Screening of an Epigenetic Drug Library Identifies 4-((hydroxyamino)carbonyl)-N-(2-hydroxyethyl)-N-Phenyl-Benzeneacetamide that Reduces Melanin Synthesis by Inhibiting Tyrosinase Activity Independently of Epigenetic Mechanisms



Abstract
:1. Introduction
2. Results
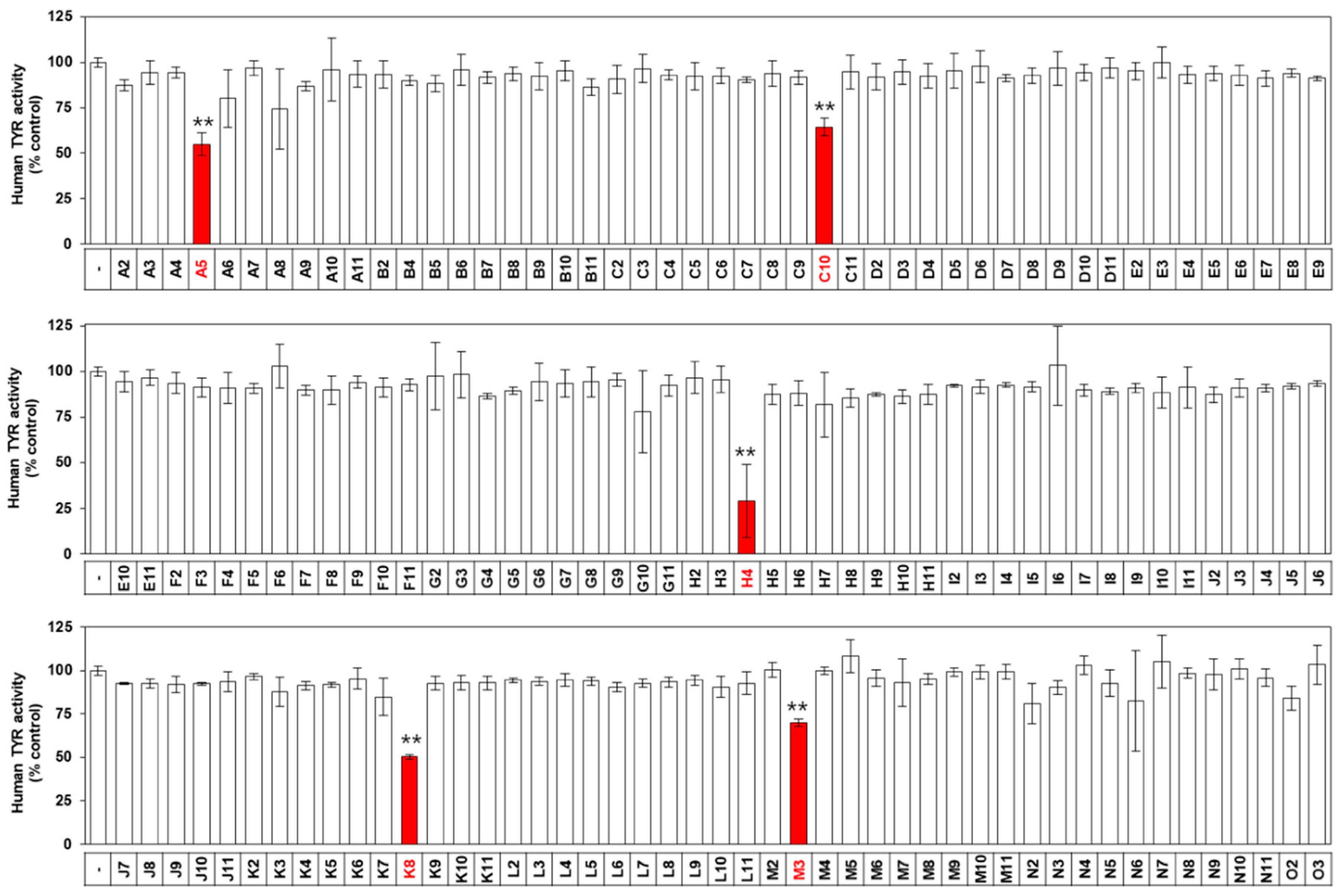
2.1. Screening of a Drug Library for Antimelanogenic Activity
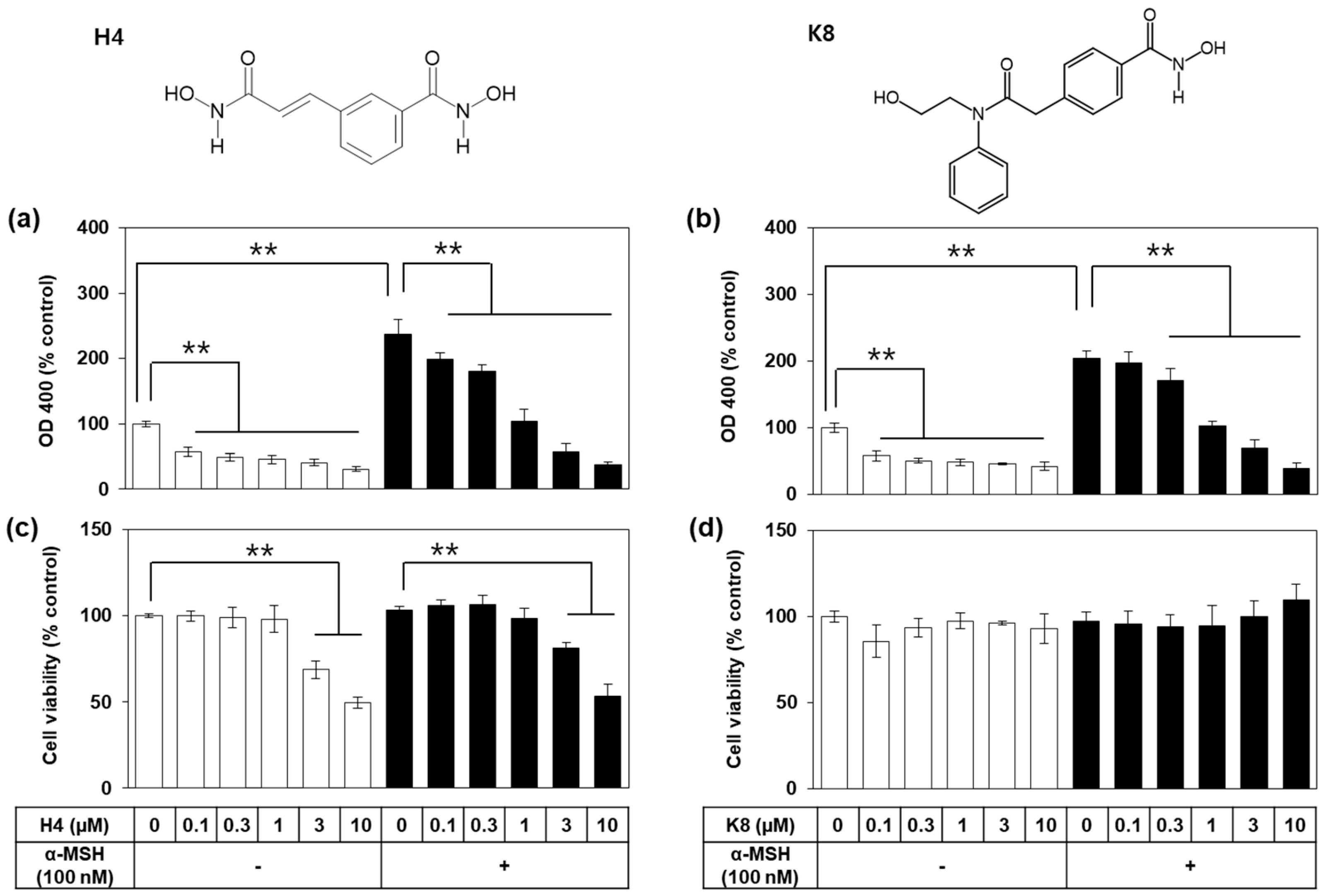
2.2. Activity Versus Cytotoxicity for H4 and K8
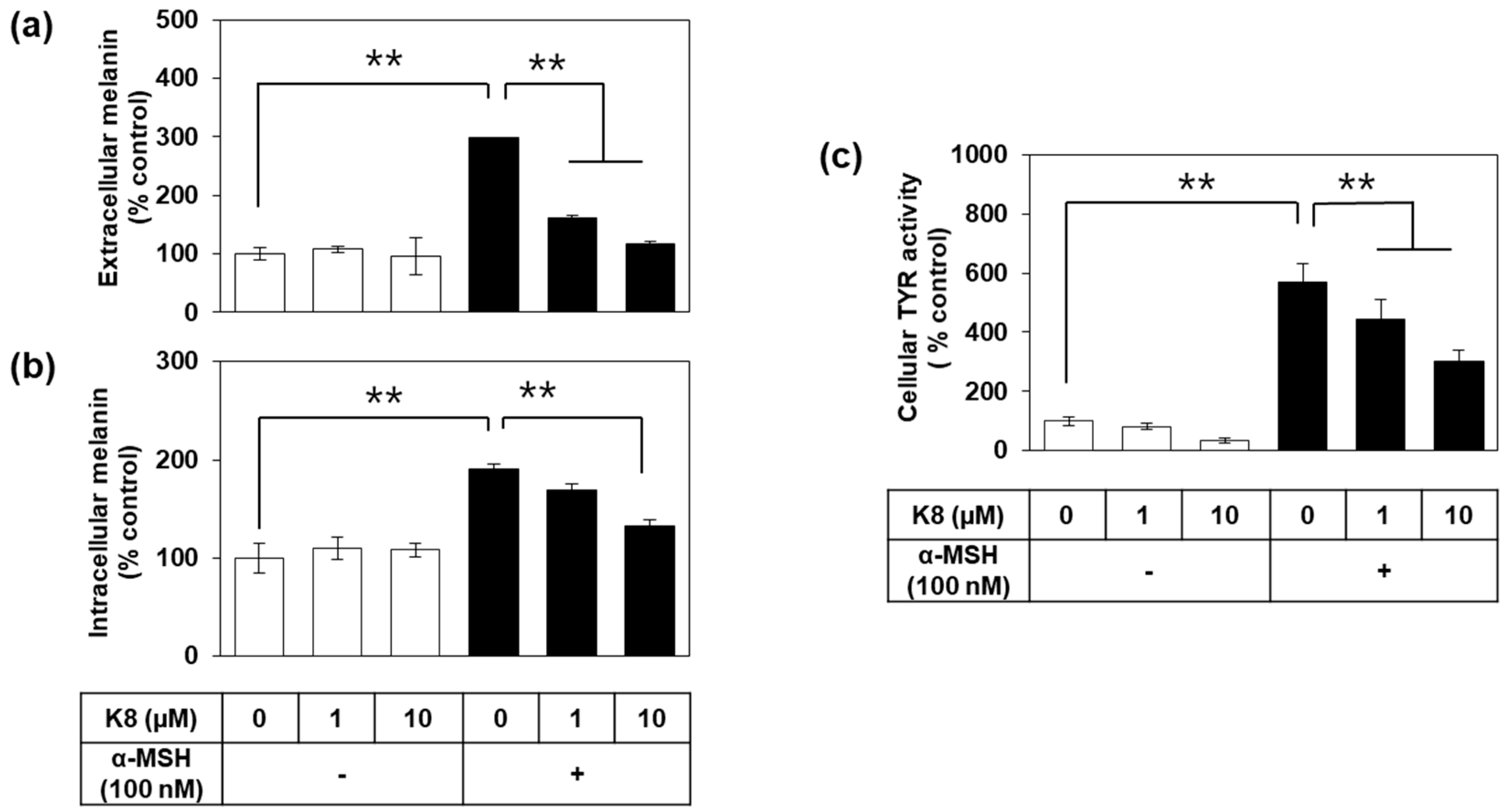
2.3. Effects of K8 on Melanin Content and TYR Activity in Cells
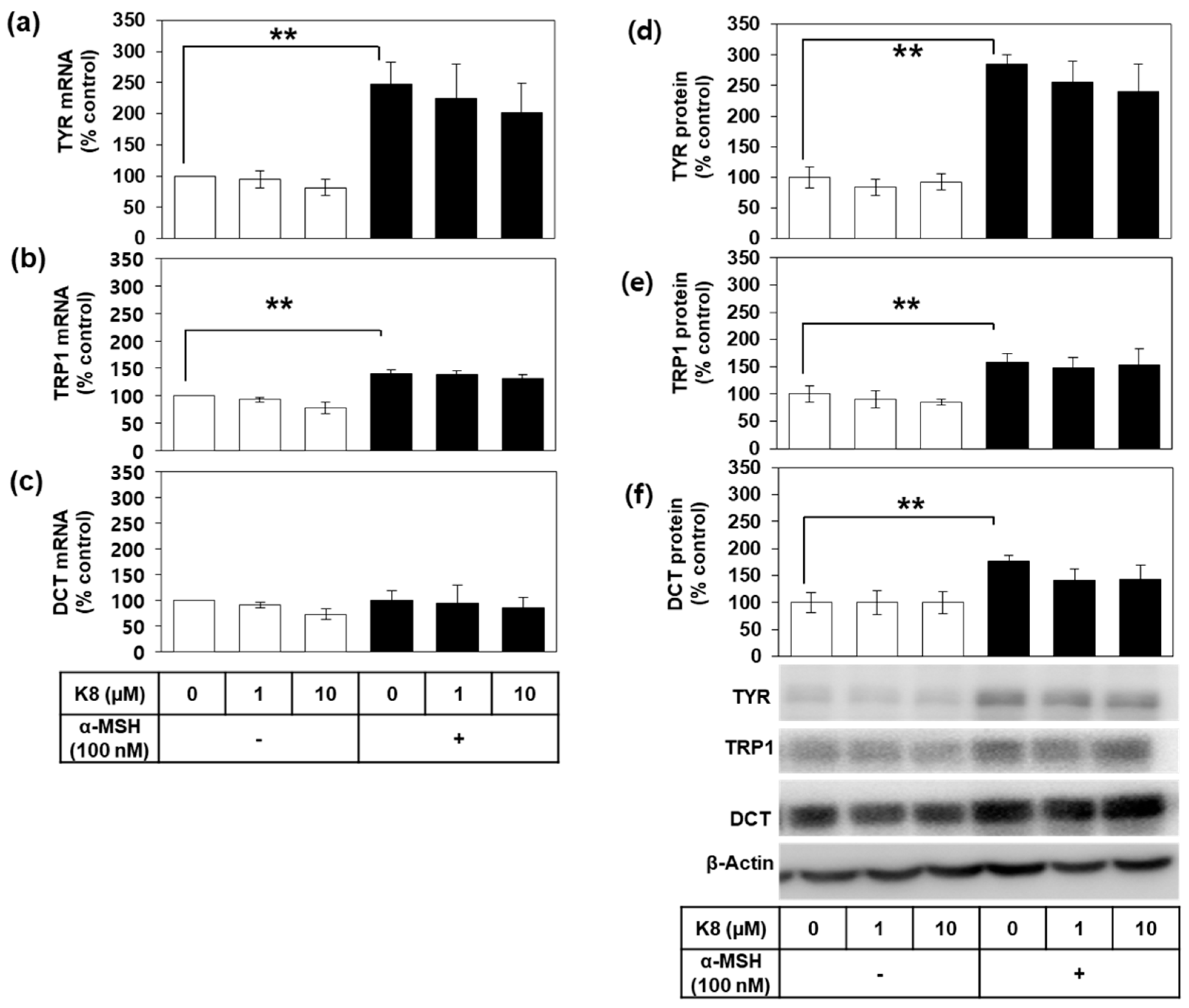
2.4. Effects of K8 on the mRNA and the Protein Expression Levels of TYR, TRP1, and DCT
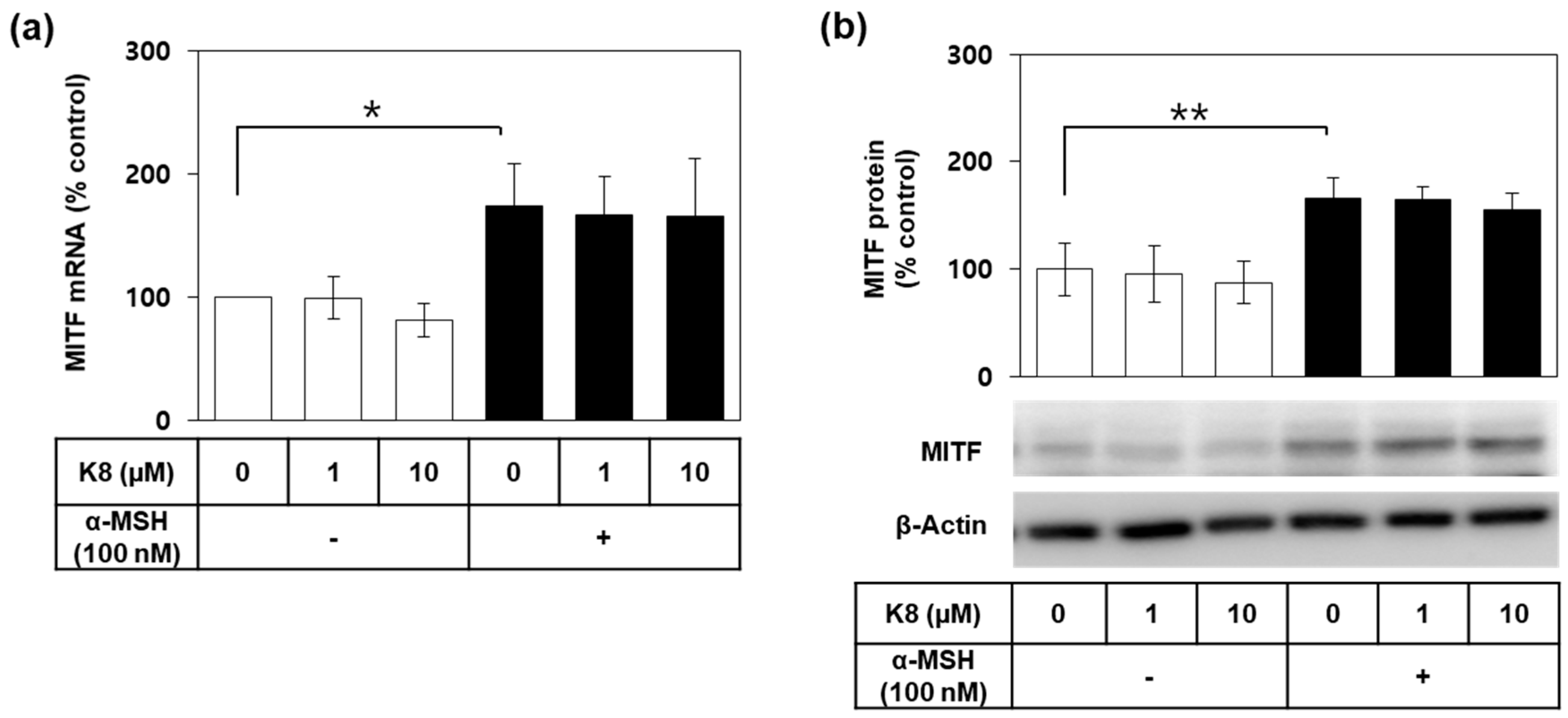
2.5. Effects of K8 on the mRNA and the Protein Expression Levels of MITF
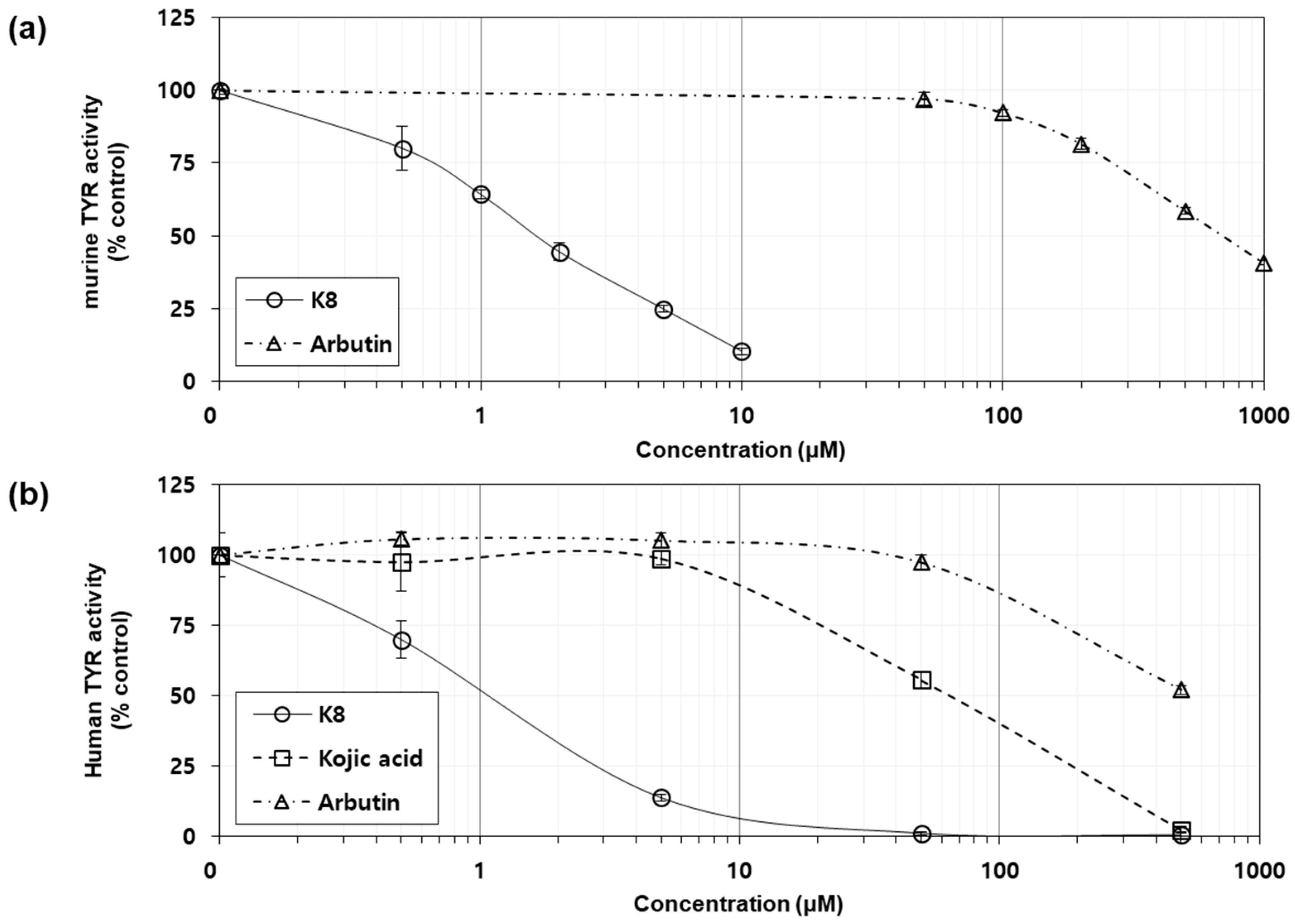
2.6. Effects of K8 on the Catalytic Activity of Murine and Human TYRs
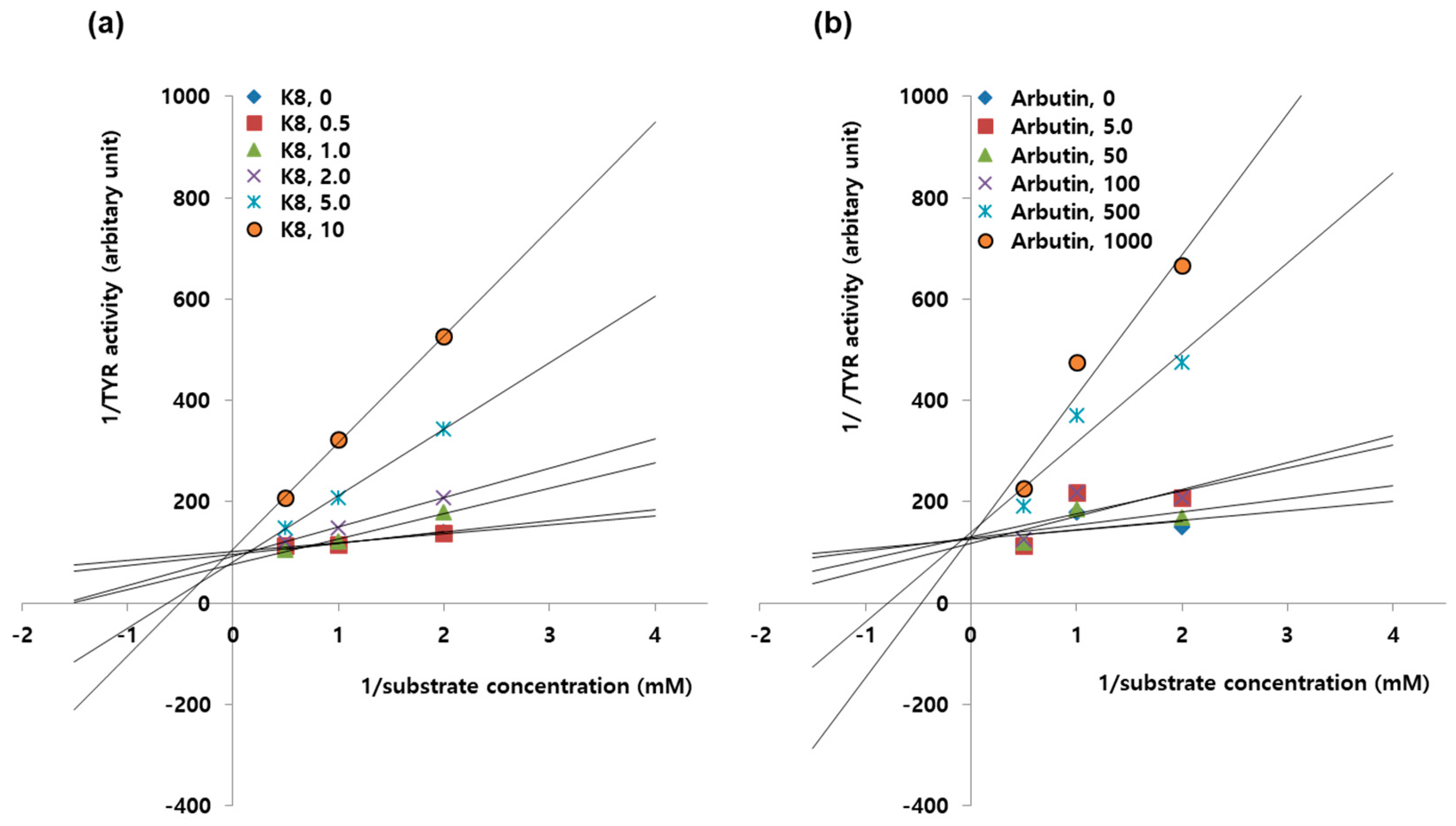
2.7. Enzyme Kinetic Study to Determine Inhibition Types of K8 and β-arbutin.
2.8. Copper Chelating Activity of K8 and Kojic Acid
2.9. Screening of a Drug Library for TYR Inhibitory Activity
3. Discussion
4. Materials and Methods
4.1. Reagents and a Drug Library
4.2. Cell Culture
4.3. Screening Assay for Overall Melanin Synthesis
4.4. Cell Viability Assay
4.5. Melanin Content Assay
4.6. Cellular TYR Activity Assay
4.7. qRT-PCR Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | GenBank Accession Number | Primer Sequences | References |
|---|---|---|---|
| TYR | NM_011661.5 | Forward: 5′-CTTCTTCTCCTCCTGGCAGATC-3′ Reverse: 5′-TGGGGGTTTTGGCTTTGTC-3′ | [60] |
| TYRP1 | NM_001282015.1 | Forward: 5′-CAGTGCAGCGTCTTCCTGAG-3′ Reverse: 5′-TTCCCGTGGGAGCACTGTAA-3′ | [61] |
| DCT | NM_010024.3 | Forward: 5′-GCAAGAGATACACGGAGGAAG-3′ Reverse: 5′-CTAAGGCATCATCATCATCACTAC-3′ | [62] |
| MITF | NM_008601.3 | Forward: 5′-GCTGGAAATGCTAGAATACAG-3′ Reverse: 5′-TTCCAGGCTGATGATGTCATC-3′ | [60] |
| GAPDH | NM_001289726.1 | Forward: 5′-GCATCTCCCTCACAATTTCCA-3′ Reverse: 5′-GTGCAGCGAACTTTATTGATGG-3′ | [63] |
4.8. Western Blotting
4.9. In Vitro TYR Catalytic Activity Assay
4.10. Copper Chelating Activity Assay
4.11. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| MITF | microphthalmia-associated transcription factor |
| TYR | tyrosinase |
| HDAC | histone deacetylase |
| CBHA | N-hydroxy-3-(3-(hydroxyamino)-3-oxo-1-propen-1-yl)-benzamide |
| HPOB | 4-((hydroxyamino)carbonyl)-N-(2-hydroxyethyl)-N-phenyl-benzeneacetamide |
| α-MSH | alpha-melanocyte-stimulating hormone |
| L-DOPA | L-3,4-dihydroxyphenylalanine |
| TYRP1 | tyrosinase-related protein 1 |
| DCT | dopachrome tautomerase |
| GAPDH | glyceraldehyde 3-phosphate dehydrogenase |
| OD | optical density |
| qRT-PCR | quantitative reverse transcription polymerase chain |
| MC1R | melanocortin 1 receptor |
References
- Costin, G.E.; Hearing, V.J. Human skin pigmentation: Melanocytes modulate skin color in response to stress. Faseb J. 2007, 21, 976–994. [Google Scholar] [CrossRef] [PubMed]
- Schiaffino, M.V. Signaling pathways in melanosome biogenesis and pathology. Int. J. Biochem. Cell Biol. 2010, 42, 1094–1104. [Google Scholar] [CrossRef] [Green Version]
- Cardinali, G.; Ceccarelli, S.; Kovacs, D.; Aspite, N.; Lotti, L.V.; Torrisi, M.R.; Picardo, M. Keratinocyte growth factor promotes melanosome transfer to keratinocytes. J. Investig. Dermatol. 2005, 125, 1190–1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epstein, J.H. Photocarcinogenesis, skin cancer, and aging. J. Am. Acad. Dermatol. 1983, 9, 487–502. [Google Scholar] [CrossRef]
- Slominski, A.; Kim, T.K.; Brozyna, A.A.; Janjetovic, Z.; Brooks, D.L.; Schwab, L.P.; Skobowiat, C.; Jozwicki, W.; Seagroves, T.N. The role of melanogenesis in regulation of melanoma behavior: Melanogenesis leads to stimulation of HIF-1alpha expression and HIF-dependent attendant pathways. Arch. Biochem. Biophys. 2014, 563, 79–93. [Google Scholar] [CrossRef] [Green Version]
- Slominski, R.M.; Zmijewski, M.A.; Slominski, A.T. The role of melanin pigment in melanoma. Exp Dermatol. 2015, 24, 258–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fistarol, S.K.; Itin, P.H. Disorders of pigmentation. J. Dtsch. Dermatol. Ges. 2010, 8, 187–201. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Tobin, D.J.; Shibahara, S.; Wortsman, J. Melanin pigmentation in mammalian skin and its hormonal regulation. Physiol. Rev. 2004, 84, 1155–1228. [Google Scholar] [CrossRef]
- Rose, P.T. Pigmentary disorders. Med. Clin. N. Am. 2009, 93, 1225–1239. [Google Scholar] [CrossRef]
- Callender, V.D.; St Surin-Lord, S.; Davis, E.C.; Maclin, M. Postinflammatory hyperpigmentation: Etiologic and therapeutic considerations. Am. J. Clin. Dermatol. 2011, 12, 87–99. [Google Scholar] [CrossRef]
- Haltaufderhyde, K.D.; Oancea, E. Genome-wide transcriptome analysis of human epidermal melanocytes. Genomics 2014, 104, 482–489. [Google Scholar] [CrossRef] [PubMed]
- Soejima, M.; Koda, Y. Population differences of two coding SNPs in pigmentation-related genes SLC24A5 and SLC45A2. Int. J. Leg. Med. 2007, 121, 36–39. [Google Scholar] [CrossRef] [PubMed]
- Ginger, R.S.; Askew, S.E.; Ogborne, R.M.; Wilson, S.; Ferdinando, D.; Dadd, T.; Smith, A.M.; Kazi, S.; Szerencsei, R.T.; Winkfein, R.J.; et al. SLC24A5 encodes a trans-Golgi network protein with potassium-dependent sodium-calcium exchange activity that regulates human epidermal melanogenesis. J. Biol. Chem. 2008, 283, 5486–5495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, A.L.; Chen, W.; Thurber, A.E.; Smit, D.J.; Smith, A.G.; Bladen, T.G.; Brown, D.L.; Duffy, D.L.; Pastorino, L.; Bianchi-Scarra, G.; et al. Analysis of cultured human melanocytes based on polymorphisms within the SLC45A2/MATP, SLC24A5/NCKX5, and OCA2/P loci. J. Investig. Dermatol. 2009, 129, 392–405. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.B.; Cho, R.J. Genetics and epigenetics of the skin meet deep sequence. J. Investig. Dermatol. 2012, 132, 923–932. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.S.; Jeong, H.S.; Kim, M.K.; Yun, H.Y.; Baek, K.J.; Kwon, N.S.; Kim, D.S. The DNA methylation inhibitor 5-azacytidine decreases melanin synthesis by inhibiting CREB phosphorylation. Pharmazie 2015, 70, 646–649. [Google Scholar]
- Yokoyama, S.; Feige, E.; Poling, L.L.; Levy, C.; Widlund, H.R.; Khaled, M.; Kung, A.L.; Fisher, D.E. Pharmacologic suppression of MITF expression via HDAC inhibitors in the melanocyte lineage. Pigment Cell Melanoma Res. 2008, 21, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.H.; Bin, B.H.; Kim, J.; Dong, S.E.; Park, P.J.; Choi, H.; Kim, B.J.; Yu, S.J.; Kang, H.; Kang, H.H.; et al. Novel inhibitory function of miR-125b in melanogenesis. Pigment Cell Melanoma Res. 2014, 27, 140–144. [Google Scholar] [CrossRef]
- Kim, K.H.; Lee, T.R.; Cho, E.G. SH3BP4, a novel pigmentation gene, is inversely regulated by miR-125b and MITF. Exp. Mol. Med. 2017, 49, e367. [Google Scholar] [CrossRef] [Green Version]
- Cooksey, C.J.; Garratt, P.J.; Land, E.J.; Pavel, S.; Ramsden, C.A.; Riley, P.A.; Smit, N.P. Evidence of the indirect formation of the catecholic intermediate substrate responsible for the autoactivation kinetics of tyrosinase. J. Biol. Chem. 1997, 272, 26226–26235. [Google Scholar] [CrossRef] [Green Version]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V. Skin whitening agents: Medicinal chemistry perspective of tyrosinase inhibitors. J. Enzym. Inhib. Med. Chem. 2017, 32, 403–425. [Google Scholar] [CrossRef] [Green Version]
- Zolghadri, S.; Bahrami, A.; Hassan Khan, M.T.; Munoz-Munoz, J.; Garcia-Molina, F.; Garcia-Canovas, F.; Saboury, A.A. A comprehensive review on tyrosinase inhibitors. J. Enzym. Inhib. Med. Chem. 2019, 34, 279–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, K.; Fukuda, M. Arbutin: Mechanism of its depigmenting action in human melanocyte culture. J. Pharmacol. Exp. 1996, 276, 765–769. [Google Scholar]
- Okajima, S.; Hamamoto, A.; Asano, M.; Isogawa, K.; Ito, H.; Kato, S.; Hirata, Y.; Furuta, K.; Takemori, H. Azepine derivative T4FAT, a new copper chelator, inhibits tyrosinase. Biochem. Biophys. Res. Commun. 2019, 509, 209–215. [Google Scholar] [CrossRef]
- Lee, J.H.; Mahendran, A.; Yao, Y.; Ngo, L.; Venta-Perez, G.; Choy, M.L.; Kim, N.; Ham, W.S.; Breslow, R.; Marks, P.A. Development of a histone deacetylase 6 inhibitor and its biological effects. Proc. Natl. Acad. Sci. USA 2013, 110, 15704–15709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maresca, V.; Flori, E.; Picardo, M. Skin phototype: A new perspective. Pigment Cell Melanoma Res. 2015, 28, 378–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasti, T.H.; Timares, L. MC1R, eumelanin and pheomelanin: Their role in determining the susceptibility to skin cancer. Photochem. Photobiol. 2015, 91, 188–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maymone, M.B.C.; Neamah, H.H.; Secemsky, E.A.; Vashi, N.A. Correlating the Dermatology Life Quality Index and Skin Discoloration Impact Evaluation Questionnaire tools in disorders of hyperpigmentation. J. Dermatol. 2018, 45, 361–362. [Google Scholar] [CrossRef]
- Pavlic, V.; Brkic, Z.; Marin, S.; Cicmil, S.; Gojkov-Vukelic, M.; Aoki, A. Gingival melanin depigmentation by Er:YAG laser: A literature review. J. Cosmet. Laser 2018, 20, 85–90. [Google Scholar] [CrossRef]
- Saxena, S.; Andersen, R.M.; Maibach, H.I. Pitfalls in clinical trials reveal need for well tolerated, more effective depigmenting agents. J. Dermatol. Treat. 2015, 26, 440–450. [Google Scholar] [CrossRef]
- Levy, L.L.; Emer, J.J. Emotional benefit of cosmetic camouflage in the treatment of facial skin conditions: Personal experience and review. Clin. Cosmet. Investig. Dermatol. 2012, 5, 173–182. [Google Scholar]
- Jow, T.; Hantash, B.M. Hydroquinone-induced depigmentation: Case report and review of the literature. Dermatitis 2014, 25, e1–e5. [Google Scholar] [CrossRef] [PubMed]
- Desmedt, B.; Courselle, P.; De Beer, J.O.; Rogiers, V.; Grosber, M.; Deconinck, E.; De Paepe, K. Overview of skin whitening agents with an insight into the illegal cosmetic market in Europe. J. Eur. Acad. Dermatol. Venereol. 2016, 30, 943–950. [Google Scholar] [CrossRef]
- Jo, D.J.; Seok, J.K.; Kim, S.Y.; Park, W.; Baek, J.H.; Kim, Y.M.; Boo, Y.C. Human skin-depigmenting effects of resveratryl triglycolate, a hybrid compound of resveratrol and glycolic acid. Int. J. Cosmet. Sci. 2018, 40, 256–262. [Google Scholar] [CrossRef]
- Ryu, J.H.; Seok, J.K.; An, S.M.; Baek, J.H.; Koh, J.S.; Boo, Y.C. A study of the human skin-whitening effects of resveratryl triacetate. Arch. Dermatol. Res. 2015, 307, 239–247. [Google Scholar] [CrossRef]
- Solano, F.; Briganti, S.; Picardo, M.; Ghanem, G. Hypopigmenting agents: An updated review on biological, chemical and clinical aspects. Pigment Cell Res. 2006, 19, 550–571. [Google Scholar] [CrossRef] [PubMed]
- Ando, H.; Kondoh, H.; Ichihashi, M.; Hearing, V.J. Approaches to identify inhibitors of melanin biosynthesis via the quality control of tyrosinase. J. Investig. Dermatol. 2007, 127, 751–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, J.D.; Peles, D.; Wakamatsu, K.; Ito, S. Current challenges in understanding melanogenesis: Bridging chemistry, biological control, morphology, and function. Pigment Cell Melanoma Res. 2009, 22, 563–579. [Google Scholar] [CrossRef] [PubMed]
- Mas, J.S.; Gerritsen, I.; Hahmann, C.; Jimenez-Cervantes, C.; Garcia-Borron, J.C. Rate limiting factors in melanocortin 1 receptor signalling through the cAMP pathway. Pigment Cell Res. 2003, 16, 540–547. [Google Scholar] [CrossRef] [PubMed]
- Flaherty, K.T.; Hodi, F.S.; Fisher, D.E. From genes to drugs: Targeted strategies for melanoma. Nat. Rev. Cancer 2012, 12, 349–361. [Google Scholar] [CrossRef]
- An, S.M.; Lee, S.I.; Choi, S.W.; Moon, S.W.; Boo, Y.C. p-Coumaric acid, a constituent of Sasa quelpaertensis Nakai, inhibits cellular melanogenesis stimulated by alpha-melanocyte stimulating hormone. Br. J. Dermatol. 2008, 159, 292–299. [Google Scholar] [CrossRef] [PubMed]
- An, S.M.; Koh, J.S.; Boo, Y.C. p-coumaric acid not only inhibits human tyrosinase activity in vitro but also melanogenesis in cells exposed to UVB. Phytother. Res. 2010, 24, 1175–1180. [Google Scholar] [PubMed]
- Boo, Y.C. p-Coumaric Acid as An Active Ingredient in Cosmetics: A Review Focusing on its Antimelanogenic Effects. Antioxidants (Basel) 2019, 8, 275. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Park, J.H.; Suh, H.J.; Lee, I.C.; Koh, J.; Boo, Y.C. Effects of resveratrol, oxyresveratrol, and their acetylated derivatives on cellular melanogenesis. Arch. Dermatol. Res. 2014, 306, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Boo, Y.C. Isolation of resveratrol from vitis viniferae caulis and its potent inhibition of human tyrosinase. Evid. Based Complement. Altern. Med. 2013, 2013, 645257. [Google Scholar] [CrossRef] [Green Version]
- Boo, Y.C. Human Skin Lightening Efficacy of Resveratrol and Its Analogs: From in Vitro Studies to Cosmetic Applications. Antioxidants (Basel) 2019, 8, 332. [Google Scholar] [CrossRef] [Green Version]
- Kwak, J.Y.; Seok, J.K.; Suh, H.J.; Choi, Y.H.; Hong, S.S.; Kim, D.S.; Boo, Y.C. Antimelanogenic effects of luteolin 7-sulfate isolated from Phyllospadix iwatensis Makino. Br. J. Dermatol. 2016, 175, 501–511. [Google Scholar] [CrossRef]
- Lee, S.W.; Kim, J.H.; Song, H.; Seok, J.K.; Hong, S.S.; Boo, Y.C. Luteolin 7-Sulfate Attenuates Melanin Synthesis through Inhibition of CREB- and MITF-Mediated Tyrosinase Expression. Antioxidants (Basel) 2019, 8, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seok, J.K.; Lee, S.W.; Choi, J.; Kim, Y.M.; Boo, Y.C. Identification of novel antimelanogenic hexapeptides via positional scanning of a synthetic peptide combinatorial library. Exp. Dermatol. 2017, 26, 742–744. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Seok, J.K.; Kim, Y.M.; Boo, Y.C. Identification of small peptides and glycinamide that inhibit melanin synthesis using a positional scanning synthetic peptide combinatorial library. Br. J. Dermatol. 2019, 181, 128–137. [Google Scholar] [CrossRef]
- Kim, M.; Park, J.; Song, K.; Kim, H.G.; Koh, J.S.; Boo, Y.C. Screening of plant extracts for human tyrosinase inhibiting effects. Int. J. Cosmet. Sci. 2012, 34, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; An, S.M.; Koh, J.S.; Jang, D.I.; Boo, Y.C. Use of non-melanocytic HEK293 cells stably expressing human tyrosinase for the screening of anti-melanogenic agents. J. Cosmet. Sci. 2011, 62, 515–523. [Google Scholar] [PubMed]
- Cheshmedzhieva, D.; Toshev, N.; Gerova, M.; Petrov, O.; Dudev, T. Hydroxamic acid derivatives as histone deacetylase inhibitors: A DFT study of their tautomerism and metal affinities/selectivities. J. Mol. Model. 2018, 24, 114. [Google Scholar] [CrossRef] [PubMed]
- Kwak, S.Y.; Yang, J.K.; Choi, H.R.; Park, K.C.; Kim, Y.B.; Lee, Y.S. Synthesis and dual biological effects of hydroxycinnamoyl phenylalanyl/prolyl hydroxamic acid derivatives as tyrosinase inhibitor and antioxidant. Bioorganic Med. Chem. Lett. 2013, 23, 1136–1142. [Google Scholar] [CrossRef] [PubMed]
- Baek, H.S.; Rho, H.S.; Yoo, J.W.; Aln, S.M.; Lee, J.Y.; Lee, J.; Kim, M.K.; Kim, D.H.; Chang, I.S. The inhibitory effect of new hydroxamic acid derivatives on melanogenesis. Bull. Korean Chem. Soc. 2008, 29, 43–46. [Google Scholar]
- Park, S.; Seok, J.K.; Kwak, J.Y.; Choi, Y.H.; Hong, S.S.; Suh, H.J.; Park, W.; Boo, Y.C. Anti-melanogenic effects of resveratryl triglycolate, a novel hybrid compound derived by esterification of resveratrol with glycolic acid. Arch. Dermatol. Res. 2016, 308, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Mitani, K.; Takano, F.; Kawabata, T.; Allam, A.E.; Ota, M.; Takahashi, T.; Yahagi, N.; Sakurada, C.; Fushiya, S.; Ohta, T. Suppression of melanin synthesis by the phenolic constituents of sappanwood (Caesalpinia sappan). Planta Med. 2013, 79, 37–44. [Google Scholar] [CrossRef]
- Garcia-Jimenez, A.; Teruel-Puche, J.A.; Berna, J.; Rodriguez-Lopez, J.N.; Tudela, J.; Garcia-Canovas, F. Action of tyrosinase on alpha and beta-arbutin: A kinetic study. PLoS ONE 2017, 12, e0177330. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Yang, R.; Zheng, Y.; Li, L.; Liu, S.; Burrows, M.; Wei, Z.; Nace, A.; Herlyn, M.; Cui, R.; Guo, W.; et al. Direct conversion of mouse and human fibroblasts to functional melanocytes by defined factors. Nat. Commun. 2014, 5, 5807. [Google Scholar] [CrossRef] [Green Version]
- Seo, G.Y.; Ha, Y.; Park, A.H.; Kwon, O.W.; Kim, Y.J. Leathesia difformis Extract Inhibits alpha-MSH-Induced Melanogenesis in B16F10 Cells via Down-Regulation of CREB Signaling Pathway. Int. J. Mol. Sci. 2019, 20, 536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsang, T.F.; Chan, B.; Tai, W.C.; Huang, G.; Wang, J.; Li, X.; Jiang, Z.H.; Hsiao, W.L.W. Gynostemma pentaphyllum saponins induce melanogenesis and activate cAMP/PKA and Wnt/beta-catenin signaling pathways. Phytomedicine 2019, 60, 153008. [Google Scholar] [CrossRef] [PubMed]
- Han, M.S.; Che, X.; Cho, G.H.; Park, H.R.; Lim, K.E.; Park, N.R.; Jin, J.S.; Jung, Y.K.; Jeong, J.H.; Lee, I.K.; et al. Functional Cooperation between Vitamin D Receptor and Runx2 in Vitamin D-Induced Vascular Calcification. PLoS ONE 2013, 8, e83584. [Google Scholar] [CrossRef] [PubMed]
- An, S.M.; Koh, J.S.; Boo, Y.C. Inhibition of melanogenesis by tyrosinase siRNA in human melanocytes. BMB Rep. 2009, 42, 178–183. [Google Scholar] [CrossRef] [PubMed]









| Code | CAS Number | Drug | Code | CAS Number | Drug |
|---|---|---|---|---|---|
| A2 | 21293-29-8 | (+)-Abscisic Acid | H4 | 174664-65-4 | CBHA |
| A3 | 3544-24-9 | 3-amino Benzamide | H5 | 251456-60-7 | M 344 |
| A4 | 929016-96-6 | SB939 | H6 | 151720-43-3 | Oxamflatin |
| A5 | 950762-95-5 | PCI 34051 | H7 | 1105698-15-4 | Salermide |
| A6 | 1219807-87-0 | 4-iodo-SAHA | H8 | 299953-00-7 | Mirin |
| A7 | 410536-97-9 | Sirtinol | H9 | 937039-45-7 | Pimelic Diphenylamide 106 |
| A8 | 328968-36-1 | C646 | H10 | 940943-37-3 | KD 5170 |
| A9 | 1239262-52-2 | Tubastatin A (trifluoroacetate salt) | H11 | 404950-80-7 | Panobinostat |
| A10 | 78824-30-3 | Garcinol | I2 | 209783-80-2 | MS-275 |
| A11 | 476-66-4 | Ellagic Acid | I3 | 926908-04-5 | HNHA |
| B2 | 287383-59-9 | Scriptaid | I4 | 48208-26-0 | RG-108 |
| B4 | 183506-66-3 | Apicidin | I5 | 10302-78-0 | 2′,3′,5′-triacetyl-5-Azacytidine |
| B5 | 83209-65-8 | HC Toxin | I6 | 979-92-0 | S-Adenosyl homocysteine |
| B6 | 1238673-32-9 | UNC0321 (trifluoroacetate salt) | I7 | 1197196-48-7 | UNC0224 |
| B7 | 72877-50-0 | (−)-Neplanocin A | I8 | 743420-02-2 | Chidamide |
| B8 | 1373232-26-8 | Cl-Amidine (hydrochloride) | I9 | 537049-40-4 | Tubacin |
| B9 | 877617-46-4 | F-Amidine (trifluoroacetate salt) | I10 | 102052-95-9 | 3-Deazaneplanocin A |
| B10 | 1256375-38-8 | JGB1741 | I11 | 58944-73-3 | Sinefungin |
| B11 | 1260635-77-5 | coumarin-SAHA | J2 | 382180-17-8 | Pyroxamide |
| C2 | 1260907-17-2 | I-BET762 | J3 | 5262-39-5 | N-Oxalylglycine |
| C3 | 1255580-76-7 | UNC0638 | J4 | 890190-22-4 | WDR5-0103 |
| C4 | 880487-62-7 | Phthalazinone pyrazole | J5 | 1396772-26-1 | EPZ005687 |
| C5 | 961-29-5 | Isoliquiritigenin | J6 | 1561178-17-3 | SGC0946 |
| C6 | 1207113-88-9 | CCG-100602 | J7 | 1415800-43-9 | UNC1215 |
| C7 | 1243583-88-1 | CAY10669 | J8 | 420831-40-9 | AK-7 |
| C8 | 3690-10-6 | Zebularine | J9 | 1346704-33-3 | GSK343 |
| C9 | 528-53-0 | Delphinidin (chloride) | J 10 | 1619994-69-2 | Bromosporine |
| C10 | 732302-99-7 | ITF 2357 | J 11 | 1619994-68-1 | GSK2801 |
| C11 | 1320288-19-4 | UNC0631 | K2 | 14513-15-6 | SIRT1/2 Inhibitor IV |
| D2 | 1320288-17-2 | UNC0646 | K3 | 2147701-33-3 | I-CBP112 (hydrochloride) |
| D3 | 1310877-95-2 | Methylstat (hydrate) | K4 | 1613695-14-9 | SGC-CBP30 |
| D4 | 120964-45-6 | 3-Deazaneplanocin A (hydrochloride) | K5 | 1481677-78-4 | UNC0642 |
| D5 | 129-46-4 | Suramin (sodium salt) | K6 | 1431612-23-5 | UNC1999 |
| D6 | 98-92-0 | Nicotinamide | K7 | 1627607-87-7 | (R)-PFI-2 (hydrochloride) |
| D7 | 207671-42-9 | 2,4-Pyridinedicarboxylic Acid | K8 | 1429651-50-2 | HPOB |
| D8 | 1403764-72-6 | PFI-1 | K9 | 96017-59-3 | 2-hexyl-4-Pentynoic Acid |
| D9 | 320-67-2 | 5-Azacytidine | K10 | 1819363-80-8 | PFI-3 |
| D10 | 1020149-73-8 | SGI-1027 | K11 | 199596-05-9 | JIB-04 |
| D11 | 2353-33-5 | Decitabine | L2 | 1477949-42-0 | CAY10683 |
| E2 | 1300031-49-5 | I-BET151 | L3 | 1346574-57-9 | GSK126 |
| E3 | 1268524-70-4 | (+)-JQ1 | L4 | 1446144-04-2 | CPI-203 |
| E4 | 1268524-71-5 | (−)-JQ1 | L5 | 154-42-7 | 6-Thioguanine |
| E5 | 160003-66-7 | BSI-201 | L6 | 1252003-15-8 | Tubastatin A |
| E6 | 86-55-5 | 1-Naphthoic Acid | L7 | 1968-05-4 | 3,3′-Diindolylmethane |
| E7 | 1716-12-7 | Sodium 4-Phenylbutyrate | L8 | 202590-98-5 | OTX015 |
| E8 | 459868-92-9 | Rucaparib (phosphate) | L9 | 2140-61-6 | 5-Methylcytidine |
| E9 | 5852-78-8 | IOX1 | L10 | 304896-21-7 | AGK7 |
| E10 | 1271738-62-5 | MI-2 (hydrochloride) | L11 | 838-07-3 | 5-Methyl-2′-deoxycytidine |
| E11 | 1934302-23-4 | MI-nc (hydrochloride) | M2 | 1380288-87-8 | EPZ5676 |
| F2 | 95058-81-4 | Gemcitabine | M3 | 852475-26-4 | MC 1568 |
| F3 | 192441-08-0 | Lomeguatrib | M4 | 40951-21-1 | α-Hydroxyglutaric Acid (sodium salt) |
| F4 | 1216744-19-2 | GSK4112 | M5 | 52248-03-0 | S-(5′-Adenosyl)-L-methionine (tosylate) |
| F5 | 876150-14-0 | Octyl-α-ketoglutarate | M6 | 1044870-39-4 | RVX-208 |
| F6 | 1596-84-5 | Daminozide | M7 | 1012054-59-9 | CUDC-101 |
| F7 | 1797832-71-3 | GSK-J1 (sodium salt) | M8 | 404951-53-7 | LAQ824 |
| F8 | 2108665-15-0 | GSK-J2 (sodium salt) | M9 | 300816-11-9 | Nullscript |
| F9 | 1797983-09-5 | GSK-J4 (hydrochloride) | M10 | 1431368-48-7 | GSK-LSD1 (hydrochloride) |
| F10 | 112522-64-2 | CI-994 | M11 | 1357389-11-7 | RGFP966 |
| F11 | 2108899-91-6 | CPTH2 (hydrochloride) | N2 | 1440209-96-0 | BRD73954 |
| G2 | 33419-42-0 | Etoposide | N3 | 501-36-0 | trans-Resveratrol |
| G3 | 111358-88-4 | Lestaurtinib | N4 | 89464-63-1 | DMOG |
| G4 | 778649-18-6 | Butyrolactone 3 | N5 | 58880-19-6 | Trichostatin A |
| G5 | 1069-66-5 | Valproic Acid (sodium salt) | N6 | 193551-00-7 | CAY10398 |
| G6 | 380315-80-0 | Tenovin-1 | N7 | 1418131-46-0 | RSC-133 |
| G7 | 1011557-82-6 | Tenovin-6 | N8 | 537034-17-6 | BML-210 |
| G8 | 156-54-7 | Sodium Butyrate | N9 | 10083-24-6 | Piceatannol |
| G9 | 1808255-64-2 | BIX01294 (hydrochloride hydrate) | N10 | 839699-72-8 | CAY10591 |
| G10 | 16611-84-0 | Anacardic Acid | N11 | 848193-68-0 | EX-527 |
| G11 | 304896-28-4 | AGK2 | O2 | 149647-78-9 | SAHA |
| H2 | 1045792-66-2 | CAY10603 | O3 | 1986-47-6 | 2-PCPA (hydrochloride) |
| H3 | 5690-03-9 | Splitomicin |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, H.; Hwang, Y.J.; Ha, J.W.; Boo, Y.C. Screening of an Epigenetic Drug Library Identifies 4-((hydroxyamino)carbonyl)-N-(2-hydroxyethyl)-N-Phenyl-Benzeneacetamide that Reduces Melanin Synthesis by Inhibiting Tyrosinase Activity Independently of Epigenetic Mechanisms. Int. J. Mol. Sci. 2020, 21, 4589. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134589
Song H, Hwang YJ, Ha JW, Boo YC. Screening of an Epigenetic Drug Library Identifies 4-((hydroxyamino)carbonyl)-N-(2-hydroxyethyl)-N-Phenyl-Benzeneacetamide that Reduces Melanin Synthesis by Inhibiting Tyrosinase Activity Independently of Epigenetic Mechanisms. International Journal of Molecular Sciences. 2020; 21(13):4589. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134589
Chicago/Turabian StyleSong, Hyerim, Yun Jeong Hwang, Jae Won Ha, and Yong Chool Boo. 2020. "Screening of an Epigenetic Drug Library Identifies 4-((hydroxyamino)carbonyl)-N-(2-hydroxyethyl)-N-Phenyl-Benzeneacetamide that Reduces Melanin Synthesis by Inhibiting Tyrosinase Activity Independently of Epigenetic Mechanisms" International Journal of Molecular Sciences 21, no. 13: 4589. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134589