Withaferin-A Can Be Used to Modulate the Keratin Network of Intermediate Filaments in Human Epidermal Keratinocytes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Cell Gross Morphology Is Not Affected by Routine Dosages of Withaferin-A up to 1 µM
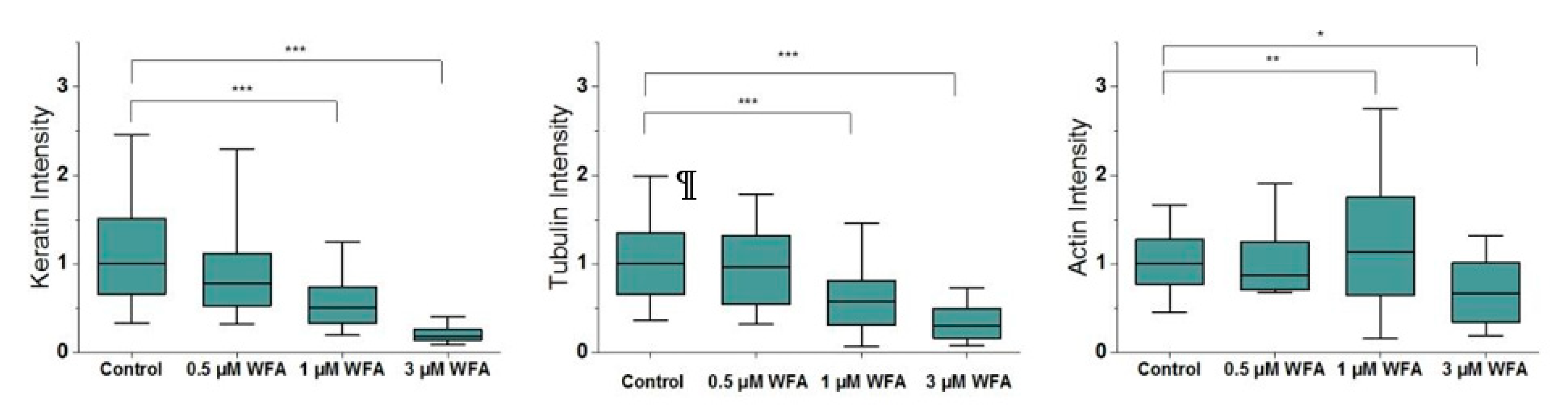
2.2. Withaferin-A Modulates the Assembly of Keratin Filaments in a Dose-Dependant Manner
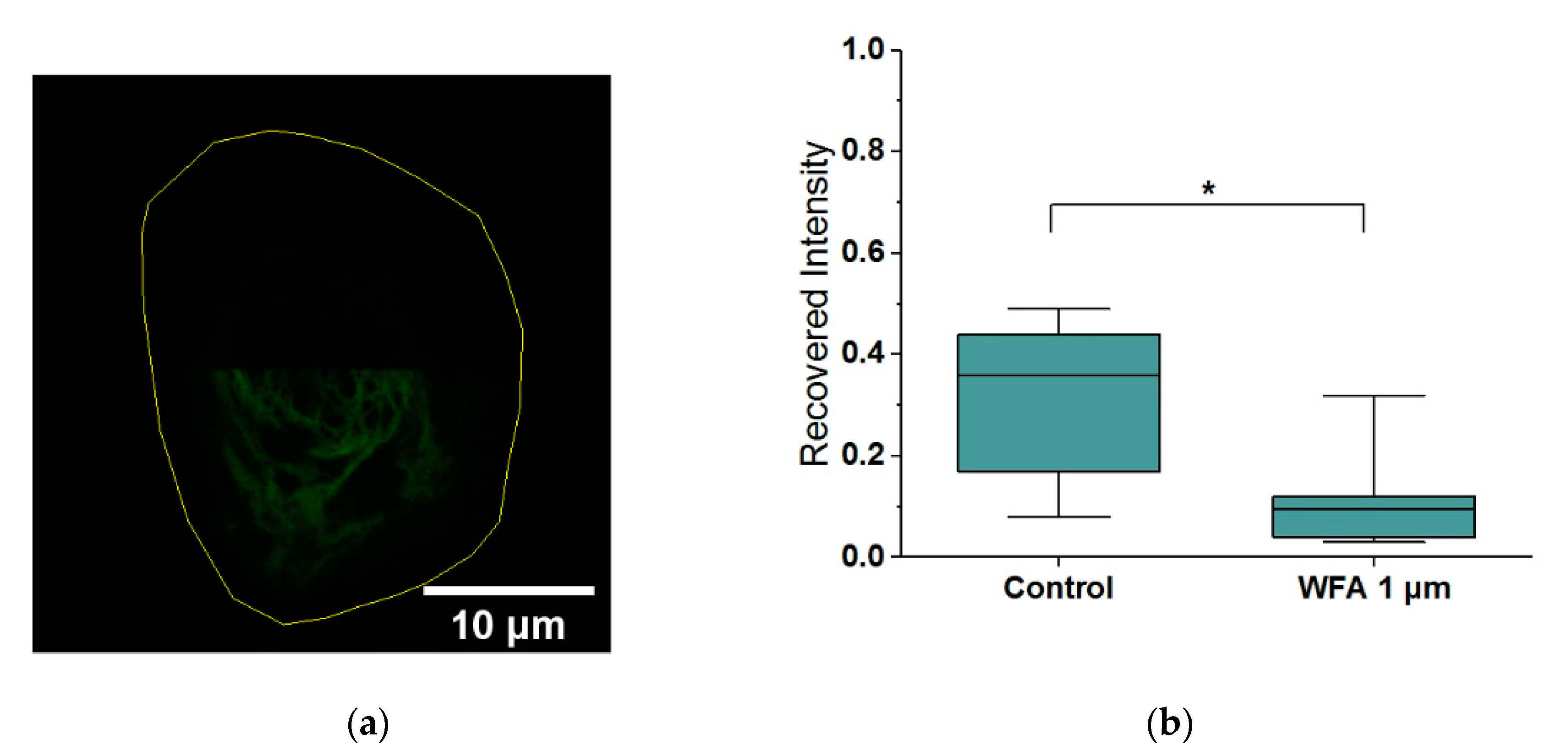
2.3. Withaferin-A Slows Keratin Filament Turnover
2.4. Withaferin-A Increases Keratinocyte Stiffness
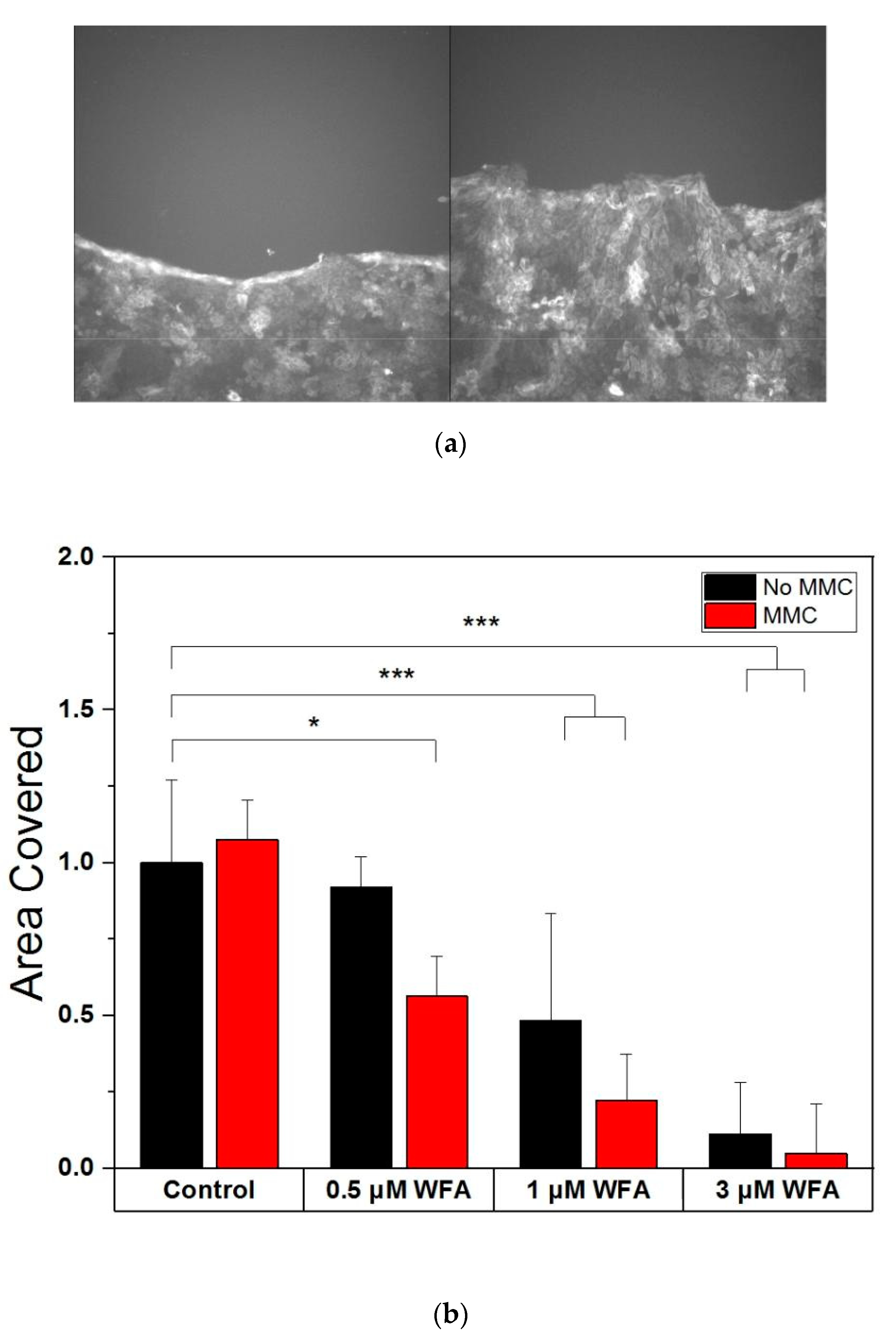
2.5. Withaferin-A Disrupts Keratinocyte Migration and Halts It at High Doses
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Drugs
4.3. Immunofluorescence Staining
4.4. Epiflourescent Imaging and Quantification
4.5. FRAP
4.6. Atomic Force Microscopy
4.7. Cell Migration
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| WFA | Withaferin-A |
| HEK | Human Epidermal Keratinocyte |
| AFM | Atomic Force Microscopy |
| MMC | Mitomycin C |
References
- Patteson, A.E.; Vahabikashi, A.; Pogoda, K.; Adam, S.A.; Goldman, A.; Goldman, R.; Janmey, P. Vimentin protects the structural integrity of the nucleus and suppresses nuclear damage caused by large deformations. bioRxiv 2019, 8, 566174. [Google Scholar]
- Charrier, E.; Janmey, P. Mechanical Properties of Intermediate Filament Proteins. Methods Enzymol. 2016, 568, 35–57. [Google Scholar] [PubMed] [Green Version]
- Magin, T.; Vijayaraj, P.; Leube, R.E. Structural and regulatory functions of keratins. Exp. Cell Res. 2007, 313, 2021–2032. [Google Scholar] [CrossRef] [PubMed]
- Kang, P.; Liao, M.; Wester, M.R.; Leeder, J.S.; Pearce, R.E. “Laminopathies:” a wide spectrum of human diseases. Exp. Cell Res. 2007, 313, 2121–2133. [Google Scholar]
- Homberg, M.; Ramms, L.; Schwarz, N.; Dreissen, G.; Leube, R.E.; Merkel, R.; Hoffmann, B.; Magin, T.M. Distinct Impact of Two Keratin Mutations Causing Epidermolysis Bullosa Simplex on Keratinocyte Adhesion and Stiffness. PLoS ONE 2015, 135, 2437–2445. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Hobbs, R.P.; Coulombe, P.A. The expanding significance of keratin intermediate filaments in normal and diseased epithelia. Curr. Opin. Cell Biol. 2013, 25, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Strnad, P.; Windoffer, R.; Leube, R.E. In vivo detection of cytokeratin filament network breakdown in cells treated with the phosphatase inhibitor okadaic acid. Cell Tissue Res. 2001, 306, 277–293. [Google Scholar] [CrossRef]
- Aggeler, J.; Seely, K. Cytoskeletal Dynamics in Rabbit Synovial Fibroblasts: 1. Effects of Acrylamide on Intermediate Filaments and Microfilaments. Cell Motil Cytoskelet. 1990, 16, 110–120. [Google Scholar] [CrossRef]
- Aspengren, S.; Wielbass, L.; Wallin, M. Effects of acrylamide, latrunculin, and nocodazole on intracellular transport and cytoskeletal organization in melanophores. Cell Motil Cytoskelet. 2006, 63, 423–436. [Google Scholar] [CrossRef]
- Strelkov, S.V.; Herrmann, H.; Geisler, N.; Wedig, T.; Zimbelmann, R.; Aebi, U.; Burkhard, P. Conserved segments 1A and 2B of the intermediate filament dimer: Their atomic structures and role in filament assembly. EMBO J. 2002, 21, 1255–1266. [Google Scholar] [CrossRef] [Green Version]
- Bargagna-Mohan, P.; Hamza, A.; Kim, Y.; Khuan, Y.; Ho, A.; Wendschlag, N.; Liu, J.; Evans, R.M.; Markovitz, D.M.; Zhan, C.; et al. The Tumor Inhibitor and Antiangiogenic Agent Withaferin A Targets the Intermediate Filament Protein Vimentin. Chem. Biol. 2011, 14, 623–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grin, B.; Mahammad, S.; Wedig, T.; Cleland, M.M.; Tsai, L.; Herrmann, H.; Goldman, R.D. Withaferin A Alters Intermediate Filament Organization, Cell Shape and Behavior. PLoS ONE 2012, 7, e39065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eidet, J.R.; Pasovic, L.; Maria, R.; Jackson, C.J.; Utheim, T.P. Objective assessment of changes in nuclear morphology and cell distribution following induction of apoptosis. Diagn Pathol. 2014, 9, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keeling, M.C.; Flores, L.R.; Dodhy, A.H.; Gavara, N. Actomyosin and vimentin cytoskeletal networks regulate nuclear shape, mechanics and chromatin organization. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sliogeryte, K.; Gavara, N. Vimentin Plays a Crucial Role in Fibroblast Ageing by Regulating Biophysical Properties and Cell Migration. Cells 2019, 8, 1164. [Google Scholar] [CrossRef] [Green Version]
- Vargas-Pinto, R.; Gong, H.; Vahabikashi, A.; Johnson, M. The effect of the endothelial cell cortex on atomic force microscopy measurements. Biophys. J. 2013, 105, 300–309. [Google Scholar] [CrossRef] [Green Version]
- Velez-Delvalle, C.; Marsch-Moreno, M.; Castro-Muñozledo, F.; Galván-Mendoza, I.J.; Kuri-Harcuch, W. Epithelial cell migration requires the interaction between the vimentin and keratin intermediate filaments. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Chang, L.; Goldman, R.D. Intermediate filaments mediate cytoskeletal crosstalk. Nat. Rev. Mol. Cell Biol. 2004, 5, 601–613. [Google Scholar] [CrossRef]
- Hohmann, T.; Dehghani, F. The Cytoskeleton—A Complex Interacting Meshwork. Cells 2019, 8, 362. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Naruse, K.; Stamenović, D.; Fredberg, J.J.; Mijailovich, S.M.; Tolić-Nørrelykke, I.M.; Polte, T.; Mannix, R.; Ingber, D.E. Mechanical behavior in living cells consistent with the tensegrity model. Proc. Natl. Acad. Sci. USA 2001, 98, 7765–7770. [Google Scholar] [CrossRef] [Green Version]
- Al-Rekabi, Z.; Haase, K.; Pelling, A.E. Microtubules Mediate Changes in Membrane Cortical Elasticity during Contractile Activation. Exp. Cell. Res. 2014, 322, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Weber, A.; Iturri, J.; Benitez, R.; Zemljic-Jokhadar, S.; Toca-Herrera, J.L. Microtubule disruption changes endothelial cell mechanics and adhesion. Sci. Rep. 2019, 9, 14903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antony, M.L.; Lee, J.; Hahm, E.R.; Kim, S.H.; Marcus, A.I.; Kumari, V.; Ji, X.; Yang, Z.; Vowell, C.L.; Wipf, P.; et al. Growth arrest by the antitumor steroidal lactone withaferin a in human breast cancer cells is associated with down-regulation and covalent binding at cysteine 303 of b-tubulin. J. Biol. Chem. 2014, 289, 1852–1865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Yan, W.; Li, Y.; Niu, L.; Ye, H.; Chen, L. The natural compound withaferin A covalently binds to Cys239 of b-tubulin to promote tubulin degradation. Mol. Pharmacol. 2019, 96, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Coulombe, P. Complementary roles of specific cysteines in keratin 14 toward the assembly, organization, and dynamics of intermediate filaments in skin keratinocytes. PLoS ONE 2012, 290, 22507–22519. [Google Scholar] [CrossRef] [Green Version]
- Windoffer, R.; Beil, M.; Magin, T.M.; Leube, R.E. Cytoskeleton in motion: The dynamics of keratin intermediate filaments in epithelia. J. Cell Biol. 2011, 194, 669–678. [Google Scholar] [CrossRef] [Green Version]
- Ramms, L.; Fabris, G.; Windoffer, R.; Schwarz, N.; Springer, R.; Zhou, C.; Lazar, J.; Stiefel, S.; Hersch, N.; Schnakenberg, U.; et al. Keratins as the main component for the mechanical integrity of keratinocytes. Proc. Natl. Acad. Sci. USA 2013, 110, 18513–18518. [Google Scholar] [CrossRef] [Green Version]
- De Pascalis, C.; Pérez-González, C.; Seetharaman, S.; Boëda, B.; Vianay, B.; Burute, M.; Leduc, C.; Borghi, N.; Trepat, X.; Etienne-Manneville, S. Intermediate filaments control collective migration by restricting tractionforces and sustaining cell-cell contacts. J. Cell Biol. 2018, 217, 3031–3044. [Google Scholar] [CrossRef] [Green Version]
- Kröger, C.; Loschke, F.; Schwarz, N.; Windoffer, R.; Leube, R.E.; Magin, T.M. Keratins control intercellular adhesion involving PKC-a-mediated desmoplakin phosphorylation. J. Cell Biol. 2013, 201, 681–692. [Google Scholar] [CrossRef] [Green Version]
- Rheinwald, J.; Green, H. Serial Cultivation of Strains of Human Epidermal Keratinocytes: The Formation of Keratinizing Colonies from Single Cells. Cell 1975, 6, 331–343. [Google Scholar] [CrossRef]
- Rheinwald, J.; Green, H. Epidermal growth factor and the multiplication of cultured human epidermal keratinocytes. Nature 1977, 265, 421–424. [Google Scholar] [CrossRef] [PubMed]
- Gan, Z.; Ding, L.; Burckhardt, C.J.; Lowery, J.; Zaritsky, A.; Sitterley, K.; Mota, A.; Costigliola, N.; Starker, C.G.; Voytas, D.F.; et al. Vimentin intermediate filaments template microtubule networks to enhance persistence in cell polarity and directed migration. Cell. Syst. 2016, 3, 252–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chadwick, R.S.; Gavara, N. Determination of the elastic moduli of thin samples and adherent cells using conical atomic force microscope tips. Nat. Nanotechnol. 2012, 7, 733–736. [Google Scholar]
- Gavara, N. Combined strategies for optimal detection of the contact point in AFM force-indentation curves obtained on thin samples and adherent cells. Sci. Rep. 2016, 6, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keeling, M.C.; Gavara, N. Withaferin-A Can Be Used to Modulate the Keratin Network of Intermediate Filaments in Human Epidermal Keratinocytes. Int. J. Mol. Sci. 2020, 21, 4450. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124450
Keeling MC, Gavara N. Withaferin-A Can Be Used to Modulate the Keratin Network of Intermediate Filaments in Human Epidermal Keratinocytes. International Journal of Molecular Sciences. 2020; 21(12):4450. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124450
Chicago/Turabian StyleKeeling, Michael C., and Núria Gavara. 2020. "Withaferin-A Can Be Used to Modulate the Keratin Network of Intermediate Filaments in Human Epidermal Keratinocytes" International Journal of Molecular Sciences 21, no. 12: 4450. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124450