2.2. Effect of Substrate Concentrations on Fungal Morphology
In this study, the effect of the concentrations of corn steep liquor (CSL) and cassava waste water (CWW) used in 2
2 factorial design (
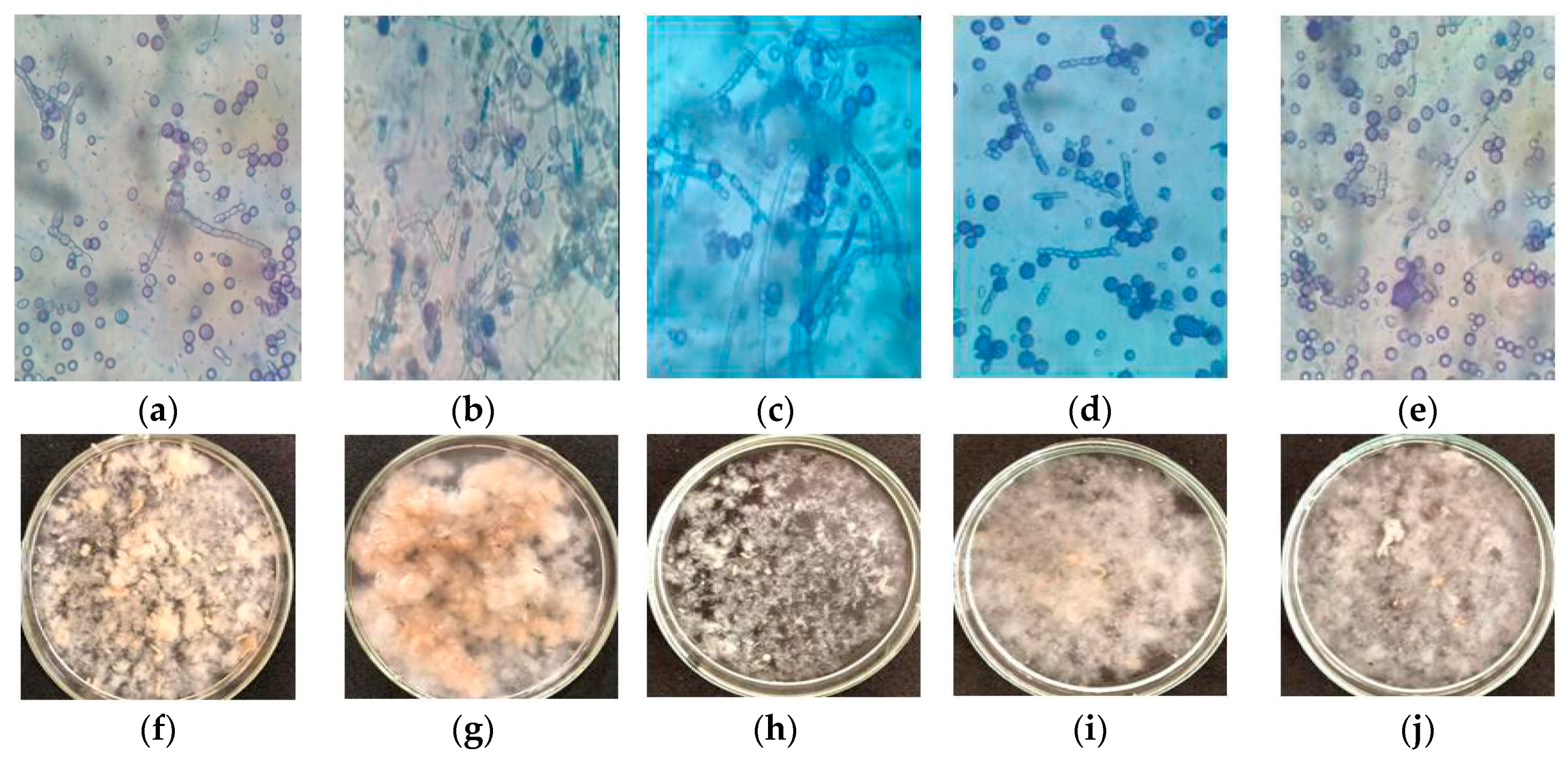
Table 2) on fungal morphology was evaluated. Microscopic and macroscopic images showed cellular differentiation in
M. subtilissimus (UCP 1262) (
Figure 1) and
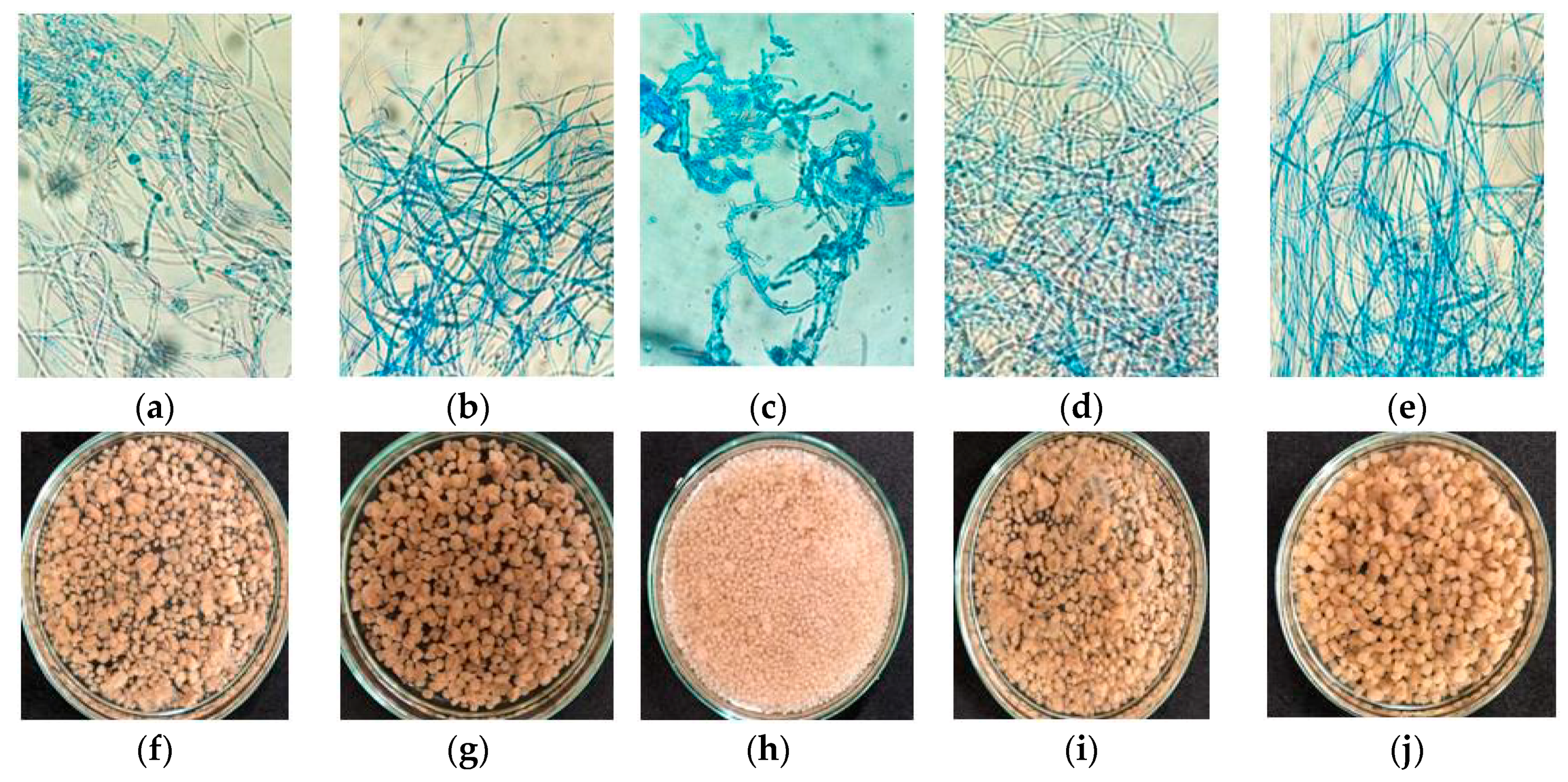
L. hyalospora (UCP 1266) (
Figure 2).
For
M. subtilissimus (UCP 1262), the yeast phase was evident in assays 3 (
Figure 1c) and 4 (
Figure 1d) (higher concentrations of CWW, 8%). In the remaining assays, the transition phase (presence of globose cells, longs/shorts, and arthroposporous hyphae) was observed (
Figure 1). In the assays 1 (
Figure 1a) and point central (
Figure 1e), yeast-like cells and fragments of short arthroposporous hyphae were observed. In assay 2 (
Figure 1b), we observed yeast cells and long arthroposporous hyphae. In addition to mycelium formation, different filamentous fungal species can grow in yeast, depending on environmental conditions. This transition is called dimorphism [
17].
Karimi and Zamani [
18] stated that in addition to factors and mechanisms such as initial spore concentration, sugar concentration, and atmospheric factors evaluated in several studies [
18,
19,
20,
21,
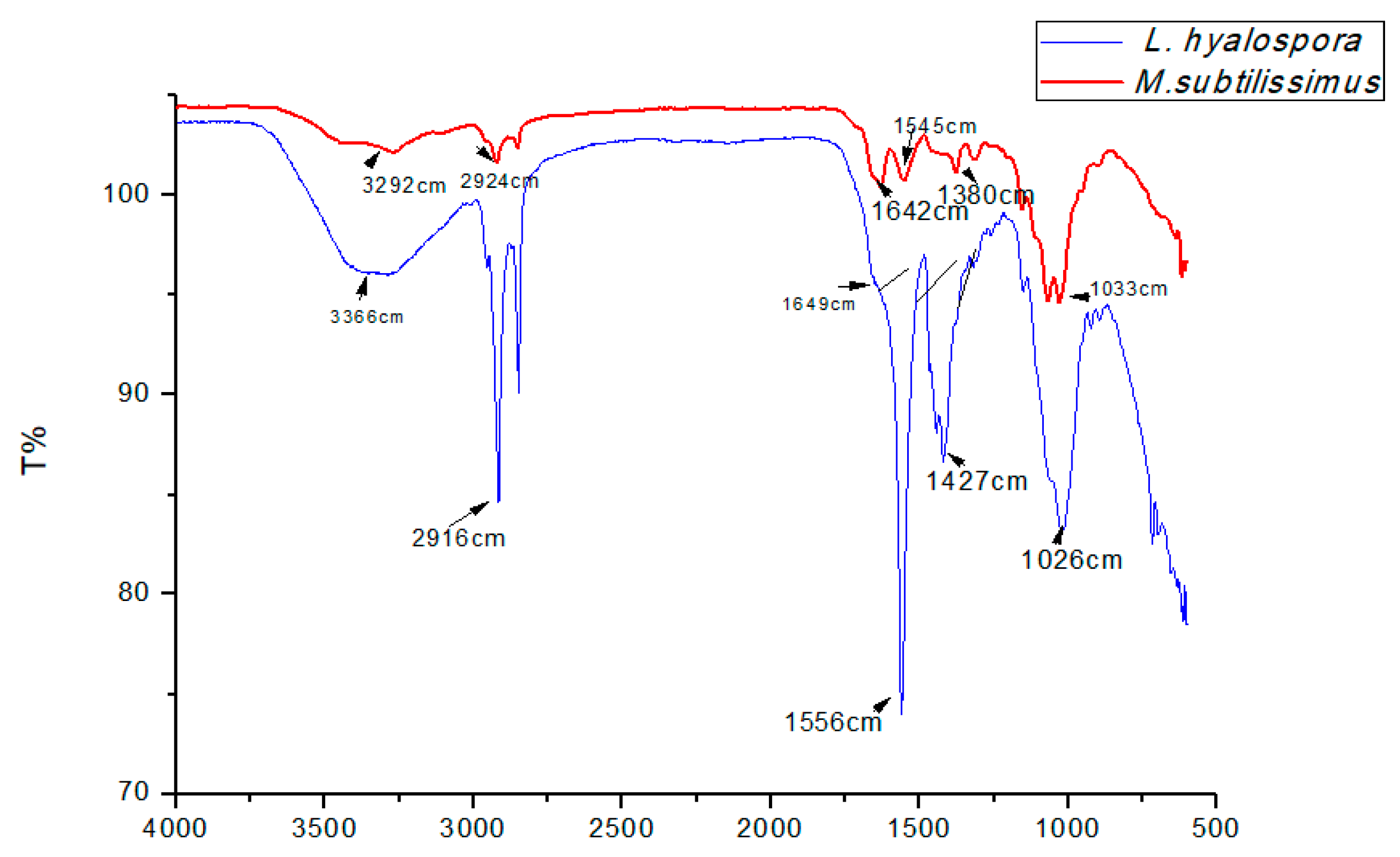
22], fungal morphology may be affected by the addition of certain chemical compounds. Among them, cyanide, acriflavine, and cycloheximide induce yeast morphology due to the inhibition of synthesis or action of cytochrome oxidase. These results can be corroborated in the current study, considering that CWW presents high cyanide concentrations (444.0 mg of cyanide per liter of CWW) [
23,
24].
In submerged cultivation with constant agitation, filamentous fungi exhibit three macroscopic morphologies—scattered mycelia, clumps (small pellets), and pellets (spherical masses of hyphae) [
25,
26]. However, due to the predominance of the yeast and transition phase, for
M. subtilissimus (UCP 1262), the granular aspect was observed under experimental conditions (
Figure 1f–j). In terms of the predominance of granular mycelium, a few areas of differentiation/fragmentation were observed in assays 1 (
Figure 1f) and 2 (
Figure 1g). Assay 3 (
Figure 1i) and center point (
Figure 1j) presented granular mycelium with fragmentation areas. Presence of scattered mycelia with significant presence of areas of differentiation and viscous liquid were observed in assay 3 (
Figure 1h).
Unlike
M. subtilissimus (UCP 1262),
L. hyalospora (UCP 1266) showed no dimorphism. No microscopically relevant differences in agro-industrial waste concentrations were observed (
Figure 2).
L. hyalospora (UCP 1266) produced different sizes and shapes of pellets that differed with agro-industrial waste concentrations. Hollow pellets over 2 mm in diameter were predominant in all assays (
Figure 2f–j), and microscopically with hyphae without cell lysis (
Figure 2a–e), except assay 3 (
Figure 2h; highest CWW level and lower CSL level), with compact and smooth pellets ranging from 1 to 2 mm in diameter and the presence of fragmented hyphae (
Figure 2c). Large and hollow pellets have been observed in several studies [
12,
27,
28,
29], where it has been suggested that autolysis in the center of these clusters of hyphae is caused by difficulties in oxygen transfer.
According to [
25], the composition of the medium can influence the pellet structure, as high concentrations of nitrogen induce pellet formation. Our results differ from those obtained by [
30], who showed that CSL did not influence pellet formation in
Penicillium chrysogenum. However, Metz and Kossen [
25] reported that the type of fungal strain is an important factor influencing pellet formation.
2.3. Effect of Substrates on Biomass and Chitosan Production by M. subtilissimus (UCP 1262) and L. hyalospora (UCP 1266)
Although both fungi showed different patterns in macro-morphology, both showed high potentials in the assimilation of alternative sources of carbon and nitrogen.
Table 3 presents the comparative analysis of the results obtained for biomass and chitosan production in each 2
2 factorial design. The highest biomass yield produced by
M. subtilissimus (UCP 1262; 4.832 mg/L) and
L. hyalospora (UCP 1266; 6.540 mg/L) was in the culture medium supplemented with CSL 6% and CWW 4%, in which
M. subtilissimus (UCP 1262) presented predominance of long and arthrosporous hyphae and
L. hyalospora (UCP 1266) formed pellets of 4 to 5 mm in diameter. The result suggests that media with higher concentrations of CSL favor the growth of the fungi studied. Similar results were reported by Berger et al. (2014), when using the CSL and CWW in the culture medium for growth by
Cunninghamella elegans. CSL is a carbohydrate- and amino acid-rich residue that favors the growth of filamentous and unicellular fungi [
23,
31,
32].
An increase in pH from 6.0 (beginning of fermentation) to 7.1–6.2 was observed in
M. subtilissimus (UCP 1262) cultivation, a result similar to that of [
23,
33,
34]. Lower pH resulted in higher biomass yields, in line with that described by [
23].
However, at the end of
L. hyalospora (UCP 1266) cultivation there was a decrease in pH from 6.0 (initial pH) to 5.1. Similar results were observed by [
35,
36]. Just as low pH favors fungal growth, it also favors chitin deacetylase activity, increasing chitin deacetylation in chitosan [
37,
38].
As for biomass production, assay 2 provided the best chitosan yields by
M. subtilissimus (UCP 1262) (32.47 mg/g), unlike [
23], which obtained the best chitosan yield by
C. elegans in higher concentration of CWW and lower concentration of CSL [
39] when cultivating
C. elegans in medium, with higher concentration of corn steep liquor—CSL (7%) obtaining the best chitosan yield. The study by [
40], when cultivating
Syncephalastrum racemosum in higher concentrations of CSL (8%), obtained the highest yield of biomass and chitosan.
Similar to
M. subtilissimus (UCP 1262), assay 2 (6% CSL and 4% CWW) provided the highest biomass production (6.540 g/L) by
L. hyalospora (UCP 1266). However, it obtained the highest chitosan production (45.03 mg/g) at the central point of the fractional factorial design-FFD (4% of CSL and 6% of CWW). Previously,
Rhizopus arrhizus obtained a higher production of the biopolymer also using 4% of CSL [
41].
Higher concentrations of cassava waste water negatively influenced the chitosan production by
M. subtilissimus (UCP 1262)
, as well as fungal morphology, predominantly forming individual spherical cells and budding multiplication. This yeast phase probably contributed negatively to chitosan production. This assumption is shown in
Figure 1 in assay 3 (2% corn steep liquor and 8% cassava waste water) and assay 4 (6% of corn steep liquor and 8% cassava waste water). The influence of dimorphism on chitosan yield was also observed by [
20], who detected in the cell wall of
M. rouxii (now
Rhizopus arrhizus) that the filament form had higher chitosan yields (9.4%) compared to the yeast phase, which had lower chitosan contents (8.4%).
The same condition that stimulated the
M. subtilissimus (UCP 1262) yeast phase, resulting in low chitosan yields, influenced the formation of compact and smooth pellets by
L. hyalospora (UCP 1266), also causing low biomass and chitosan yields. The best pellet diameter range for chitosan production was 4 to 5 mm, with a content of 45.03 g/kg of dry biomass. In the study of Sparringa et al. [
42] the highest production of glycosamine (107 g/kg), obtained from chitosan deacetylated produced by
Rhizopus oligosporus NRRL 2710, resulted in pellet formation with 16–35 mm. Sparringa and Owens [
43] analyzed the pellet size of affected
R. oryzae ATCC 20,344 glycosamine production. In addition, high glycosamine content (0.19 g/g) was obtained with pellet formation of 5.0 mm, and reduction to 1 mm negatively influenced glycosamine yield to 0.15 g/g.
Ultrastructural aspects of biomasses with higher chitosan yields were also observed by scanning electron microscopy.
Figure 3 shows the mycelial branches of the
Mucor subtilissimus (UCP 1262) (6% CSL/4% CWW) with loose mycelium and hyphae with a thickness that was thinner, tubular, contorted, and with morphological aspect of yeast cells (
Figure 3a) and
Lichtheimia hyalospora (UCP 1266) (assay central point: 4% CSL/6% CWW) with compact mycelium hyphae with a thickness that was thinner, tubular and contorted. (
Figure 3b). This study suggests that
L. hyalospora (UCP 1266) mycelium fragmentation is a result of pellet formation. As other genera of the order Mucorales, they grow rapidly and have tubular-shaped hyphae and no septation. Liao et al. [
44] observed morphological aspects of
Rhizopus oryzae by SEM grown in agroindustrial wastes (soybean meal, wheat, and rice) to understand the relationship between morphology and production of glycosamine, lipids, and amino acids. According to [
45], the culture medium composition and culture conditions influence the metabolic regulation and, consequently, the morphology. The increase in biomass and chitosan is a reliable indicator of the development of the studied fungi.
Table 4 presents the best results from various studies for biomass and chitosan production by Mucoralean fungi, which suggests that the chitosan content of the fungi depends on the fermentation time, culture medium, and cultivation conditions.
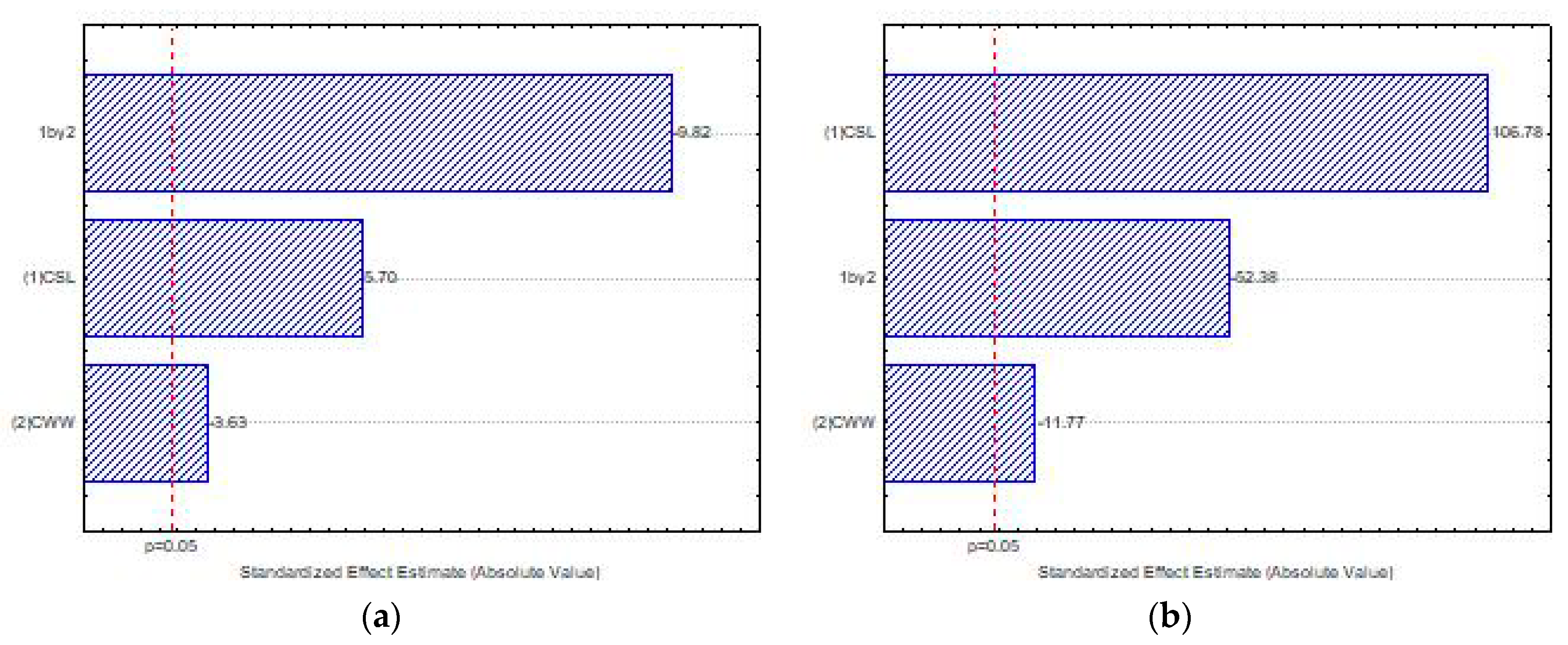
Figure 4 presents the influence of the independent variables, corn steep liquor—CSL (1) and cassava waste water-CWW (2), and the interaction between these variables (1 × 2) on biomass production by
Mucor subtilissimus (UCP 1262) and
Lichtheimia hyalospora (UCP 1266), using factorial design, with statistical significance of
p < 0.05. The Pareto chart illustrates that the increase in the concentration of CSL (1) positively influenced the growth of the microorganisms. However, lower levels of CWW (2) were suggested for biomass production (
Figure 4a,b). The interaction of the independent variables (1 × 2) showed that the maximum level of CSL (1) and the minimum level of CWW (2) had an antagonistic interaction, with significant influence on the biomass production in both microorganisms (
Figure 4a,b).
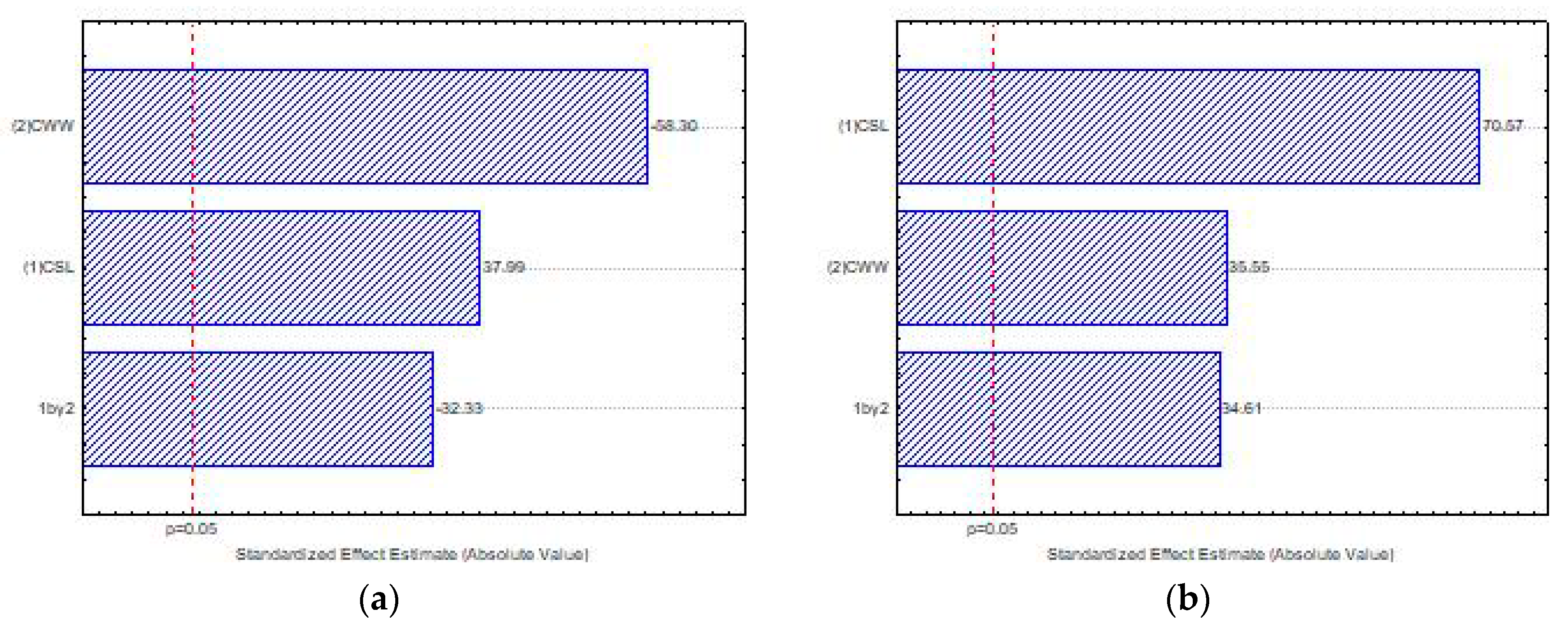
The Pareto graph (
Figure 5a) proves that higher levels of the independent variable CSL (1) significantly influenced the chitosan production. However, lower levels of the CWW (2) significantly influenced the production of chitosan by
M. subtilissimus (UCP 1262). The interaction of the independent variables (1 × 2) significantly influenced, antagonistically, the chitosan production by
M. subtilissimus.
The Pareto (
Figure 5b) shows that higher concentrations of both independent variables—CSL and CWW—exhibited a significant influence on the chitosan production by
L. hyalospora (UCP 1266). Consequently, the interaction of the variables (1 × 2) was significant, suggesting that the synergic effect of these substrates stimulated the biopolymer production.
2.5. Optimization of Chitosan Production by L. hyalospora (UCP 1266)
Due to the results obtained from the previous factorial design,
L. hyalospora (UCP 1266) was selected for a further DCCR 2
2, as it presented the highest chitosan productivity in the culture media with CSL and CWW. For this, the levels of CSL were increased and the higher levels of CWW around the central point of the previous factorial design were maintained. Since the studied concentrations may have allowed a higher activity of chitin deacetylase, the enzyme was responsible for the deacetylation of chitin in cell wall [
55].
The results of the experiments that evaluated the influence of different concentrations of CSL and CWW on the production of biomass and chitosan by
L. hyalospora (UCP 1266) are shown in
Table 6.
From the calculation of the coefficients, we obtained an equation adjusted to the experimental data, with the effects of the two independent variables
x and
y (CSL and CWW) on the production of chitosan Equation (1).
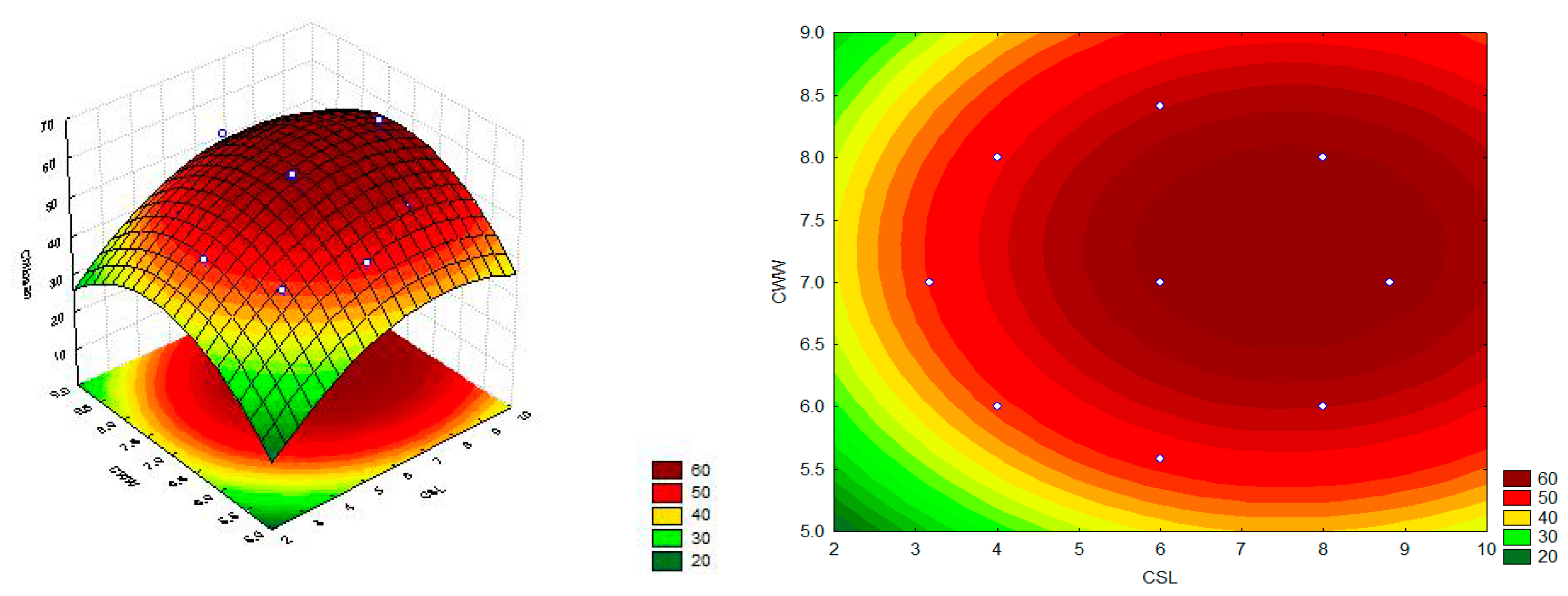
According to the response surface graph (
Figure 7) and Equation (1), the highest chitosan production by
L. hyalospora (UCP 1266) (63.18 mg/g) was achieved in medium containing the highest concentration of CSL (+1.41) and intermediate concentration of CWW (center point—0) (assay 6). The coefficient of determination (
R2) of the obtained model was 0.85, showing good suitability of the experimental data.
The analysis of these results suggests a sustainable culture medium with concentrations of cassava waste water and corn steep liquor as nutritional sources of carbon and nitrogen for optimized production of biomass and chitosan by L. hyalospora UCP 1266. The use of low cost substrates as nutritional sources in culture media decreases the final value of the by-products, mainly in industrial production. Microbial biomass offers economic advantages for industrial scale production on chitosan obtained from crustacean shells, as it does not require high solvent amounts and high temperatures during the extraction process, in addition to obtaining the polysaccharide in a short time.

 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}