Latz, M.I. Role of TRP Channels in Dinoflagellate Mechanotransduction. Biol. Bull. 2017, 233, 151–167. [Google Scholar] [CrossRef]
- Palmer, C.P.; Zhou, X.L.; Lin, J.; Loukin, S.H.; Kung, C.; Saimi, Y. A TRP homolog in Saccharomyces cerevisiae forms an intracellular Ca2+-permeable channel in the yeast vacuolar membrane. Proc. Natl. Acad. Sci. USA 2001, 98, 7801–7805. [Google Scholar] [CrossRef] [Green Version]
- Fujiu, K.; Nakayama, Y.; Iida, H.; Sokabe, M.; Yoshimura, K. Mechanoreception in motile flagella of Chlamydomonas. Nat. Cell Biol. 2011, 13, 630–632. [Google Scholar] [CrossRef]
- Brailoiu, E.; Churamani, D.; Cai, X.; Schrlau, M.G.; Brailoiu, G.C.; Gao, X.; Hooper, R.; Boulware, M.J.; Dun, N.J.; Marchant, J.S.; et al. Essential requirement for two-pore channel 1 in NAADP-mediated calcium signaling. J. Cell Biol. 2009, 186, 201–209. [Google Scholar] [CrossRef]
- Ward, J.M.; Mäser, P.; Schroeder, J.I. Plant ion channels: Gene families, physiology, and functional genomics analyses. Annu. Rev. Physiol. 2009, 71, 59–82. [Google Scholar] [CrossRef] [Green Version]
- Howe, K.L.; Contreras-Moreira, B.; De Silva, N.; Maslen, G.; Akanni, W.; Allen, J.; Alvarez-Jarreta, J.; Barba, M.; Bolser, D.M.; Cambell, L.; et al. Ensembl Genomes 2020—Enabling non-vertebrate genomic research. Nucleic Acids Res. 2020, 48, D689–D695. [Google Scholar] [CrossRef] [Green Version]
- Rosindell, J.; Harmon, L.J. OneZoom: A Fractal Explorer for the Tree of Life. PLoS Biol. 2012, 10, e1001406. [Google Scholar] [CrossRef] [Green Version]
- Wilczynska, Z.; Happle, K.; Müller-Taubenberger, A.; Schlatterer, C.; Malchow, D.; Fisher, P.R. Release of Ca2+ from the Endoplasmic Reticulum Contributes to Ca2+ Signaling in Dictyostelium discoideum. Eukaryot. Cell 2005, 4, 1513–1525. [Google Scholar] [CrossRef] [Green Version]
- Montell, C. Physiology, Phylogeny, and Functions of the TRP Superfamily of Cation Channels. Sci. STKE 2001, 2001, re1. [Google Scholar] [CrossRef]
- Venkatachalam, K.; Long, A.A.; Elsaesser, R.; Nikolaeva, D.; Broadie, K.; Montell, C. Motor Deficit in a Drosophila Model of Mucolipidosis Type IV due to Defective Clearance of Apoptotic Cells. Cell 2008, 135, 838–851. [Google Scholar] [CrossRef] [Green Version]
- Wong, C.-O.; Li, R.; Montell, C.; Venkatachalam, K. Drosophila TRPML is required for TORC1 activation. Curr. Biol. 2012, 22, 1616–1621. [Google Scholar] [CrossRef] [Green Version]
- Wong, C.-O.; Palmieri, M.; Li, J.; Akhmedov, D.; Chao, Y.; Broadhead, G.T.; Zhu, M.X.; Berdeaux, R.; Collins, C.A.; Sardiello, M.; et al. Diminished MTORC1-Dependent JNK Activation Underlies the Neurodevelopmental Defects Associated with Lysosomal Dysfunction. Cell Rep. 2015, 12, 2009–2020. [Google Scholar] [CrossRef] [Green Version]
- Curcio-Morelli, C.; Charles, F.A.; Micsenyi, M.C.; Cao, Y.; Venugopal, B.; Browning, M.F.; Dobrenis, K.; Cotman, S.L.; Walkley, S.U.; Slaugenhaupt, S.A. Macroautophagy is defective in mucolipin-1-deficient mouse neurons. Neurobiol. Dis. 2010, 40, 370–377. [Google Scholar] [CrossRef] [Green Version]
- Hersh, B.M.; Hartwieg, E.; Horvitz, H.R. The Caenorhabditis elegans mucolipin-like gene cup-5 is essential for viability and regulates lysosomes in multiple cell types. Proc. Natl. Acad. Sci. USA 2002, 99, 4355–4360. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, M.; Begovic, E.; Chapman, J.; Putnam, N.H.; Hellsten, U.; Kawashima, T.; Kuo, A.; Mitros, T.; Salamov, A.; Carpenter, M.L.; et al. The Trichoplax genome and the nature of placozoans. Nature 2008, 454, 955–960. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, M.; Simakov, O.; Chapman, J.; Fahey, B.; Gauthier, M.E.A.; Mitros, T.; Richards, G.S.; Conaco, C.; Dacre, M.; Hellsten, U.; et al. The Amphimedon queenslandica genome and the evolution of animal complexity. Nature 2010, 466, 720–726. [Google Scholar] [CrossRef]
- Putnam, N.H.; Srivastava, M.; Hellsten, U.; Dirks, B.; Chapman, J.; Salamov, A.; Terry, A.; Shapiro, H.; Lindquist, E.; Kapitonov, V.V.; et al. Sea anemone genome reveals ancestral eumetazoan gene repertoire and genomic organization. Science 2007, 317, 86–94. [Google Scholar] [CrossRef] [Green Version]
- Cai, X. Unicellular Ca2+ signaling “toolkit” at the origin of metazoa. Mol. Biol. Evol. 2008, 25, 1357–1361. [Google Scholar] [CrossRef] [Green Version]
- Jegla, T.J.; Zmasek, C.M.; Batalov, S.; Nayak, S.K. Evolution of the human ion channel set. Comb. Chem. High Throughput Screen. 2009, 12, 2–23. [Google Scholar] [CrossRef]
- Lima, W.C.; Leuba, F.; Soldati, T.; Cosson, P. Mucolipin controls lysosome exocytosis in Dictyostelium. J. Cell Sci. 2012, 125, 2315–2322. [Google Scholar] [CrossRef] [Green Version]
- Li, S.C.; Kane, P.M. The yeast lysosome-like vacuole: Endpoint and crossroads. Biochim. Biophys. Acta 2009, 1793, 650–663. [Google Scholar] [CrossRef] [Green Version]
- Hedrich, R.; Marten, I. TPC1-SV channels gain shape. Mol. Plant 2011, 4, 428–441. [Google Scholar] [CrossRef]
- Docampo, R.; de Souza, W.; Miranda, K.; Rohloff, P.; Moreno, S.N.J. Acidocalcisomes? Conserved from bacteria to man. Nat. Rev. Microbiol. 2005, 3, 251–261. [Google Scholar] [CrossRef]
- Venkatachalam, K.; Wong, C.-O.; Zhu, M.X. The Role of TRPMLs in Endolysosomal Trafficking and Function. Cell Calcium 2015, 58, 48–56. [Google Scholar] [CrossRef] [Green Version]
- Denis, V.; Cyert, M.S. Internal Ca2+ release in yeast is triggered by hypertonic shock and mediated by a TRP channel homologue. J. Cell Biol. 2002, 156, 29–34. [Google Scholar] [CrossRef]
- Ruta, L.L.; Nicolau, I.; Popa, C.V.; Farcasanu, I.C. Manganese Suppresses the Haploinsufficiency of Heterozygous trpy1Δ/TRPY1Saccharomyces cerevisiae Cells and Stimulates the TRPY1-Dependent Release of Vacuolar Ca2+ under H2O2 Stress. Cells 2019, 8, 79. [Google Scholar] [CrossRef] [Green Version]
- Gómez, M.; González, A.; Sáez, C.A.; Morales, B.; Moenne, A. Copper-induced activation of TRP channels promotes extracellular calcium entry, activation of CaMs and CDPKs, copper entry and membrane depolarization in Ulva compressa. Front. Plant Sci. 2015, 6, 182. [Google Scholar] [CrossRef] [Green Version]
- González, A.; Sáez, C.A.; Morales, B.; Moenne, A. Copper-induced activation of TRP channels promotes extracellular calcium entry and activation of CaMK, PKA, PKC, PKG and CBLPK leading to increased expression of antioxidant enzymes in Ectocarpus siliculosus. Plant Physiol. Biochem. 2018, 126, 106–116. [Google Scholar] [CrossRef]
- Yoon, H.S.; Hackett, J.D.; Ciniglia, C.; Pinto, G.; Bhattacharya, D. A molecular timeline for the origin of photosynthetic eukaryotes. Mol. Biol. Evol. 2004, 21, 809–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, E.H. Chlamydomonas as a Model Organism. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 363–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegemann, P. Algal sensory photoreceptors. Annu. Rev. Plant Biol. 2008, 59, 167–189. [Google Scholar] [CrossRef]
- Merchant, S.S.; Prochnik, S.E.; Vallon, O.; Harris, E.H.; Karpowicz, S.J.; Witman, G.B.; Terry, A.; Salamov, A.; Fritz-Laylin, L.K.; Maréchal-Drouard, L.; et al. The Chlamydomonas genome reveals the evolution of key animal and plant functions. Science 2007, 318, 245–250. [Google Scholar] [CrossRef] [Green Version]
- McGoldrick, L.L.; Singh, A.K.; Demirkhanyan, L.; Lin, T.-Y.; Casner, R.G.; Zakharian, E.; Sobolevsky, A.I. Structure of the thermo-sensitive TRP channel TRP1 from the alga Chlamydomonas reinhardtii. Nat. Commun. 2019, 10, 1–12. [Google Scholar] [CrossRef]
- Huang, K.; Diener, D.R.; Mitchell, A.; Pazour, G.J.; Witman, G.B.; Rosenbaum, J.L. Function and dynamics of PKD2 in Chlamydomonas reinhardtii flagella. J. Cell Biol. 2007, 179, 501–514. [Google Scholar] [CrossRef] [Green Version]
- Latorre, R.; Zaelzer, C.; Brauchi, S. Structure-functional intimacies of transient receptor potential channels. Q. Rev. Biophys. 2009, 42, 201–246. [Google Scholar] [CrossRef]
- Carpaneto, A.; Cantu’, A.M.; Busch, H.; Gambale, F. Ion channels in the vacuoles of the seagrass Posidonia oceanica. FEBS Lett. 1997, 412, 236–240. [Google Scholar] [CrossRef] [Green Version]
- Wickett, N.J.; Mirarab, S.; Nguyen, N.; Warnow, T.; Carpenter, E.; Matasci, N.; Ayyampalayam, S.; Barker, M.S.; Burleigh, J.G.; Gitzendanner, M.A.; et al. Phylotranscriptomic analysis of the origin and early diversification of land plants. Proc. Natl. Acad. Sci. USA 2014, 111, E4859–E4868. [Google Scholar] [CrossRef] [Green Version]
- Hedrich, R.; Mueller, T.D.; Becker, D.; Marten, I. Structure and Function of TPC1 Vacuole SV Channel Gains Shape. Mol. Plant 2018, 11, 764–775. [Google Scholar] [CrossRef] [Green Version]
- Hedrich, R.; Neher, E. Cytoplasmic calcium regulates voltage-dependent ion channels in plant vacuoles. Nature 1987, 329, 833–836. [Google Scholar] [CrossRef]
- Schulz-Lessdorf, B.; Hedrich, R. Protons and calcium modulate SV-type channels in the vacuolar-lysosomal compartment—Channel interaction with calmodulin inhibitors. Planta 1995, 197, 655–671. [Google Scholar] [CrossRef]
- Weiser, T.; Blum, W.; Bentrup, F.W. Calmodulin regulates the Ca2+-dependent slow-vacuolar ion channel in the tonoplast of Chenopodium rubrum suspension cells. Planta 1991, 185, 440–442. [Google Scholar] [CrossRef]
- Latz, A.; Becker, D.; Hekman, M.; Müller, T.; Beyhl, D.; Marten, I.; Eing, C.; Fischer, A.; Dunkel, M.; Bertl, A.; et al. TPK1, a Ca2+-regulated Arabidopsis vacuole two-pore K(+) channel is activated by 14-3-3 proteins. Plant J. 2007, 52, 449–459. [Google Scholar] [CrossRef]
- Allen, G.J.; Sanders, D. Calcineurin, a Type 2B Protein Phosphatase, Modulates the Ca2+-Permeable Slow Vacuolar Ion Channel of Stomatal Guard Cells. Plant Cell 1995, 7, 1473–1483. [Google Scholar] [CrossRef]
- Bethke, P.C.; Jones, R.L. Ca2+-Calmodulin Modulates Ion Channel Activity in Storage Protein Vacuoles of Barley Aleurone Cells. Plant Cell 1994, 6, 277–285. [Google Scholar] [CrossRef]
- Beyhl, D.; Hörtensteiner, S.; Martinoia, E.; Farmer, E.E.; Fromm, J.; Marten, I.; Hedrich, R. The fou2 mutation in the major vacuolar cation channel TPC1 confers tolerance to inhibitory luminal calcium. Plant J. 2009, 58, 715–723. [Google Scholar] [CrossRef] [Green Version]
- Carpaneto, A.; Cantù, A.M.; Gambale, F. Redox agents regulate ion channel activity in vacuoles from higher plant cells. FEBS Lett. 1999, 442, 129–132. [Google Scholar] [CrossRef] [Green Version]
- Carpaneto, A.; Cantù, A.M.; Gambale, F. Effects of cytoplasmic Mg2+ on slowly activating channels in isolated vacuoles of Beta vulgaris. Planta 2001, 213, 457–468. [Google Scholar] [CrossRef]
- Dobrovinskaya, O.R.; Muñiz, J.; Pottosin, I.I. Asymmetric block of the plant vacuolar Ca2+-permeable channel by organic cations. Eur. Biophys. J. 1999, 28, 552–563. [Google Scholar] [CrossRef]
- Dobrovinskaya, O.R.; Muñiz, J.; Pottosin, I.I. Inhibition of vacuolar ion channels by polyamines. J. Membr. Biol. 1999, 167, 127–140. [Google Scholar] [CrossRef]
- Carpaneto, A. Nickel inhibits the slowly activating channels of radish vacuoles. Eur. Biophys. J. 2003, 32, 60–66. [Google Scholar] [CrossRef]
- Peiter, E.; Maathuis, F.J.M.; Mills, L.N.; Knight, H.; Pelloux, J.; Hetherington, A.M.; Sanders, D. The vacuolar Ca2+-activated channel TPC1 regulates germination and stomatal movement. Nature 2005, 434, 404–408. [Google Scholar] [CrossRef]
- Choi, W.-G.; Toyota, M.; Kim, S.-H.; Hilleary, R.; Gilroy, S. Salt stress-induced Ca2+ waves are associated with rapid, long-distance root-to-shoot signaling in plants. Proc. Natl. Acad. Sci. USA 2014, 111, 6497–6502. [Google Scholar] [CrossRef] [Green Version]
- Kiep, V.; Vadassery, J.; Lattke, J.; Maaß, J.-P.; Boland, W.; Peiter, E.; Mithöfer, A. Systemic cytosolic Ca2+ elevation is activated upon wounding and herbivory in Arabidopsis. New Phytol. 2015, 207, 996–1004. [Google Scholar] [CrossRef]
- Bonaventure, G.; Gfeller, A.; Rodríguez, V.M.; Armand, F.; Farmer, E.E. The fou2 gain-of-function allele and the wild-type allele of Two Pore Channel 1 contribute to different extents or by different mechanisms to defense gene expression in Arabidopsis. Plant Cell Physiol. 2007, 48, 1775–1789. [Google Scholar] [CrossRef]
- Bonaventure, G.; Gfeller, A.; Proebsting, W.M.; Hörtensteiner, S.; Chételat, A.; Martinoia, E.; Farmer, E.E. A gain-of-function allele of TPC1 activates oxylipin biogenesis after leaf wounding in Arabidopsis. Plant J. 2007, 49, 889–898. [Google Scholar] [CrossRef]
- Lenglet, A.; Jaślan, D.; Toyota, M.; Mueller, M.; Müller, T.; Schönknecht, G.; Marten, I.; Gilroy, S.; Hedrich, R.; Farmer, E.E. Control of basal jasmonate signalling and defence through modulation of intracellular cation flux capacity. New Phytol. 2017, 216, 1161–1169. [Google Scholar] [CrossRef] [Green Version]
- Jaślan, D.; Dreyer, I.; Lu, J.; O’Malley, R.; Dindas, J.; Marten, I.; Hedrich, R. Voltage-dependent gating of SV channel TPC1 confers vacuole excitability. Nat. Commun. 2019, 10, 2659. [Google Scholar] [CrossRef] [Green Version]
- Fabiato, A. Simulated calcium current can both cause calcium loading in and trigger calcium release from the sarcoplasmic reticulum of a skinned canine cardiac Purkinje cell. J. Gen. Physiol. 1985, 85, 291–320. [Google Scholar] [CrossRef] [Green Version]
- Fabiato, A. Time and calcium dependence of activation and inactivation of calcium-induced release of calcium from the sarcoplasmic reticulum of a skinned canine cardiac Purkinje cell. J. Gen. Physiol. 1985, 85, 247–289. [Google Scholar] [CrossRef] [Green Version]
- Ward, J.M.; Schroeder, J.I. Calcium-Activated K+ Channels and Calcium-Induced Calcium Release by Slow Vacuolar Ion Channels in Guard Cell Vacuoles Implicated in the Control of Stomatal Closure. Plant Cell 1994, 6, 669–683. [Google Scholar] [CrossRef]
- Ivashikina, N.; Hedrich, R. K+ currents through SV-type vacuolar channels are sensitive to elevated luminal sodium levels. Plant J. 2005, 41, 606–614. [Google Scholar] [CrossRef]
- Gradogna, A.; Scholz-Starke, J.; Gutla, P.V.K.; Carpaneto, A. Fluorescence combined with excised patch: Measuring calcium currents in plant cation channels. Plant J. 2009, 58, 175–182. [Google Scholar] [CrossRef]
- Rienmüller, F.; Beyhl, D.; Lautner, S.; Fromm, J.; Al-Rasheid, K.A.S.; Ache, P.; Farmer, E.E.; Marten, I.; Hedrich, R. Guard cell-specific calcium sensitivity of high density and activity SV/TPC1 channels. Plant Cell Physiol. 2010, 51, 1548–1554. [Google Scholar] [CrossRef] [Green Version]
- Pottosin, I.I.; Tikhonova, L.I.; Hedrich, R.; Schönknecht, G. Slowly activating vacuolar channels can not mediate Ca2+-induced Ca2+ release. Plant J. 1997, 12, 1387–1398. [Google Scholar] [CrossRef]
- Dadacz-Narloch, B.; Beyhl, D.; Larisch, C.; López-Sanjurjo, E.J.; Reski, R.; Kuchitsu, K.; Müller, T.D.; Becker, D.; Schönknecht, G.; Hedrich, R. A novel calcium binding site in the slow vacuolar cation channel TPC1 senses luminal calcium levels. Plant Cell 2011, 23, 2696–2707. [Google Scholar] [CrossRef] [Green Version]
- Ishibashi, K.; Suzuki, M.; Imai, M. Molecular cloning of a novel form (two-repeat) protein related to voltage-gated sodium and calcium channels. Biochem. Biophys. Res. Commun. 2000, 270, 370–376. [Google Scholar] [CrossRef]
- Rahman, T.; Cai, X.; Brailoiu, G.C.; Abood, M.E.; Brailoiu, E.; Patel, S. Two-pore channels provide insight into the evolution of voltage-gated Ca2+ and Na+ channels. Sci. Signal. 2014, 7, ra109. [Google Scholar] [CrossRef] [Green Version]
- Brailoiu, E.; Hooper, R.; Cai, X.; Brailoiu, G.C.; Keebler, M.V.; Dun, N.J.; Marchant, J.S.; Patel, S. An Ancestral Deuterostome Family of Two-pore Channels Mediates Nicotinic Acid Adenine Dinucleotide Phosphate-dependent Calcium Release from Acidic Organelles. J. Biol. Chem. 2010, 285, 2897–2901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchant, J.S.; Patel, S. Two-pore channels at the intersection of endolysosomal membrane traffic. Biochem. Soc. Trans. 2015, 43, 434–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cang, C.; Aranda, K.; Ren, D. A non-inactivating high-voltage-activated two-pore Na+ channel that supports ultra-long action potentials and membrane bistability. Nat. Commun. 2014, 5, 5015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calcraft, P.J.; Ruas, M.; Pan, Z.; Cheng, X.; Arredouani, A.; Hao, X.; Tang, J.; Rietdorf, K.; Teboul, L.; Chuang, K.-T.; et al. NAADP mobilizes calcium from acidic organelles through two-pore channels. Nature 2009, 459, 596–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagostena, L.; Festa, M.; Pusch, M.; Carpaneto, A. The human two-pore channel 1 is modulated by cytosolic and luminal calcium. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Galione, A. NAADP receptors. Cold Spring Harb. Perspect. Biol. 2011, 3, a004036. [Google Scholar] [CrossRef] [Green Version]
- Gerndt, S.; Chen, C.-C.; Chao, Y.-K.; Yuan, Y.; Burgstaller, S.; Scotto Rosato, A.; Krogsaeter, E.; Urban, N.; Jacob, K.; Nguyen, O.N.P.; et al. Agonist-mediated switching of ion selectivity in TPC2 differentially promotes lysosomal function. eLife 2020, 9. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, W.; Li, P.; Calvo, R.; Southall, N.; Hu, X.; Bryant-Genevier, M.; Feng, X.; Geng, Q.; Gao, C.; et al. Agonist-specific voltage-dependent gating of lysosomal two-pore Na+ channels. eLife 2019, 8. [Google Scholar] [CrossRef]
- Ruas, M.; Galione, A.; Parrington, J. Two-Pore Channels: Lessons from Mutant Mouse Models. Messenger (Los Angel) 2015, 4, 4–22. [Google Scholar] [CrossRef] [Green Version]
- Grimm, C.; Holdt, L.M.; Chen, C.-C.; Hassan, S.; Müller, C.; Jörs, S.; Cuny, H.; Kissing, S.; Schröder, B.; Butz, E.; et al. High susceptibility to fatty liver disease in two-pore channel 2-deficient mice. Nat. Commun. 2014, 5, 4699. [Google Scholar] [CrossRef]
- Sakurai, Y.; Kolokoltsov, A.A.; Chen, C.-C.; Tidwell, M.W.; Bauta, W.E.; Klugbauer, N.; Grimm, C.; Wahl-Schott, C.; Biel, M.; Davey, R.A. Ebola virus. Two-pore channels control Ebola virus host cell entry and are drug targets for disease treatment. Science 2015, 347, 995–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelu, J.J.; Webb, S.E.; Galione, A.; Miller, A.L. TPC2-mediated Ca2+ signaling is required for the establishment of synchronized activity in developing zebrafish primary motor neurons. Dev. Biol. 2018, 438, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Kelu, J.J.; Webb, S.E.; Galione, A.; Miller, A.L. Characterization of ADP-ribosyl cyclase 1-like (ARC1-like) activity and NAADP signaling during slow muscle cell development in zebrafish embryos. Dev. Biol. 2019, 445, 211–225. [Google Scholar] [CrossRef] [PubMed]
- Chang, F.-S.; Wang, Y.; Dmitriev, P.; Gross, J.; Galione, A.; Pears, C. A two-pore channel protein required for regulating mTORC1 activity on starvation. BMC Biol. 2020, 18, 8. [Google Scholar] [CrossRef] [PubMed]
- Boccaccio, A.; Scholz-Starke, J.; Hamamoto, S.; Larisch, N.; Festa, M.; Gutla, P.V.K.; Costa, A.; Dietrich, P.; Uozumi, N.; Carpaneto, A. The phosphoinositide PI(3,5)P₂ mediates activation of mammalian but not plant TPC proteins: Functional expression of endolysosomal channels in yeast and plant cells. Cell. Mol. Life Sci. 2014, 71, 4275–4283. [Google Scholar] [CrossRef]
Figure 1.
Phylogenetic tree indicating the number of genes encoding TRPML and TPC proteins. Numbers of TRP, TRPML, and TPC proteins of selected species of the Metazoa, Fungi, and Viridiplantae as well as selected ancestral species. The tree was constructed based on information included in the following resources [2,12,48,50,51,52,53,54,55,56,57,58,59,60,61], and constructed manually based on this information. Branch color coding of the tree indicates a first (orange), second (bright green), and third (blue) gene duplication of TPRML. Full species names can be found in the text.
Figure 1.
Phylogenetic tree indicating the number of genes encoding TRPML and TPC proteins. Numbers of TRP, TRPML, and TPC proteins of selected species of the Metazoa, Fungi, and Viridiplantae as well as selected ancestral species. The tree was constructed based on information included in the following resources [2,12,48,50,51,52,53,54,55,56,57,58,59,60,61], and constructed manually based on this information. Branch color coding of the tree indicates a first (orange), second (bright green), and third (blue) gene duplication of TPRML. Full species names can be found in the text.

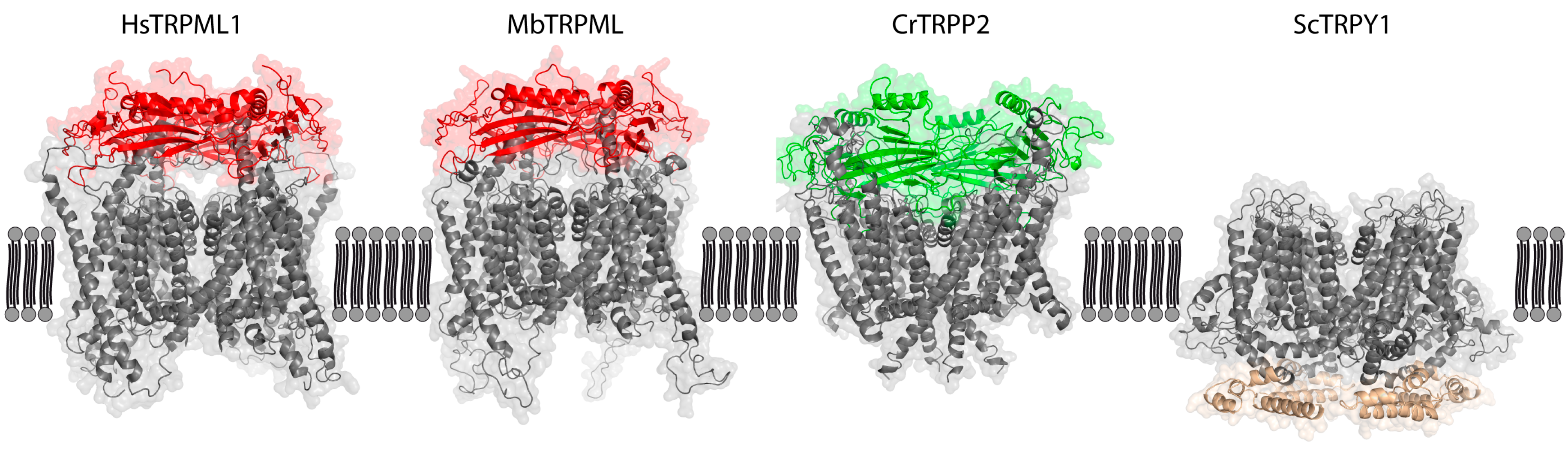
Figure 2.
Main structural differences in loops linking TMD between TRPML ancestors of animal (red), plant (green), and fungi TRPY1 (brown) kingdoms. Amino acid sequences were obtained for Monosiga brevicollis MbTRPML (A9UQ01), Chlamydomonas reinhardtii CrTRPP2 (ABR14113), and Saccharomyces cerevisiae ScTRPY1 (Q12324), and aligned to resolved proteins using SWISS-MODEL (https://swissmodel.expasy.org/). Homology models were constructed using 6BCO (Mus musculus TRPM4) as a template for ScTRPY1, 6T9N (Homo sapiens Polycystin-2) for CrTRPP2 and 6E7P (Homo sapiens TRPML1) for MbTRPML, and visualized using PyMOL. Only MbTRPML aligned well with human TRPML1. Red and green structures indicate the mucolipin or polycystin domains of TRPML and TRPP, respectively, while the TRPY1-unique intracellular domain of ScTRPY1 is colored brown. Having constructed the homology models, we interrogated the possibility of vacuolar channel sorting. Putative lysosomal targeting signals (LTS) were identified using a custom Prosite scan (https://prosite.expasy.org/cgi-bin/prosite/), searching for motifs DxxLL, [DE] xxxL [LI], DxxLM, DxxMV, IMxxYxxL (plant), and YxxL (yeast). Only cytosolic LTS were considered relevant. TRPY1 bears three yeast LTS (Y107-L110, Y293-L296, and Y473-L476, YxxL), while CrTRPP2 bears two yeast and one cross-species LTS (Y44-L47 and Y95-L98, YxxL; E1459-L1464, [DE] xxxL [LI]). MbTRPML bears a single cross-species LTS (E500-I505, [DE] xxxL [LI]).
Figure 2.
Main structural differences in loops linking TMD between TRPML ancestors of animal (red), plant (green), and fungi TRPY1 (brown) kingdoms. Amino acid sequences were obtained for Monosiga brevicollis MbTRPML (A9UQ01), Chlamydomonas reinhardtii CrTRPP2 (ABR14113), and Saccharomyces cerevisiae ScTRPY1 (Q12324), and aligned to resolved proteins using SWISS-MODEL (https://swissmodel.expasy.org/). Homology models were constructed using 6BCO (Mus musculus TRPM4) as a template for ScTRPY1, 6T9N (Homo sapiens Polycystin-2) for CrTRPP2 and 6E7P (Homo sapiens TRPML1) for MbTRPML, and visualized using PyMOL. Only MbTRPML aligned well with human TRPML1. Red and green structures indicate the mucolipin or polycystin domains of TRPML and TRPP, respectively, while the TRPY1-unique intracellular domain of ScTRPY1 is colored brown. Having constructed the homology models, we interrogated the possibility of vacuolar channel sorting. Putative lysosomal targeting signals (LTS) were identified using a custom Prosite scan (https://prosite.expasy.org/cgi-bin/prosite/), searching for motifs DxxLL, [DE] xxxL [LI], DxxLM, DxxMV, IMxxYxxL (plant), and YxxL (yeast). Only cytosolic LTS were considered relevant. TRPY1 bears three yeast LTS (Y107-L110, Y293-L296, and Y473-L476, YxxL), while CrTRPP2 bears two yeast and one cross-species LTS (Y44-L47 and Y95-L98, YxxL; E1459-L1464, [DE] xxxL [LI]). MbTRPML bears a single cross-species LTS (E500-I505, [DE] xxxL [LI]).

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Jaślan, D.; Böck, J.; Krogsaeter, E.; Grimm, C. Evolutionary Aspects of TRPMLs and TPCs. Int. J. Mol. Sci. 2020, 21, 4181. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21114181
AMA Style
Jaślan D, Böck J, Krogsaeter E, Grimm C. Evolutionary Aspects of TRPMLs and TPCs. International Journal of Molecular Sciences. 2020; 21(11):4181. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21114181
Chicago/Turabian StyleJaślan, Dawid, Julia Böck, Einar Krogsaeter, and Christian Grimm. 2020. "Evolutionary Aspects of TRPMLs and TPCs" International Journal of Molecular Sciences 21, no. 11: 4181. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21114181
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.