Chronic Sildenafil Treatment Improves Vasomotor Function in a Mouse Model of Accelerated Aging

 , and
, and 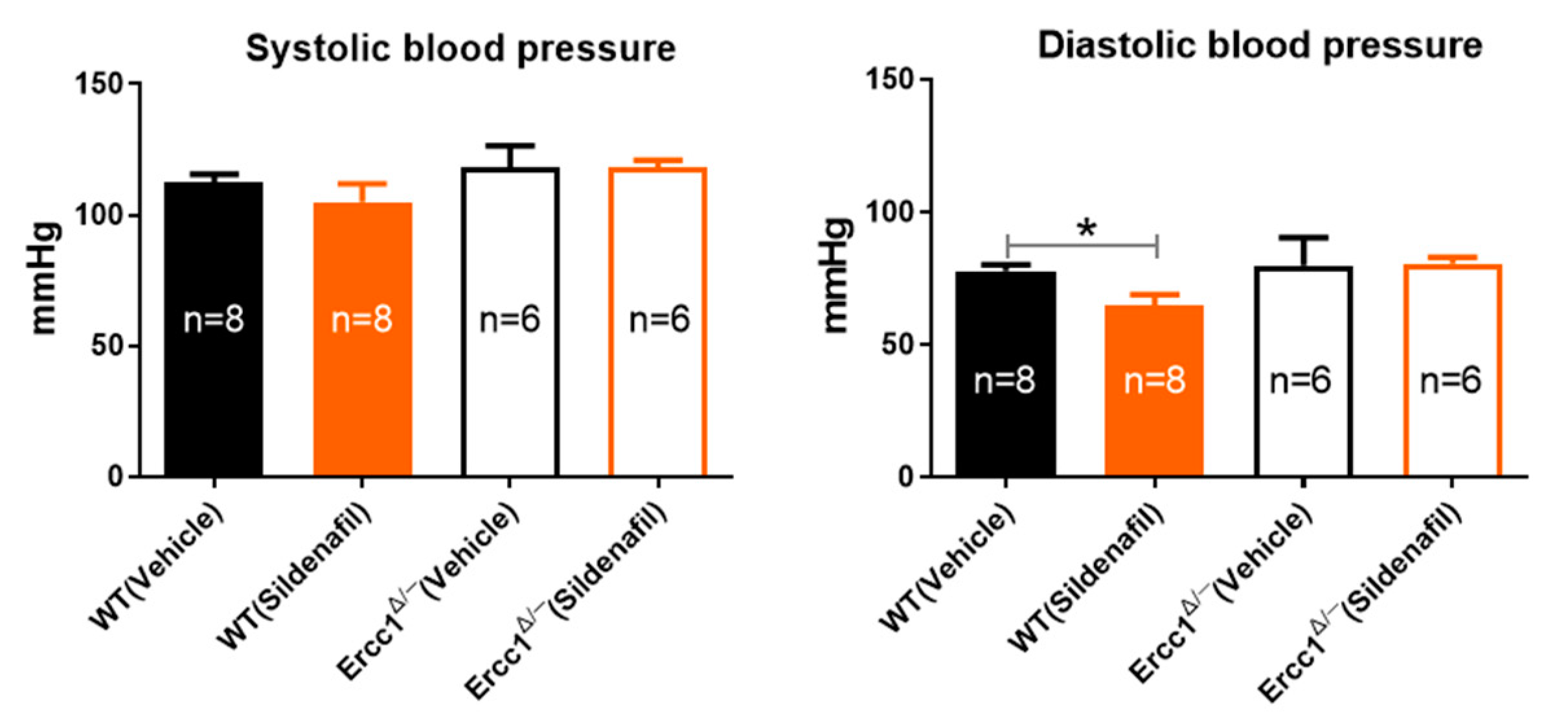{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Unchanged Systolic and Diastolic Blood Pressure in Accelerated-Aging Model
2.2. Reduced Cutaneous Vasodilation in Ercc1Δ/− Mice
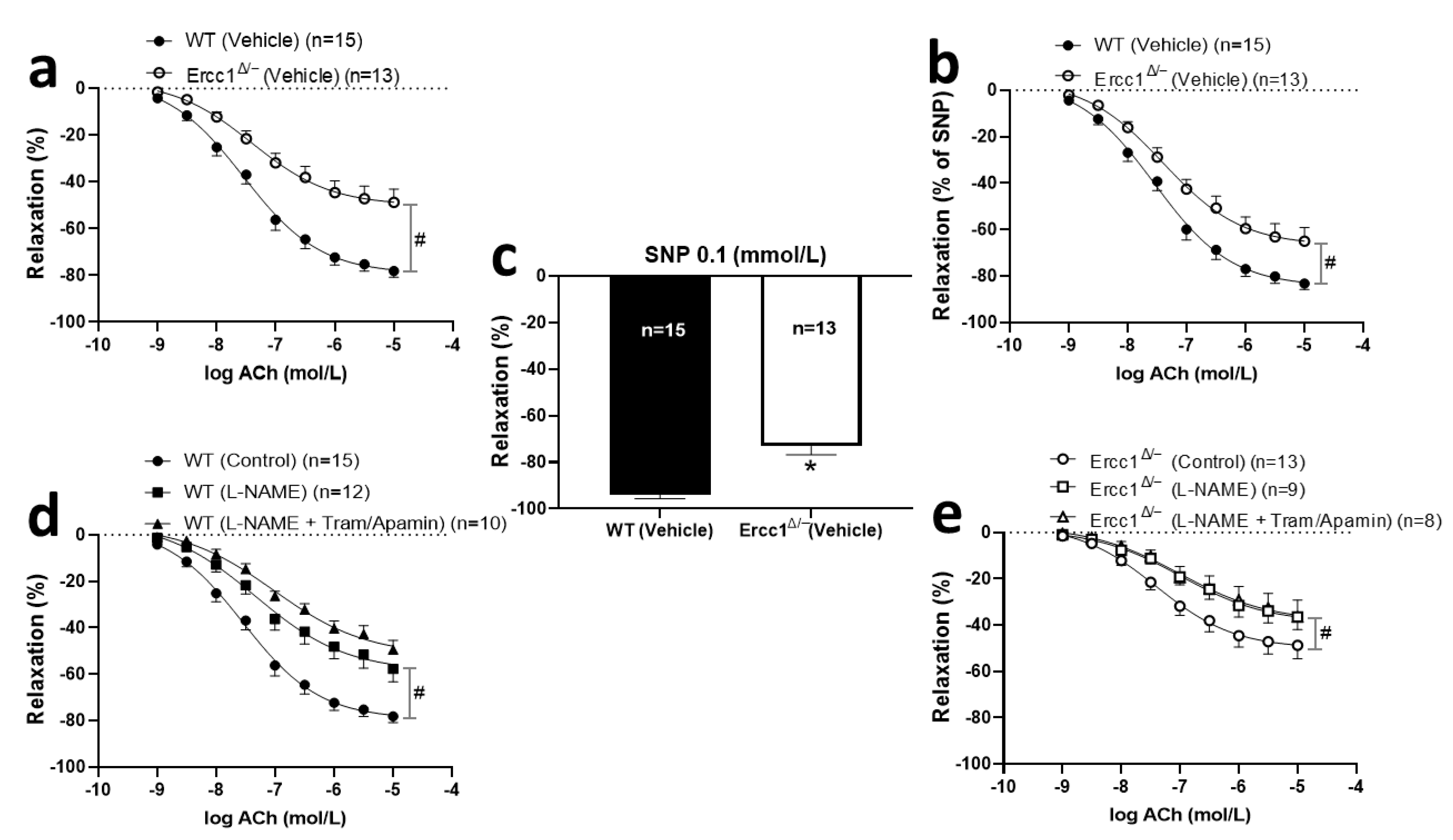
2.3. Diminished Endothelium-Dependent and -Independent Response in Ercc1Δ/− Mice
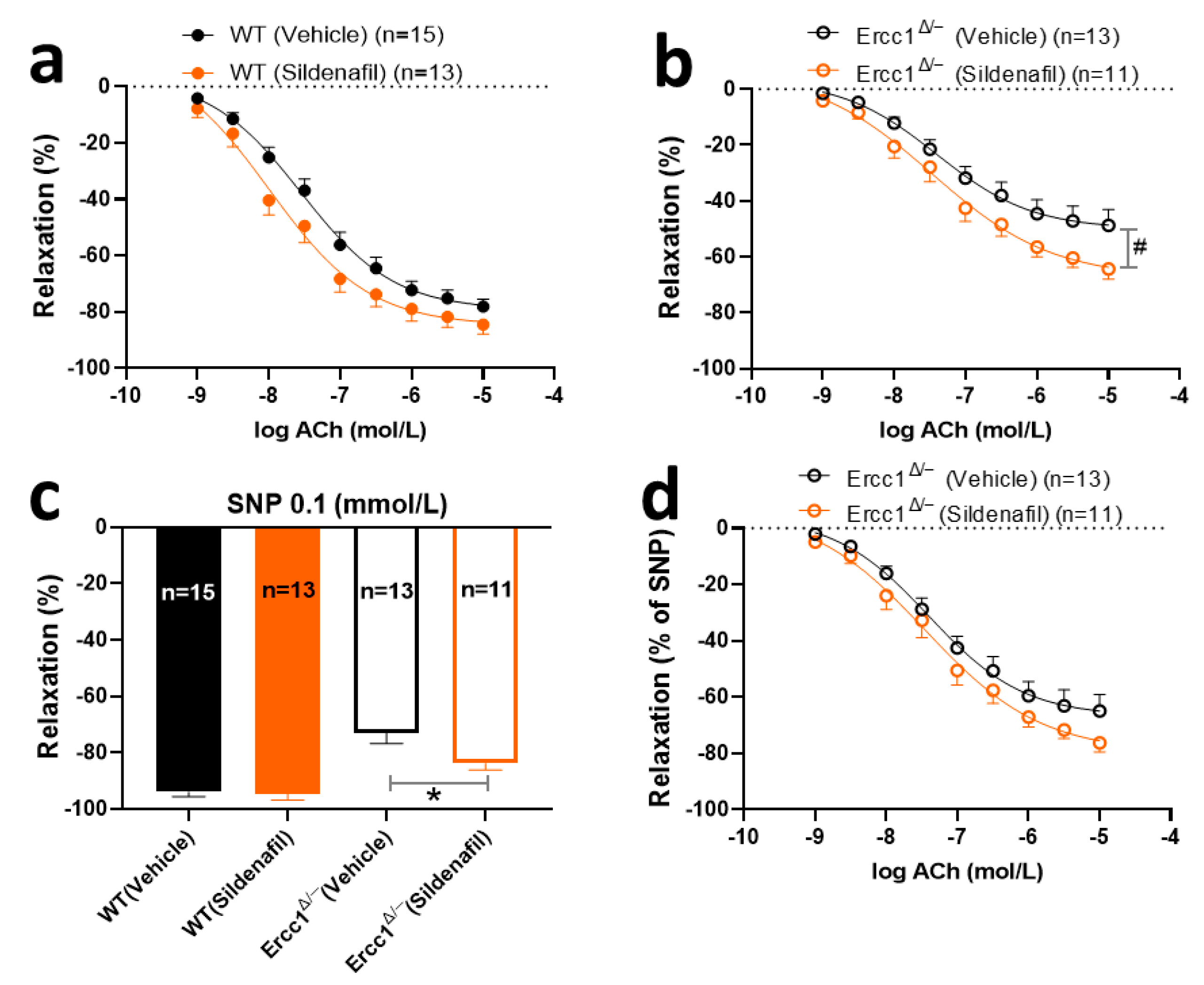
2.4. The Effects of Chronic Treatment with Sildenafil on Endothelium-Dependent and -Independent Response
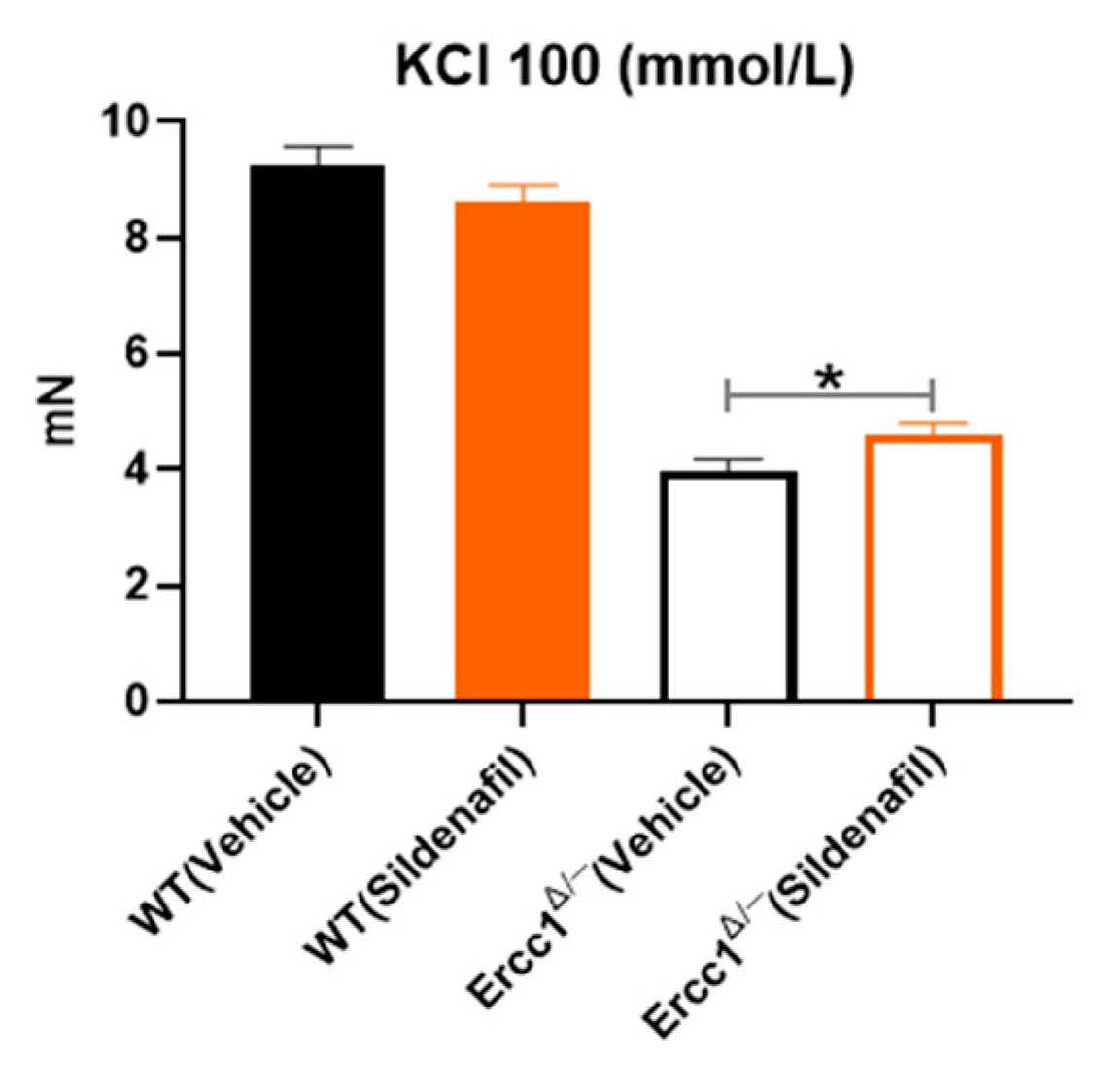
2.5. Effect of Sildenafil on Vasoconstriction Mediated by KCl 100
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Study Design
4.3. Blood Pressure and Vasodilator Function (In Vivo)
4.4. Sacrifice and Wire Myography
4.5. Data Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| cGMP | Cyclic guanosine monophosphate |
| sGC | Soluble guanylyl cyclase |
| eNOS | Endothelial nitric oxide synthase |
| PDE | Phosphodiesterase |
| NO | Nitric oxide |
| cAMP | Cyclic adenosine monophosphate |
| VSMC | Vascular smooth muscle cell |
| ERCC | Excision repair cross-complementing |
| EDH | Endothelium-dependent hyperpolarization |
| EDHF | Endothelium-derived hyperpolarizing factor |
| SNP | Sodium nitroprusside |
| ACh | Acetylcholine |
| L-NAME | L-NG-Nitro arginine methyl ester |
| LMNA | Lamin AC |
References
- North, B.J.; Sinclair, D.A. The intersection between aging and cardiovascular disease. Circ. Res. 2012, 110, 1097–1108. [Google Scholar] [CrossRef] [PubMed]
- Vanhoutte, P.M.; Shimokawa, H.; Feletou, M.; Tang, E.H.C. Endothelial dysfunction and vascular disease–a 30th anniversary update. Acta Physiol. 2017, 219, 22–96. [Google Scholar] [CrossRef] [PubMed]
- Widmer, R.J.; Lerman, A. Endothelial dysfunction and cardiovascular disease. Glob. Cardiol. Sci. Pract. 2014, 3, 291–308. [Google Scholar] [CrossRef] [PubMed]
- Golshiri, K.; Ataei Ataabadi, E.; Portilla-Fernandez, E.; Danser, J.; Roks, A. The importance of the nitric oxide-cGMP pathway in age-related cardiovascular disease: Focus on phosphodiesterase-1 and soluble guanylate cyclase. Basic Clin. Pharm. Toxicol 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friebe, A.; Sandner, P.; Schmidtko, A. cGMP: A unique 2nd messenger molecule–recent developments in cGMP research and development. Naunyn-Schmiedeberg’s Arch. Pharm. 2020, 393, 287–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durik, M.; Kavousi, M.; van der Pluijm, I.; Isaacs, A.; Cheng, C.; Verdonk, K.; Loot, A.E.; Oeseburg, H.; Bhaggoe, U.M.; Leijten, F.; et al. Nucleotide excision DNA repair is associated with age-related vascular dysfunction. Circulation 2012, 126, 468–478. [Google Scholar] [CrossRef]
- Bautista Nino, P.K.; Durik, M.; Danser, A.H.; de Vries, R.; Musterd-Bhaggoe, U.M.; Meima, M.E.; Kavousi, M.; Ghanbari, M.; Hoeijmakers, J.H.; O’Donnell, C.J.; et al. Phosphodiesterase 1 regulation is a key mechanism in vascular aging. Clin. Sci. 2015, 129, 1061–1075. [Google Scholar] [CrossRef]
- Donato, A.J.; Machin, D.R.; Lesniewski, L.A. Mechanisms of Dysfunction in the Aging Vasculature and Role in Age-Related Disease. Circ. Res. 2018, 123, 825–848. [Google Scholar] [CrossRef]
- Lugnier, C. Cyclic nucleotide phosphodiesterase (PDE) superfamily: A new target for the development of specific therapeutic agents. Pharm. Ther. 2006, 109, 366–398. [Google Scholar] [CrossRef]
- Andersson, K.E. PDE5 inhibitors-pharmacology and clinical applications 20 years after sildenafil discovery. Br. J. Pharm. 2018, 175, 2554–2565. [Google Scholar] [CrossRef] [Green Version]
- Goud, A.P.; Goud, P.T.; Diamond, M.P.; Gonik, B.; Abu-Soud, H.M. Activation of the cGMP signaling pathway is essential in delaying oocyte aging in diabetes mellitus. Biochemistry 2006, 45, 11366–11378. [Google Scholar] [CrossRef] [PubMed]
- Kukreja, R.C.; Salloum, F.N.; Das, A. Cyclic guanosine monophosphate signaling and phosphodiesterase-5 inhibitors in cardioprotection. J. Am. Coll. Cardiol. 2012, 9, 1921–1927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoeijmakers, J.H. DNA damage, aging, and cancer. N. Engl. J. Med. 2009, 361, 1475–1485. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Campo, L.; Sánchez-López, A.; Salaices, M.; von Kleeck, R.A.; Expósito, E.; González-Gómez, C.; Cussó, L.; Guzmán-Martínez, G.; Ruiz-Cabello, J.; Desco, M.; et al. Vascular smooth muscle cell-specific progerin expression in a mouse model of Hutchinson-Gilford progeria syndrome promotes arterial stiffness: Therapeutic effect of dietary nitrite. Aging Cell 2019, 18, e12936. [Google Scholar] [CrossRef] [Green Version]
- Tonoyama, Y.; Shinya, M.; Toyoda, A.; Kitano, T.; Oga, A.; Nishimaki, T.; Katsumura, T.; Oota, H.; Wan, M.T.; Yip, B.W.P.; et al. Abnormal nuclear morphology is independent of longevity in a zmpste24-deficient fish model of Hutchinson-Gilford progeria syndrome (HGPS). Comp. Biochem. Physiol. Part C Toxicol. Pharm. 2018, 209, 54–62. [Google Scholar] [CrossRef]
- Bautista-Nino, P.K.; Portilla-Fernandez, E.; Rubio-Beltrán, E.; van der Linden, J.J.; de Vries, R.; van Veghel, R.; de Boer, M.; Durik, M.; Ridwan, Y.; Brandt, R.; et al. Local endothelial DNA repair deficiency causes aging-resembling endothelial-specific dysfunction. Clin. Sci. 2020, 134, 727–746. [Google Scholar] [CrossRef] [Green Version]
- Breitenstein, S.; Roessig, L.; Sandner, P.; Lewis, K.S. Novel sGC Stimulators and sGC Activators for the Treatment of Heart Failure. Handb. Exp. Pharm. 2017, 243, 225–247. [Google Scholar]
- Vermeij, W.P.; Dollé, M.E.T.; Reiling, E.; Jaarsma, D.; Payan-Gomez, C.; Bombardieri, C.R.; Wu, H.; Roks, A.J.; Botter, S.M.; van der Eerden, B.C.; et al. Restricted diet delays accelerated ageing and genomic stress in DNA-repair-deficient mice. Nature 2016, 537, 427–431. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Van Thiel, B.S.; Bautista-Niño, P.K.; Reiling, E.; Durik, M.; Leijten, F.P.J.; Ridwan, Y.; Brandt, R.M.C.; Van Steeg, H.; Dollé, M.E.T.; et al. Dietary restriction but not angiotensin II type 1 receptor blockade improves DNA damage-related vasodilator dysfunction in rapidly aging Ercc1Delta/-mice. Clin. Sci. 2017, 131, 1941–1953. [Google Scholar] [CrossRef] [Green Version]
- Maassen VanDenBrink, A.; Bax, W.A.; Ramrattan, N.N.; Ferrari, M.D.; Saxena, P.R. Human isolated coronary artery contraction to sumatriptan: A post hoc analysis. Cephalalgia 1999, 19, 651–654. [Google Scholar] [CrossRef] [PubMed]
- Del Campo, L.; Sánchez-López, A.; González-Gómez, C.; Andrés-Manzano, M.J.; Dorado, B.; Andrés, V. Vascular Smooth Muscle Cell-Specific Progerin Expression Provokes Contractile Impairment in a Mouse Model of Hutchinson-Gilford Progeria Syndrome that Is Ameliorated by Nitrite Treatment. Cells 2020, 9, 656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujii, N.; Meade, R.D.; Alexander, L.M.; Akbari, P.; Foudil-Bey, I.; Louie, J.C.; Boulay, P.; Kenny, G.P.; Inoue, Y. iNOS-dependent sweating and eNOS-dependent cutaneous vasodilation are evident in younger adults, but are diminished in older adults exercising in the heat. J. Appl. Physiol. 2016, 120, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Oue, A.; Asashima, C.; Oizumi, R.; Ichinose-Kuwahara, T.; Kondo, N.; Inoue, Y. Age-related attenuation of conduit artery blood flow response to passive heating differs between the arm and leg. Eur. J. Appl. Physiol. 2018, 118, 2307–2318. [Google Scholar] [CrossRef] [PubMed]
- Vermeij, W.P.; Hoeijmakers, J.H.; Pothof, J. Genome Integrity in Aging: Human Syndromes, Mouse Models, and Therapeutic Options. Annu. Rev. Pharm. Toxicol. 2016, 56, 427–445. [Google Scholar] [CrossRef]
- McMurray, J.G.; Feldman, R.A.; Auerbach, S.M.; DeRiesthal, H.; Wilson, N.; On behalf of the Multicenter Study Group. Long-term safety and effectiveness of sildenafil citrate in men with erectile dysfunction. Clin. Risk Manag. 2007, 3, 975–981. [Google Scholar]
- Selfridge, J.; Hsia, K.T.; Redhead, N.J.; Melton, D.W. Correction of liver dysfunction in DNA repair-deficient mice with an ERCC1 transgene. Nucleic. Acids Res. 2001, 29, 4541–4550. [Google Scholar] [CrossRef] [Green Version]
- Dollé, M.E.T.; Kuiper, R.V.; Roodbergen, M.; Robinson, J.; De Vlugt, S.; Wijnhoven, S.W.; Beems, R.B.; De La Fonteyne, L.; De With, P.; Van Der Pluijm, I.; et al. Broad segmental progeroid changes in short-lived Ercc1-/Δ7 mice. Pathobiol. Aging Age-Relat. Dis. 2011, 1, 7219. [Google Scholar]
- Ishikura, F.; Beppu, S.; Hamada, T.; Khandheria, B.K.; Seward, J.B.; Nehra, A. Effects of Sildenafil Citrate (Viagra) Combined With Nitrate on the Heart. Circulation 2000, 102, 2516–2521. [Google Scholar] [CrossRef] [Green Version]
- Ramani, G.V.; Park, M.H. Update on the clinical utility of sildenafil in the treatment of pulmonary arterial hypertension. Drug Des. Devel. 2010, 4, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Weeda, G.; Donker, I.; De Wit, J.; Morreau, H.; Janssens, R.; Vissers, C.J.; Nigg, A.; Van Steeg, H.; Bootsma, D.; Hoeijmakers, J.H. Disruption of mouse ERCC1 results in a novel repair syndrome with growth failure, nuclear abnormalities and senescence. Curr. Biol. 1997, 7, 427–439. [Google Scholar] [CrossRef] [Green Version]
- Au, A.; Williams, C.L.; Pointer, M.A.; Awumey, E.M. Ups and downs of Viagra: Revisiting ototoxicity in the mouse model. PLoS ONE 2013, 8, e79226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bridges, L.E.; Williams, C.L.; Pointer, M.A.; Awumey, E.M. Mesenteric artery contraction and relaxation studies using automated wire myography. J. Vis. Exp. 2011, 55, e3119. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Golshiri, K.; Ataei Ataabadi, E.; Brandt, R.; van der Pluijm, I.; de Vries, R.; Danser, A.H.J.; Roks, A. Chronic Sildenafil Treatment Improves Vasomotor Function in a Mouse Model of Accelerated Aging. Int. J. Mol. Sci. 2020, 21, 4667. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134667
Golshiri K, Ataei Ataabadi E, Brandt R, van der Pluijm I, de Vries R, Danser AHJ, Roks A. Chronic Sildenafil Treatment Improves Vasomotor Function in a Mouse Model of Accelerated Aging. International Journal of Molecular Sciences. 2020; 21(13):4667. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134667
Chicago/Turabian StyleGolshiri, Keivan, Ehsan Ataei Ataabadi, Renata Brandt, Ingrid van der Pluijm, René de Vries, A. H. Jan Danser, and Anton Roks. 2020. "Chronic Sildenafil Treatment Improves Vasomotor Function in a Mouse Model of Accelerated Aging" International Journal of Molecular Sciences 21, no. 13: 4667. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134667