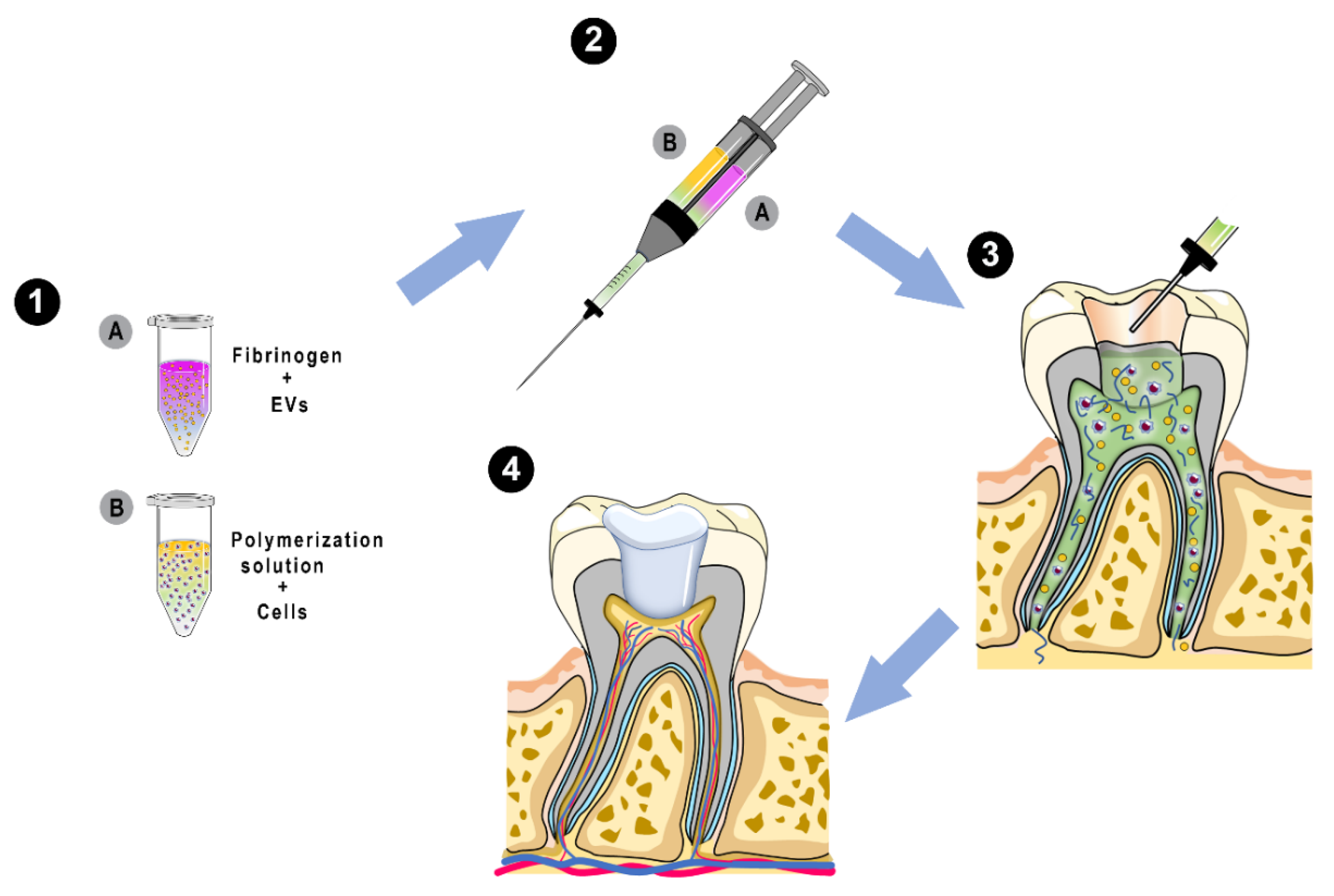
Extracellular Vesicles-Loaded Fibrin Gel Supports Rapid Neovascularization for Dental Pulp Regeneration

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Confirmation that Isolated Dental Pulp Cells Are MSCs
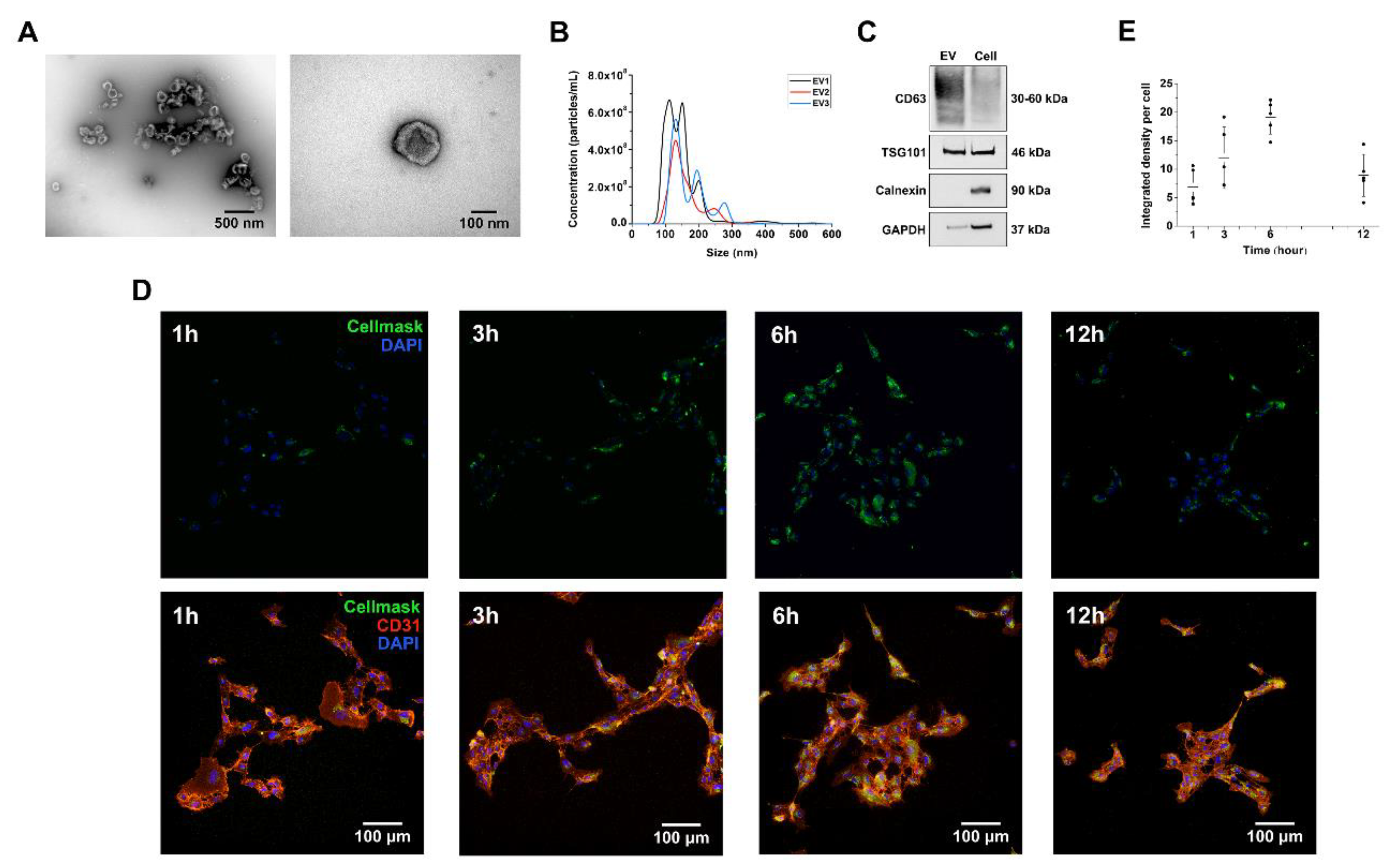
2.2. DPSC-Derived Vesicles Display the Key Characteristics of EVs
2.3. DPSC-Derived EVs Are Internalized by Human Umbilical Vein Endothelial Cells (HUVEC)
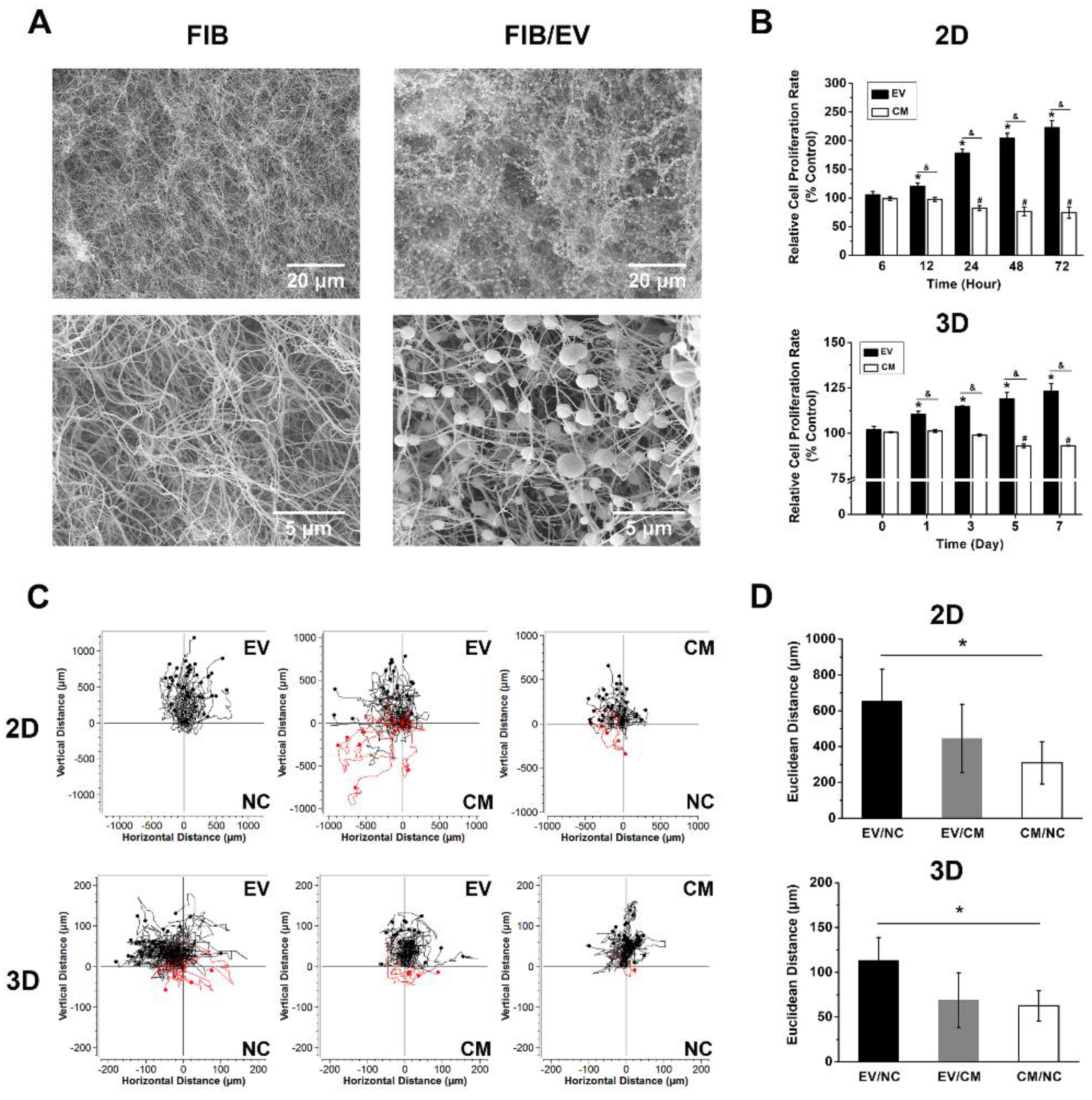
2.4. Fibrin Gels Can Be Homogeneously Loaded with EVs
2.5. DPSC-Derived EVs Enhance Cell Growth in Monolayers and within Fibrin Gels
2.6. DPSC-Derived EVs Enhance HUVEC Migration in Monolayers and Fibrin Gels
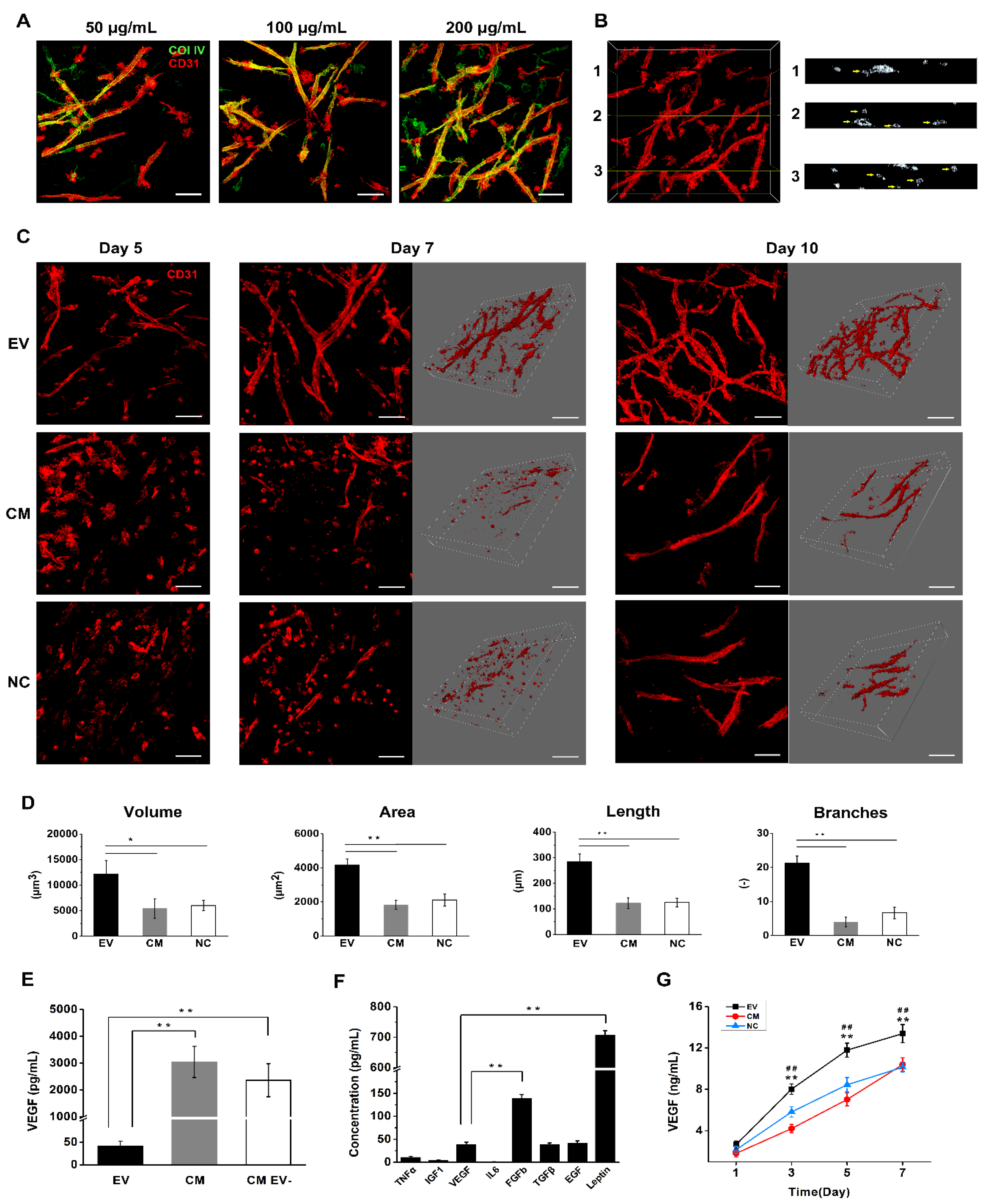
2.7. DPSC-Derived EVs Induce Vascular Tube Formation in Fibrin Gels
2.8. DPSC-Derived EVs Contain VEGF and Several Additional Proangiogenic Factors
2.9. EV-Loaded Fibrin Gels Promote the Secretion of VEGF
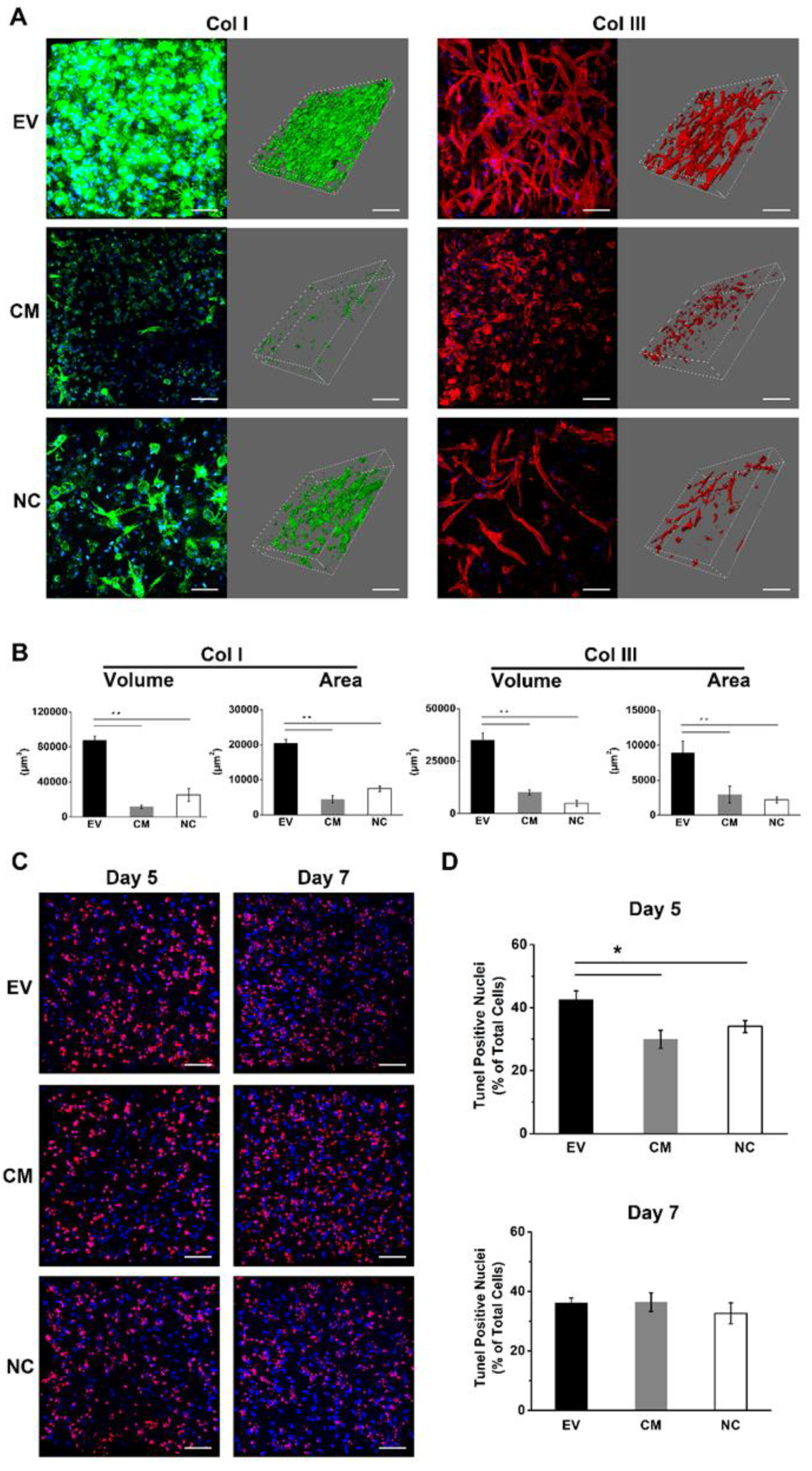
2.10. EV-Loaded Fibrin Gels Stimulate the Deposition of Collagen I and III
2.11. EV-Loaded Fibrin Gels Accelerate the Rate of Apoptosis during Initial Angiogenesis
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Characterization
4.2. Isolation of DPSC-Derived Extracellular Vesicles
4.3. Identification of DPSC-Derived EVs
4.4. EV Labeling and Internalization Assay
4.5. Preparation and Characterization of EV-Loaded Hydrogels
4.6. Cell Proliferation Assay
4.7. Chemotaxis Assay
4.8. Angiogenesis Assay
4.9. Apoptosis Assay
4.10. Release of Angiogenic Factors
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AD-MSCs | Adipose-derived mesenchymal stromal cells |
| BM-MSCs | Bone marrow-derived mesenchymal stromal cells |
| CO2 | Carbon dioxide |
| CM | Conditioned medium |
| DPSCs | Dental pulp stem cells |
| ECM | Extracellular matrix |
| ELISA | Enzyme-linked immunosorbent assay |
| EVs | Extracellular vesicles |
| HUMSCs | Human umbilical cord mesenchymal stromal cells |
| HUVECs | Human umbilical cord veins |
| miRNA | microRNA |
| mRNA | messenger RNA |
| MSCs | Mesenchymal stromal cells |
| NTA | Nanoparticle tracking analysis |
| SEM | Scanning electron microscopy |
| TEM | Transmission electron microscopy |
| TPLSM | Two-photon laser-scanning microscopy |
| VEGF | Vascular endothelial growth factor |
References
- Albuquerque, M.; Valera, M.; Nakashima, M.; Nör, J.; Bottino, M. Tissue-engineering-based strategies for regenerative endodontics. J. Dent. Res. 2014, 93, 1222–1231. [Google Scholar] [CrossRef]
- Auger, F.A.; Gibot, L.; Lacroix, D. The pivotal role of vascularization in tissue engineering. Annu. Rev. Biomed. Eng. 2013, 15, 177–200. [Google Scholar] [CrossRef]
- Dissanayaka, W.L.; Zhang, C. The role of vasculature engineering in dental pulp regeneration. J. Endod. 2017, 43, 102–106. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Shabbir, A.; Cox, A.; Rodriguez-Menocal, L.; Salgado, M.; Badiavas, E.V. Mesenchymal stem cell exosomes induce proliferation and migration of normal and chronic wound fibroblasts, and enhance angiogenesis in vitro. Stem Cells Dev. 2015, 24, 1635–1647. [Google Scholar] [CrossRef]
- Shi, Q.; Qian, Z.; Liu, D.; Sun, J.; Wang, X.; Liu, H.; Xu, J.; Guo, X. GMSC-derived exosomes combined with a chitosan/silk hydrogel sponge accelerates wound healing in a diabetic rat skin defect model. Front. Physiol. 2017, 8, 904. [Google Scholar] [CrossRef] [PubMed]
- Xian, X.; Gong, Q.; Li, C.; Guo, B.; Jiang, H. Exosomes with highly angiogenic potential for possible use in pulp regeneration. J. Endod. 2018, 44, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Nishikawa, M.; Shinotsuka, H.; Matsui, Y.; Ohara, S.; Imai, T.; Takakura, Y. Visualization and in vivo tracking of the exosomes of murine melanoma B16-BL6 cells in mice after intravenous injection. J. Biotechnol. 2013, 165, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Smyth, T.; Kullberg, M.; Malik, N.; Smith-Jones, P.; Graner, M.W.; Anchordoquy, T.J. Biodistribution and delivery efficiency of unmodified tumor-derived exosomes. J. Control. Release 2015, 199, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Wiklander, O.P.; Nordin, J.Z.; O’Loughlin, A.; Gustafsson, Y.; Corso, G.; Mäger, I.; Vader, P.; Lee, Y.; Sork, H.; Seow, Y. Extracellular vesicle in vivo biodistribution is determined by cell source, route of administration and targeting. J. Extracell. Vesicles 2015, 4, 26316. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Liu, X.; Li, H.; Chen, C.; Hu, B.; Niu, X.; Li, Q.; Zhao, B.; Xie, Z.; Wang, Y. Exosomes/tricalcium phosphate combination scaffolds can enhance bone regeneration by activating the PI3K/Akt signaling pathway. Stem Cell Res. Ther. 2016, 7, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Yang, Y.; Li, Y.; Niu, X.; Zhao, B.; Wang, Y.; Bao, C.; Xie, Z.; Lin, Q.; Zhu, L. Integration of stem cell-derived exosomes with in situ hydrogel glue as a promising tissue patch for articular cartilage regeneration. Nanoscale 2017, 9, 4430–4438. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Srivastava, D.; Grover, S.; Sharma, V. Biomaterials in tooth tissue engineering: A review. J. Clin. Diagn. Res. JCDR 2014, 8, 309. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, K.A.; Marques, M.M.; Tedesco, T.K.; Carvalho, G.L.; Gonçalves, F.; Caballero-Flores, H.; Morimoto, S.; Moreira, M.S. Screening of hydrogel-based scaffolds for dental pulp regeneration-A systematic review. Arch. Oral Biol. 2018, 98, 182–194. [Google Scholar] [CrossRef] [PubMed]
- Morin, K.T.; Tranquillo, R.T. In vitro models of angiogenesis and vasculogenesis in fibrin gel. Exp. Cell Res. 2013, 319, 2409–2417. [Google Scholar] [CrossRef] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Gangadaran, P.; Rajendran, R.; Lee, H.; Kalimuthu, S.; Hong, C.; Jeong, S.; Lee, S.-W.; Lee, J.; Ahn, B.-C. Extracellular vesicles from mesenchymal stem cells activates VEGF receptors and accelerates recovery of hindlimb ischemia. J. Control. Release 2017, 264, 112–126. [Google Scholar] [CrossRef]
- Pöschl, E.; Schlötzer-Schrehardt, U.; Brachvogel, B.; Saito, K.; Ninomiya, Y.; Mayer, U. Collagen IV is essential for basement membrane stability but dispensable for initiation of its assembly during early development. Development 2004, 131, 1619–1628. [Google Scholar] [CrossRef] [Green Version]
- Mallat, Z.; Tedgui, A. Apoptosis in the vasculature: Mechanisms and functional importance. Br. J. Pharmacol. 2000, 130, 947–962. [Google Scholar] [CrossRef] [Green Version]
- Bian, S.; Zhang, L.; Duan, L.; Wang, X.; Min, Y.; Yu, H. Extracellular vesicles derived from human bone marrow mesenchymal stem cells promote angiogenesis in a rat myocardial infarction model. J. Mol. Med. 2014, 92, 387–397. [Google Scholar] [CrossRef]
- Lopatina, T.; Mazzeo, A.; Bruno, S.; Tetta, C.; Natalia, K.; Romagnoli, R.; Salizzoni, M.; Porta, M. The angiogenic potential of adipose mesenchymal stem cell-derived extracellular vesicles is modulated by basic fibroblast growth factor. J. Stem Cell Res. Ther. 2014, 4, 245–251. [Google Scholar]
- Zhang, B.; Wu, X.; Zhang, X.; Sun, Y.; Yan, Y.; Shi, H.; Zhu, Y.; Wu, L.; Pan, Z.; Zhu, W. Human umbilical cord mesenchymal stem cell exosomes enhance angiogenesis through the Wnt4/β-catenin pathway. Stem Cells Transl. Med. 2015, 4, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Merckx, G.; Hosseinkhani, B.; Kuypers, S.; Deville, S.; Irobi, J.; Nelissen, I.; Michiels, L.; Lambrichts, I.; Bronckaers, A. Angiogenic effects of human dental pulp and bone marrow-derived mesenchymal stromal cells and their extracellular vesicles. Cells 2020, 9, 312. [Google Scholar] [CrossRef] [Green Version]
- Duval, K.; Grover, H.; Han, L.-H.; Mou, Y.; Pegoraro, A.F.; Fredberg, J.; Chen, Z. Modeling physiological events in 2D vs. 3D cell culture. Physiology 2017, 32, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Deepthi, S.; Jayakumar, R. Alginate nanobeads interspersed fibrin network as in situ forming hydrogel for soft tissue engineering. Bioact. Mater. 2018, 3, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Janmey, P.A.; Winer, J.P.; Weisel, J.W. Fibrin gels and their clinical and bioengineering applications. J. R. Soc. Interface 2008, 6, 1–10. [Google Scholar] [CrossRef]
- Mulcahy, L.A.; Pink, R.C.; Carter, D.R.F. Routes and mechanisms of extracellular vesicle uptake. J. Extracell. Vesicles 2014, 3, 24641. [Google Scholar] [CrossRef] [Green Version]
- Margolis, L.; Sadovsky, Y. The biology of extracellular vesicles: The known unknowns. PLoS Biol. 2019, 17, e3000363. [Google Scholar] [CrossRef]
- Ludwig, N.; Yerneni, S.S.; Azambuja, J.H.; Gillespie, D.G.; Menshikova, E.V.; Jackson, E.K.; Whiteside, T.L. Tumor-derived exosomes promote angiogenesis via adenosine A 2B receptor signaling. Angiogenesis 2020, 1–12. [Google Scholar]
- Banfi, A.; Holnthoner, W.; Martino, M.M.; Ylä-Herttuala, S. Editorial: Vascularization for Regenerative Medicine. Front. Bioeng. Biotechnol. 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Dissanayaka, W.; Hargreaves, K.M.; Jin, L.; Samaranayake, L.P.; Zhang, C. The interplay of dental pulp stem cells and endothelial cells in an injectable peptide hydrogel on angiogenesis and pulp regeneration in vivo. Tissue Eng. Part A 2014, 21, 550–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaessmeyer, S.; Sehl, J.; In, M.; Merle, R.; Richardson, K.; Plendl, J. Subcellular Interactions during Vascular Morphogenesis in 3D Cocultures between Endothelial Cells and Fibroblasts. Int. J. Mol. Sci. 2017, 18, 2590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreimendahl, F.; Köpf, M.; Thiebes, A.L.; Duarte Campos, D.F.; Blaeser, A.; Schmitz-Rode, T.; Apel, C.; Jockenhoevel, S.; Fischer, H. Three-dimensional printing and angiogenesis: Tailored agarose-type I collagen blends comprise three-dimensional printability and angiogenesis potential for tissue-engineered substitutes. Tissue Eng. Part C Methods 2017, 23, 604–615. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, Z.; Lu, W.W.; Zhen, W.; Yang, D.; Peng, S. Novel biomaterial strategies for controlled growth factor delivery for biomedical applications. Npg Asia Mater. 2017, 9, 435. [Google Scholar] [CrossRef]
- Potente, M.; Gerhardt, H.; Carmeliet, P. Basic and therapeutic aspects of angiogenesis. Cell 2011, 146, 873–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagedorn, M.; Balke, M.; Schmidt, A.; Bloch, W.; Kurz, H.; Javerzat, S.; Rousseau, B.; Wilting, J.; Bikfalvi, A. VEGF coordinates interaction of pericytes and endothelial cells during vasculogenesis and experimental angiogenesis. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2004, 230, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Cao, R.; Brakenhielm, E.; Wahlestedt, C.; Thyberg, J.; Cao, Y. Leptin induces vascular permeability and synergistically stimulates angiogenesis with FGF-2 and VEGF. Proc. Natl. Acad. Sci. USA 2001, 98, 6390–6395. [Google Scholar] [CrossRef] [Green Version]
- Ide, S.; Tokuyama, R.; Davaadorj, P.; Shimozuma, M.; Kumasaka, S.; Tatehara, S.; Satomura, K. Leptin and vascular endothelial growth factor regulate angiogenesis in tooth germs. Histochem. Cell Biol. 2011, 135, 281–292. [Google Scholar] [CrossRef]
- Ngo, V.A.; Jung, J.-Y.; Koh, J.-T.; Oh, W.-M.; Hwang, Y.-C.; Lee, B.-N. Leptin induces odontogenic differentiation and angiogenesis in human dental pulp cells via activation of the mitogen-activated protein kinase signaling pathway. J. Endod. 2018, 44, 585–591. [Google Scholar] [CrossRef]
- Bai, Y.; Li, P.; Yin, G.; Huang, Z.; Liao, X.; Chen, X.; Yao, Y. BMP-2, VEGF and bFGF synergistically promote the osteogenic differentiation of rat bone marrow-derived mesenchymal stem cells. Biotechnol. Lett. 2013, 35, 301–308. [Google Scholar] [CrossRef]
- Lee, J.H.; Um, S.; Jang, J.H.; Seo, B.M. Effects of VEGF and FGF-2 on proliferation and differentiation of human periodontal ligament stem cells. Cell Tissue Res. 2012, 348, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Aurora, A.B.; Johnson, B.A.; Qi, X.; McAnally, J.; Hill, J.A.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. The endothelial-specific microRNA miR-126 governs vascular integrity and angiogenesis. Dev. Cell 2008, 15, 261–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suárez, Y.; Sessa, W.C. MicroRNAs as novel regulators of angiogenesis. Circ. Res. 2009, 104, 442–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Culav, E.M.; Clark, C.H.; Merrilees, M.J. Connective tissues: Matrix composition and its relevance to physical therapy. Phys. Ther. 1999, 79, 308–319. [Google Scholar] [CrossRef] [Green Version]
- Linde, A. Session II: Cells and Extracellular Matrices of the Dental Pulp—CT Hanks, Chairman: The Extracellular Matrix of the Dental Pulp and Dentin. J. Dent. Res. 1985, 64, 523–529. [Google Scholar] [CrossRef]
- Duval, H.; Harris, M.; Li, J.; Johnson, N. New insights into the function and regulation of endothelial cell apoptosis. Angiogenesis 2003, 6, 171–183. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Thiebes, A.L.; Kreimendahl, F.; Ruetten, S.; Buhl, E.M.; Wolf, M.; Jockenhoevel, S.; Apel, C. Extracellular Vesicles-Loaded Fibrin Gel Supports Rapid Neovascularization for Dental Pulp Regeneration. Int. J. Mol. Sci. 2020, 21, 4226. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124226
Zhang S, Thiebes AL, Kreimendahl F, Ruetten S, Buhl EM, Wolf M, Jockenhoevel S, Apel C. Extracellular Vesicles-Loaded Fibrin Gel Supports Rapid Neovascularization for Dental Pulp Regeneration. International Journal of Molecular Sciences. 2020; 21(12):4226. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124226
Chicago/Turabian StyleZhang, Siyuan, Anja Lena Thiebes, Franziska Kreimendahl, Stephan Ruetten, Eva Miriam Buhl, Michael Wolf, Stefan Jockenhoevel, and Christian Apel. 2020. "Extracellular Vesicles-Loaded Fibrin Gel Supports Rapid Neovascularization for Dental Pulp Regeneration" International Journal of Molecular Sciences 21, no. 12: 4226. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124226