Ketamine and Ro 25-6981 Reverse Behavioral Abnormalities in Rats Subjected to Dietary Zinc Restriction


Abstract
:1. Introduction
2. Results
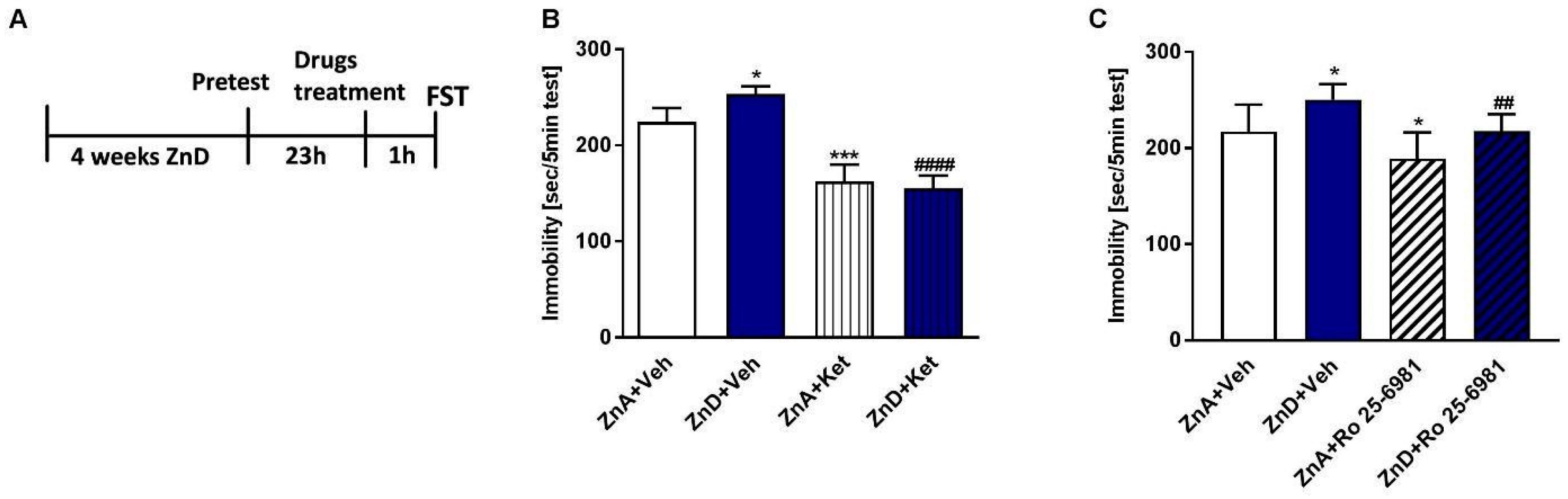
2.1. Effects of Ro and Ketamine on Immobility Time and Locomotor Activity of ZnD Rats
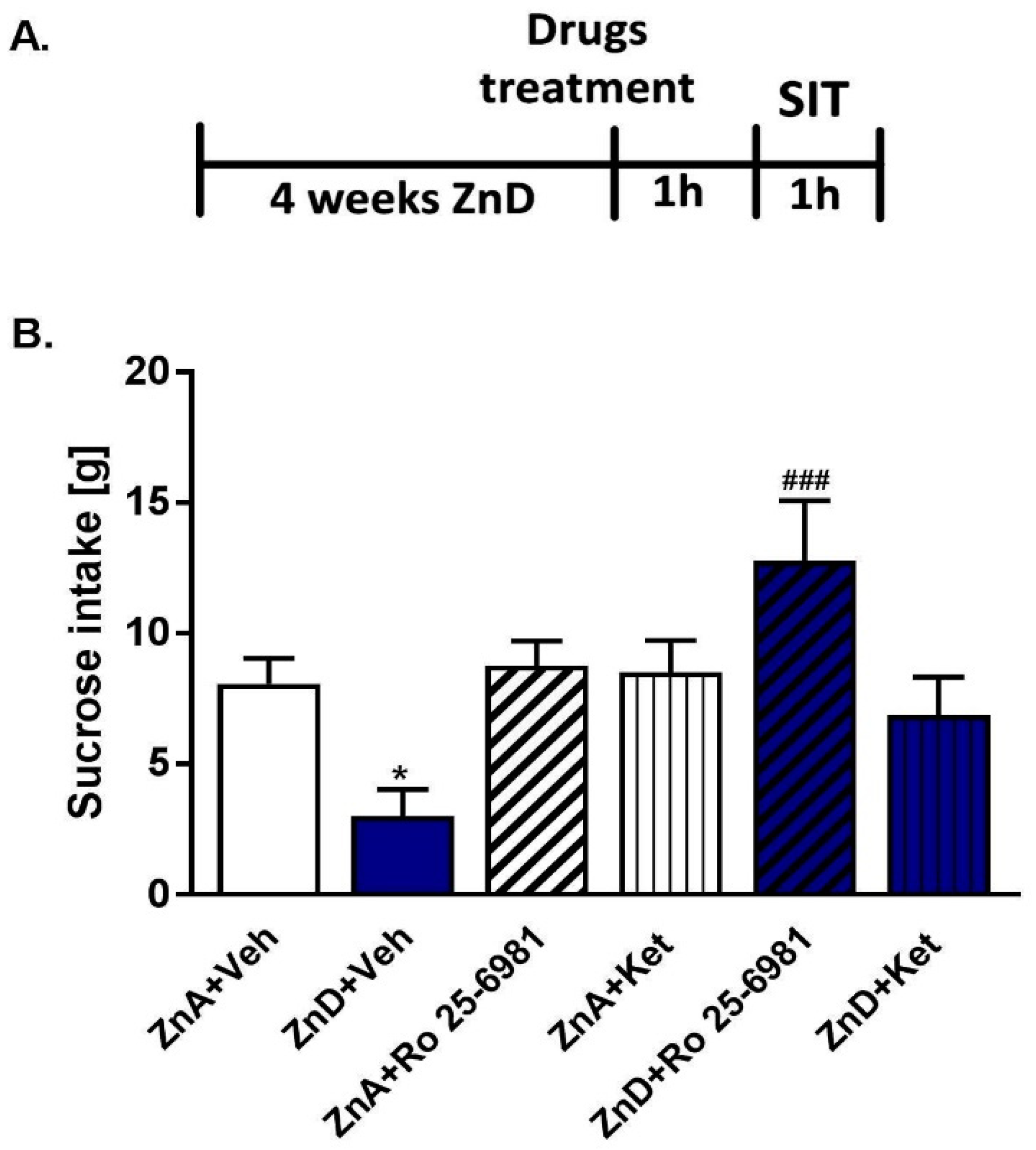
2.2. Effects of Ro and Ketamine on Sucrose Intake in Rats Subjected to Dietary Zn Restriction
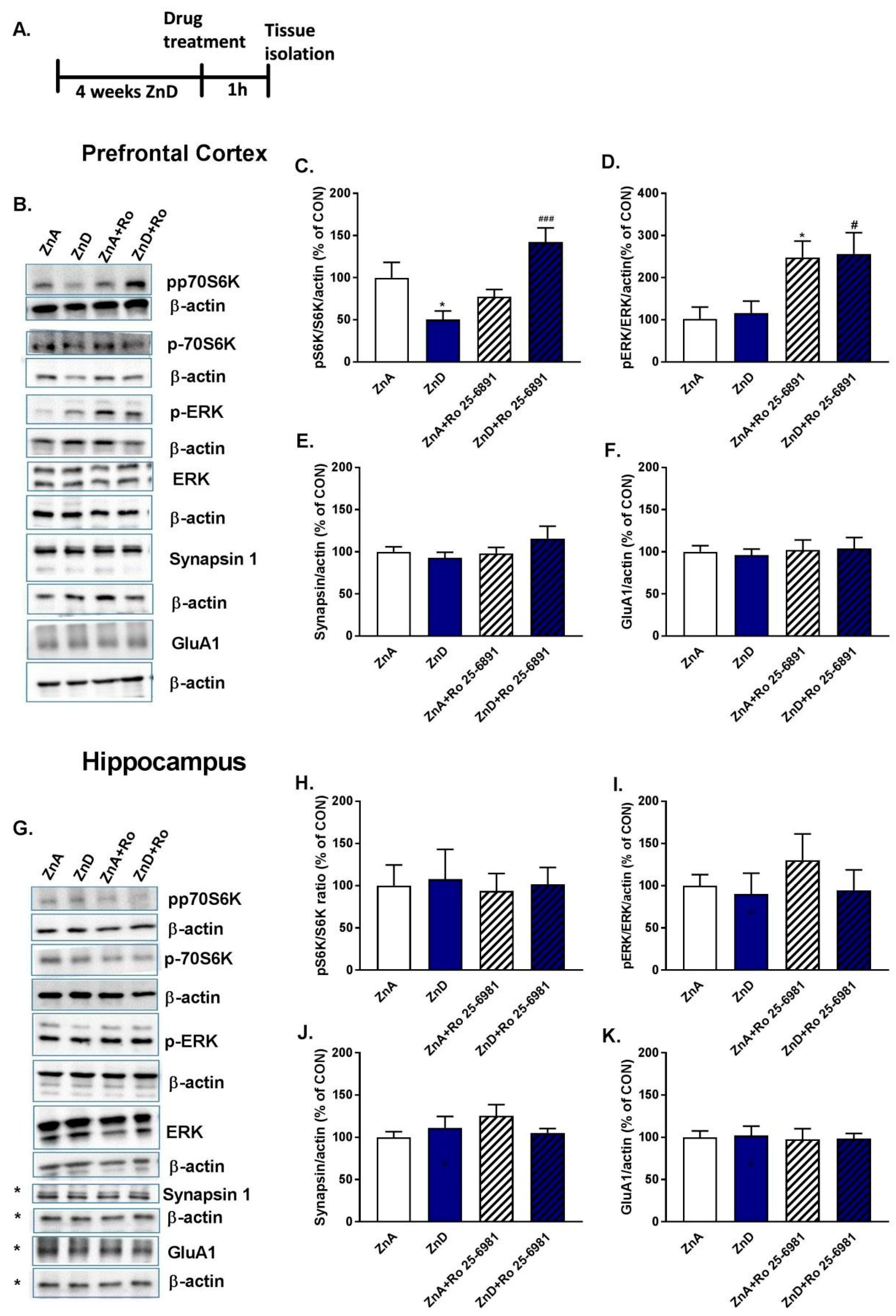
2.3. Effects of Ro and ZnD on the Levels of Synaptic Proteins and ERK/p-70S6K Kinases
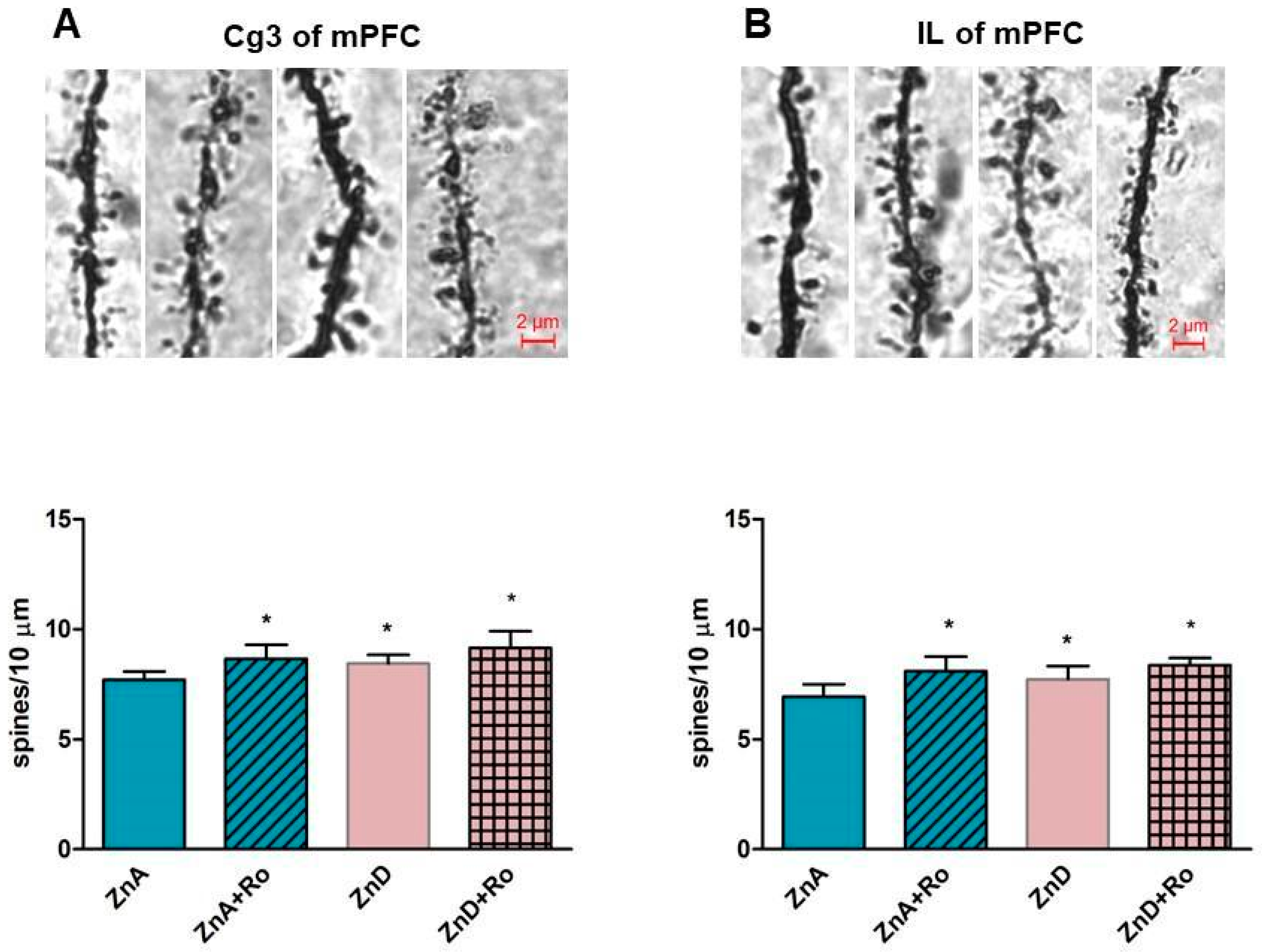
2.4. Effects of Ro and ZnD on the Density of Dendritic Spines in the Medial Prefrontal Cortex (mPFC)
3. Discussion
4. Material and Methods
4.1. Animals
4.2. Drug Administration
4.3. Forced Swim Test (FST)
4.4. Locomotor Activity
4.5. Zinc Restriction
4.6. Sucrose Intake Test
4.7. Synaptosome Preparation and Western Blotting
4.8. Morphological Analysis of Dendritic Spines
4.9. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Busfield, J. Challenging claims that mental illness has been increasing and mental well-being declining. Soc. Sci. Med. 2012, 75, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Hidaka, B.H. Depression as a disease of modernity: Explanations for increasing prevalence. J. Affect. Disord. 2012, 140, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Wittchen, H.U.; Jacobi, F.; Rehm, J.; Gustavsson, A.; Svensson, M.; Jonsson, B.; Olesen, J.; Allgulander, C.; Alonso, J.; Faravelli, C.; et al. The size and burden of mental disorders and other disorders of the brain in Europe 2010. Eur. Neuropsychopharmacol. 2011, 21, 655–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenblat, J.D.; McIntyre, R.S.; Alves, G.S.; Fountoulakis, K.N.; Carvalho, A.F. Beyond Monoamines-Novel Targets for Treatment-Resistant Depression: A Comprehensive Review. Curr. Neuropharmacol. 2015, 13, 636–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szewczyk, B.; Szopa, A.; Serefko, A.; Poleszak, E.; Nowak, G. The role of magnesium and zinc in depression: Similarities and differences. Magnes. Res. 2018, 31, 78–89. [Google Scholar] [CrossRef]
- Vashum, K.P.; McEvoy, M.; Milton, A.H.; McElduff, P.; Hure, A.; Byles, J.; Attia, J. Dietary zinc is associated with a lower incidence of depression: Findings from two Australian cohorts. J. Affect. Disord. 2014, 166, 249–257. [Google Scholar] [CrossRef]
- Jacka, F.N.; Maes, M.; Pasco, J.A.; Williams, L.J.; Berk, M. Nutrient intakes and the common mental disorders in women. J. Affect. Disord. 2012, 141, 79–85. [Google Scholar] [CrossRef]
- Hansen, C.R., Jr.; Malecha, M.; Mackenzie, T.B.; Kroll, J. Copper and zinc deficiencies in association with depression and neurological findings. Biol. Psychiatry 1983, 18, 395–401. [Google Scholar]
- Maes, M.; Bosmans, E.; De Jongh, R.; Kenis, G.; Vandoolaeghe, E.; Neels, H. Increased serum IL-6 and IL-1 receptor antagonist concentrations in major depression and treatment resistant depression. Cytokine 1997, 9, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Lehto, S.M.; Ruusunen, A.; Tolmunen, T.; Voutilainen, S.; Tuomainen, T.P.; Kauhanen, J. Dietary zinc intake and the risk of depression in middle-aged men: A 20-year prospective follow-up study. J. Affect. Disord. 2013, 150, 682–685. [Google Scholar] [CrossRef]
- Marcellini, F.; Giuli, C.; Papa, R.; Gagliardi, C.; Dedoussis, G.; Herbein, G.; Fulop, T.; Monti, D.; Rink, L.; Jajte, J.; et al. Zinc status, psychological and nutritional assessment in old people recruited in five European countries: Zincage study. Biogerontology 2006, 7, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Markiewicz-Zukowska, R.; Gutowska, A.; Borawska, M.H. Serum zinc concentrations correlate with mental and physical status of nursing home residents. PLoS ONE 2015, 10, e0117257. [Google Scholar] [CrossRef]
- Siwek, M.; Szewczyk, B.; Dudek, D.; Styczen, K.; Sowa-Kucma, M.; Mlyniec, K.; Siwek, A.; Witkowski, L.; Pochwat, B.; Nowak, G. Zinc as a marker of affective disorders. Pharmacol. Rep. 2013, 65, 1512–1518. [Google Scholar] [CrossRef]
- McLoughlin, I.J.; Hodge, J.S. Zinc in depressive disorder. Acta Psychiatr. Scand. 1990, 82, 451–453. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; De Vos, N.; Demedts, P.; Wauters, A.; Neels, H. Lower serum zinc in major depression in relation to changes in serum acute phase proteins. J. Affect. Disord. 1999, 56, 189–194. [Google Scholar] [CrossRef]
- Siwek, M.; Dudek, D.; Schlegel-Zawadzka, M.; Morawska, A.; Piekoszewski, W.; Opoka, W.; Zieba, A.; Pilc, A.; Popik, P.; Nowak, G. Serum zinc level in depressed patients during zinc supplementation of imipramine treatment. J. Affect. Disord. 2010, 126, 447–452. [Google Scholar] [CrossRef]
- Mlyniec, K.; Davies, C.L.; Budziszewska, B.; Opoka, W.; Reczynski, W.; Sowa-Kucma, M.; Doboszewska, U.; Pilc, A.; Nowak, G. Time course of zinc deprivation-induced alterations of mice behavior in the forced swim test. Pharmacol. Rep. 2012, 64, 567–575. [Google Scholar] [CrossRef]
- Mlyniec, K.; Nowak, G. Zinc deficiency induces behavioral alterations in the tail suspension test in mice. Effect of antidepressants. Pharmacol. Rep. 2012, 64, 249–255. [Google Scholar] [CrossRef]
- Whittle, N.; Lubec, G.; Singewald, N. Zinc deficiency induces enhanced depression-like behaviour and altered limbic activation reversed by antidepressant treatment in mice. Amino Acids 2009, 36, 147–158. [Google Scholar] [CrossRef]
- Doboszewska, U.; Sowa-Kucma, M.; Mlyniec, K.; Pochwat, B.; Holuj, M.; Ostachowicz, B.; Pilc, A.; Nowak, G.; Szewczyk, B. Zinc deficiency in rats is associated with up-regulation of hippocampal NMDA receptor. Prog. Neuropsychopharmacol. Biol. Psychiatry 2015, 56, 254–263. [Google Scholar] [CrossRef]
- Doboszewska, U.; Szewczyk, B.; Sowa-Kucma, M.; Mlyniec, K.; Rafalo, A.; Ostachowicz, B.; Lankosz, M.; Nowak, G. Antidepressant activity of fluoxetine in the zinc deficiency model in rats involves the NMDA receptor complex. Behav. Brain Res. 2015, 287, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Tassabehji, N.M.; Corniola, R.S.; Alshingiti, A.; Levenson, C.W. Zinc deficiency induces depression-like symptoms in adult rats. Physiol. Behav. 2008, 95, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Yang, J.; Luo, A.; Hashimoto, K. Molecular and cellular mechanisms underlying the antidepressant effects of ketamine enantiomers and its metabolites. Transl. Psychiatry 2019, 9, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarate, C.A., Jr.; Niciu, M.J. Ketamine for depression: Evidence, challenges and promise. World Psychiatry 2015, 14, 348–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berman, R.M.; Cappiello, A.; Anand, A.; Oren, D.A.; Heninger, G.R.; Charney, D.S.; Krystal, J.H. Antidepressant effects of ketamine in depressed patients. Biol. Psychiatry 2000, 47, 351–354. [Google Scholar] [CrossRef]
- Zarate, C.A., Jr.; Singh, J.B.; Carlson, P.J.; Brutsche, N.E.; Ameli, R.; Luckenbaugh, D.A.; Charney, D.S.; Manji, H.K. A randomized trial of an N-methyl-D-aspartate antagonist in treatment-resistant major depression. Arch. Gen. Psychiatry 2006, 63, 856–864. [Google Scholar] [CrossRef]
- Daly, E.J.; Singh, J.B.; Fedgchin, M.; Cooper, K.; Lim, P.; Shelton, R.C.; Thase, M.E.; Winokur, A.; Van Nueten, L.; Manji, H.; et al. Efficacy and Safety of Intranasal Esketamine Adjunctive to Oral Antidepressant Therapy in Treatment-Resistant Depression: A Randomized Clinical Trial. JAMA Psychiatry 2018, 75, 139–148. [Google Scholar] [CrossRef]
- DiazGranados, N.; Ibrahim, L.A.; Brutsche, N.E.; Ameli, R.; Henter, I.D.; Luckenbaugh, D.A.; Machado-Vieira, R.; Zarate, C.A., Jr. Rapid resolution of suicidal ideation after a single infusion of an N-methyl-D-aspartate antagonist in patients with treatment-resistant major depressive disorder. J. Clin. Psychiatry 2010, 71, 1605–1611. [Google Scholar] [CrossRef] [Green Version]
- Canuso, C.M.; Singh, J.B.; Fedgchin, M.; Alphs, L.; Lane, R.; Lim, P.; Pinter, C.; Hough, D.; Sanacora, G.; Manji, H.; et al. Efficacy and Safety of Intranasal Esketamine for the Rapid Reduction of Symptoms of Depression and Suicidality in Patients at Imminent Risk for Suicide: Results of a Double-Blind, Randomized, Placebo-Controlled Study. Am. J. Psychiatry 2018, 175, 620–630. [Google Scholar] [CrossRef]
- De Berardis, D.; Fornaro, M.; Valchera, A.; Cavuto, M.; Perna, G.; Di Nicola, M.; Serafini, G.; Carano, A.; Pompili, M.; Vellante, F.; et al. Eradicating Suicide at Its Roots: Preclinical Bases and Clinical Evidence of the Efficacy of Ketamine in the Treatment of Suicidal Behaviors. Int. J. Mol. Sci. 2018, 19, 2888. [Google Scholar] [CrossRef] [Green Version]
- De Berardis, D.; Tomasetti, C.; Pompili, M.; Serafini, G.; Vellante, F.; Fornaro, M.; Valchera, A.; Perna, G.; Volpe, U.; Martinotti, G.; et al. An Update on Glutamatergic System in Suicidal Depression and on the Role of Esketamine. Curr. Top. Med. Chem. 2020, 20, 554–584. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Liu, R.J.; Dwyer, J.M.; Banasr, M.; Lee, B.; Son, H.; Li, X.Y.; Aghajanian, G.; Duman, R.S. Glutamate N-methyl-D-aspartate receptor antagonists rapidly reverse behavioral and synaptic deficits caused by chronic stress exposure. Biol. Psychiatry 2011, 69, 754–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, B.K.; Qin, J.; Nie, Y.; Chen, J.C. Sustained antidepressant action of the N-methyl-D-aspartate receptor antagonist MK-801 in a chronic unpredictable mild stress model. Exp. Ther. Med. 2018, 16, 5376–5383. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, M.G.; Ames, H.M.; Christensen, M.K.; Sorensen, J.C. Zincergic innervation of medial prefrontal cortex by basolateral projection neurons. Neuroreport 2007, 18, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Frederickson, C.J.; Danscher, G. Zinc-containing neurons in hippocampus and related CNS structures. Prog. Brain Res. 1990, 83, 71–84. [Google Scholar] [CrossRef]
- Maret, W. Zinc biochemistry: From a single zinc enzyme to a key element of life. Adv. Nutr. 2013, 4, 82–91. [Google Scholar] [CrossRef] [Green Version]
- Suh, S.W.; Chen, J.W.; Motamedi, M.; Bell, B.; Listiak, K.; Pons, N.F.; Danscher, G.; Frederickson, C.J. Evidence that synaptically-released zinc contributes to neuronal injury after traumatic brain injury. Brain Res. 2000, 852, 268–273. [Google Scholar] [CrossRef]
- Mlyniec, K.; Gawel, M.; Nowak, G. Study of antidepressant drugs in GPR39 (zinc receptor(-)/(-)) knockout mice, showing no effect of conventional antidepressants, but effectiveness of NMDA antagonists. Behav. Brain Res. 2015, 287, 135–138. [Google Scholar] [CrossRef]
- Li, N.; Lee, B.; Liu, R.J.; Banasr, M.; Dwyer, J.M.; Iwata, M.; Li, X.Y.; Aghajanian, G.; Duman, R.S. mTOR-dependent synapse formation underlies the rapid antidepressant effects of NMDA antagonists. Science 2010, 329, 959–964. [Google Scholar] [CrossRef] [Green Version]
- Pochwat, B.; Rafalo-Ulinska, A.; Domin, H.; Misztak, P.; Nowak, G.; Szewczyk, B. Involvement of extracellular signal-regulated kinase (ERK) in the short and long-lasting antidepressant-like activity of NMDA receptor antagonists (zinc and Ro 25-6981) in the forced swim test in rats. Neuropharmacology 2017, 125, 333–342. [Google Scholar] [CrossRef]
- Doboszewska, U.; Szewczyk, B.; Sowa-Kucma, M.; Noworyta-Sokolowska, K.; Misztak, P.; Golebiowska, J.; Mlyniec, K.; Ostachowicz, B.; Krosniak, M.; Wojtanowska-Krosniak, A.; et al. Alterations of Bio-elements, Oxidative, and Inflammatory Status in the Zinc Deficiency Model in Rats. Neurotox. Res. 2016, 29, 143–154. [Google Scholar] [CrossRef] [Green Version]
- Paoletti, P.; Vergnano, A.M.; Barbour, B.; Casado, M. Zinc at glutamatergic synapses. Neuroscience 2009, 158, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Pochwat, B.; Nowak, G.; Szewczyk, B. An update on NMDA antagonists in depression. Expert Rev. Neurother 2019, 19, 1055–1067. [Google Scholar] [CrossRef]
- Ghosal, S.; Duman, C.H.; Liu, R.J.; Wu, M.; Terwilliger, R.; Girgenti, M.J.; Wohleb, E.; Fogaca, M.V.; Teichman, E.M.; Hare, B.; et al. Ketamine rapidly reverses stress-induced impairments in GABAergic transmission in the prefrontal cortex in male rodents. Neurobiol. Dis. 2019, 134, 104669. [Google Scholar] [CrossRef] [PubMed]
- Talbot, J.N.; Geffert, L.M.; Jorvig, J.E.; Goldstein, R.I.; Nielsen, C.L.; Wolters, N.E.; Amos, M.E.; Munro, C.A.; Dallman, E.; Mereu, M.; et al. Rapid and sustained antidepressant properties of an NMDA antagonist/monoamine reuptake inhibitor identified via transporter-based virtual screening. Pharmacol. Biochem. Behav. 2016, 150, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Frazzini, V.; Granzotto, A.; Bomba, M.; Massetti, N.; Castelli, V.; d’Aurora, M.; Punzi, M.; Iorio, M.; Mosca, A.; Delli Pizzi, S.; et al. The pharmacological perturbation of brain zinc impairs BDNF-related signaling and the cognitive performances of young mice. Sci. Rep. 2018, 8, 9768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christoffel, D.J.; Golden, S.A.; Russo, S.J. Structural and synaptic plasticity in stress-related disorders. Rev. Neurosci. 2011, 22, 535–549. [Google Scholar] [CrossRef] [Green Version]
- Krzystyniak, A.; Baczynska, E.; Magnowska, M.; Antoniuk, S.; Roszkowska, M.; Zareba-Koziol, M.; Das, N.; Basu, S.; Pikula, M.; Wlodarczyk, J. Prophylactic Ketamine Treatment Promotes Resilience to Chronic Stress and Accelerates Recovery: Correlation with Changes in Synaptic Plasticity in the CA3 Subregion of the Hippocampus. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [Green Version]
- Duman, C.H.; Duman, R.S. Spine synapse remodeling in the pathophysiology and treatment of depression. Neurosci. Lett. 2015, 601, 20–29. [Google Scholar] [CrossRef] [Green Version]
- Goldin, M.; Segal, M. Protein kinase C and ERK involvement in dendritic spine plasticity in cultured rodent hippocampal neurons. Eur. J. Neurosci. 2003, 17, 2529–2539. [Google Scholar] [CrossRef]
- Ota, K.T.; Liu, R.J.; Voleti, B.; Maldonado-Aviles, J.G.; Duric, V.; Iwata, M.; Dutheil, S.; Duman, C.; Boikess, S.; Lewis, D.A.; et al. REDD1 is essential for stress-induced synaptic loss and depressive behavior. Nat. Med. 2014, 20, 531–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, V.; Zhang, M.X.; Swank, M.W.; Kunz, J.; Wu, G.Y. Regulation of dendritic morphogenesis by Ras-PI3K-Akt-mTOR and Ras-MAPK signaling pathways. J. Neurosci. 2005, 25, 11288–11299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Ren, Q.; Qu, Y.; Zhang, J.C.; Ma, M.; Dong, C.; Hashimoto, K. Mechanistic Target of Rapamycin-Independent Antidepressant Effects of (R)-Ketamine in a Social Defeat Stress Model. Biol. Psychiatry 2018, 83, 18–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Shirayama, Y.; Zhang, J.C.; Ren, Q.; Yao, W.; Ma, M.; Dong, C.; Hashimoto, K. R-ketamine: A rapid-onset and sustained antidepressant without psychotomimetic side effects. Transl. Psychiatry 2015, 5, e632. [Google Scholar] [CrossRef]
- Szewczyk, B.; Pochwat, B.; Rafalo, A.; Palucha-Poniewiera, A.; Domin, H.; Nowak, G. Activation of mTOR dependent signaling pathway is a necessary mechanism of antidepressant-like activity of zinc. Neuropharmacology 2015, 99, 517–526. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 2nd ed.; Academic Press: San Diego, CA, USA, 1986. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
 | Experimental Group | Locomotor Activity (10 min) |
| ZnA + NaCl | 100 ± 9.28 | |
| ZnA + Ro 25–6981 ZnA + Ketamine | 137.8 ± 39.01 76.22 ± 14.6 | |
| ZnD + NaCl | 70.76 ± 5.03 | |
| ZnD + Ro 25–6981 ZnD + Ketamine | 124.56 ± 12.3 66.93 ± 4.87 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pochwat, B.; Domin, H.; Rafało-Ulińska, A.; Szewczyk, B.; Nowak, G. Ketamine and Ro 25-6981 Reverse Behavioral Abnormalities in Rats Subjected to Dietary Zinc Restriction. Int. J. Mol. Sci. 2020, 21, 4791. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134791
Pochwat B, Domin H, Rafało-Ulińska A, Szewczyk B, Nowak G. Ketamine and Ro 25-6981 Reverse Behavioral Abnormalities in Rats Subjected to Dietary Zinc Restriction. International Journal of Molecular Sciences. 2020; 21(13):4791. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134791
Chicago/Turabian StylePochwat, Bartłomiej, Helena Domin, Anna Rafało-Ulińska, Bernadeta Szewczyk, and Gabriel Nowak. 2020. "Ketamine and Ro 25-6981 Reverse Behavioral Abnormalities in Rats Subjected to Dietary Zinc Restriction" International Journal of Molecular Sciences 21, no. 13: 4791. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134791