Mediterranean Diet: Lipids, Inflammation, and Malaria Infection
by
 , and
, and
Adriana R. Silva
1,2,3,*,† ,
,
Bianca P. T. Moraes
1,2,4,† and
and
Cassiano F. Gonçalves-de-Albuquerque
1,2,4,5,* 1
Laboratório de Imunofarmacologia, Instituto Oswaldo Cruz, Fundação Oswaldo Cruz (FIOCRUZ), Rio de Janeiro 21040-900, Brazil
2
Programa de Neurociências da Universidade Federal Fluminense (UFF), Niterói 24020-141, Brazil
3
Programa de Pós-Graduação em Biologia Celular e Molecular, Instituto Oswaldo Cruz, FIOCRUZ, Rio de Janeiro 21040-900, Brazil
4
Laboratório de Imunofarmacologia, Universidade Federal do Estado do Rio de Janeiro (UNIRIO), Rio de Janeiro 20210-010, Brazil
5
Programa de Pós-Graduação em Biologia Molecular e Celular, UNIRIO, Rio de Janeiro 20210-010, Brazil
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Int. J. Mol. Sci. 2020, 21(12), 4489; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124489
Submission received: 31 December 2019
/
Revised: 5 February 2020
/
Accepted: 15 February 2020
/
Published: 24 June 2020
(This article belongs to the Special Issue Metabolic and Vascular Effects of Mediterranean Diet)
Abstract
:The Mediterranean diet (MedDiet) consists of consumption of vegetables and healthy oils and have beneficial effects on metabolic and inflammatory diseases. Our goal here is to discuss the role of fatty acid content in MedDiet, mostly omega-3, omega-6, and omega-9 on malaria. Malaria affects millions of people around the globe. The parasite Plasmodium causes the disease. The metabolic and inflammatory alterations in the severe forms have damaging consequences to the host. The lipid content in the MedDiet holds anti-inflammatory and pro-resolutive features in the host and have detrimental effects on the Plasmodium. The lipids from the diet impact the balance of pro- and anti-inflammation, thus, lipids intake from the diet is critical to parasite elimination and host tissue damage caused by an immune response. Herein, we go into the cellular and molecular mechanisms and targets of the MedDiet fatty acids in the host and the parasite, reviewing potential benefits of the MedDiet, on inflammation, malaria infection progression, and clinical outcome.
1. Introduction
Over the years, many scientific articles, newspapers, and magazines have promoted the beneficial role of the Mediterranean diet (MedDiet), including association with a decreased incidence of cardiovascular events, diabetes, metabolic syndrome, and breast cancer [1,2,3,4,5].
The MedDiet is a food pattern of countries bordering the Mediterranean Sea, a “Mediterranean lifestyle” with three primary components—olive oil, wine, and bread. The MedDiet dietary pattern includes high consumption of fruits, vegetables, legumes, olive oil, and whole grains, moderate-to-high intake of fish (and seafood), moderate alcohol intake, and low consumption of meat products, dairy and sweets [6,7,8,9] (Table 1). MedDiet has low total fat (<30%), low saturated fat (<10%), and high monounsaturated fatty acid:saturated fatty acid (MUFA:SFA) ratio [8]. Polyunsaturated fatty acid (PUFA) composes the MedDiet, and includes the omega-6 linolenic acid (LA) and marine-derived omega-3 fatty acids, such as α-linolenic acid (ALA), eicosapentaenoic acid (EPA), docosapentaenoic acid (DPA), and docosahexaenoic acid (DHA). Mediterranean seafood species are a dietary source of omega-3 fatty acids, particularly the pelagic fish species [10], but plants and dry fruit might provide a reasonable amount of omega-3 fatty acids. The main source of MUFA is the olive oil [11]. Lipofilic vitamins sources include fish (vitamin D), vegetable and fruits, and minor grade dairy products (vitamins E and A) [12].
Olive oil and fish stand out as health food and the lipid composition of omega-9 and omega-3 fatty acids can account for the wide variety of beneficial effects [1,5,17]. The amount of lipids in the MedDiet is the same as the occidental diet, and the key difference is the type of lipid with a higher amount of unsaturated fatty acid in the MedDiet. Many reports describe the effects of unsaturated fatty acids modulating both inflammation and metabolism, denoting the benefits of the MedDiet [18,19].
Besides the anti-inflammatory effects of fatty acids, the pro-resolutive role of fatty acids are related to the control of the development of inflammatory, infectious, and parasitic diseases. The aim of this review is to summarize the effects of omega-3 eicosapentaenoic acid, docosahexaenoic acid and docosapentaenoic acid, the omega-6 arachidonic acid, and the omega-9 oleic acid, on immune cells (structures in Figure 1), preventing exacerbated inflammation and its deleterious effect, and point out that the control of activation of immune cells and some intracellular signaling proteins and transcription factors are the molecular mechanisms of the beneficial role of these fatty acids. As fatty acid participation in malaria is not broadly reviewed and understood, we both discuss the modulation of parasite fatty acid metabolism; and host diet supplementation with the fatty acid components of MedDiet, relating that to the control of the host response leading to a better outcome. There is a high demand for the discovery of adjuvant therapies or approaches as diet supplementation for parasitic disease management, prevention of severe malaria symptoms, and the improvement of clinical handling.
2. Malaria and Inflammation
2.1. Malaria Cycle, Vaccine, and Treatment
Malaria has gone through tireless efforts to eradicate [20], and besides the advances in diagnosis and treatment, it remains one of the most severe and life-threatening infectious diseases [21]. The parasite plasmodium causes malaria. Both Plasmodium falciparum and vivax are the most prevalent in Africa (99.7%) and in America (74.1%), affecting 219 million people in 2017, where young children in sub-Saharan Africa are the most affected [22].
Approximately 40 species of the mosquito genus Anopheles are considered to be 41 dominant vector species (DVS) of the human malaria parasite [23]. Research mainly focuses on preventing mosquito infection with plasmodium, and preventing transmission of the disease [24]. The long-lasting insecticide-treated bed nets and indoor residual spraying is responsible for decreasing malaria infection in Africa between 2000 and 2015 [25].
During the mosquito–human life cycle, the Plasmodium undergoes many morphological states [26], and during the symptomatic phase, the parasites might replicate exponentially in patients. Female Anopheles mosquito transmits sporozoites, that reach the bloodstream and migrate to hepatocytes. On hepatic cells, the parasite multiplies generating merozoites. Merozoites enter into the bloodstream infecting red blood cells (RBC), starting the intra-erythrocytic cycle. A small part of the schizogony generates the gametocytes required for transmission to the mosquito [21,26,27].
There is no effective malaria vaccine [24]. The only registered malaria vaccine confers only short-term and modest protection [28]. BCG-vaccinated volunteers presented a higher frequency of moderate or severe clinical symptoms than control. It also generates an earlier natural killer cell and monocyte activation, which might have activity against heterologous pathogens [29]. Malaria treatment also remains a challenge. Artemisinin-based combination therapies are the gold standard treatment, when correctly employed [24]. Considerable effort has been made for the development of new malaria treatment as the potential use of “metacaspases”, the unique proteases absent in humans [30]. Tafenoquine was recently registered to treat Plasmodium vivax. In a controlled trial in Ethiopia, Peru, Brazil, Cambodia, Thailand, and the Philippines, a single-dose of tafenoquine reduced the risk of Plasmodium vivax recurrence in patients with regular glucose-6-phosphate dehydrogenase activity [31].
2.2. Malaria Physiopathology
Malaria is classified into uncomplicated and complicated malaria. The first one can be asymptomatic or have mild signs, like fever, with no organ dysfunction [32]. The last one is caused by an inadequate immune response, presenting fever, organ dysfunction, respiratory distress, hepatorenal failure, cerebral malaria, shock, and death [32,33]. In this regard, an immune response is linked to the severity of malaria as well as sepsis. The immune checkpoint blockade is a striking success in specific diseases, suggesting that it would be useful for preventing and treating a range of infectious diseases [34].
A balance between host pro-inflammatory and anti-inflammatory immune responses is a crucial determinant for the pathogenesis of severe malaria. Weaker pro-inflammatory responses would confer protection against inflammation-driven organ damage, but could allow parasite persistence and proliferation, while exuberant pro-inflammatory responses are predicted to control the infection but due to maladapted inflammatory responses could trigger lethal immunopathology, including cerebral malaria [35,36,37]. Controlling deleterious excessive immune response is a strategy to obtain desired therapeutic or preventive strategies in cerebral malaria [38].
The host immune system recognized parasite molecules during infected erythrocyte rupture by the receptor, such as the toll-like receptors (TLRs), triggering the production of pro-inflammatory mediators [35]. Parasite factors stimulate the synthesis and release of cytokines, such as tumor necrosis factor-alpha (TNF-α), interleukin (IL)-6, and IL-1 by macrophages and other cells, resulting in fever, chills, and hyperkinetic hemodynamic changes [39].
The glycosylphosphatidylinositol (GPI) of Plasmodium falciparum is a malaria pathogen-associated molecular pattern (PAMP) and a toxin. Purified GPI induces the expression of pro-inflammatory cytokines TNF-α, IL-1, and IL-12 [40], the inducible nitric oxide synthase (iNOS or NOS2) [41], and adhesion molecules on the endothelium, increasing endothelial-cell binding by parasitized red blood cells (pRBC) [42]. GPI alone is sufficient to induce malarial shock-like syndrome [40]. The proinflammatory cytokine IL-1α has a role in the liver pathology of the experimental malaria model. Plasmodium chabaudi recruits IL-1α-producing neutrophils to the liver. The IL-1α production, independent of the nucleotide-binding oligomerization domain, Leucine-rich Repeat, and Pyrin domain containing 3 (NLRP3) inflammasome activation, increases the production of TNF-α, causing inflammation and hepatocytes apoptosis [43]. A recent report showed the dependence of reactive oxygen species (ROS) produced by xanthine oxidase, to induce the production of inflammatory cytokine in primary human monocyte-derived macrophages in response to plasma patients with cerebral malaria [44]. Infections with intestinal parasites did not alter the patterns of cytokine production in individuals from populations living with endemic malaria, when infected with Plasmodium falciparum and Plasmodium vivax [45]. This finding differed from other studies where children with malaria coinfected with Gram negative bacteria, presented a lower parasite burden, with production of IL-4, IL-10, IL-12, and interferon-gamma (IFN-γ) lowering parasitemia, without worsening anemia [46].
2.3. The Role of Monocytes in Malaria
The hallmark of Plasmodium falciparum infection is exponential parasite growth, with a strong host immune response and microvascular obstruction, due to the adherence of mature parasites to the endothelium, leading to a life-threatening condition [47]. During the blood-stage of malaria infection, the inflammatory response is amplified, with cell activation and pro-inflammatory mediator production, by reciprocal activation loops of white blood cells and endothelial cells [48]. During severe malaria, besides cytokine production, there is an increase in adhesion molecules expression, release of angiopoietin 2 (Ang-2), decrease of nitric oxide (NO), and adhesion of parasitized RBC (pRBC) and leukocytes to brain vasculature, causing microvascular dysfunction that ends up in an activation of microglial cells and astrocytes [49]. Monocytes are vital to control plasmodium burden and to protect the host from the infection because they are associated with pathogenesis and drive the inflammation and sequestration of pRBC in the brain [27]. Monocytes do not phagocytose erythrocytes infected with mature gametocytes [50], but they phagocytize merozoites and asexual pRBC [27]. Opsonized Plasmodium with specific IgG enhances the phagocytic activity and offers protection against malaria infection [51,52,53,54].
Plasmodium-specific IgE and activated monocytes have a role in disease severity [55]. Ortega-Pajares and Rogerson, 2018 reviewed the role of “primed” monocytes at the molecular level in malaria infection in a mechanism called trained immunity in which monocytes have “memory” [27]. In the experimental malaria model in mice, monocytes are the main sequestered leukocyte, inducing and aggravating brain inflammation by recruiting CD4+ and CD8+ T cells [56,57] and also by the chemokine (C-X-C motif) ligand 10 (CXCL10) production [57].
Recent work used a model to predict parasite multiplication rate, and host response in subjects infected with P. falciparum. The authors predicted that parasite-growth inhibition was associated with a lower expression of CXCL10 and IFN-α-associated genes [58]. These results were consistent with findings that CXCL10 deletion and neutralization decreases the parasite load in mice [57]. Hence this model could be a useful tool to study malaria infection course.
In an experimental cerebral malaria study, the induction of IL-33 has led to the polarization to the pro-resolutive macrophages of M2 phenotype and forkhead box P3 (Foxp3) Treg cells [59]. Additionally, IL-33-treated mice presented a reduction in parasitemia at the initial step of infection, but they presented hyperparasitemia and died at the late stages of infection. IL-33 is a new player in experimental cerebral malaria development because the microvascular pathology was dependent on IL-33/ST2 signaling [60].
2.4. The Role of T Cells in Malaria
Blocking immune response prevents T cells from exhaustion, increasing the T cell effector function and allowing interactions between immune cells [61]. The balance of the immune response depends on the optimal T cell activation to the successful resolution of the microbial infections, as well. The negative regulators of T cell activation of the cytotoxic T-lymphocyte-associated protein 4 (CTLA-4) and programmed cell death (PD-1)/programmed death-ligand (PD-L), damps responses during chronic infections [62]. The CTLA-4 and PD-1/PD-L1 inhibitory pathways independently regulate host resistance to plasmodium-induced acute immune pathology [62]. A multimeric form of PDL2 fused with the Fc region of immunoglobulin (PDL2–Fc) attenuates lethal malaria infection [63]. Likewise, a combined blockade of the inhibitory molecules PDL1 and lymphocyte activation gene 3 protein (LAG3), accelerate the clearance of acute non-lethal Plasmodium yoelii malaria [64]. Antibody-mediated triggering of the T cell co-stimulatory molecule OX40 signaling also improved parasite clearance in malarial infections [65]. Additionally, blocking T cell immunoglobulin and mucin domain-containing protein 3 (TIM3) signaling with an antibody, resulted in accelerated parasite clearance and reduced symptoms of cerebral disease in Plasmodium berghei-infected mice [66]. Antibody-mediated blockade of either CTLA4 or PDL1, but not the PD-1/PD-L2, increased the incidence of cerebral malaria in resistance mice in Plasmodium berguei ANKA (PbA) infection [62]. Thus, checkpoint blockade could complement malarial drugs to generate long-term immunity and protect populations in endemic areas.
Diseases caused by intracellular parasites, such as Plasmodium, depend on the IFN-γ-producing Th1 cells to activate dendritic cells and macrophages for capturing, antigen presentation, and for killing pathogens [67]. However, the Th1 cells produce inflammatory cytokines, damaging tissues. IL-10 is a regulatory molecule in malaria, which could prevent excessive inflammation [68]. Studies from mice infected with malaria, IL-10 dampen the inflammatory response, prevent tissue damage, and protect animals [69]. Conversely, the IL-10 anti-inflammatory effect was associated with severe anemia and high parasitemia in experimental malaria [70,71]. Therefore, IL-10 protects host tissues against damaging inflammation, but by doing so, it can foster parasite growth [72].
The splenic plasmacytoid dendritic cells might act as a Trojan horse and carry the live Plasmodium within it. Rapamycin treatment reverses it by inducing an autophagic flux shift of the aggressive Th1 immune pathway, which is reported to incur host damage to a better well-balanced cytokine profile Th2 pathway (increase in IL10/TNF-α, regulatory T cell Treg/Th17) [73]. The IL-10 also has a role in cross-reactive memory response. Mice primed against Plasmodium chabaudi (Pcc) and infected with and Plasmodium Berghei ANKA (PbA) present high levels of IFN-γ and IL-10. Blocking IL-10 receptor increases the activated CD4+ and γδ T cells and the IFN-γ production in response to PbA, with a reduction of parasitemia [74]. Although IFN-γ protects against malaria infection, it has been linked to cerebral malaria and immunopathology [75,76].
Leukotriene B4 (LTB4) was found to be increased in serum mice with cerebral-malaria-induced IFN-γ through dendritic cells [77,78]. Treatment with aspirin caused an overproduction of the omega-6 arachidonic acid (AA) metabolite LTB4 [79]. Hence it might be associated with overproduction of IFN-γ in experimental and human cerebral malaria [80]. Treatment with the AA metabolite leukotriene C4 (LTC4) induced erythrocyte cell death [81]. So LTC4 might induce a clearance of pRBC against malaria infection; contrariwise, the excessive erythrocyte cell death might cause anemia [80].
Naturally acquired immunity to malaria requires both innate and acquired immune responses. Antibodies generated to recognize Plasmodium antigens are lost without the development of immunological B cell memory. Infections during pregnancy might expose the fetus to microbial antigens triggering inflammation and fetal immune response, affecting the B cell. It could prime the infant immune system and its response against a challenge might be augmented [82]. Ly and Hansen reviewed the importance of a proper response and the generation of a long-term B cell memory in malaria infection [83].
2.5. Cerebral Malaria and Inflammation
Sequestration of pRBC within the brain microvasculature is a hallmark of cerebral malaria. Plasmodium falciparum-pRBC isolated in children with complicated malaria, increased binding to brain microvascular endothelial cells, with better adhesion compared to uncomplicated malaria cases. The var gene transcripts predict to bind host endothelial protein C receptor and intercellular adhesion molecule 1 (ICAM-1) [84]. Post mortem analysis showed that children who died of malaria had sequestered pRBC in the retinal microvessels [85]. Binding of pRBC to host endothelial protein C receptor (EPCR) inhibits its interaction with activated protein C, leading to alterations in coagulation, inflammation, and endothelial barrier [86,87,88,89]. Reinforcing the idea that cerebral malaria pathology is based on increased cytoadherence of pRBC in the brain microvasculature [84]. Malaria PbA infection increased lipid peroxidation, and the release of TNF and-α IFN-Υ, and capsazepine (antagonist of transient receptor potential channels, of the vanilloid subtype—TRPV1)-treated group had lower levels of these products [90]. The TRPV1 knockout (KO) exhibited lower concentrations of plasma and brain cytokines, especially TNF-α and IL-6, less cerebral swelling with higher levels of junctional adhesion molecule A, and claudin-5, in their brain vasculature. The TRPV1 KO mice presented less neuronal damage, and they were protected against Plasmodium-induced mortality and morbidity, but the Plasmodium load was not affected [91].
A new approach based on the analysis of the genetic effects of inflammatory response gene variants might offer a new rationale for use, in the prevention and management of severe malaria in infected people who develop severe clinical syndromes. Some inflammatory response genes can be associated with malaria, like TNF-α, NOS2, Interferon-alpha/beta receptor alpha chain gene (IFNAR1), heme oxygenase 1 (HMOX1), TLRs, CD36, and CD40LG. Nevertheless, we must consider that pro-inflammatory genetic variants in early infection stages are useful in resolving malaria, but at later stages these genetic variants relates to enhanced vulnerability to organ damage, resulting from inflammation [35].
3. Inflammation and Fatty Acids
3.1. Omega-9
Non-esterified fatty acids (NEFA) play many essential functions in living organisms, but they can also be toxic to cells. MUFA oleic acid, an omega-9, can be pro-inflammatory or anti-inflammatory, depending on its dose or concentration. The omega-9, oleic acid, is a PUFA with the first double bond on the ninth carbon from the terminal methyl end (Figure 1). Our group has given contributions to this field. Increased plasma concentrations of this omega-9 and other fatty acids increase during diseases. Disease severity in sepsis and leptospirosis can be associated with high plasma levels of NEFA and low blood albumin concentration, denoting NEFA toxicity [92].
Omega-9 induced acute lung injury in experimental models. Increased omega-9 (oleic acid) plasma concentrations cause lung injury by interfering with sodium transport controlled by Na/K-ATPase, leading to acute respiratory distress syndrome (ARDS). Dosage of omega-9 completely inhibited lung Na/K-ATPase activity-induced lung edema and leukocyte accumulation, lipid body formation in leukocytes, and the production of the omega-6 AA metabolites, such as LTB4 and prostaglandin E2 (PGE2) [93]. The omega-9 intratracheal injection caused edema, cell migration and activation, production of lipid mediators, and cytokines on a mechanism dependent on intracellular signaling triggered by the mitogen-activated kinase (MAP) ERK1/2 activation, because of pharmacological inhibition of ERK1/2 phosphorylation blocked neutrophil migration, edema, lipid body formation, and IL-6 production [94].
Plasma NEFA increases in severe systemic inflammatory response syndrome as sepsis, a syndrome characterized by both inflammatory and metabolic alterations, including massive cytokine production, oxidative stress, lipid metabolism disturbances, and organ dysfunction. Although omega-9 (oleic acid) can cause inflammation unexpectedly, 24 h after either intravenous or intragastric administration of the omega-9, the primary NEFA plasma levels were dose-dependently reduced. This effect of omega-9 on the decrease of blood NEFA levels can account for the health benefits of unsaturated fatty-acid-enriched diets, such as the MedDiet, reducing the risk of cardiovascular diseases and cancer [92]. The MedDiet is based on olive oil consumption. Omega-9, oleic acid, is the main component of olive oil. The supplementation for 14 days with this omega-9 ameliorated clinical outcome, increased the survival rate, decreased liver and kidney injury, and diminished NEFA plasma levels in mice subjected to the experimental sepsis model of cecal ligation and puncture (CLP). Omega-9 was effective in diminishing reactive oxygen species (ROS) production and NEFA plasma levels. The beneficial effect of omega-9 supplementation was followed by an increase in translation of the enzyme carnitine palmitoyltransferase IA (CPT1A) and uncoupling protein 2 (UCP2) liver-enhanced expression. These phenomena were compatible with the finding that the decrease in 5′ AMP-activated protein kinase (AMPK) expression found in the liver of septic mice was prevented by omega-9. We suggest that a pretreatment with omega-9 decrease NEFA concentration and increase CPT1A and UCP2 and AMPK levels, and a decrease ROS production (Figure 2). The prevention of metabolic dysfunction by omega-9 has a beneficial role in sepsis, supporting the benefits of diets high in MUFA [95].
Besides the role of omega-9, oleic acid, supplementation on metabolic dysfunction during sepsis, omega-9 treatment increased levels of the anti-inflammatory cytokine IL-10 and decreased levels of the pro-inflammatory cytokines TNF-α and IL-1β, in the peritoneum of septic mice with restored corticosterone levels. The anti-inflammatory role of the omega-9 included a significant inhibitory effect on neutrophil migration, from circulation to the peritoneal cavity and on leukocyte, rolling on the endothelium. Omega-9 also increased bacterial elimination in the peritoneal lavage, suggesting a direct effect on bacteria and an indirect one, on improving the ability of inflammatory leukocytes to fight against infection. Furthermore, omega-9 restored liver and adipose tissue expression of the transcription factor peroxisome-proliferator-activated receptor gamma (PPARγ) expression in septic animals (Figure 2). PPARγ controls inflammation by both negatively modulating the production of pro-inflammatory cytokines and enhancing the synthesis of anti-inflammatory cytokines during sepsis [96]. PPARγ also protected the brain against microvascular dysfunction in sepsis, including diminished leukocyte/endothelial cell interaction and increased functional capillary density in the brain, improving cerebral perfusion [97]. The beneficial anti-inflammatory role of omega-9 in sepsis possibly can occur through a mechanism dependent on PPARγ expression and can join the many described beneficial properties of NEFA-enriched diets in inflammatory diseases [98].
Inflammation associated with metabolic dysregulation is also a common feature in aging. Plasma saturated, PUFA, MUFA, and circulating TNF-α and IL-6 concentrations increased with age, whereas IL-10 and TGF-β1 concentrations decreased with age. Higher levels of the saturated fatty acids C18:0 and C24:0 are linked to lower TGF-β1 concentrations, and higher C16:0 are linked to higher TNF-α concentration. C16:0 also induces the production of cytokines in vitro. Monocytes incubated with C16:0 produced pro-inflammatory cytokines during phorbol myristate acetate (PMA)-induced differentiation. In contrast, the effect of omega-9 was opposite, priming a macrophage phenotype associated with the resolution of inflammation [99].
3.2. Omega-3
Omega-3 are PUFA with the first double bond on the third carbon from the terminal methyl end found (Figure 1) in fish and flaxseed oils. The major fatty acids of the omega-3 family in the fish oil are EPA and DHA. ALA is found in flaxseed oil. Omega-3 protects the heart, liver, and brain [100]. The substrates that generate active metabolites produced by COX, lipoxygenases, and cytochrome P450 monooxygenases, are precursors of the anti-inflammatory signaling molecules eicosanoids. These are players in the control of inflammation [101] and display a wide range of beneficial effects in humans and animals. Omega-3 controls inflammation and tissue homeostasis [102]. Some studies value the link between omega-3 and low-grade inflammation, because omega-3 might displace the eicosanoid metabolism to the formation of mediators with low or no inflammatory activity, decreasing endothelial activation [101]. Diet supplementation with fish oil or omega-3 has a beneficial effect on skin inflammation. In UV-irradiated skin, EPA altered the lipidome, diminishing the production of pro-inflammatory lipids, and DHA inhibited the migration of Langerhans cells [103]. DHA is a substrate for the generation of mediators, as maresins are also involved in tissue regeneration. Macrophages are crucial in producing these mediators and regulating inflammatory response, initiating and promoting its resolution. These mediators promote the uptake and clearance of apoptotic cells, accelerating the resolution of inflammation, culminating in tissue repair and regeneration [104].
Besides EPA and DHA, another important omega-3 PUFAs has recently emerged as specialized pro-resolving mediators like resolvins, maresins, and protectins. DPA, a 22:5 long-chain PUFA [105], can be found with EPA and DHA in fish oils in a percentage of: EPA 10–13%, DPA 2–5%, and DHA 9–11% [106,107]. It also is an elongated metabolite of EPA and is an intermediary metabolite between EPA and DHA [107]. DPA products also have both tissue-protective and anti-inflammatory responses [108]. The protectins derived from DPA reduce human neutrophil–endothelial cell interactions, neutrophil chemotaxis and recruitment, and increased macrophage phagocytosis [108,109]. During sterile inflammation, DPA protectins carry potent leukocyte directed actions, reduce the production of inflammatory cytokines, including monocyte chemoattractant protein-1 (MCP-1/CXCL-2) [110]. Maresin 1 stimulated macrophage phagocytosis and clearance of human apoptotic neutrophils, similar to DHA-derived Maresin 1 [111]. It indicates that biological effects displayed by n-3 fatty acids that has been attributed to EPA and DHA might have be due to DPA as well [112]. DPA even showed a stronger anti-inflammatory effect than EPA and DHA, in an ulcerative colitis model due to its easier incorporation into inflammatory cells [113].
A switch from acute to chronic inflammation can occur when the resolution of inflammation is disrupted, as in chronic heart failure (CHF), where T lymphocytes play a critical role. The omega-3-derived specialized pro-resolving lipid mediator resolvin is low in plasma of patients with CHF, because of low expression in 15-lipoxygenase on their leukocytes. CD8+ and CD4+ T cells were unresponsive in CHF patients. They also show a decrease in the resolving 1 receptor (GPR32), which might be linked to the progression of chronic inflammation, raising the role of omega-3-derived lipids and their signaling pathways as potential targets to CHF treatment [114].
Lipoxygenase omega-3 metabolite resolvin play a role in Trypanossoma cruzi infection, causing a decrease in inflammation and fibrosis in chronic heart disease in mice, consequently increasing the host survival. Resolvin D1 also effective regulates the cell activation in patients with Chagas disease. During Leishmania spp. infections, resolvin D1 control the cutaneous manifestation of diseases [115]. During infection, resolvin enhanced phagocytic clearance of bacteria, downregulated inflammation caused by infection, accelerated resolution of inflammation by promoting neutrophil apoptosis, besides modulating neutrophil migration ability and increased survival. Resolvin was adjuvant to ciprofloxacin and vancomycin in inducing antibacterial effect, indicating a promising new strategy in managing infectious disease [116].
Omega-3 effects are potentiated by non-steroidal anti-inflammatory drugs (NSAID) like aspirin. Their combination promotes the generation of bioactive lipids that better control inflammation. Mice treated with omega-3 and aspirin produce those metabolites. For instance, aspirin stimulates human endothelial cells with upregulated COX-2, to convert C20:5 omega-3 to 18R-hydroxyeicosapentaenoic acid (HEPE) and 15R-HEPE, which are inhibitors of neutrophil transendothelial migration and infiltration. These are the benefits of omega-3 supplementation, which is crucial in modulating inflammation, cancer, and vascular diseases [117].
3.3. Omega-6
The omega-6 AA (Figure 1) is the main fatty acid present in the membrane phospholipids of cells responsible for causing inflammation. AA is a substrate for COX and lipoxygenase, and these lead to potential inflammatory mediators called eicosanoids, like prostaglandins and leukotrienes (Figure 2), respectively [118]. Omega-6 are the precursors of many pro-inflammatory signaling molecules that trigger inflammation [101].
Dietary fatty acids alter immune responses caused by a respiratory infection in pulmonary fibroblasts and bronchial epithelial cells. AA cause IL-6 and CXCL8 release, and it is even higher with AA and the bacterial compound lipoteichoic acid (LTA) together. The production of cytokines by pulmonary fibroblasts depends on prostaglandin, and synergism depend on p38 mitogen-activated protein (MAP) kinase signaling. The high content of dietary omega-6, such as in obese patient diet, could lead to more severe inflammation [119].
Lipoxins (LX) are also produced from AA by lipoxygenase activity in the inflammatory site and present both anti-inflammatory and pro-resolving bioactions [120]. LX reduced neutrophil migration in vitro and increased the neutrophil clearance in vivo [121]. Lipoxin A4 (LXA4) regulates leukocyte trafficking and responses [122], it likewise modulates the activation of vascular, smooth muscle, and epithelial cells [123], and reduces organ fibrosis [124]. LXA4 also attenuates nociception through actions on astrocyte-expressed lipoxin receptors, reducing pain [125]. Despite LX anti-inflammatory properties and pro-resolutive effects [126], they do not impair host defense against infection [127]. LX have beneficial effects in infectious diseases caused by Mycobacterium tuberculosis infections, because they promote necrotic death of infected macrophages and inhibit the initiation of cell-mediated immunity [128,129]. In Trypanosoma cruzi infection, 15-epi-LXA4 reduced parasitemia and increased survival [130]. Conversely, in Toxoplasma gondii infection, LX reduced mortality but increased parasite load [131]. In sepsis, LX increased survival by reducing systemic inflammation [132]. The binding of LXA4 to its receptor ALX can modulate the expression of adhesion molecules through inhibition of the NFκB pathway in endothelial cells (Figure 2) [133,134,135].
3.4. Omega-3 Versus Omega-6
Common sense believes dietary intake of the omega-6 AA or its precursor linoleic acid (LA) potentiates inflammation because the synthesis of the pro-inflammatory AA metabolites prevail over the production of anti-inflammatory AA metabolites or resolutive mediators (Figure 2). Some references, however, have shown there is no correlation. Nevertheless, the competition of both substrates and high omega-6 diet content would favor the production of pro-inflammatory and an inhibition of the anti-inflammatory and pro-resolutive effect of omega-3 [118].
An alternative use of NSAID inhibits the formation of AA metabolites because of their role on COX inhibition, and therefore, its deleterious side effects would be a diet rich in EPA/DHA. Supplementation with fish oil inhibits inflammatory cytokines, such as TNF-α and IL-1β and pro-inflammatory and pro-aggregatory eicosanoids such as thromboxane-2 and PGE2. Eating omega-6 seed oils have the potential to increase those phenomena and omega-3 suppress inflammation. A balance, favoring the enrichment of a diet with omega-3 might profit numerous chronic inflammatory diseases, such as rheumatoid arthritis, atherosclerosis, dyslipidemia, diabetes, obesity and heart failure [136], sepsis, and malaria.
Supplementation of an appropriate omega-6/omega-3 ratio modulated T helper (Th) cells and Treg cells that are involved in the exacerbated immune response related to the progression of sepsis. In an experimental model of sepsis fed with a diet rich in fish oil with an omega-6/omega-3 PUFA ratio of 2:1, decreased inflammatory mediator production in the plasma and peritoneal lavage (Figure 2), compared to omega-6/omega-3 PUFA ratio of 7:1 rich diet. This was followed by a decrease in Th1, Th2, and Th17 cells (Figure 2), and a reduction of neutrophil infiltration. A diet rich in omega-6/omega-3 PUFA ratio of 2:1, upregulated the transcription factor PPARγ. The activation of PPARγ might be the molecular mechanism involved in which a fish oil-enriched diet maintains balanced Th polarization, mitigates inflammation, and alleviates exacerbated inflammation in sepsis [96].
Besides the omega-3 effect on T cells, dietary omega-3 long-chain PUFA control inflammation in mice by modulating dendritic cell function during experimental autoimmune uveitis (EAU). An omega-6 long-chain PUFA diet has not shown the same effect. Transfer of dendritic cells from mice fed omega-3 long-chain PUFA diet to mice with EAU decreased the T cell proliferation and the production of IFN-γ and IL-17 by T cells, mitigating the disease development. Splenocytes were isolated from the mice and cocultured with CD4+ T cells isolated from mice with EAU [137]. Omega-3 long-chain PUFA seem to have a molecular target in T cells, the T-bet and RORγt, Th1- and Th17-related transcription factors, respectively. T-bet and RORγt transcription was diminished in the retina and lymph node cells of mice fed with rich omega-3 long-chain diet, but not with an omega-6 long-chain one [138].
4. Fatty Acids and Malaria
4.1. Fatty Acid Metabolism of the Parasite as a Target of Antimalarial Compounds
The malaria parasite goes through several morphological stages during its life cycle [26]. The intense proliferation demands nutrients, including fatty acids. Parasites obtain fatty acids by two pathways, scavenging from the vertebrate host and mosquito vector or by producing them through the fatty acid synthase type II (FAS-II) [139,140]. This enzyme was expressed in the mosquito midgut and liver-stage parasites [140], the blood-stage did not require the FAS-II [141,142,143]. P. falciparum FAS-II knockout parasites failed to complete sporogony, revealing that the FAS-ll pathway is essential for the human infection phase [140]. Mostly due to the differences in FAS-I and the Plasmodium FAS-II, the last emerges as a promising drug target, especially for the clinically silent liver state infection. In this regard, synthesized 2-, 5-, 6-, and 9-hexadecanoic acids (HDAs) were developed. Plasmodium falciparum blood stages 5-HAD, displayed the highest antiplasmodial activity, and 2-HDA was the most significant inhibitor of the FAS II enzymes [33].
A series of 2-alkynoic fatty acids (2-AFA) with chain lengths between 14 and 18 carbon atoms were tested against Plasmodium falciparum and Plasmodium berghei. Treatment with 2-ODA and 2-TDA was effective in treating infection and killing parasites in hepatic cell cultures. Among them, 2-ODA was the most effective inhibitor, with the three critical enzymes in the FAS II elongation pathway being even more potent than primaquine [144].
Four fatty acids from lichens, the evernic, vulpic, psoromic, and (+)-usnic acids, act indirectly via binding to allosteric sites on the protein surface of the FAS-II enzymes (PfFabI, PfFabG, and PfFabZ elongation enzymes), thus, having an antiplasmodial effect [145].
Fatty acid biosynthesis is an interactive process, beginning with the condensation of acetyl-CoA, with a growing fatty acid chain. Plasmodium falciparum enoyl acyl carrier protein (ACP) reductase (PfENR) is responsible for the final step in each fatty acid synthesis cycle [146]. Fabl is an enzyme that catalyzes the last step of biosynthesis in the FAS-II pathway and is also known as an enoyl-ACP reductase. In the malaria Plasmodium berghei mouse model, the use of triclosan, a potent inhibitor of FabI, actively suppressed parasitemia [147]. Additionally, triclosan shows activity against Plasmodium asexual blood-stage parasites and shows efficacy in vivo, whereas FabI-deficient Plasmodium berghei sporozoites failed to complete liver-stage development in vitro [141].
Turkish marine sponge Agelas oroides, produce a vast diversity of pyrrole and imidazole alkaloids [148], yielding a variety of pure metabolites that have inhibitory effects against Plasmodium falciparum enoyl-ACP reductase (PfFabI) enzyme [149].
A rhodanine (2-thioxothiazolidin-4-one) class of inhibitory compounds of enoyl acyl carrier protein (ACP) reductase inhibited Plasmodium growth in RBC cultures [150], even in chloroquine-sensitive Plasmodium falciparum [151].
The continuous growth of parasite resistance requires novel modes of action. Fatty acids have also been used on insecticide new formulation strategies. An insecticide comprising straight-chain octanoic, nonanoic, and decanoic saturated fatty acids (C8, C9, and C10) called C8910, displayed insecticidal effects on different strains of Anopheles funestus [152]. Supplemented birds with olive oil and vitamin E increased the transmission to the vector, indicating that vertebrate host oxidative status influences parasite transmission to the vector [153].
4.2. Omega-3, Omega-6, and Malaria
Since 1957, Godfrey has shown that diet alterations might influence the development of malaria disease in mice. The Plasmodium berghei infection was utterly suppressed in the mice, when given a high cod liver oil diet and also in those given vitamin “E” [154]. Feeding mice 4 weeks before inoculation with Plasmodium yoelii with the fish oils (menhaden, anchovy, or salmon) in a vitamin E-deficient diet, actively protected against malaria in mice, reducing parasitemia and improving survival [155,156]. Mice supplemented with SFA, EPA, and DHA fortified with alfa-tocopherol and that infected with Plasmodium yoelii, reduced parasitemia and improved survival rates [156,157].
Mice fed with fish oil diets rich in EPA and DHA with vitamin E did not survive, but when fed without vitamin E they had considerably improved survival, and the effect lasted for months [158]. Yet, C-18 fatty acids (omega-9 oleic acid, elaidic acid, and linoleic acid) displayed a considerably inhibitory activity, both in infected cells and in free parasites Plasmodium falciparum, Plasmodium vinckei petteri, and Plasmodium yoelii. Vitamin E addition to a culture of trophozoite-infected cells did not reverse the inhibitory effects of fatty acids [159].
EPA, DHA, and AA enhanced neutrophil antimalarial activity with increased superoxide radical generation in response to Plasmodium falciparum [160]. Fatty acids stimulate the generation of free radicals by leukocyte to kill Plasmodium falciparum. Patients with active Plasmodium falciparum-positive malaria with lower concentrations of lipid peroxides and nitric oxide allow an infection to persist, whereas elevated levels of EPA might indicate an attempt to overcome the infection [161]. The animal malaria model treated with DHA showed a decrease in the duration of malaria symptoms [162]. Both, treatments with linolenic acid (omega-3) and linoleic acid (omega-6), showed a potent inhibition of the parasitemia in Plasmodium berghei infection [163].
Diet supplementation with omega-3 makes the host response more prompt to a resolutive response, accelerating the resolution of inflammation. Nevertheless, the administration of the omega-6 metabolite LX also negatively modulates inflammation by shifting to a more anti-inflammatory phenotype. The lipoxin epimer 15-epi-LXA4 and LXA4 itself, as discussed before, have numerous anti-inflammatory properties [164,165,166]. The administration of 15-epi-LXA4 reduced brain inflammation by inhibiting IL-12 production, and accumulation of IFN-producing cells in the brains of infected mice decreases lymphocyte infiltration and enhances survival [36]. Similarly, treatment with LXA4 reduced vascular occlusion and reduced Plasmodium berghei-infected erythrocytes adhered to brain vasculature, besides an improved functional capillary density [49]. Moreover, LXA4-induced heme oxygenase 1 (HO-1), an enzyme associated with tissue protection [167], in endothelial dysfunction in vivo [168] and in vitro [169], which are essential to the prevention of severe malaria development. HO-1 is related to the inhibition of adhesion molecules preserving endothelial functions and BBB integrity [170,171]. Additionally, HO-1 in brain tissue was associated with mouse survival and decreased cerebral edema [172].
Lung injury in severe malaria is a concern. Neutrophil accumulation and alveolar-capillary membrane alterations cause lung damage, leading to physiological dysfunction. In Plasmodium berghei infection, pretreatment with LXA4, prevented lung edema and preserved lung morphology and function, compared to nontreated Plasmodium berghei-infected mice. During Plasmodium berghei infection, pretreatment or posttreatment with LXA4 alleviated lung neutrophil accumulation, reduced alveolar collapse, thickening and edema, and decreased the production of CXCL1 in plasma [173]. LXA4 also affected neutrophil migration in vitro, stimulated by plasma from mice infected with Plasmodium berghei, by disrupting neutrophil cytoskeleton that inhibits F-actin polarization. In conclusion, LXA4 exerted therapeutic effects in malaria-induced ALI by inhibiting lung dysfunction [173].
A healthy diet stimulating the increase of the daily consumption of omega-3 has been broadly discussed as a link between increased consumption of omega-3 to a less inflammatory phenotype. It is valid for malaria as well, but interestingly and unexpectedly, the administration of a high-fat saturated diet to mice for just 4 days dramatically mitigates Plasmodium berguei ANKA infection. Oxidative stress seems to be involved in biochemical mechanisms. Products of fatty acid β-oxidation impact Plasmodium survival in hepatocytes, even transient dietary changes impact the Plasmodium berguei ANKA infection and the development of the disease [174].
5. Concluding Remarks
The MedDiet has beneficial effects on cardiovascular diseases, diabetes, metabolic syndrome, certain types of cancer, and inflammatory and infectious diseases [1,5,98,175,176]. MedDiet consists of a large number of bioactive ingredients, including high content of olive oil and fish rich in omega-3 and omega-9. Both fatty acids have anti-inflammatory [2] and pro-resolutive effects [104], acting pharmacologically and on biochemical targets [177]. The omega-3 and omega-9 show metabolic and immune regulatory effects, modulating intracellular pathways and transcription factor activation. The lipid mediators generated through omega-3 or omega-6 metabolism have a critical role in controlling inflammatory, infectious, and parasitic diseases [98], including malaria. Besides omega-3 and omega-9, saturated fatty acid and omega-6 have emerged with potential beneficial effects on experimental malaria by controlling excessive inflammation, impacting malaria infection development, and clinical outcome. Dosing the amount and type of fatty acid in MedDiet is critical to seal the inflammatory, infectious, and parasitic disease progression. Further controlled studies will unearth the potential benefits of increased consumption of specific fatty acid components of the MedDiet.
Author Contributions
Conceptualization, writing—original draft preparation and editing, funding acquisition—all authors (B.P.T.M., A.R.S. and C.F.G.-d.-A.); review—A.R.S. and C.F.G.-d.-A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Instituto Oswaldo Cruz, Fundação Oswaldo Cruz (FIOCRUZ), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) Grant 001, Programa de Neurociências da Universidade Federal Fluminense (UFF), Universidade Federal do Estado do Rio de Janeiro (Unirio), Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Tuttolomondo, A.; Simonetta, I.; Daidone, M.; Mogavero, A.; Ortello, A.; Pinto, A. Metabolic and Vascular Effect of the Mediterranean Diet. Int. J. Mol. Sci. 2019, 20, 4716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, J.; Fialho, M.; Santos, R.; Peixoto-Placido, C.; Madeira, T.; Sousa-Santos, N.; Virgolino, A.; Santos, O.; Vaz Carneiro, A. Is olive oil good for you? A systematic review and meta-analysis on anti-inflammatory benefits from regular dietary intake. Nutrition 2020, 69, 110559. [Google Scholar] [CrossRef] [PubMed]
- Newman, T.M.; Vitolins, M.Z.; Cook, K.L. From the Table to the Tumor: The Role of Mediterranean and Western Dietary Patterns in Shifting Microbial-Mediated Signaling to Impact Breast Cancer Risk. Nutrients 2019, 11, 2565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muralidharan, J.; Galie, S.; Hernandez-Alonso, P.; Bullo, M.; Salas-Salvado, J. Plant-Based Fat, Dietary Patterns Rich in Vegetable Fat and Gut Microbiota Modulation. Front. Nutr. 2019, 6, 157. [Google Scholar] [CrossRef] [Green Version]
- Kargin, D.; Tomaino, L.; Serra-Majem, L. Experimental Outcomes of the Mediterranean Diet: Lessons Learned from the Predimed Randomized Controlled Trial. Nutrients 2019, 11, 2991. [Google Scholar] [CrossRef] [Green Version]
- Ferro-Luzzi, A.; Branca, F. Mediterranean diet, Italian-style: Prototype of a healthy diet. Am. J. Clin. Nutr. 1995, 61 (Suppl. 6), 1338S–1345S. [Google Scholar] [CrossRef]
- Mastorakou, D.; Rabaeus, M.; Salen, P.; Pounis, G.; de Lorgeril, M. Mediterranean Diet: A Health-Protective Dietary Pattern for Modern Times. In Analysis in Nutrition Research, 1st ed.; Pounis, G., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 233–258. [Google Scholar]
- Simopoulos, A.P. Fish and Fish Oil in the Mediterranean Diets. In Fish and Fish Oil in Health and Disease Prevention; Susan, K., Raatz, D.M.B., Eds.; Academic Press—Elsevier: Amsterdam, The Netherlands, 2016; p. 380. [Google Scholar]
- Willett, W.C.; Sacks, F.; Trichopoulou, A.; Drescher, G.; Ferro-Luzzi, A.; Helsing, E.; Trichopoulos, D. Mediterranean diet pyramid: A cultural model for healthy eating. Am. J. Clin. Nutr. 1995, 61 (Suppl. 6), 1402S–1406S. [Google Scholar] [CrossRef]
- Galli, C.; Marangoni, F. N-3 fatty acids in the Mediterranean diet. Prostaglandins Leukot Essent Fatty Acids 2006, 75, 129–133. [Google Scholar] [CrossRef]
- Mazzocchi, A.; Leone, L.; Agostoni, C.; Pali-Scholl, I. The Secrets of the Mediterranean Diet. Does [Only] Olive Oil Matter? Nutrients 2019, 11, 2941. [Google Scholar] [CrossRef] [Green Version]
- Castiglione, D.; Platania, A.; Conti, A.; Falla, M.; D’Urso, M.; Marranzano, M. Dietary Micronutrient and Mineral Intake in the Mediterranean Healthy Eating, Ageing, and Lifestyle (MEAL) Study. Antioxidants 2018, 7, 79. [Google Scholar] [CrossRef] [Green Version]
- Bedard, A.; Riverin, M.; Dodin, S.; Corneau, L.; Lemieux, S. Sex differences in the impact of the Mediterranean diet on cardiovascular risk profile. Br. J. Nutr. 2012, 108, 1428–1434. [Google Scholar] [CrossRef] [PubMed]
- Stamm, O.; Latscha, U.; Janecek, P.; Campana, A. Development of a special electrode for continuous subcutaneous pH measurement in the infant scalp. Am. J. Obstet. Gynecol. 1976, 124, 193–195. [Google Scholar] [CrossRef]
- Bedard, A.; Dodin, S.; Corneau, L.; Lemieux, S. The impact of abdominal obesity status on cardiovascular response to the mediterranean diet. J. Obes. 2012, 2012, 969124. [Google Scholar] [CrossRef] [PubMed]
- Mayr, H.L.; Thomas, C.J.; Tierney, A.C.; Kucianski, T.; George, E.S.; Ruiz-Canela, M.; Hebert, J.R.; Shivappa, N.; Itsiopoulos, C. Randomization to 6-month Mediterranean diet compared with a low-fat diet leads to improvement in Dietary Inflammatory Index scores in patients with coronary heart disease: The AUSMED Heart Trial. Nutr. Res. 2018, 55, 94–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alkhatib, A.; Tsang, C.; Tuomilehto, J. Olive Oil Nutraceuticals in the Prevention and Management of Diabetes: From Molecules to Lifestyle. Int. J. Mol. Sci. 2018, 19, 2024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yubero-Serrano, E.M.; Lopez-Moreno, J.; Gomez-Delgado, F.; Lopez-Miranda, J. Extra virgin olive oil: More than a healthy fat. Eur. J. Clin. Nutr. 2019, 72 (Suppl. 1), 8–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casas, R.; Estruch, R.; Sacanella, E. Influence of Bioactive Nutrients on the Atherosclerotic Process: A Review. Nutrients 2018, 10, 1630. [Google Scholar] [CrossRef] [Green Version]
- World-Health-Organization. World Malaria Report. 2015. Available online: https://apps.who.int/iris/bitstream/handle/10665/200018/9789241565158_eng.pdf;jsessionid=0F94417AFADC9C6AEA7DD8DDC7B6624E?sequence=1 (accessed on 30 December 2019).
- Cowman, A.F.; Healer, J.; Marapana, D.; Marsh, K. Malaria: Biology and Disease. Cell 2016, 167, 610–624. [Google Scholar] [CrossRef] [Green Version]
- World-Health-Organization. World Malaria Report. 2018. Available online: http://apps.who.int/iris/bitstream/handle/10665/275867/9789241565653-eng.pdf?ua=1 (accessed on 30 December 2019).
- Sinka, M.E.; Bangs, M.J.; Manguin, S.; Coetzee, M.; Mbogo, C.M.; Hemingway, J.; Patil, A.P.; Temperley, W.H.; Gething, P.W.; Kabaria, C.W.; et al. The dominant Anopheles vectors of human malaria in Africa, Europe and the Middle East: Occurrence data, distribution maps and bionomic precis. Parasit Vectors 2010, 3, 117. [Google Scholar] [CrossRef] [Green Version]
- Ashley, E.A.; Pyae Phyo, A.; Woodrow, C.J. Malaria. Lancet 2018, 391, 1608–1621. [Google Scholar] [CrossRef]
- Bhatt, S.; Weiss, D.J.; Cameron, E.; Bisanzio, D.; Mappin, B.; Dalrymple, U.; Battle, K.; Moyes, C.L.; Henry, A.; Eckhoff, P.A.; et al. The effect of malaria control on Plasmodium falciparum in Africa between 2000 and 2015. Nature 2015, 526, 207–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Josling, G.A.; Llinas, M. Sexual development in Plasmodium parasites: Knowing when it’s time to commit. Nat. Rev. Microbiol. 2015, 13, 573–587. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Pajares, A.; Rogerson, S.J. The Rough Guide to Monocytes in Malaria Infection. Front. Immunol. 2018, 9, 2888. [Google Scholar] [CrossRef] [PubMed]
- Olotu, A.; Fegan, G.; Wambua, J.; Nyangweso, G.; Leach, A.; Lievens, M.; Kaslow, D.C.; Njuguna, P.; Marsh, K.; Bejon, P. Seven-Year Efficacy of RTS,S/AS01 Malaria Vaccine among Young African Children. N. Engl. J. Med. 2016, 374, 2519–2529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walk, J.; de Bree, L.C.J.; Graumans, W.; Stoter, R.; van Gemert, G.J.; van de Vegte-Bolmer, M.; Teelen, K.; Hermsen, C.C.; Arts, R.J.W.; Behet, M.C.; et al. Outcomes of controlled human malaria infection after BCG vaccination. Nat. Commun. 2019, 10, 874. [Google Scholar] [CrossRef] [Green Version]
- Pandey, K.C.; Kumari, V.; Dixit, R.; Tiwari, R.; Katyal, A. Metacaspases: Potential Drug Target against Protozoan Parasites. Front. Pharmacol. 2019, 10, 790. [Google Scholar] [CrossRef]
- Lacerda, M.V.G.; Llanos-Cuentas, A.; Krudsood, S.; Lon, C.; Saunders, D.L.; Mohammed, R.; Yilma, D.; Batista Pereira, D.; Espino, F.E.J.; Mia, R.Z.; et al. Single-Dose Tafenoquine to Prevent Relapse of Plasmodium vivax Malaria. N. Engl. J. Med. 2019, 380, 215–228. [Google Scholar] [CrossRef]
- Milner, D.A., Jr. Malaria Pathogenesis. Cold Spring Harb. Perspect. Med. 2018, 8. [Google Scholar] [CrossRef] [Green Version]
- Anstey, N.M.; Douglas, N.M.; Poespoprodjo, J.R.; Price, R.N. Plasmodium vivax: Clinical spectrum, risk factors and pathogenesis. Adv. Parasitol. 2012, 80, 151–201. [Google Scholar]
- Goncalves-de-Albuquerque, C.F.; Rohwedder, I.; Silva, A.R.; Ferreira, A.S.; Kurz, A.R.M.; Cougoule, C.; Klapproth, S.; Eggersmann, T.; Silva, J.D.; de Oliveira, G.P.; et al. The Yin and Yang of Tyrosine Kinase Inhibition during Experimental Polymicrobial Sepsis. Front. Immunol. 2018, 9, 901. [Google Scholar] [CrossRef] [Green Version]
- Penha-Goncalves, C. Genetics of Malaria Inflammatory Responses: A Pathogenesis Perspective. Front. Immunol. 2019, 10, 1771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shryock, N.; McBerry, C.; Salazar Gonzalez, R.M.; Janes, S.; Costa, F.T.; Aliberti, J. Lipoxin A(4) and 15-epi-lipoxin A(4) protect against experimental cerebral malaria by inhibiting IL-12/IFN-gamma in the brain. PLoS ONE 2013, 8, e61882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunnington, A.J.; Walther, M.; Riley, E.M. Piecing together the puzzle of severe malaria. Sci. Transl. Med. 2013, 5, 211ps18. [Google Scholar] [CrossRef] [PubMed]
- Shastri, A.; Bonifati, D.M.; Kishore, U. Innate immunity and neuroinflammation. Mediat. Inflamm. 2013, 2013, 342931. [Google Scholar] [CrossRef] [PubMed]
- Sypniewska, P.; Duda, J.F.; Locatelli, I.; Althaus, C.R.; Althaus, F.; Genton, B. Clinical and laboratory predictors of death in African children with features of severe malaria: A systematic review and meta-analysis. BMC Med. 2017, 15, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schofield, L.; Hackett, F. Signal transduction in host cells by a glycosylphosphatidylinositol toxin of malaria parasites. J. Exp. Med. 1993, 177, 145–153. [Google Scholar] [CrossRef] [Green Version]
- Tachado, S.D.; Gerold, P.; McConville, M.J.; Baldwin, T.; Quilici, D.; Schwarz, R.T.; Schofield, L. Glycosylphosphatidylinositol toxin of Plasmodium induces nitric oxide synthase expression in macrophages and vascular endothelial cells by a protein tyrosine kinase-dependent and protein kinase C-dependent signaling pathway. J. Immunol. 1996, 156, 1897–1907. [Google Scholar]
- Schofield, L.; Novakovic, S.; Gerold, P.; Schwarz, R.T.; McConville, M.J.; Tachado, S.D. Glycosylphosphatidylinositol toxin of Plasmodium up-regulates intercellular adhesion molecule-1, vascular cell adhesion molecule-1, and E-selectin expression in vascular endothelial cells and increases leukocyte and parasite cytoadherence via tyrosine kinase-dependent signal transduction. J. Immunol. 1996, 156, 1886–1896. [Google Scholar]
- De Menezes, M.N.; Salles, E.M.; Vieira, F.; Amaral, E.P.; Zuzarte-Luis, V.; Cassado, A.; Epiphanio, S.; Alvarez, J.M.; Alves-Filho, J.C.; Mota, M.M.; et al. IL-1alpha promotes liver inflammation and necrosis during blood-stage Plasmodium chabaudi malaria. Sci. Rep. 2019, 9, 7575. [Google Scholar] [CrossRef]
- Ty, M.C.; Zuniga, M.; Gotz, A.; Kayal, S.; Sahu, P.K.; Mohanty, A.; Mohanty, S.; Wassmer, S.C.; Rodriguez, A. Malaria inflammation by xanthine oxidase-produced reactive oxygen species. EMBO Mol. Med. 2019, 11, e9903. [Google Scholar] [CrossRef]
- Sanchez-Arcila, J.C.; Perce-da-Silva, D.S.; Vasconcelos, M.P.; Rodrigues-da-Silva, R.N.; Pereira, V.A.; Aprigio, C.J.; Lima, C.A.; Fonseca e Fonseca, B.P.; Banic, D.M.; Lima-Junior, J.C.; et al. Intestinal parasites coinfection does not alter plasma cytokines profile elicited in acute malaria in subjects from endemic area of Brazil. Mediat. Inflamm. 2014, 2014, 857245. [Google Scholar] [CrossRef] [PubMed]
- Davenport, G.C.; Hittner, J.B.; Otieno, V.; Karim, Z.; Mukundan, H.; Fenimore, P.W.; Hengartner, N.W.; McMahon, B.H.; Kempaiah, P.; Ong’echa, J.M.; et al. Reduced Parasite Burden in Children with Falciparum Malaria and Bacteremia Coinfections: Role of Mediators of Inflammation. Mediat. Inflamm. 2016, 2016, 4286576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wassmer, S.C.; Grau, G.E. Severe malaria: what’s new on the pathogenesis front? Int. J. Parasitol. 2017, 47, 145–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deroost, K.; Pham, T.T.; Opdenakker, G.; Van den Steen, P.E. The immunological balance between host and parasite in malaria. FEMS Microbiol. Rev. 2016, 40, 208–257. [Google Scholar] [CrossRef]
- Souza, M.C.; Padua, T.A.; Henriques, M.G. Endothelial-Leukocyte Interaction in Severe Malaria: Beyond the Brain. Mediat. Inflamm. 2015, 2015, 168937. [Google Scholar] [CrossRef] [Green Version]
- Bansal, G.P.; Weinstein, C.S.; Kumar, N. Insight into phagocytosis of mature sexual (gametocyte) stages of Plasmodium falciparum using a human monocyte cell line. Acta Trop. 2016, 157, 96–101. [Google Scholar] [CrossRef] [Green Version]
- Chan, J.A.; Stanisic, D.I.; Duffy, M.F.; Robinson, L.J.; Lin, E.; Kazura, J.W.; King, C.L.; Siba, P.M.; Fowkes, F.J.; Mueller, I.; et al. Patterns of protective associations differ for antibodies to P. falciparum-infected erythrocytes and merozoites in immunity against malaria in children. Eur. J. Immunol. 2017, 47, 2124–2136. [Google Scholar] [CrossRef] [Green Version]
- Lambert, L.H.; Bullock, J.L.; Cook, S.T.; Miura, K.; Garboczi, D.N.; Diakite, M.; Fairhurst, R.M.; Singh, K.; Long, C.A. Antigen reversal identifies targets of opsonizing IgGs against pregnancy-associated malaria. Infect. Immun. 2014, 82, 4842–4853. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Feng, G.; Beeson, J.; Hogarth, P.M.; Rogerson, S.J.; Yan, Y.; Jaworowski, A. CD14(hi)CD16+ monocytes phagocytose antibody-opsonised Plasmodium falciparum infected erythrocytes more efficiently than other monocyte subsets, and require CD16 and complement to do so. BMC Med. 2015, 13, 154. [Google Scholar] [CrossRef] [Green Version]
- Hommel, M.; Chan, J.A.; Umbers, A.J.; Langer, C.; Rogerson, S.J.; Smith, J.D.; Beeson, J.G. Evaluating antibody functional activity and strain-specificity of vaccine candidates for malaria in pregnancy using in vitro phagocytosis assays. Parasit Vectors 2018, 11, 69. [Google Scholar] [CrossRef] [Green Version]
- Kumsiri, R.; Troye-Blomberg, M.; Pattanapanyasat, K.; Krudsood, S.; Maneerat, Y. IgE low affinity receptor (CD23) expression, Plasmodium falciparum specific IgE and tumor necrosis factor-alpha production in Thai uncomplicated and severe falciparum malaria patients. Acta Trop. 2016, 154, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Pai, S.; Qin, J.; Cavanagh, L.; Mitchell, A.; El-Assaad, F.; Jain, R.; Combes, V.; Hunt, N.H.; Grau, G.E.; Weninger, W. Real-time imaging reveals the dynamics of leukocyte behaviour during experimental cerebral malaria pathogenesis. PLoS Pathog. 2014, 10, e1004236. [Google Scholar] [CrossRef] [PubMed]
- Ioannidis, L.J.; Nie, C.Q.; Ly, A.; Ryg-Cornejo, V.; Chiu, C.Y.; Hansen, D.S. Monocyte- and Neutrophil-Derived CXCL10 Impairs Efficient Control of Blood-Stage Malaria Infection and Promotes Severe Disease. J. Immunol. 2016, 196, 1227–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgiadou, A.; Lee, H.J.; Walther, M.; van Beek, A.E.; Fitriani, F.; Wouters, D.; Kuijpers, T.W.; Nwakanma, D.; D’Alessandro, U.; Riley, E.M.; et al. Modelling pathogen load dynamics to elucidate mechanistic determinants of host-Plasmodium falciparum interactions. Nat. Microbiol. 2019, 4, 1592–1602. [Google Scholar] [CrossRef] [PubMed]
- Besnard, A.G.; Guabiraba, R.; Niedbala, W.; Palomo, J.; Reverchon, F.; Shaw, T.N.; Couper, K.N.; Ryffel, B.; Liew, F.Y. IL-33-mediated protection against experimental cerebral malaria is linked to induction of type 2 innate lymphoid cells, M2 macrophages and regulatory T cells. PLoS Pathog. 2015, 11, e1004607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palomo, J.; Reverchon, F.; Piotet, J.; Besnard, A.G.; Couturier-Maillard, A.; Maillet, I.; Tefit, M.; Erard, F.; Mazier, D.; Ryffel, B.; et al. Critical role of IL-33 receptor ST2 in experimental cerebral malaria development. Eur. J. Immunol. 2015, 45, 1354–1365. [Google Scholar] [CrossRef] [Green Version]
- Wykes, M.N.; Lewin, S.R. Immune checkpoint blockade in infectious diseases. Nat. Rev. Immunol. 2018, 18, 91–104. [Google Scholar] [CrossRef]
- Hafalla, J.C.; Claser, C.; Couper, K.N.; Grau, G.E.; Renia, L.; de Souza, J.B.; Riley, E.M. The CTLA-4 and PD-1/PD-L1 inhibitory pathways independently regulate host resistance to Plasmodium-induced acute immune pathology. PLoS Pathog. 2012, 8, e1002504. [Google Scholar] [CrossRef]
- Karunarathne, D.S.; Horne-Debets, J.M.; Huang, J.X.; Faleiro, R.; Leow, C.Y.; Amante, F.; Watkins, T.S.; Miles, J.J.; Dwyer, P.J.; Stacey, K.J.; et al. Programmed Death-1 Ligand 2-Mediated Regulation of the PD-L1 to PD-1 Axis Is Essential for Establishing CD4(+) T Cell Immunity. Immunity 2016, 45, 333–345. [Google Scholar] [CrossRef] [Green Version]
- Butler, N.S.; Moebius, J.; Pewe, L.L.; Traore, B.; Doumbo, O.K.; Tygrett, L.T.; Waldschmidt, T.J.; Crompton, P.D.; Harty, J.T. Therapeutic blockade of PD-L1 and LAG-3 rapidly clears established blood-stage Plasmodium infection. Nat. Immunol. 2011, 13, 188–195. [Google Scholar] [CrossRef] [Green Version]
- Zander, R.A.; Obeng-Adjei, N.; Guthmiller, J.J.; Kulu, D.I.; Li, J.; Ongoiba, A.; Traore, B.; Crompton, P.D.; Butler, N.S. PD-1 Co-inhibitory and OX40 Co-stimulatory Crosstalk Regulates Helper T Cell Differentiation and Anti-Plasmodium Humoral Immunity. Cell Host Microbe 2015, 17, 628–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, N.; Zou, Y.; Piao, X.; Liu, S.; Wang, L.; Li, S.; Chen, Q. T-Cell Immunoglobulin- and Mucin-Domain-Containing Molecule 3 Signaling Blockade Improves Cell-Mediated Immunity Against Malaria. J. Infect. Dis. 2016, 214, 1547–1556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tubo, N.J.; Jenkins, M.K. CD4+ T Cells: Guardians of the phagosome. Clin. Microbiol. Rev. 2014, 27, 200–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couper, K.N.; Blount, D.G.; Riley, E.M. IL-10: The master regulator of immunity to infection. J. Immunol. 2008, 180, 5771–5777. [Google Scholar] [CrossRef]
- Freitas do Rosario, A.P.; Langhorne, J. T cell-derived IL-10 and its impact on the regulation of host responses during malaria. Int. J. Parasitol. 2012, 42, 549–555. [Google Scholar] [CrossRef]
- Kobayashi, F.; Ishida, H.; Matsui, T.; Tsuji, M. Effects of in vivo administration of anti-IL-10 or anti-IFN-gamma monoclonal antibody on the host defense mechanism against Plasmodium yoelii yoelii infection. J. Vet. Med. Sci. 2000, 62, 583–587. [Google Scholar] [CrossRef] [Green Version]
- Omer, F.M.; de Souza, J.B.; Riley, E.M. Differential induction of TGF-beta regulates proinflammatory cytokine production and determines the outcome of lethal and nonlethal Plasmodium yoelii infections. J. Immunol. 2003, 171, 5430–5436. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Ng, S.; Engwerda, C. The Role of IL-10 in Malaria: A Double Edged Sword. Front. Immunol. 2019, 10, 229. [Google Scholar] [CrossRef] [Green Version]
- Sengupta, A.; Keswani, T.; Sarkar, S.; Ghosh, S.; Mukherjee, S.; Bhattacharyya, A. Autophagic induction modulates splenic plasmacytoid dendritic cell mediated immune response in cerebral malarial infection model. Microbes Infect. 2019, 21, 475–484. [Google Scholar] [CrossRef]
- Nakamae, S.; Kimura, D.; Miyakoda, M.; Sukhbaatar, O.; Inoue, S.I.; Yui, K. Role of IL-10 in inhibiting protective immune responses against infection with heterologous Plasmodium parasites. Parasitol. Int. 2019, 70, 5–15. [Google Scholar] [CrossRef] [Green Version]
- Grau, G.E.; Heremans, H.; Piguet, P.F.; Pointaire, P.; Lambert, P.H.; Billiau, A.; Vassalli, P. Monoclonal antibody against interferon gamma can prevent experimental cerebral malaria and its associated overproduction of tumor necrosis factor. Proc. Natl. Acad. Sci. USA 1989, 86, 5572–5574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanez, D.M.; Manning, D.D.; Cooley, A.J.; Weidanz, W.P.; van der Heyde, H.C. Participation of lymphocyte subpopulations in the pathogenesis of experimental murine cerebral malaria. J. Immunol. 1996, 157, 1620–1624. [Google Scholar] [PubMed]
- Toda, A.; Terawaki, K.; Yamazaki, S.; Saeki, K.; Shimizu, T.; Yokomizo, T. Attenuated Th1 induction by dendritic cells from mice deficient in the leukotriene B4 receptor 1. Biochimie 2010, 92, 682–691. [Google Scholar] [CrossRef] [PubMed]
- Arcoleo, F.; Milano, S.; D’Agostino, P.; Cillari, E. Effect of exogenous leukotriene B4 (LTB4) on BALB/c mice splenocyte production of Th1 and Th2 lymphokines. Int. J. Immunopharmacol. 1995, 17, 457–463. [Google Scholar] [CrossRef]
- Babu, K.S.; Salvi, S.S. Aspirin and asthma. Chest 2000, 118, 1470–1476. [Google Scholar] [CrossRef]
- Rogerio, A.P.; Anibal, F.F. Role of leukotrienes on protozoan and helminth infections. Mediat. Inflamm. 2012, 2012, 595694. [Google Scholar] [CrossRef] [Green Version]
- Foller, M.; Mahmud, H.; Gu, S.; Wang, K.; Floride, E.; Kucherenko, Y.; Luik, S.; Laufer, S.; Lang, F. Participation of leukotriene C(4) in the regulation of suicidal erythrocyte death. J. Physiol. Pharmacol. 2009, 60, 135–143. [Google Scholar]
- Yeo, K.T.; Embury, P.; Anderson, T.; Mungai, P.; Malhotra, I.; King, C.; Kazura, J.; Dent, A. HIV, Cytomegalovirus, and Malaria Infections during Pregnancy Lead to Inflammation and Shifts in Memory B Cell Subsets in Kenyan Neonates. J. Immunol. 2019, 202, 1465–1478. [Google Scholar] [CrossRef]
- Ly, A.; Hansen, D.S. Development of B Cell Memory in Malaria. Front. Immunol. 2019, 10, 559. [Google Scholar] [CrossRef]
- Storm, J.; Jespersen, J.S.; Seydel, K.B.; Szestak, T.; Mbewe, M.; Chisala, N.V.; Phula, P.; Wang, C.W.; Taylor, T.E.; Moxon, C.A.; et al. Cerebral malaria is associated with differential cytoadherence to brain endothelial cells. EMBO Mol. Med. 2019, 11, e9164. [Google Scholar] [CrossRef]
- Barrera, V.; Hiscott, P.S.; Craig, A.G.; White, V.A.; Milner, D.A.; Beare, N.A.; MacCormick, I.J.; Kamiza, S.; Taylor, T.E.; Molyneux, M.E.; et al. Severity of retinopathy parallels the degree of parasite sequestration in the eyes and brains of malawian children with fatal cerebral malaria. J. Infect. Dis. 2015, 211, 1977–1986. [Google Scholar] [CrossRef]
- Gillrie, M.R.; Avril, M.; Brazier, A.J.; Davis, S.P.; Stins, M.F.; Smith, J.D.; Ho, M. Diverse functional outcomes of Plasmodium falciparum ligation of EPCR: Potential implications for malarial pathogenesis. Cell. Microbiol. 2015, 17, 1883–1899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampath, S.; Brazier, A.J.; Avril, M.; Bernabeu, M.; Vigdorovich, V.; Mascarenhas, A.; Gomes, E.; Sather, D.N.; Esmon, C.T.; Smith, J.D. Plasmodium falciparum adhesion domains linked to severe malaria differ in blockade of endothelial protein C receptor. Cell. Microbiol. 2015, 17, 1868–1882. [Google Scholar] [CrossRef] [Green Version]
- Bernabeu, M.; Danziger, S.A.; Avril, M.; Vaz, M.; Babar, P.H.; Brazier, A.J.; Herricks, T.; Maki, J.N.; Pereira, L.; Mascarenhas, A.; et al. Severe adult malaria is associated with specific PfEMP1 adhesion types and high parasite biomass. Proc. Natl. Acad. Sci. USA 2016, 113, E3270–E3279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kessler, A.; Dankwa, S.; Bernabeu, M.; Harawa, V.; Danziger, S.A.; Duffy, F.; Kampondeni, S.D.; Potchen, M.J.; Dambrauskas, N.; Vigdorovich, V.; et al. Linking EPCR-Binding PfEMP1 to Brain Swelling in Pediatric Cerebral Malaria. Cell Host Microbe 2017, 22, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, E.S.; Brito, C.X.; Teixeira, S.A.; Barboza, R.; dos Reis, A.S.; Azevedo-Santos, A.P.; Muscara, M.; Costa, S.K.; Marinho, C.R.; Brain, S.D.; et al. TRPV1 antagonism by capsazepine modulates innate immune response in mice infected with Plasmodium berghei ANKA. Mediat. Inflamm. 2014, 2014, 506450. [Google Scholar] [CrossRef] [Green Version]
- Pereira, D.M.S.; Teixeira, S.A.; Murillo, O.; Peixoto, E.P.M.; Araujo, M.C.; Sousa, N.C.F.; Monteiro-Neto, V.; Calixto, J.B.; Cunha, T.M.; Marinho, C.R.F.; et al. TRPV1 Contributes to Cerebral Malaria Severity and Mortality by Regulating Brain Inflammation. Oxidative Med. Cell. Longev. 2019, 2019, 9451671. [Google Scholar] [CrossRef] [Green Version]
- Goncalves de Albuquerque, C.F.; Burth, P.; Younes Ibrahim, M.; Garcia, D.G.; Bozza, P.T.; Castro Faria Neto, H.C.; Castro Faria, M.V. Reduced plasma nonesterified fatty acid levels and the advent of an acute lung injury in mice after intravenous or enteral oleic acid administration. Mediat. Inflamm. 2012, 2012, 601032. [Google Scholar] [CrossRef] [Green Version]
- Goncalves-de-Albuquerque, C.F.; Burth, P.; Silva, A.R.; de Moraes, I.M.; de Jesus Oliveira, F.M.; Santelli, R.E.; Freire, A.S.; Bozza, P.T.; Younes-Ibrahim, M.; de Castro-Faria-Neto, H.C.; et al. Oleic acid inhibits lung Na/K-ATPase in mice and induces injury with lipid body formation in leukocytes and eicosanoid production. J. Inflamm. 2013, 10, 34. [Google Scholar] [CrossRef] [Green Version]
- Goncalves-de-Albuquerque, C.F.; Silva, A.R.; Burth, P.; de Moraes, I.M.; Oliveira, F.M.; Younes-Ibrahim, M.; dos Santos Mda, C.; D’Avila, H.; Bozza, P.T.; Faria Neto, H.C.; et al. Oleic acid induces lung injury in mice through activation of the ERK pathway. Mediat. Inflamm. 2012, 2012, 956509. [Google Scholar] [CrossRef] [Green Version]
- Goncalves-de-Albuquerque, C.F.; Medeiros-de-Moraes, I.M.; Oliveira, F.M.; Burth, P.; Bozza, P.T.; Castro Faria, M.V.; Silva, A.R.; Castro-Faria-Neto, H.C. Omega-9 Oleic Acid Induces Fatty Acid Oxidation and Decreases Organ Dysfunction and Mortality in Experimental Sepsis. PLoS ONE 2016, 11, e0153607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araujo, C.V.; Campbell, C.; Goncalves-de-Albuquerque, C.F.; Molinaro, R.; Cody, M.J.; Yost, C.C.; Bozza, P.T.; Zimmerman, G.A.; Weyrich, A.S.; Castro-Faria-Neto, H.C.; et al. A PPARgamma agonist enhances bacterial clearance through neutrophil extracellular trap formation and improves survival in sepsis. Shock 2016, 45, 393–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araujo, C.V.; Estato, V.; Tibirica, E.; Bozza, P.T.; Castro-Faria-Neto, H.C.; Silva, A.R. PPAR gamma activation protects the brain against microvascular dysfunction in sepsis. Microvasc. Res. 2012, 84, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Medeiros-de-Moraes, I.M.; Goncalves-de-Albuquerque, C.F.; Kurz, A.R.M.; Oliveira, F.M.J.; de Abreu, V.H.P.; Torres, R.C.; Carvalho, V.F.; Estato, V.; Bozza, P.T.; Sperandio, M.; et al. Omega-9 Oleic Acid, the Main Compound of Olive Oil, Mitigates Inflammation during Experimental Sepsis. Oxidative Med. Cell. Longev. 2018, 2018, 6053492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pararasa, C.; Ikwuobe, J.; Shigdar, S.; Boukouvalas, A.; Nabney, I.T.; Brown, J.E.; Devitt, A.; Bailey, C.J.; Bennett, S.J.; Griffiths, H.R. Age-associated changes in long-chain fatty acid profile during healthy aging promote pro-inflammatory monocyte polarization via PPARgamma. Aging Cell 2016, 15, 128–139. [Google Scholar] [CrossRef]
- Ye, J.; Ghosh, S. Omega-3 PUFA vs. NSAIDs for Preventing Cardiac Inflammation. Front. Cardiovasc. Med. 2018, 5, 146. [Google Scholar] [CrossRef]
- Dessi, M.; Noce, A.; Bertucci, P.; Manca di Villahermosa, S.; Zenobi, R.; Castagnola, V.; Addessi, E.; Di Daniele, N. Atherosclerosis, dyslipidemia, and inflammation: The significant role of polyunsaturated Fatty acids. ISRN Inflamm. 2013, 2013, 191823. [Google Scholar] [CrossRef] [Green Version]
- Ishihara, T.; Yoshida, M.; Arita, M. Omega-3 fatty acid-derived mediators that control inflammation and tissue homeostasis. Int. Immunol. 2019, 9, 559–567. [Google Scholar] [CrossRef] [Green Version]
- Kendall, A.C.; Pilkington, S.M.; Murphy, S.A.; Del Carratore, F.; Sunarwidhi, A.L.; Kiezel-Tsugunova, M.; Urquhart, P.; Watson, R.E.B.; Breitling, R.; Rhodes, L.E.; et al. Dynamics of the human skin mediator lipidome in response to dietary omega-3 fatty acid supplementation. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2019, 33, 13014–13027. [Google Scholar]
- Dalli, J.; Serhan, C.N. Pro-Resolving Mediators in Regulating and Conferring Macrophage Function. Front. Immunol. 2017, 8, 1400. [Google Scholar] [CrossRef] [Green Version]
- Weylandt, K.H. Docosapentaenoic acid derived metabolites and mediators—The new world of lipid mediator medicine in a nutshell. Eur. J. Pharmacol. 2016, 785, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Guo, X.F.; Sinclair, A.J. Short update on docosapentaenoic acid: A bioactive long-chain n-3 fatty acid. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 88–91. [Google Scholar] [CrossRef] [PubMed]
- Morin, C.; Hiram, R.; Rousseau, E.; Blier, P.U.; Fortin, S. Docosapentaenoic acid monoacylglyceride reduces inflammation and vascular remodeling in experimental pulmonary hypertension. Am. J. Physiol. Heart Circ. Physiol. 2014, 307, H574–H586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalli, J.; Colas, R.A.; Serhan, C.N. Novel n-3 immunoresolvents: Structures and actions. Sci. Rep. 2013, 3, 1940. [Google Scholar] [CrossRef]
- Aursnes, M.; Tungen, J.E.; Vik, A.; Colas, R.; Cheng, C.Y.; Dalli, J.; Serhan, C.N.; Hansen, T.V. Total synthesis of the lipid mediator PD1n-3 DPA: Configurational assignments and anti-inflammatory and pro-resolving actions. J. Nat. Prod. 2014, 77, 910–916. [Google Scholar] [CrossRef]
- Pistorius, K.; Souza, P.R.; De Matteis, R.; Austin-Williams, S.; Primdahl, K.G.; Vik, A.; Mazzacuva, F.; Colas, R.A.; Marques, R.M.; Hansen, T.V.; et al. PDn-3 DPA Pathway Regulates Human Monocyte Differentiation and Macrophage Function. Cell Chem. Biol. 2018, 25, 749–760. [Google Scholar] [CrossRef]
- Tungen, J.E.; Aursnes, M.; Dalli, J.; Arnardottir, H.; Serhan, C.N.; Hansen, T.V. Total synthesis of the anti-inflammatory and pro-resolving lipid mediator MaR1n-3 DPA utilizing an sp(3)-sp(3) Negishi cross-coupling reaction. Chemistry 2014, 20, 14575–14578. [Google Scholar] [CrossRef] [Green Version]
- Skulas-Ray, A.C.; Flock, M.R.; Richter, C.K.; Harris, W.S.; West, S.G.; Kris-Etherton, P.M. Red Blood Cell Docosapentaenoic Acid (DPA n-3) is Inversely Associated with Triglycerides and C-reactive Protein (CRP) in Healthy Adults and Dose-Dependently Increases Following n-3 Fatty Acid Supplementation. Nutrients 2015, 7, 6390–6404. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Z.; Dai, Z.; Cao, Y.; Shen, Q.; Zhang, Y. Docosapentaenoic acid (DPA, 22:5n-3) ameliorates inflammation in an ulcerative colitis model. Food Funct. 2019, 10, 4199–4209. [Google Scholar] [CrossRef]
- Chiurchiu, V.; Leuti, A.; Saracini, S.; Fontana, D.; Finamore, P.; Giua, R.; Padovini, L.; Incalzi, R.A.; Maccarrone, M. Resolution of inflammation is altered in chronic heart failure and entails a dysfunctional responsiveness of T lymphocytes. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2019, 33, 909–916. [Google Scholar] [CrossRef]
- Lopez-Munoz, R.A.; Molina-Berrios, A.; Campos-Estrada, C.; Abarca-Sanhueza, P.; Urrutia-Llancaqueo, L.; Pena-Espinoza, M.; Maya, J.D. Inflammatory and Pro-resolving Lipids in Trypanosomatid Infections: A Key to Understanding Parasite Control. Front. Microbiol. 2018, 9, 1961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, C.D.; Schwarze, J. The role of pro-resolution lipid mediators in infectious disease. Immunology 2014, 141, 166–173. [Google Scholar] [CrossRef] [Green Version]
- Serhan, C.N.; Clish, C.B.; Brannon, J.; Colgan, S.P.; Chiang, N.; Gronert, K. Novel functional sets of lipid-derived mediators with antiinflammatory actions generated from omega-3 fatty acids via cyclooxygenase 2-nonsteroidal antiinflammatory drugs and transcellular processing. J. Exp. Med. 2000, 192, 1197–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Innes, J.K.; Calder, P.C. Omega-6 fatty acids and inflammation. Prostaglandins Leukot Essent Fatty Acids 2018, 132, 41–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutting, S.; Zakarya, R.; Bozier, J.; Xenaki, D.; Horvat, J.C.; Wood, L.G.; Hansbro, P.M.; Oliver, B.G. Dietary Fatty Acids Amplify Inflammatory Responses to Infection through p38 MAPK Signaling. Am. J. Respir. Cell Mol. Biol. 2019, 60, 554–568. [Google Scholar] [CrossRef] [PubMed]
- Ryan, A.; Godson, C. Lipoxins: Regulators of resolution. Curr. Opin. Pharmacol. 2010, 10, 166–172. [Google Scholar] [CrossRef]
- Wu, B.; Walker, J.; Spur, B.; Rodriguez, A.; Yin, K. Effects of Lipoxin A4 on antimicrobial actions of neutrophils in sepsis. Prostaglandins Leukot Essent Fatty Acids 2015, 94, 55–64. [Google Scholar] [CrossRef]
- Romano, M.; Cianci, E.; Simiele, F.; Recchiuti, A. Lipoxins and aspirin-triggered lipoxins in resolution of inflammation. Eur. J. Pharmacol. 2015, 760, 49–63. [Google Scholar] [CrossRef]
- Brennan, E.P.; Mohan, M.; McClelland, A.; de Gaetano, M.; Tikellis, C.; Marai, M.; Crean, D.; Dai, A.; Beuscart, O.; Derouiche, S.; et al. Lipoxins Protect Against Inflammation in Diabetes-Associated Atherosclerosis. Diabetes 2018, 67, 2657–2667. [Google Scholar] [CrossRef] [Green Version]
- Brennan, E.P.; Cacace, A.; Godson, C. Specialized pro-resolving mediators in renal fibrosis. Mol. Asp. Med. 2017, 58, 102–113. [Google Scholar] [CrossRef]
- Svensson, C.I.; Zattoni, M.; Serhan, C.N. Lipoxins and aspirin-triggered lipoxin inhibit inflammatory pain processing. J. Exp. Med. 2007, 204, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Chiang, N.; Van Dyke, T.E. Resolving inflammation: Dual anti-inflammatory and pro-resolution lipid mediators. Nat. Rev. Immunol. 2008, 8, 349–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basil, M.C.; Levy, B.D. Specialized pro-resolving mediators: Endogenous regulators of infection and inflammation. Nat. Rev. Immunol. 2016, 16, 51–67. [Google Scholar] [CrossRef] [PubMed]
- Tobin, D.M.; Roca, F.J.; Oh, S.F.; McFarland, R.; Vickery, T.W.; Ray, J.P.; Ko, D.C.; Zou, Y.; Bang, N.D.; Chau, T.T.; et al. Host genotype-specific therapies can optimize the inflammatory response to mycobacterial infections. Cell 2012, 148, 434–446. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Divangahi, M.; Gan, H.; Shin, D.S.; Hong, S.; Lee, D.M.; Serhan, C.N.; Behar, S.M.; Remold, H.G. Lipid mediators in innate immunity against tuberculosis: Opposing roles of PGE2 and LXA4 in the induction of macrophage death. J. Exp. Med. 2008, 205, 2791–2801. [Google Scholar] [CrossRef] [Green Version]
- Molina-Berrios, A.; Campos-Estrada, C.; Henriquez, N.; Faundez, M.; Torres, G.; Castillo, C.; Escanilla, S.; Kemmerling, U.; Morello, A.; Lopez-Munoz, R.A.; et al. Protective role of acetylsalicylic acid in experimental Trypanosoma cruzi infection: Evidence of a 15-epi-lipoxin A(4)-mediated effect. PLoS Negl. Trop. Dis. 2013, 7, e2173. [Google Scholar] [CrossRef]
- Aliberti, J.; Serhan, C.; Sher, A. Parasite-induced lipoxin A4 is an endogenous regulator of IL-12 production and immunopathology in Toxoplasma gondii infection. J. Exp. Med. 2002, 196, 1253–1262. [Google Scholar] [CrossRef]
- Walker, J.; Dichter, E.; Lacorte, G.; Kerner, D.; Spur, B.; Rodriguez, A.; Yin, K. Lipoxin a4 increases survival by decreasing systemic inflammation and bacterial load in sepsis. Shock 2011, 36, 410–416. [Google Scholar] [CrossRef]
- Merched, A.J.; Serhan, C.N.; Chan, L. Nutrigenetic disruption of inflammation-resolution homeostasis and atherogenesis. J. Nutr. Nutr. 2011, 4, 12–24. [Google Scholar] [CrossRef] [Green Version]
- Campos-Estrada, C.; Liempi, A.; Gonzalez-Herrera, F.; Lapier, M.; Kemmerling, U.; Pesce, B.; Ferreira, J.; Lopez-Munoz, R.; Maya, J.D. Simvastatin and Benznidazole-Mediated Prevention of Trypanosoma cruzi-Induced Endothelial Activation: Role of 15-epi-lipoxin A4 in the Action of Simvastatin. PLoS Negl. Trop. Dis. 2015, 9, e0003770. [Google Scholar] [CrossRef]
- Gonzalez-Herrera, F.; Cramer, A.; Pimentel, P.; Castillo, C.; Liempi, A.; Kemmerling, U.; Machado, F.S.; Maya, J.D. Simvastatin Attenuates Endothelial Activation through 15-Epi-Lipoxin A4 Production in Murine Chronic Chagas Cardiomyopathy. Antimicrob. Agents Chemother. 2017, 61, e02137-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DiNicolantonio, J.J.; O’Keefe, J.H. Importance of maintaining a low omega-6/omega-3 ratio for reducing inflammation. Open Heart 2018, 5, e000946. [Google Scholar] [CrossRef] [PubMed]
- Uchi, S.H.; Yanai, R.; Kobayashi, M.; Hatano, M.; Kobayashi, Y.; Yamashiro, C.; Nagai, T.; Tokuda, K.; Connor, K.M.; Sonoda, K.H.; et al. Dendritic cells mediate the anti-inflammatory action of omega-3 long-chain polyunsaturated fatty acids in experimental autoimmune uveitis. PLoS ONE 2019, 14, e0219405. [Google Scholar] [CrossRef]
- Shoda, H.; Yanai, R.; Yoshimura, T.; Nagai, T.; Kimura, K.; Sobrin, L.; Connor, K.M.; Sakoda, Y.; Tamada, K.; Ikeda, T.; et al. Dietary Omega-3 Fatty Acids Suppress Experimental Autoimmune Uveitis in Association with Inhibition of Th1 and Th17 Cell Function. PLoS ONE 2015, 10, e0138241. [Google Scholar] [CrossRef]
- Tarun, A.S.; Vaughan, A.M.; Kappe, S.H. Redefining the role of de novo fatty acid synthesis in Plasmodium parasites. Trends Parasitol. 2009, 25, 545–550. [Google Scholar] [CrossRef] [PubMed]
- van Schaijk, B.C.; Kumar, T.R.; Vos, M.W.; Richman, A.; van Gemert, G.J.; Li, T.; Eappen, A.G.; Williamson, K.C.; Morahan, B.J.; Fishbaugher, M.; et al. Type II fatty acid biosynthesis is essential for Plasmodium falciparum sporozoite development in the midgut of Anopheles mosquitoes. Eukaryot. Cell 2014, 13, 550–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, M.; Kumar, T.R.; Nkrumah, L.J.; Coppi, A.; Retzlaff, S.; Li, C.D.; Kelly, B.J.; Moura, P.A.; Lakshmanan, V.; Freundlich, J.S.; et al. The fatty acid biosynthesis enzyme FabI plays a key role in the development of liver-stage malarial parasites. Cell Host Microbe 2008, 4, 567–578. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, A.M.; O’Neill, M.T.; Tarun, A.S.; Camargo, N.; Phuong, T.M.; Aly, A.S.; Cowman, A.F.; Kappe, S.H. Type II fatty acid synthesis is essential only for malaria parasite late liver stage development. Cell. Microbiol. 2009, 11, 506–520. [Google Scholar] [CrossRef] [Green Version]
- Tasdemir, D.; Sanabria, D.; Lauinger, I.L.; Tarun, A.; Herman, R.; Perozzo, R.; Zloh, M.; Kappe, S.H.; Brun, R.; Carballeira, N.M. 2-Hexadecynoic acid inhibits plasmodial FAS-II enzymes and arrests erythrocytic and liver stage Plasmodium infections. Bioorg. Med. Chem. 2010, 18, 7475–7485. [Google Scholar] [CrossRef] [Green Version]
- Carballeira, N.M.; Bwalya, A.G.; Itoe, M.A.; Andricopulo, A.D.; Cordero-Maldonado, M.L.; Kaiser, M.; Mota, M.M.; Crawford, A.D.; Guido, R.V.; Tasdemir, D. 2-Octadecynoic acid as a dual life stage inhibitor of Plasmodium infections and plasmodial FAS-II enzymes. Bioorg. Med. Chem. Lett. 2014, 24, 4151–4157. [Google Scholar] [CrossRef]
- Lauinger, I.L.; Vivas, L.; Perozzo, R.; Stairiker, C.; Tarun, A.; Zloh, M.; Zhang, X.; Xu, H.; Tonge, P.J.; Franzblau, S.G.; et al. Potential of lichen secondary metabolites against Plasmodium liver stage parasites with FAS-II as the potential target. J. Nat. Prod. 2013, 76, 1064–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freundlich, J.S.; Anderson, J.W.; Sarantakis, D.; Shieh, H.M.; Yu, M.; Valderramos, J.C.; Lucumi, E.; Kuo, M.; Jacobs, W.R., Jr.; Fidock, D.A.; et al. Synthesis, biological activity, and X-ray crystal structural analysis of diaryl ether inhibitors of malarial enoyl acyl carrier protein reductase. Part 1: 4’-substituted triclosan derivatives. Bioorg. Med. Chem. Lett. 2005, 15, 5247–5252. [Google Scholar] [CrossRef] [PubMed]
- Surolia, A.; Ramya, T.N.; Ramya, V.; Surolia, N. ‘FAS’t inhibition of malaria. Biochem. J. 2004, 383, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Sauleau, P.; Moriou, C.; Al Mourabit, A. Metabolomics approach to chemical diversity of the Mediterranean marine sponge Agelas oroides. Nat. Prod. Res. 2017, 31, 1625–1632. [Google Scholar] [CrossRef] [PubMed]
- Tasdemir, D.; Topaloglu, B.; Perozzo, R.; Brun, R.; O’Neill, R.; Carballeira, N.M.; Zhang, X.; Tonge, P.J.; Linden, A.; Ruedi, P. Marine natural products from the Turkish sponge Agelas oroides that inhibit the enoyl reductases from Plasmodium falciparum, Mycobacterium tuberculosis and Escherichia coli. Bioorg. Med. Chem. 2007, 15, 6834–6845. [Google Scholar] [CrossRef]
- Kumar, G.; Parasuraman, P.; Sharma, S.K.; Banerjee, T.; Karmodiya, K.; Surolia, N.; Surolia, A. Discovery of a rhodanine class of compounds as inhibitors of Plasmodium falciparum enoyl-acyl carrier protein reductase. J. Med. Chem. 2007, 50, 2665–2675. [Google Scholar] [CrossRef]
- Kumar, G.; Banerjee, T.; Kapoor, N.; Surolia, N.; Surolia, A. SAR and pharmacophore models for the rhodanine inhibitors of Plasmodium falciparum enoyl-acyl carrier protein reductase. IUBMB Life 2010, 62, 204–213. [Google Scholar] [CrossRef]
- Samuel, M.; Oliver, S.V.; Wood, O.R.; Coetzee, M.; Brooke, B.D. Evaluation of the toxicity and repellence of an organic fatty acids mixture (C8910) against insecticide susceptible and resistant strains of the major malaria vector Anopheles funestus Giles (Diptera: Culicidae). Parasit Vectors 2015, 8, 321. [Google Scholar] [CrossRef] [Green Version]
- Delhaye, J.; Glaizot, O.; Christe, P. The effect of dietary antioxidant supplementation in a vertebrate host on the infection dynamics and transmission of avian malaria to the vector. Parasitol. Res. 2018, 117, 2043–2052. [Google Scholar] [CrossRef] [Green Version]
- Godfrey, D.G. Antiparasitic action of dietary cod liver oil upon Plasmodium berghei and its reversal by vitamin E. Exp. Parasitol. 1957, 6, 555–565. [Google Scholar] [CrossRef]
- Levander, O.A.; Ager, A.L., Jr.; Morris, V.C.; May, R.G. Plasmodium yoelii: Comparative antimalarial activities of dietary fish oils and fish oil concentrates in vitamin E-deficient mice. Exp. Parasitol. 1990, 70, 323–329. [Google Scholar] [CrossRef]
- Fujikawa, M.; Kamitani, T.; Tunru, I.; Yamazaki, K.; Hamazaki, T. Antimalarial effects of purified and α-tocopherol—Fortified n-3 polyunsaturated fatty acids. J. Nutr. Biochem. 1993, 4, 153–157. [Google Scholar] [CrossRef]
- Endres, S.; Ghorbani, R.; Kelley, V.E.; Georgilis, K.; Lonnemann, G.; van der Meer, J.W.; Cannon, J.G.; Rogers, T.S.; Klempner, M.S.; Weber, P.C.; et al. The effect of dietary supplementation with n-3 polyunsaturated fatty acids on the synthesis of interleukin-1 and tumor necrosis factor by mononuclear cells. N. Engl. J. Med. 1989, 320, 265–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fevang, P.; Saav, H.; Hostmark, A.T. Dietary fish oils and long-term malaria protection in mice. Lipids 1995, 30, 437–441. [Google Scholar] [CrossRef] [PubMed]
- Krugliak, M.; Deharo, E.; Shalmiev, G.; Sauvain, M.; Moretti, C.; Ginsburg, H. Antimalarial effects of C18 fatty acids on Plasmodium falciparum in culture and on Plasmodium vinckei petteri and Plasmodium yoelii nigeriensis in vivo. Exp. Parasitol. 1995, 81, 97–105. [Google Scholar] [CrossRef]
- Kumaratilake, L.M.; Ferrante, A.; Robinson, B.S.; Jaeger, T.; Poulos, A. Enhancement of neutrophil-mediated killing of Plasmodium falciparum asexual blood forms by fatty acids: Importance of fatty acid structure. Infect. Immun. 1997, 65, 4152–4157. [Google Scholar] [CrossRef] [Green Version]
- Arun Kumar, C.; Das, U.N. Lipid peroxides, nitric oxide and essential fatty acids in patients with Plasmodium falciparum malaria. Prostaglandins Leukot Essent Fatty Acids 1999, 61, 255–258. [Google Scholar] [CrossRef]
- Hamazaki, K.; Syafruddin, D.; Tunru, I.S.; Azwir, M.F.; Asih, P.B.; Sawazaki, S.; Hamazaki, T. The effects of docosahexaenoic acid-rich fish oil on behavior, school attendance rate and malaria infection in school children—a double-blind, randomized, placebo-controlled trial in Lampung, Indonesia. Asia Pac. J. Clin. Nutr. 2008, 17, 258–263. [Google Scholar]
- Melariri, P.; Campbell, W.; Etusim, P.; Smith, P. In Vitro and in Vivo Antimalarial Activity of Linolenic and Linoleic Acids and their Methyl Esters. Adv. Stud. Biol. 2012, 4, 333–349. [Google Scholar]
- Villegas-Mendez, A.; Greig, R.; Shaw, T.N.; de Souza, J.B.; Gwyer Findlay, E.; Stumhofer, J.S.; Hafalla, J.C.; Blount, D.G.; Hunter, C.A.; Riley, E.M.; et al. IFN-gamma-producing CD4+ T cells promote experimental cerebral malaria by modulating CD8+ T cell accumulation within the brain. J. Immunol. 2012, 189, 968–979. [Google Scholar] [CrossRef] [Green Version]
- Ademolue, T.W.; Aniweh, Y.; Kusi, K.A.; Awandare, G.A. Patterns of inflammatory responses and parasite tolerance vary with malaria transmission intensity. Malar. J. 2017, 16, 145. [Google Scholar] [CrossRef] [PubMed]
- Clark, I.A.; Alleva, L.M.; Budd, A.C.; Cowden, W.B. Understanding the role of inflammatory cytokines in malaria and related diseases. Travel Med. Infect. Dis. 2008, 6, 67–81. [Google Scholar] [CrossRef] [PubMed]
- Gozzelino, R.; Jeney, V.; Soares, M.P. Mechanisms of cell protection by heme oxygenase-1. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 323–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.Z.; Zhang, Y.C.; Cheng, J.S.; Ni, Q.; Li, P.J.; Wang, S.W.; Han, W.; Zhang, Y.L. BML-111, a lipoxin receptor agonist, ameliorates ‘two-hit’-induced acute pancreatitis-associated lung injury in mice by the upregulation of heme oxygenase-1. Artif. Cells Nanomed. Biotechnol. 2014, 42, 110–120. [Google Scholar] [CrossRef]
- Lv, W.; Lv, C.; Yu, S.; Yang, Y.; Kong, H.; Xie, J.; Sun, H.; Andersson, R.; Xu, D.; Chen, B.; et al. Lipoxin A4 attenuation of endothelial inflammation response mimicking pancreatitis-induced lung injury. Exp. Biol. Med. 2013, 238, 1388–1395. [Google Scholar] [CrossRef]
- Belhaj, A.; Dewachter, L.; Kerbaul, F.; Brimioulle, S.; Dewachter, C.; Naeije, R.; Rondelet, B. Heme oxygenase-1 and inflammation in experimental right ventricular failure on prolonged overcirculation-induced pulmonary hypertension. PLoS ONE 2013, 8, e69470. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, P.; Zhang, Z.; Hu, Q.; He, J.; Liu, H.; Zhao, J.; Liang, Y.; He, Z.; Li, X.; et al. LXA4 ameliorates cerebrovascular endothelial dysfunction by reducing acute inflammation after subarachnoid hemorrhage in rats. Neuroscience 2019, 408, 105–114. [Google Scholar] [CrossRef]
- Pamplona, A.; Ferreira, A.; Balla, J.; Jeney, V.; Balla, G.; Epiphanio, S.; Chora, A.; Rodrigues, C.D.; Gregoire, I.P.; Cunha-Rodrigues, M.; et al. Heme oxygenase-1 and carbon monoxide suppress the pathogenesis of experimental cerebral malaria. Nat. Med. 2007, 13, 703–710. [Google Scholar] [CrossRef]
- Padua, T.A.; Torres, N.D.; Candea, A.L.P.; Costa, M.F.S.; Silva, J.D.; Silva-Filho, J.L.; Costa, F.T.M.; Rocco, P.R.M.; Souza, M.C.; Henriques, M.G. Therapeutic effect of Lipoxin A4 in malaria-induced acute lung injury. J. Leukoc. Biol. 2018, 103, 657–670. [Google Scholar] [CrossRef]
- Zuzarte-Luis, V.; Mello-Vieira, J.; Marreiros, I.M.; Liehl, P.; Chora, A.F.; Carret, C.K.; Carvalho, T.; Mota, M.M. Dietary alterations modulate susceptibility to Plasmodium infection. Nat. Microbiol. 2017, 2, 1600–1607. [Google Scholar] [CrossRef]
- Luisi, M.L.E.; Lucarini, L.; Biffi, B.; Rafanelli, E.; Pietramellara, G.; Durante, M.; Vidali, S.; Provensi, G.; Madiai, S.; Gheri, C.F.; et al. Effect of Mediterranean Diet Enriched in High Quality Extra Virgin Olive Oil on Oxidative Stress, Inflammation and Gut Microbiota in Obese and Normal Weight Adult Subjects. Front. Pharmacol. 2019, 10, 1366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serra-Majem, L.; Roman-Vinas, B.; Sanchez-Villegas, A.; Guasch-Ferre, M.; Corella, D.; La Vecchia, C. Benefits of the Mediterranean diet: Epidemiological and molecular aspects. Mol. Asp. Med. 2019, 67, 1–55. [Google Scholar] [CrossRef] [PubMed]
- Schwingshackl, L.; Morze, J.; Hoffmann, G. Mediterranean diet and health status: Active ingredients and pharmacological mechanisms. Br. J. Pharmacol. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Structures of typical omega-6, omega-3, and omega-9 fatty acids.
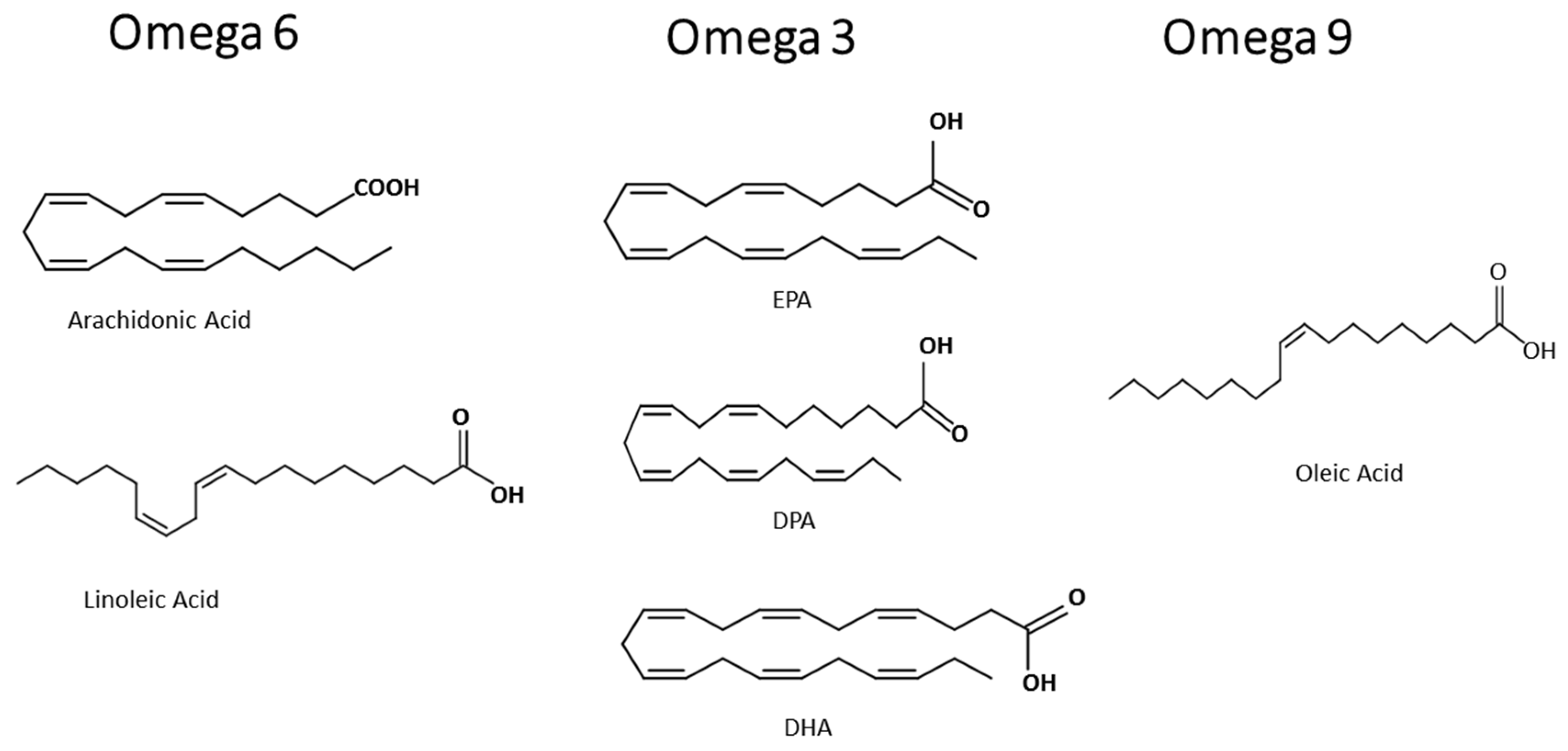
Figure 2.
Effects of omega-6, omega-3, and omega-9 on inflammation in host cells. IL—interleukin, CXCL8—C-X-C motif chemokine ligand 8, NFκB nuclear factor kappa B, Th—T helper cells, IL—interleukin, TNF-α—tumor necrosis factor alpha, CPT1A—carnitine palmitoyltransferase 1A, UCP2—uncoupling protein 2, AMPK—5′ adenosine monophosphate-activated protein kinase, ROS—reactive oxygen species, and NEFA—non-esterified fatty acid.
Figure 2.
Effects of omega-6, omega-3, and omega-9 on inflammation in host cells. IL—interleukin, CXCL8—C-X-C motif chemokine ligand 8, NFκB nuclear factor kappa B, Th—T helper cells, IL—interleukin, TNF-α—tumor necrosis factor alpha, CPT1A—carnitine palmitoyltransferase 1A, UCP2—uncoupling protein 2, AMPK—5′ adenosine monophosphate-activated protein kinase, ROS—reactive oxygen species, and NEFA—non-esterified fatty acid.
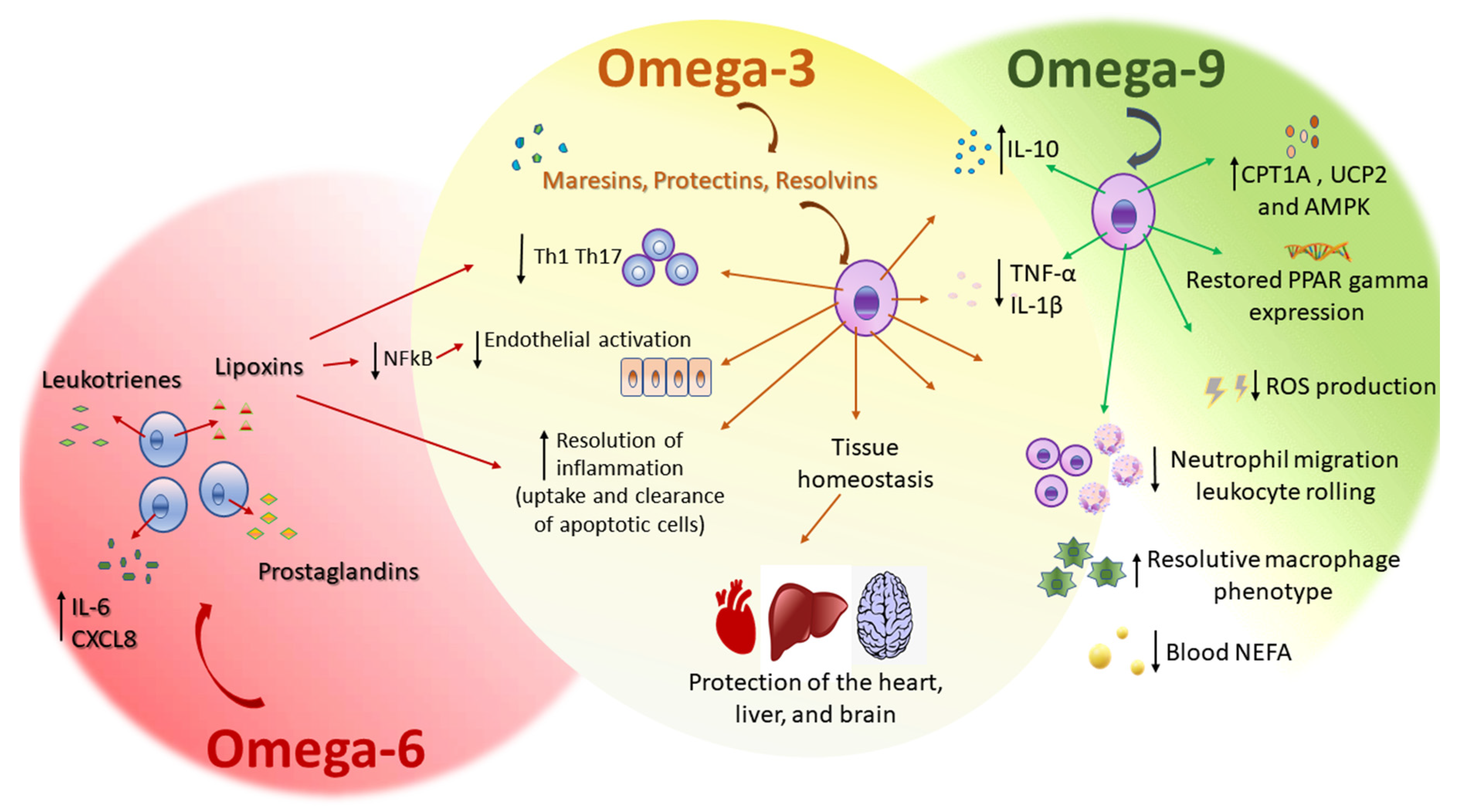

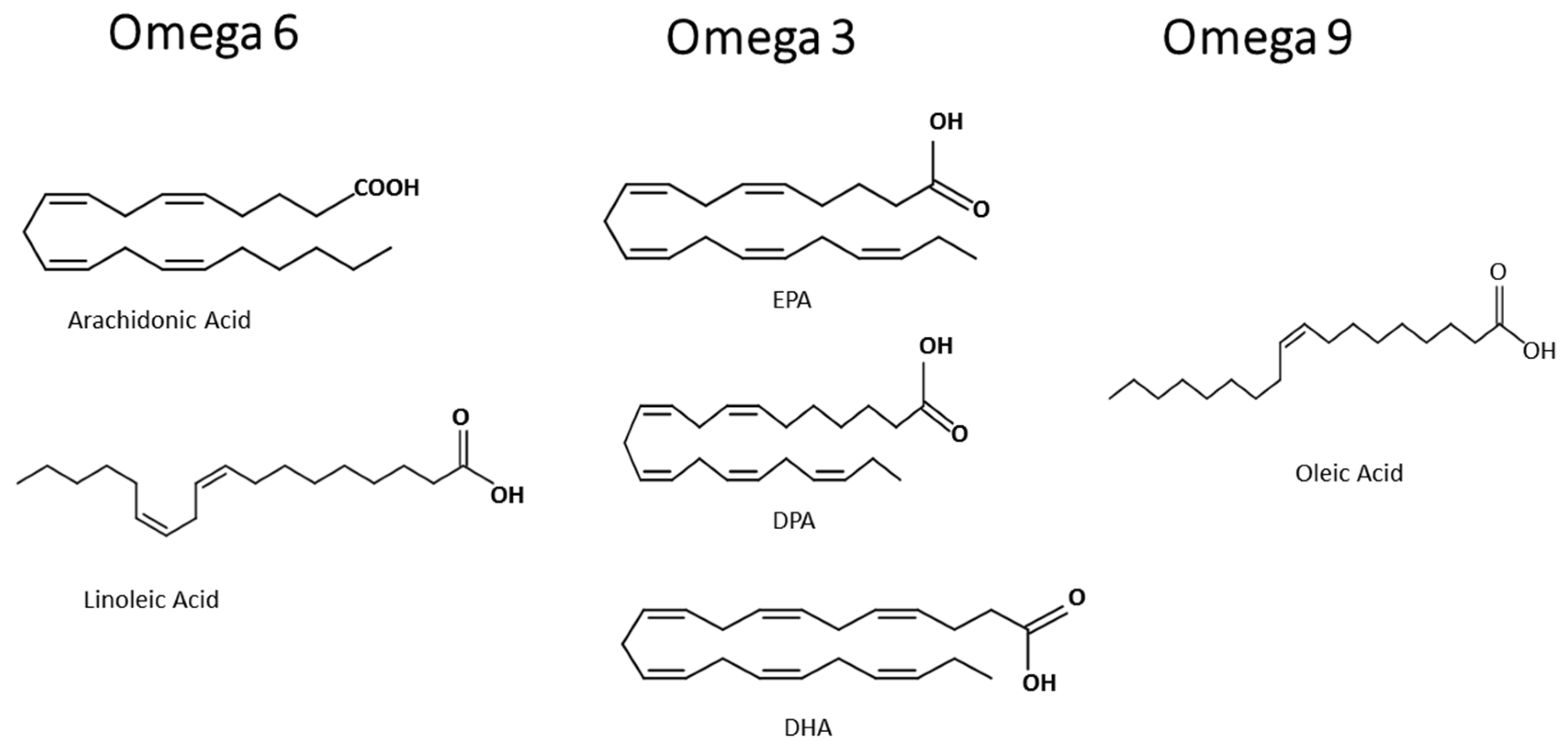{kind=link}
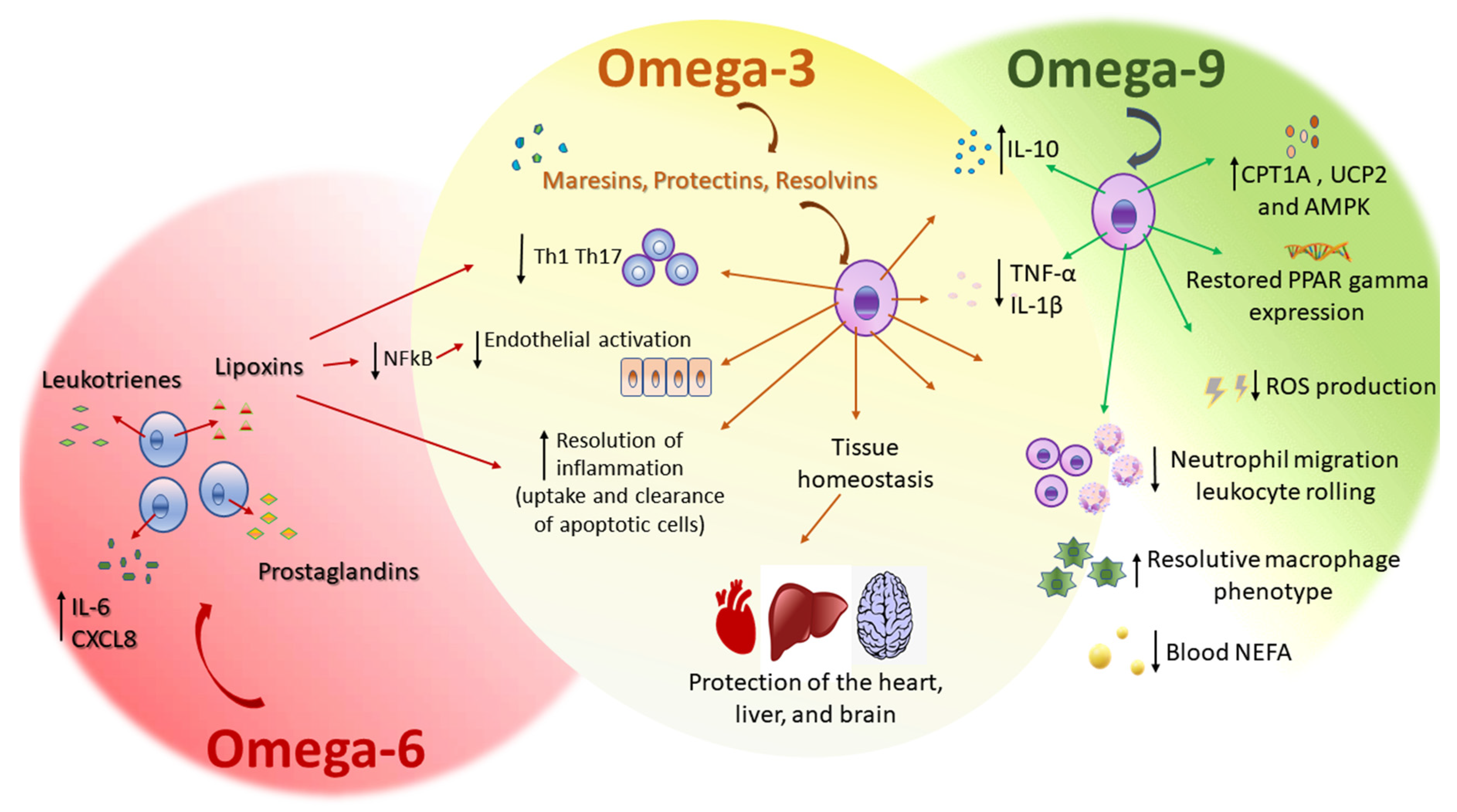{kind=link}
Table 1.
Daily nutritional suggested composition for the experimental MedDiet (2240 kcal/d).
| Nutrients | Amount |
|---|---|
| Carbohydrate (g) | 194.0 |
| Fiber (g) | 41.1 |
| Protein (g) | 87.3 |
| Fat (g) | 106.0 |
| SFA (g) | 20.9 |
| MUFA (g) | 52.5 |
| PUFA (g) | 24.8 |
| Cholesterol (mg) | 289.7 |
| Alcohol (g) | 11.1 |
| MUFA to SFA ratio | 2.7 |
| Omega-3 (g) | 5.9 |
| Omega-6 (g) | 18.7 |
| Cholesterol (mg) | 255.8 |
| Beta carotene (μg) | 3039 |
| Vitamin A (RE) | 2161.6 |
| Vitamin D (μg) | 3.2 |
| Vitamin E (mg) | 29.3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Silva, A.R.; Moraes, B.P.T.; Gonçalves-de-Albuquerque, C.F. Mediterranean Diet: Lipids, Inflammation, and Malaria Infection. Int. J. Mol. Sci. 2020, 21, 4489. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124489
AMA Style
Silva AR, Moraes BPT, Gonçalves-de-Albuquerque CF. Mediterranean Diet: Lipids, Inflammation, and Malaria Infection. International Journal of Molecular Sciences. 2020; 21(12):4489. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124489
Chicago/Turabian StyleSilva, Adriana R., Bianca P. T. Moraes, and Cassiano F. Gonçalves-de-Albuquerque. 2020. "Mediterranean Diet: Lipids, Inflammation, and Malaria Infection" International Journal of Molecular Sciences 21, no. 12: 4489. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124489
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.