Functional Markers for Precision Plant Breeding


1
School of Biotechnology, Sher-e-Kashmir University of Agricultural Sciences & Technology of Jammu, Chatha, Jammu 190008, India
2
Department of Plant Sciences, University of Tennessee, Knoxville, TN 37996, USA
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2020, 21(13), 4792; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134792
Submission received: 4 May 2020
/
Revised: 19 June 2020
/
Accepted: 2 July 2020
/
Published: 6 July 2020
(This article belongs to the Special Issue Functional Genomics for Plant Breeding)
Abstract
:Advances in molecular biology including genomics, high-throughput sequencing, and genome editing enable increasingly faster and more precise cultivar development. Identifying genes and functional markers (FMs) that are highly associated with plant phenotypic variation is a grand challenge. Functional genomics approaches such as transcriptomics, targeting induced local lesions in genomes (TILLING), homologous recombinant (HR), association mapping, and allele mining are all strategies to identify FMs for breeding goals, such as agronomic traits and biotic and abiotic stress resistance. The advantage of FMs over other markers used in plant breeding is the close genomic association of an FM with a phenotype. Thereby, FMs may facilitate the direct selection of genes associated with phenotypic traits, which serves to increase selection efficiencies to develop varieties. Herein, we review the latest methods in FM development and how FMs are being used in precision breeding for agronomic and quality traits as well as in breeding for biotic and abiotic stress resistance using marker assisted selection (MAS) methods. In summary, this article describes the use of FMs in breeding for development of elite crop cultivars to enhance global food security goals.
1. Introduction
At the core of traditional plant breeding is phenotypic selection of superior genotypes. In reality, selections are the product of genetic recombination (genotype) and environment interactions. In plant breeding, genetic variability is the base for the improvement and the development of new cultivars. Genetic variability is repetitively produced in crosses and selections for desired traits. As we select beyond morphological traits to include genetic markers such as cytological and biochemical markers, more than 12 years may be needed to develop a variety. Herein, cataloging new genes and allele variants was initially accomplished using these markers and tracked in subsequent crosses. These days, these traditional markers have largely been supplanted by those from genomics approaches [1,2,3]. Genomic markers are not affected by environmental factors, sample collection stages, and the developmental stages of the plant [4].
Under a 50% higher demand for food by 2030 [5] from limited genetic and environmental resources, plant breeders are challenged to increase their output of superior varieties that are adapted to changing climates [6]. Further, the genetic base has narrowed with the introduction of high yielding varieties with a commensurate yield plateau [7]. Under these circumstances, plant breeders must explore and implement a wide range of genomic resources. There is a plethora of new breeding technologies and approaches that address global food security challenges, which simultaneously include both sustainability and humanitarian goals [8].
Precision breeding is a plant breeding approach in which a phenotypic trait of interest is selected by means of identifying a functional marker (FM) that is directly derived from the genomic region of a trait-controlling gene [9]. In precision breeding, selections are based on the polymorphic genic regions linked with a trait of interest. Availability of genomic resources are of utmost important for development of FMs and their use in precision breeding [10]. At the core of genomic innovations are complete genome sequences of cultivated crop species. In addition, systems biology approaches are used to infer relationships among transcripts, genes, proteins, and phenotypes [11]. Hence, hosts of molecular markers have been developed and are being used in plant breeding programs for genotype identification, phylogenetic studies, population structure and diversity analysis, and to better understand genome evolution [12,13].
2. Brief History of Molecular Marker Development
The genomics revolution during the 1990s greatly improved our understanding of the genetic make-up of a wide array of living organisms, including plants. With the advent of molecular marker technology, plant breeding became more efficient, including identifying quantitative trait loci (QTL) for use in genetic mapping [13]. Different types of molecular markers have been developed: restriction fragment length polymorphism (RFLP) [14], random amplification of polymorphic DNA (RAPD) [15,16], amplified fragment length polymorphism (AFLP) [17], microsatellite or simple sequence repeat (SSR) [18,19], sequence characterized amplified region (SCARs) [20], cleaved amplified polymorphic sequences (CAPS) [21], single nucleotide polymorphism (SNP) [22], and diversity arrays technology (DArT) markers [23]. Among these, SNP markers have gained the widest use [24]. Most of these marker types are random DNA markers (RDMs), which may be lost during recombination. In contrast, if candidate trait genes are known, then DNA markers (functional markers; FMs) within the gene or closely linked to the gene can be developed and used in agricultural crops. Herein lies the power of FMs.
FMs are defined as DNA markers that have been derived from functionally characterized sequence motifs [25]. Therefore, SNPs as FMs are more useful in plant breeding compared with RDMs and genic molecular markers (GMMs) (Figure 1). Although GMMs may be present within a gene of interest, functionally, they may not be linked to the phenotypic trait of interest, which may lead to false selection in MAS. FMs have been called “perfect markers” in contrast with RDMs as “non-perfect markers” such as RFLPs, AFLPs, or SSR markers [26]. Thus, this review assesses the state-of-the-art of FMs and their use in plant breeding.
3. Functional Markers
FMs in genic regions are optimally directly linked with a phenotypic trait, i.e., one that functionally characterizes the observed phenotypic variation [25]. FMs are also known as precision markers or diagnostic markers. FMs enable efficient and quick characterization and screening of germplasm for allelic diversity with accuracy since they are not subjected to recombination. Once genetic effects have been assigned to functional sequence motifs, FMs can be used for fixation of beneficial alleles in the breeding population [26]. As the polymorphism occurs within the target gene and fixes the favorable allele in the breeding population, the utility of FMs can also be made in selection of complex traits [27].
The development of FMs first requires the identification of a gene of interest affecting the phenotypic trait, its nucleotide sequence, and functional characterization [13]. Gene identification may be attained by a number of approaches such as expression profiling, map-based cloning, QTL mapping, expression profiling, and transposon tagging [28,29] (Figure 2). Plant transformation for overexpression or knockdown analysis is necessary for functional characterization of a candidate gene [30]. The second step towards FM development involves the study of allelic variation within characterized genes. Then, allele sequencing must be performed between genotypes to identify those polymorphisms that underlie causative variation of the phenotypic trait [31]. The development of FMs requires critical understanding about the sequence of allele/gene linked to the trait. These polymorphisms among alleles for genes of interest may be due to insertions or deletions (InDels), SNPs, different number of repeat motifs within SSRs, and partial or complete loss of the gene [28]. Hence, FMs may be based on a wide range of types of coding and non-coding DNA [32].
Over the last two decades with advancement of different next generation sequencing (NGS) platforms, numerous QTLs have been identified by developing various mapping populations such as recombinant inbred lines (RILs), near isogenic lines (NILs), doubled haploids (DHs), etc. Development of mapping population by crossing two diverse parents is time and cost consuming. To overcome these barriers, an alternate approach, association mapping, has been used for detecting QTL in natural or breeding germplasm. Association mapping (AM) is a powerful approach that identifies polymorphisms near or within a gene of interest that controls the phenotypic differences between genotypes [33]. Linkage disequilibrium (LD) tends to be maintained over many generations between loci that are genetically linked to one another, which enables marker assisted selection (MAS) [34]. Association mapping also facilitates the search for functional variation in genes of interest; the larger and more diverse the samples are, the greater the potential associations are [35]. Genic SNPs and maintenance of LD are quite helpful for FM development and subsequent marker-assisted backcross breeding (MABB) activities [36].
High-throughput genotyping technologies and large-scale genomic resources have led to ample SNP candidates for high-resolution linkage map production [37]. Furthermore, genome-wide association studies (GWAS) are now a viable alternative to QTL mapping for dissecting important quantitative traits. GWAS may provide efficient assessment representative sets of individuals and genes. GWAS leverage LD to enable high density genotyping that spans the entire genome of an organism to identify genomic regions linked to the phenotypic traits. Recently, GWAS have been successfully applied for identification of genomic regions associated with important traits in rice, barley, corn, wheat, and other crop species. GWAS in these crops have provided important information about the marker-trait associations, which can be successfully used in plant breeding programs [37]. Recently, GWAS have also been used in an association mapping population for identification of SNPs associated with total sugar content and sorbitol for improving fruit quality of lettuce [36] and peaches [38].
Advances in next-generation sequencing technologies have driven the costs of DNA sequencing down to the point that genotyping-by-sequencing (GBS) is now feasible for plant species with both high genetic diversity and large genomes [39] to the degree that they have become breeding tools. GBS involves the use of restriction enzymes for targeted complexity reduction of the genome followed by multiplexing and sequencing. GBS can generate numerous SNP markers covering much of the genome in a cost-effective manner [39,40]. Therefore, these genome-wide SNPs can be used in genomic selection, GWAS, and genetic diversity studies. In one example, GBS-derived SNPs associated with the functional allele E3Ha for maturity in soybean has been identified [41].
Similarly, transcriptome sequencing provides a new tool for genomic studies on model or non-model organisms. RNA-sequencing (RNA-Seq) is a powerful tool for transcription profiling, providing rapid access to a collection of expressed sequences. Transcriptome sequencing of an organism provides facile insights into the gene space, enabling gene discovery and FM development as well as study gene expression patterns and comparative biology [42]. RNA-Seq may enable molecular marker development, including FMs in non-model plant species that may not have a reference genome yet sequenced [43]. RNA-Seq has been successfully applied in different domains of life from yeast to plants. SNP detection is an important part of molecular genetic research because SNP loci can be exploited to construct high-density genetic maps and enable GWAS [44,45,46,47,48,49].
Functional SNPs are useful for improving breeding efficiency. With the progress in functional genomics research, increasing numbers of FMs responsible for agriculturally important traits have been identified, which provide valuable genetic resources for molecular breeding. Resequencing and SNP genotyping are two key strategies used in GWAS for identification of functional SNPs and development of FMs (Figure 3). Mapping populations (RILs, DHs, segregation, etc.) are used for appropriate QTL identification, in which genome resequencing of different lines generates saturated SNPs. The SNPs located at the QTL are regarded as GWAS SNPs, because the candidate genes at the QTL locus are predicted according to GWAS analysis. On the other hand, in association mapping, diverse germplasm is useful, and SNP genotyping on the basis of genomic resequencing provides a strong tool for the detection of SNPs in large accession collections. The comparison of GWAS SNPs from populations helps identify functional SNPs linked to the phenotypic trait.
Functional genomic techniques such as RNAi, site-directed mutagenesis, gene knockout analysis, and transposon tagging are useful in gene discovery [50]. With the possible exception of naturally occurring transposon systems in maize, most methods (transposon, T-DNA, antisense, and RNAi) rely on transgenic introduction of foreign DNA, which is not possible in most of the important crops. The development of transgenic is the main impediment in the use of functional genomics in the development of FMs in crops. However, RNAi has been exploited in the functional analysis of the 22-kD maize zein storage protein [51] and lysine rich in maize [52], the functional role of GhACT1 gene in fiber elongation in cotton [53]. However, RNAi generates unpredictable outcomes, and the whole procedure is laborious, as it requires vector construction, transformation, and transgenic analysis [54]. RNAi has several limitations such as partial and short-term suppression of genes rather than rendering a complete loss-of-function. In addition, off-target effects may cause false positive observations [55]. Still, RNAi experiments with a range of target gene suppression, which enables knowledge about the effect of gene expression in target tissues, i.e., gene function [56].
The emergence of different genome editing tools such as clustered regularly interspaced short palindromic repeat (CRISPR-Cas9), zinc finger nucleases (ZFNs), and transcription activator-like effector nucleases (TALENs) is important, as these are breakthrough technologies to knock-out genes. CRISPR-Cas9 approaches are especially relatively facile, affordable, and efficient research tools [57]. CRISPR-Cas9 produces double stranded breaks (DSBs) to the target loci, which can be repaired via homology directed repair (HDR) or non-homologous end joining (NHEJ) mechanisms. In most of the cases, NHEJ causes deletion mutations or random insertion of variable lengths, resulting in knockout mutants with frameshift mutations. Most often, coding regions of genes are disrupted, leading to a loss of an endogenous protein. This tool has numerous advantageous compared with RNAi technology, such as complete loss of function with relatively low off target activities [56], heritable and permanent or stable knock outs, and efficient characterization of non-coding RNAs (ncRNAs) by disrupting their DNA coding sequences. In addition, CRISPR-mediated knockouts in various crops [56,58] are likened to natural mutants and can be used directly in breeding and rapid crop domestication.
Point mutations for knock-in mutations may also result in gene gain-of-function; thus, using CRISPR-Cas9 in this way is also a breeding tool. HDR-mediated knock-ins have been achieved in rice, cotton, tobacco, poplar, sweet potato, and several other plant species [59].
Compared to different functional genomic approaches, targeting induced local lesions in genomes (TILLING) is a non-genetically modified organism (GMO) techniques and applied to any plant species regardless of genome size, ploidy level and mode of propagation. TILLING offers many advantages in cases where the transformation is difficult or if the investigation of a continuing series of unknown genes in a specific crop is desired. This technique requires no complicated manipulations or expensive apparatus. It enables one to screen the mutant pools easily for investigating the functions of specific genes, avoiding both confounding gene separation steps and tedious tissue culture procedures as are involved in anti-sense RNA and RNAi. It allows rapid and inexpensive detection of induced point mutations in populations of mutagenized individuals. Moreover, TILLING involves a series of alleles in a targeted locus compared to functional genomic approaches. In TILLING, the use of ethyl methanesulfonate (EMS) chemical mutagen produces G/C to A/T transition, which provides high frequency of point mutations distributed randomly in the genomes. Endonuclease cut effectively with the multiple mismatches in a DNA duplex and the heteroduplex DNA of unknown sequence to that of a known sequence reveals the positions of polymorphic sites. Therefore, both nucleotide changes and small insertions/deletions are identified. It can be performed with fewer expenses than the full sequencing methods currently used for most SNP discovery. Moreover, a well developed and tested protocol of TILLING is available for a number of crops, such as lotus [60], barley, common bean, field mustard, maize [61], oat, pea, peanut, potato, rice [62], rape seed, sorghum, soybean, Medicago spp., tomato, and wheat [63]. TILLING is an attractive strategy for a wide range of applications from the basic functional genomic study to practical crop breeding.
Regardless of the source of DNA, once FMs are developed, the next step is the functional validation of markers regarding the link to a gene of interest [64]. The validation of newly developed FMs for their functionality can be carried out by gene expression studies, including virus induced gene silencing (VIGS) and gene knock-down and knock-out analyses as noted above [65,66,67]. However, the VIGS technique has an advantage over the other techniques when it is useful to silence multiple genes within gene families, which has additional power in the analysis of polyploid species [65,68,69].
4. Advantages of FMs over Other Markers
RDMs are the most prevalent DNA markers used for indirect selection. RDMs are derived from the DNA sequence polymorphisms in the regions adjacent to the gene of interest; they do not always lead to predictive selection for traits of interest. However, the problems associated with the use of RDMs can now be overcome by the use of FMs that are 100% predictive of the corresponding phenotype. Once genetic effects have been assigned to functional sequence motifs, FMs fix alleles in several genetic backgrounds without additional calibration [70,71]. This is advantageous in marker applications, particularly in plant breeding, to select parental materials to build segregating populations as well as subsequent selection of advanced breeding lines [25].
FMs reside within the target genes themselves and are directly linked to the morphological traits that can be used with great reliability and efficiency to identify favorable alleles in a breeding program [36]. FMs also reduce the chances of loss and false selection of information using MAS [65,72]. Additionally, in MAS, QTL validation is needed when applied to different genetic backgrounds; however, FMs avoid this validation [30,65,73]. FMs can facilitate the selection of exceptional phenotypic traits that would enable a breeder to identify rare recombinants in a large population [74]. In addition, FMs are useful for screening for alleles in natural as well as breeding populations, fixation of alleles in the population, construction of linked FM haplotypes, and combination of FM alleles associated with complex traits [25].
5. FMs in Precision Plant Breeding
Genome sequencing costs continue to decrease, which has facilitated FM development. Since FMs outperform RDMs, FMs are increasingly being used in MABB activities for quantitative and quality traits [74,75]. In precision breeding, FMs can be used in germplasm evaluation, genetic diversity analysis, MAS, MABB, marker assisted recurrent selection (MARS), and genomic selection (GS) for improvement of important traits [29,76,77,78].
5.1. Germplasm Evaluation and Genetic Diversity
Plant genetic resources (PGR) are the basic material required for the improvement of crop species. These are the important sources of gene(s) for yield enhancement, quality improvement, disease and insect pests, as well as abiotic stresses. Before the advent of plant genomic approaches, the main genetic markers for evaluation of germplasm were various morphological traits [17]. Sequencing revolution enables the development of numerous molecular markers such as RDMs, GMMs, and FMs [79]. To screen the germplasm for allelic variation for particular phenotypic traits, FMs enable direct linkage with the gene of interest and can be used directly in breeding programs [80]. FMs enable the characterization and screening of the germplasm for allelic diversity with more accuracy. Compared to RDMs, FMs identify agronomically important genes directly from germplasm such as landraces, traditional cultivars, wild relatives, and plant genetic resources. The identified traits in the germplasm are linked with particular FMs and can be utilized in crossing programs for development of new cultivars. This also enables the plant breeder to develop new genetic resources to act as bridging genotypes for transferring the valuable genes to cultivated ones [81]. Besides this, the International Union for the Protection of New Varieties of Plants (UPOV) also endorsed the use of FMs for trait-specific characterization of germplasm and varieties for distinctness, uniformity, and stability (DUS) characters [82].
The knowledge of genetic variability present within and among the germplasm is the basis for the improvement and the development of crop varieties [83]. Genetic diversity information of PGR is essential and prerequisite for breeding programs. Today, vast PGRs that are available are characterized based on the phenotypic traits, but few of these have been characterized at the molecular level. Availability of high throughput genotypic techniques made large-scale use of molecular markers in genetic diversity analysis, identification of variety, conformity of F1 hybrid, and in plant variety protection [84,85]. For identification of potential and diverse genotypes for breeding, a broad genetic base is required [2,86]. The genomics era has provided several genomic resources such as RDM, GMM, and FM to assess the genetic variability in the available genetic resources [87]. Trait-specific FM genetic diversity is essential to broaden the genetic base to enable precision breeding [88]. FMs hold the promise for identification of alleles/genes involving polymorphisms causing functional genetic variation [89]. FMs can be effectively used to characterize the genetic diversity among closely related plant species based on functionally characterized genes linked with the phenotypic traits [90]. Alternatively, genetic diversity among genotypes can be assessed in the functional parts of the genome by DNA-based profiling methods [91,92,93].
5.2. Marker Assisted Selection
MAS is a molecular breeding technique in which direct and indirect phenotypic selections of a genotype are made on the basis of a molecular marker that may consist of RDMs, GMMs, and/or FMs [2]. MAS is used in crop improvement to overcome the difficulties that arise from the conventional plant breeding methods [94,95]. FMs used in MAS help in selection and identification of genotypes in a segregating population, which are directly linked with morphological traits (Figure 4). FMs have the uniqueness for confirmation of candidate genes governing the desired phenotypic traits, which can be used directly in the plant breeding methods through MAS. Several FMs have been developed and used for improvement of important morphological, quality, and biotic and abiotic stress resistance traits in different crops such as wheat, maize, rice, fruits, and vegetables. In these crops, FMs have aided in the improvement of various qualitative and quantitative traits such as flowering time, photoperiod response, plant height, seed length, seed weight, aroma, amylose content, oil content, and resistance to various diseases and insect pests [64,96,97]. These traits may be controlled by recessive or dominant alleles of a gene of interest.
MABB is used to transfer a phenotypic trait controlled by a recessive or dominant allele into an elite crop variety through repeated backcrosses and selections using molecular markers. In MABB, selection is operative both at genotypic as well as phenotypic levels. In MABB, the desired traits are transferred from the donor parent (non-recurrent parent) to a widely-adapted variety (recurrent parent). Repeated backcrossing and selection are practiced on the basis of molecular markers and the phenotypic traits until most of the genes stemming from the donor are eliminated. In MABB, FMs are used to target the gene controlling the desired trait of interest (foreground selection) and in recovering the genome of an elite variety (background selection) if a sufficient number of markers are available. FMs have been successfully employed to select for a number of genes controlling desired traits that have been transferred to adapted varieties via MABB [72]. The efficiency of MABB can be enhanced by using FMs to transfer the desired gene(s) controlling simple or complex trait(s) into cultivated varieties [98]. Moreover, FMs can be used for multiplex screening assays for foreground selection to identify introgressed genes [99,100].
MARS plant breeding methods for transferring complex traits into varieties take a long time under continuous recurrent selection. The phenotypic selection of complex traits may be difficult because of ambiguous phenotypic selection and analysis of complex traits owing to the large number of genes that contribute to the traits. In MARS, favorable alleles of complex traits are accumulated with the help of genetic markers and are thus made more efficient. Among the various genetic markers, FMs help the accumulation of favorable alleles of a complex trait most efficiently [101]. For improvement of complex traits, several recurrent selection cycles are required to accumulate favorable QTL alleles in the breeding population [102], which is aided by use of FMs. MARS thus decreases the number of needed breeding cycles while increasing the precision of selecting complex traits. When parents used in MARS are crossed when informed by FMs, an ideal genotype is possible after only a few successive generations of backcrossing. FMs enable genetic gain for the improvement of complex traits and the development of inbred lines of a hybrid [98].
5.3. Gene Pyramiding
Gene pyramiding is a method of assemblage of different desirable genes from various donor parents into a single plant (Figure 5). In gene pyramiding, genes controlling different traits are simultaneously transferred into a single cultivar. Gene pyramiding is used for improving few unsatisfactory traits of a widely-grown elite variety, and these unsatisfactory genes are replaced with better genes. Although gene pyramiding is possible through conventional plant breeding methods, phenotypic selection and identification of a single plant containing more than one gene are very difficult. There are chances of loss of gene of interest from recombination and number of meiotic cycles, which may convolute plant breeding [103]. FMs can improve the prospects of gene pyramiding for different traits [104] as demonstrated by the plethora of FMs linked with a multitude of morphological traits, quality traits, and resistance to biotic and abiotic stresses for use in many crops (Table 1) [64,105].
5.4. Genomic Selection
Genomic selection (GS) was developed by Meuwissen et al. [106], which is an advanced method of MAS. GS has aided the improvement of complex traits such as grain yield and its components, quality traits, and abiotic stress resistance, which vary rapidly with generation of desired phenotypes by selection [107]. GS is also known as genomics-assisted breeding (GAB), which uses phenotypic data, genotypic data, and modeling to predict the genomic estimated breeding values (GEBVs) for each individual [108,109]. In this method, GEBVs are used to predict the genetic values of selected candidates predicted from high density of markers that depends on all the major and the minor molecular effects [110]. GS requires: (i) a diverse population used for development of a training population; (ii) genotypic as well as phenotyping analysis of the training population; (iii) genotypes with high values of GEBV to be selected on the basis of their genotypic data; (iv) a testing population composed of progeny of the genotypes used as study material that are taken as input for the GS model to yield GEBVs; (v) high GEBV values are selected again in genotypes; and iv) the selected genotypes are used as parents for continuous crossing and selection [111]. In GS, FMs have advantages over other markers as few numbers of trait-specific FMs are required for predicting GEBV values. GS has been used in several crops including wheat [112], maize [113], and Brassica rapa [114]. Increased prediction of GEBVs is facilitated by using FMs [115].
6. FMs for the Improvement of Agronomic Traits, Quality Traits, and Stress Resistance
Advances in sequencing techniques enable the identification of SNPs and indels linked with various economically important traits; FM development is thus enabled [64]. Indels may cause phenotypic variation from extensive genomic effects, which are accompanied by chances of elimination from natural selection [116]. Hence, SNP-derived FMs have advantages over indel-derived markers because the widely distributed nature of FMs throughout the genome [117,118]. FMs have been developed for various agronomic, quality traits, and biotic and abiotic stress resistances, which have been pyramided in different crops using MAS, MABB, MARS, and GS approaches [64] (Table 1).
6.1. FMs for Agronomic Traits
FMs perfectly discriminate alleles of a targeted gene, and FMs have been deployed for improvement of important agronomic traits through MABB. In wheat, more than 97 FMs have been developed and used in MAS [119]. Two FMs for TaSUS2-2B and TaZds-D1 genes encoding grain weight have been developed, which can be used in MABB for wheat improvement [120]. Similarly, FM markers that are based on SNPs present in the eighth exon of the TaGW2 gene have been developed for enhancing grain weight in wheat [121]. In wheat, the pre-sprouting of spikes results in low grain yield production and development of low quality products. An SNP-based CAPS marker was developed for TaSDr gene responsible for low spike sprouting in wheat crop under field conditions [122]. Two semi-dwarf genes, such as Rht1 and Rht2, encode a protein involved in GA-signal transduction located on chromosome 4B and 4D of wheat and played a significant role in the Green Revolution. Rht1 and Rht2 are the result of point mutations to change the tall genotypes of wheat to semi-dwarf ones. FMs were developed for screening the semi-dwarf genotypes in wheat germplasm [123].
In rice, the erect panicle trait significantly contributes to increased yield for which an FM has been developed that is transferable to other rice varieties through molecular breeding [118]. Similarly, the semi-dwarf gene sd1 on chromosome 1 in rice also played a role in the Green Revolution. This is the most widely deployed gene in modern rice breeding programs in the world for development of semi-dwarf varieties. The sd1 gene FM was developed owing to a 280 bp deletion within the coding region of the Os20ox2 gene that encodes the non-functional protein that leads to reduction in gibberellic acid (GA3) production in dwarf rice plants [124]. An FM enabled the transfer of sd1 gene in Ranbir Basmati rice through MABB [125]. In the two-line system of hybrid rice production, photoperiod-thermo-sensitive genic male (PGMS and TGMS) sterility is essential. An FM has been developed for the pms3 (p/tms12-1) gene to transfer male sterility in other rice varieties using MABB [126].
In maize, an FM for reduced plant height along with early flowering has been developed for the Dwarf 8 gene from a deletion of bases in the coding region of the gene. This marker has been used for screening of maize germplasm for early flowering time and reduced plant height [31]. In barley, an FM has been developed because of deletions in the intron region in VRN-H 1 gene. This gene is responsible for regulation of spring growth habit and vernalization in barley [127]. In mustard crops, trichomes provide protection against number of insect pests and diseases. An FM for BrpHL1, a trichome-related gene, is used in mustard breeding [128]. Similarly, FMs have been developed for agronomically-important traits of legumes and vegetable crops (Table 1).
6.2. FM for Quality Traits
Quality traits are important for a variety of reasons, such as to meet consumer preferences. FMs have been successfully applied in MAS to improve the food quality of crops [72]. A wheat FM, YP7A, was developed for the Psy1 gene, which is involved with yellow grain pigment [128]. Similarly, an FM has been developed for TaZds-D1, which is also responsible for increased yellow pigmentation in wheat [152]. Gluten proteins determine bread-making quality of wheat. An SNP-based FM for Glu-B3 encoding low molecular weight gluten protein has been developed for improving the quality of wheat products [151].
In rice, particularly basmati, aroma is one of the most important components for determining the price in the world market. Basmati, the aromatic rice praised for its unique quality, is a connoisseur’s delight, which has a pleasant aroma and is nature’s gift. A set of FMs linked to a recessive badh2 gene responsible for 1-acetyl-2-pyrroline (AP) production has been identified and exploited for screening rice germplasm for fragrance [154]. Grain length and grain breadth also determine the grain qualities of rice. Long grain length adds an aesthetic value in rice for appearance and fetches a higher price in the world market. For grain length in rice, an SNP-based FM has been developed for gene GS3 encoding trans-membrane protein for use in rice breeding. The GS3 gene governs grain elongation after cooking longitudinally, an important trait affecting the physical appearance of the rice grains [220]. In rice, intermediate amylose content is preferred by the consumers, and an FM has been developed for gene Wx-in encoding moderate amylose content for screening of rice germplasm [159]. Similarly, in sorghum, an InDel based FM has been developed for the SbBADH2 gene responsible for fragrance. This FM has been used in identification of sorghum genotypes possessing high fragrance, which can be further used in sorghum breeding [165].
Maize is an important cereal crop of the world, which is used as both food and feed crop. The quality improvement in maize is important to enhance the nutritional values for humans and animals. To increase the nutritional values of maize, plant breeders are successfully exploiting the modern molecular techniques for screening the genotypes and their use in breeding. In this context, SNP-based FMs for ZmcrtRB3, which encodes a carotenoid hydroxylase, have been developed in maize [161]. This gene affects a-carotene content in maize and is successfully transferred to other maize varieties to enhance the pro-vitamin A content. Similarly, FMs have been developed for crtRB1 and LcyE encoding pro-vitamin A content in maize [163]. Sweetness is an important quality trait in maize and is encoded by the sugary1 gene. For successful screening and transfer of this trait, an FM has been developed [162]. Maize oil is a desirable best vegetable oil because of its high smoking point while frying. A gene DGAT1-2 has been identified for governing oleic acid quantity and oil content in maize. An FM has been developed for this candidate gene to screen maize germplasm [160].
In soybean, an FM for Gmbadh2-1 and Gmbadh2-2 genes, which are responsible for fragrance production, has been developed [166].
Celery is an important vegetable crop, which is rich in minerals and vitamins such as A, C, K, potassium, folate, and flavonoids. In celery, high flavonoid content is encoded by AgFNSI, and an FM has been developed for screening germplasm for this trait to be used in breeding [168].
6.3. FMs for Biotic Stress Resistance
Crop yield losses are caused by various pests, including diseases and insects. Several disease resistance genes have been identified and transferred into elite cultivars with the help of FMs (Table 1). Wheat diseases include leaf rust, yellow rust, brown rust, and powdery mildew. FMs for disease resistance genes have been developed for pyramiding of different resistance genes into various wheat varieties [221]. In wheat, FMs for resistance to leaf rust, yellow rust, and powdery mildew, such as Lr 34, Yr 18, and Pm 38, have been developed and used for pyramiding into elite wheat varieties for durable resistance to these diseases [174,222]. Similarly, an FM has been developed for gene Sr45 encoding wheat stem rust resistance to be used in MABB [175].
Rice crop yield may be decreased by various diseases and insect pests such as bacterial leaf blight, blast, brown spot, and brown hopper. With the accessibility of complete genome sequences of rice subspecies indica and japonica, a number of FMs, such as Xa3, xa5, xa13, Xa21, and Xa38, have been developed for bacterial leaf blight [103,176,177,178,179]. Similarly, FMs have been developed for blast resistance genes such as Pit, Pi54 (Pikh), Pi35, and PigmR [182,183,184,185]. These FMs have been used in MABB for foreground selection of various resistance genes for development of rice cultivars such as “Samba Mahsuri” [178,223], “Improved Pusa Basmati 1” [224], “Pusa 1121” [180], “Improved Tapaswini” [225], and “Ranbir Basmati” [125].
In tomato, an FM for ACY, a gene that confers tomato yellow leaf curl virus resistance was developed and used to produce leaf curl virus resistance tomato cultivars [190]. FMs for bacterial wilt and fusarium wilt have been developed in tomato to be used in breeding [191].
In watermelon, FMs have been developed for Zucchini Yellow Mosaic Virus (ZYMV) resistance gene eIF4E (197) and powdery mildew resistance gene Pm [198]. Similarly, an FM for controlling Cauliflower Mosaic Virus (CaMV) resistance has been developed for the cmv6.1 gene in cucumber [200]. Oumouloud et al. [193] developed an FM for the Fom 1 gene responsible for controlling fusarium disease in melon. An FM has been developed for Rpf gene encoding leaf scald resistance in sugarcane, which can be used for development of sugarcane varieties resistance to this disease [194].
6.4. FMs for Abiotic Stress Tolerance
Abiotic stress is one of the major unpredictable and uncontrolled factors in crop production. Abiotic stress resistance is a complex trait controlled by polygenes and depends upon the time and the severity of the stress components. All these unprecedentedly and uncontrolled factors of abiotic stress make it difficult to characterize and develop FMs for abiotic stresses. Very few FMs have been developed for abiotic stresses compared to biotic stresses; however, one goal is to discover functional variations linked to the abiotic stress traits (Table 1). In wheat, an SNP linked with the dehydration tolerance has been identified in the TaMYB2 transcription factor gene; an allele-specific FM has been developed for use in MABB [208]. Similarly, two SNPs were identified in the DREB1 gene in wheat, and an allele-specific FM has been developed [210]. Pandey et al. [209] developed SNP-based FMs for the TaAQP gene encoding drought tolerance, which was validated in wheat varieties. Similarly, an FM for the TtASR1 gene encoding salt tolerance in wheat has been developed, which should be helpful to screen wheat germplasm for salt tolerance [211].
In rice, the phosphate (P) uptake 1 (Pup1) gene confers P-deficiency stress tolerance in field-grown rice. A Pup1 gene-specific SNP, indel-, and CAPS-based FMs have been developed to transfer this trait in P-deficiency-susceptible varieties through MAS [213]. Most areas of Asian countries are flooded during rice growing period in rainy seasons, and a tolerance gene Submergence-A1 (SubA1) for flooding has been identified. An FM for the SubA1 gene has been developed and successfully used to transfer flooding tolerance in the “Swarna” variety of rice [212].
In foxtail millet, an SNP-based FM has been developed for the SiDREB2 gene, which is responsible for a dehydration response [218].
Similarly, an FM for the SbMATE gene, which confers aluminum stress tolerance in sorghum, has been developed [216]. The SbMATE FM helps in the screening of sorghum germplasm for aluminum stress tolerance, which can be used in breeding.
In cowpea, an FM for CPRD12 gene conferring drought and salt tolerance has been developed [217].
7. Future Prospects and Conclusions
FM development for row crops has been implemented in plant breeding, but there has been meager FM research in horticultural and forage crops. Since FMs are derived chiefly from coding DNA within genes, these markers hold prospects for directly selecting a phenotypic trait. FMs also may be feasible for interspecies transferability and can be used in those species for which limited genomic resources are available. For successful implementation of precision breeding, the integration of advanced genomic tools with conventional plant breeding methods is essential. Moreover, development of cost-effective FMs is important for the efficient execution of precision breeding in crops. FMs can be effectively used in screening of germplasm, diversity analysis, QTL mapping, gene identification and isolation, and phylogenetic studies. In these studies, the use of FMs increases the accuracy and the efficiency of plant breeding for cultivar development with desired traits. In the future, allele-specific development of FMs for newly identified genes is essential for enabling efficient direct selection. Moreover, modern genomic and breeding approaches such as GWAS and GS have not been fully exploited for crop improvement but can be increasingly deployed in both model and non-model crop species with the availability of these NGS-based markers. In spite of available genomic resources, more work is needed to identify and develop FMs and to implement them in MAS for food security and sustainability goals. FMs may be used in the future for new plant breeding techniques using biotechnology in precision breeding.
In the future, we expect that new plant breeding techniques (NPBTs) using gene editing, cisgenesis, and epigenetic approaches will play an increasingly greater role in variety development. They can potentially confer traits that may be difficult via traditional breeding. Some examples include stress tolerance, shelf life, color, yield, and nutritional content. The results of conventional plant breeding are sometimes difficult to predict and require several years to fix traits in varieties. NPBTs may allow the breeders to develop improved varieties more precisely and more quickly. We expect that FMs can also come into play as NPBTs hit their stride to allow for efficient introgression of novel traits.
Author Contributions
R.K.S. designed and wrote the manuscript. C.N.S.J. advised, conceived the concept, and improved the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
A USDA Hatch grant and funding from the Ivan Racheff Endowment to CNS enabled the writing of this review paper.
Conflicts of Interest
The authors declare that they have no competing interests.
Compliance with Ethical Standards
All applicable ethical standards were followed.
References
- Gupta, P.K.; Rustgi, S.; Kulwal, P.L. Linkage disequilibrium and association studies in higher plants: Present status and future prospects. Plant Mol. Biol. 2005, 57, 461–485. [Google Scholar] [CrossRef] [PubMed]
- Collard, B.C.; Jahufer, M.Z.; Brouwer, J.B.; Pang, E.C.K. An introduction to markers, quantitative trait loci (QTL) mapping and marker-assisted selection for crop improvement: The basic concepts. Euphytica 2005, 142, 169–196. [Google Scholar] [CrossRef]
- Kumar, M.; Chaudhary, V.; Sharma, R.; Sirohi, U.; Singh, J. Advances in biochemical and molecular marker techniques and their applications in genetic studies of orchid: A review. Int. J. Chem. Stud. 2018, 6, 806–822. [Google Scholar]
- Winter, P.; Kahl, G. Molecular marker technologies for plant improvement. World J. Microbiol. Biotechnol. 1995, 11, 438–448. [Google Scholar] [CrossRef] [PubMed]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [Green Version]
- Frona, D.; Szenderak, J.; Harangi-Rakos, M. The challenge of feeding the World. Sustainability 2019, 11, 5816. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Xie, W.; He, H.; Yu, H.; Chen, W.; Li, J.; Yu, R.; Yao, Y.; Zhang, W.; He, Y.; et al. A high-density SNP genotyping array for rice biology and molecular breeding. Mol. Plant 2014, 7, 541–553. [Google Scholar] [CrossRef] [Green Version]
- Oliver, M.J. Why we need GMO crops in agriculture. Mo. Med. 2014, 111, 493–507. [Google Scholar]
- Conner, T. Precision breeding: A new genetic technique providing international opportunities for crop improvement. Seed Quest 2004. Available online: https://www.seedquest.com (accessed on 15 April 2020).
- Bohra, A.; Bharadwaj, C.; Radhakrishnan, T.; Singh, N.P.; Varshney, R.K. Translational genomics and molecular breeding for enhancing precision and efficiency in crop improvement programs: Some examples in legumes. Indian J. Genet. 2019, 79, 227–240. [Google Scholar] [CrossRef]
- Schmutz, J.; Steven, B.; Cannon Schlueter, J.; Jianxin, M.; Therese, M.; William, N.; David, H.L.; Qijian, S.; Jay, T.J.; Jianlin, C. Genome sequence of the palaeopolyploid soybean. Nature 2010, 463, 178–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhutmal, R.R.; Mundhe, A.G.; More, A.W. Molecular marker techniques: A Review. Int. J. Curr. Microbiol. Appl. Sci. 2018, 6, 816–825. [Google Scholar]
- Amom, T.; Nongdam, P. The use of molecular marker methods in plants: A review. Int. J. Curr. Res. Rev. 2017, 9, 1–9. [Google Scholar] [CrossRef]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphism. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar] [PubMed]
- Williams, J.; Kubelik, A.; Livak, K.; Rafalski, J.; Tingey, S. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res. 1990, 18, 6531–6535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welsh, J.; McClelland, M. Fingerprinting genomes using PCR with arbitrary primers. Nucleic Acids Res. 1990, 18, 7213–7218. [Google Scholar] [CrossRef] [Green Version]
- Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; van de Lee, T.; Hornes, M.; Frijters, A.; Pot, J.; Peleman, J.; Kuiper, M. AFLP: A new technique for DNA fingerprinting. Nucleic Acids Res. 1995, 23, 4407–4414. [Google Scholar] [CrossRef] [Green Version]
- Tautz, D. Hypervariability of simple sequences as a general source of polymorphic DNA markers. Nucleic Acids Res. 1989, 17, 6463–6471. [Google Scholar] [CrossRef]
- Litt, M.; Luty, J.A. A hypervariable microsatellite revealed by in vitro amplification of a dinucleotide repeat within the cardiac muscle actin gene. Am. J. Hum. Genet. 1989, 44, 397–401. [Google Scholar]
- Paran, I.; Michelmore, R.W. Development of reliable PCR-based markers linked to downy mildew resistance genes in lettuce. Theor. Appl. Genet. 1993, 85, 985–993. [Google Scholar] [CrossRef]
- Konieczny, A.; Ausubel, F.M. A procedure for mapping Arabidopsis mutations using co-dominant ecotype-specific PCR-based markers. Plant J. 1993, 4, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.K.; Roy, J.K.; Prasad, M. Single nucleotide polymorphisms: A new paradigm for molecular marker technology and DNA polymorphism detection with emphasis on their use in plants. Curr. Sci. 2001, 80, 524–535. [Google Scholar]
- Jaccoud, D.; Peng, K.; Feinstein, D.; Kilian, A. Diversity arrays: A solid state technology for sequence information independent genotyping. Nucleic Acids Res. 2001, 29, E25. [Google Scholar] [CrossRef] [Green Version]
- Sousa, T.V.; Caixeta, E.T.; Alkimim, E.R.; Oliveira, A.C.B.; Pereira, A.A.; Sakiyama, N.S.; Zambolim, L.; Resende, M.D.V. Early selection enabled by the implementation of genomic selection in Coffea arabica breeding. Front. Plant Sci. 2019, 9, 1934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, J.R.; Lubberstedt, T. Functional markers in plants. Trends Plant Sci. 2003, 8, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Varshney, R.K.; Graner, A.; Sorrells, M.E. Genomics-assisted breeding for crop improvement. Trends Plant Sci. 2005, 10, 621–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagge, M.; Xia, X.; Lubberstedt, T. Functional markers in wheat. Curr. Opin. Plant Biol. 2007, 10, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Borevitz, J.O.; Nordborg, M. The impact of genomics on the study of natural variation in Arabidopsis. Plant Physiol. 2003, 132, 718–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salgotra, R.K.; Gupta, B.B.; Stewart, C.N. From genomics to functional markers in the era of next-generation sequencing. Biotechnol. Lett. 2014, 36, 417–426. [Google Scholar] [CrossRef]
- Ingvardsen, C.; Schejbel, B.; Lubberstedt, T. Functional markers in resistance breeding. In Progress in Botany; Springer: Berlin/Heidelberg, Germany, 2008; Volume 69, pp. 61–87. [Google Scholar]
- Thornsberry, J.M.; Goodman, M.M.; Doebley, J.; Kresovich, S.; Nielsen, D.; Buckler, E.S. Dwarf8 polymorphisms associate with variation in flowering time. Nat. Gen. 2001, 28, 286–289. [Google Scholar] [CrossRef]
- McCouch, S.R.; Chen, X.; Panaud, O.; Temnykh, S.; Xu, Y.; Cho, Y.G.; Huang, N.; Ishii, T.; Blair, M. Microsatellite marker development, mapping and applications in rice genetics and breeding. Plant Mol. Biol. 1997, 35, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Soto-Cerda, B.J.; Cloutier, S. Association mapping in plant genomes. In Genetic Diversity in Plants; Mahmut, C., Ed.; IntechOpen Limited: London, UK, 2012; ISBN 978-953-51-0185-7. [Google Scholar]
- Rostoks, N.T.; Ramsay, L.; Mackenzie, K.; Cardle, L.; Bhat, P.R.; Roose, M.L.; Svensson, J.T.; Stein, N.; Varshney, R.K.; Marshall, D.F.; et al. Recent history of artificial outcrossing facilitates whole-genome association mapping in elite inbred crop varieties. Proc. Natl. Acad. Sci. USA 2006, 103, 18656–18661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breseghello, F.; Sorrels, M.E. Association analysis as a strategy for improvement of qualitative traits in plants. Crop Sci. 2006, 46, 1323–1330. [Google Scholar] [CrossRef]
- Simko, I.; Pechenick, D.A.; McHale, L.K.; Truco, M.J.; Ochoa, O.E.; Michelmore, R.W.; Scheffler, B.E. Association mapping and marker-assisted selection of the lettuce dieback resistance gene Tvr1. BMC Plant Biol. 2009, 9, 135. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Nie, X.; Shen, C.; You, C.; Li, W.; Zhao, W.; Zhang, X.; Lin, Z. Population structure and genetic basis of the agronomic traits of upland cotton in China revealed by a genome-wide association study using high-density SNPs. Plant Biotechnol. J. 2017, 15, 1374–1386. [Google Scholar] [CrossRef]
- Forcada, C.; Guajardo, V.; Chin-Wo, S.R.; Moreno, M.Á. Association mapping analysis for fruit quality traits in Prunus persica using SNP markers. Front. Plant Sci. 2019, 9, 2005. [Google Scholar] [CrossRef] [Green Version]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple genotyping-by sequencing (GBS) approach for high diversity species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef] [Green Version]
- Sonah, H.; Bastien, M.; Iquira, E.; Tardivel, A.; Tardivel, A.; Legare, G.; Boyle, B.; Normandeau, E.; Laroche, J.; Larose, S.; et al. An improved genotyping by sequencing (GBS) approach offering increased versatility and efficiency of SNP discovery and genotyping. PLoS ONE 2013, 8, e54603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tardivel, A.; Sonah, H.; Belzile, F.; O’Donoughue, L.S. Rapid identification of alleles at the soybean maturity gene E3 using genotyping by sequencing and a haplotype-based approach. Plant Genome 2014, 7, 1–9. [Google Scholar] [CrossRef]
- Garg, R.; Jain, M. RNA-Seq for transcriptome analysis in non-model plants. Methods Mol. Biol. 2013, 43–58. [Google Scholar] [CrossRef]
- Chen, X.R.; Xing, Y.P.; Li, Y.P.; Tong, Y.H.; Xu, J.Y. RNA-Seq reveals infection-related gene expression changes in Phytophthora capsici. PLoS ONE 2013, 8, e74588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novaes, E.; Drost, D.R.; Farmerie, W.G.; Pappas, G.J.; Grattapaglia, D.; Sederoff, R.R.; Kirst, M. High-throughput gene and SNP discovery in Eucalyptus grandis, an uncharacterized genome. BMC Genom. 2008, 9, 312. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Rupe, M.A.; Danilevskaya, O.N.; Yang, X.; Hu, Z. Genome-wide mRNA profiling reveals heterochronic allelic variation and a new imprinted gene in hybrid maize endosperm. Plant J. 2003, 36, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Duran, C.; Appleby, N.; Clark, T.; Wood, D.; Imelfort, M.; Batley, J.; David, E. AutoSNPdb: An annotated single nucleotide polymorphism database for crop plants. Nucleic Acids Res. 2009, 37, 951–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Wang, L.; Zhan, Q.; Liu, Y.; Yang, X. Transcriptome characterization and functional marker development in Sorghum sudanense. PLoS ONE 2016, 11, e0154947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Son, S.; Shin, M.; Fujii, N.; Hoshino, T.; Park, S. Transcriptome-wide mining, characterization, and development of microsatellite markers in Lychnis kiusiana (Caryophyllaceae). BMC Plant Biol. 2019, 19, 14. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Chu, P.; Wu, Y.; Chan, W.R.; Wang, Y.H. Identification of functional SSR markers in freshwater ornamental Shrimps Neocaridina denticulata using transcriptome sequencing. Mar. Biotechnol. 2020. [Google Scholar] [CrossRef]
- Abdurakhmonov, I.Y.; Ayubov, M.S.; Ubaydullaeva, K.A.; Buriev, Z.T.; Shermatov, S.E.; Ruziboev, H.S.; Shapulatov, U.M.; Saha, S.; Ulloa, M.; Yu, J.Z.; et al. RNA interference for functional genomics and improvement of cotton (Gossypium sp.). Front. Plant Sci. 2016, 7, 202. [Google Scholar] [CrossRef] [Green Version]
- Segal, G.; Song, R.; Messing, J. A new opaque variant of maize by a single dominant RNA-interference-inducing transgene. Genetics 2003, 165, 387–397. [Google Scholar]
- Angaji, S.A.; Hedayati, S.S.; Poor, R.H.; Poor, S.S.; Shiravi, S.; Madani, S. Application of RNA interference in plants. Plant Omics J. 2010, 3, 77–84. [Google Scholar]
- Machado, A.; Wu, Y.; Yang, Y.; Llewellyn, D.J.; Dennis, E.S. The MYB transcription factor GhMYB25 regulates early fiber and trichome development. Plant J. 2009, 59, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Que, Q.; Jorgensen, R.A. Homology-based control of gene expression patterns in transgenic petunia flowers. Dev. Genet. 1998, 22, 100–109. [Google Scholar] [CrossRef]
- Sigoillot, F.D.; Lyman, S.; Huckins, J.F.; Adamson, B.; Chung, E.; Quattrochi, B.; King, R.W. A bioinformatics method identifies prominent off targeted transcripts in RNAi screens. Nat. Methods 2012, 9, 363–366. [Google Scholar] [CrossRef] [Green Version]
- Schaeffer, S.M.; Nakata, P.A. CRISPR/Cas9-mediated genome editing and gene replacement in plants: Transitioning from lab to field. Plant Sci. 2015, 240. [Google Scholar] [CrossRef] [PubMed]
- Romay, G.; Bragard, C. Antiviral defenses in plants through genome editing. Front. Microbiol. 2017, 8, 47. [Google Scholar] [CrossRef] [Green Version]
- Fan, D.; Liu, T.; Li, C.; Jiao, B.; Li, S.; Hou, Y.; Luo, K. Efficient CRISPR/Cas9-mediated targeted mutagenesis in Populus in the first generation. Sci. Rep. 2015, 5, 12217. [Google Scholar] [CrossRef]
- Liu, D.; Hu, R.; Palla, K.J.; Tuskan, G.A.; Yang, X. Advances and perspectives on the use of CRISPR/Cas9 systems in plant genomics research. Curr. Opin. Plant Biol. 2016, 30, 70–77. [Google Scholar] [CrossRef] [Green Version]
- Perry, J.A.; Wang, T.L.; Welham, T.J.; Gardner, S.; Pike, J.M.; Yoshida, S.; Parniske, M. A TILLING reverse genetics tool and a web-accessible collection of mutants of the legume Lotus japonicus. Plant Physiol. 2003, 131, 866–871. [Google Scholar] [CrossRef] [Green Version]
- Till, B.J.; Reynolds, S.H.; Weil, C.; Springer, N.; Burtner, C.; Young, K.; Bowers, E.; Codomo, C.A.; Enns, L.C.; Odden, A.R.; et al. Discovery of induced point mutations in maize genes by TILLING. BMC Plant Biol. 2004, 4, 12. [Google Scholar] [CrossRef] [Green Version]
- McCallum, C.M.; Comai, L.; Greene, E.A.; Henikoff, S. Targeted screening for induced mutations. Nat. Biotechnol. 2000, 18, 455–457. [Google Scholar] [CrossRef]
- Slade, A.J.; Fuerstenberg, S.I.; Loeffler, D.; Steine, M.N.; Facciotti, D. A reverse genetic, nontransgenic approach to wheat crop improvement by TILLING. Nat. Biotechnol. 2005, 23, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Kage, U.; Kumar, A.; Dhokane, D.; Karre, S.; Kushalappa, A.C. Functional molecular markers for crop improvement. Crit. Rev. Biotechnol. 2016, 36, 917–930. [Google Scholar] [CrossRef] [PubMed]
- Rodenburg, R.J. The functional genomics laboratory: Functional validation of genetic variants. J. Inherit. Metab. Dis. 2018, 41, 297–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burch-Smith, T.M.; Anderson, J.C.; Martin, G.B.; Dinesh-Kumar, S.P. Applications and advantages of virus—Induced gene silencing for gene function studies in plants. Plant J. 2004, 39, 734–746. [Google Scholar] [CrossRef]
- Tadege, M.; Ratet, P.; Mysore, K.S. Insertional mutagenesis: A swiss army knife for functional genomics of Medicago truncatula. Trends Plant Sci. 2005, 10, 229–235. [Google Scholar] [CrossRef]
- Gupta, P.K. Beyond CRISPR: Single base editors for human health and crop improvement. Curr. Sci. 2019, 116, 386–397. [Google Scholar] [CrossRef]
- Robertson, D. VIGS vectors for gene silencing: Many targets, many tools. Annu. Rev. Plant Biol. 2004, 55, 495–519. [Google Scholar] [CrossRef]
- Cakir, C.; Gillespie, M.E.; Scofield, S.R. Rapid determination of gene function by virus-induced gene silencing in wheat and barley. Crop Sci. 2010, 50, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.K.; Kumar, J.; Mir, R.R.; Kumar, A. Marker-assisted selection as a component of conventional plant breeding. In Plant Breeding Reviews; Janick, J., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2010; pp. 145–217. [Google Scholar]
- Lau, W.C.P.; Rafii, M.Y.; Ismail, M.R.; Puteh, A.; Latif, M.A.; Ramli, A. Review of functional markers for improving cooking, eating, and the nutritional qualities of rice. Front. Plant Sci. 2015, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Nawaz, M.A.; Yang, S.H.; Rehman, H.M.; Baloch, F.S.; Lee, J.D.; Park, J.H.; Chung, G. Genetic diversity and population structure of Korean wild soybean (Glycine soja Sieb. and Zucc.) inferred from microsatellite markers. Biochem. Syst. Ecol. 2017, 71, 87–96. [Google Scholar] [CrossRef]
- Edmeades, G.O.; McMaster, G.S.; White, J.W.; Campos, H. Genomics and the physiologist: Bridging the gap between genes and crop response. Field Crops Res. 2004, 90, 5–18. [Google Scholar] [CrossRef]
- Yang, H.; Li, C.; Lam, H.M.; Clements, J.; Yan, G.; Zhao, S. Sequencing consolidates molecular markers with plant breeding practice. Theor. Appl. Genet. 2015, 128, 779–795. [Google Scholar] [CrossRef]
- Xu, Y.; McCouch, S.R.; Zhang, Q. How can we use genomics to improve cereals with rice as a reference genome? Plant Mol. Biol. 2005, 59, 7–26. [Google Scholar] [CrossRef] [PubMed]
- Yi, M.; Than, K.; Vanavichit, A.; Chai-arree, W.; Toojinda, T. Marker assisted back cross breeding to improve cooking quality traits in Myanmar rice cultivar Manawthukha. Field Crop Res. 2009, 113, 178–186. [Google Scholar] [CrossRef]
- Jin, L.; Lu, Y.; Shao, Y.; Xiao, P.; Shen, S.; Corke, H.; Bao, J. Molecular marker assisted selection for improvement of the eating, cooking and sensory quality of rice (Oryza sativa L.). J. Cereal Sci. 2010, 51, 159–164. [Google Scholar] [CrossRef]
- Yu, J.; Hu, S.; Wang, J.; Wong, G.K.; Li, S.; Liu, B.; Deng, Y.; Dai, L.; Zhou, Y.; Zhang, X.; et al. A draft sequence of the rice genome (Oryza sativa L. ssp. indica). Science 2002, 296, 79–92. [Google Scholar] [CrossRef]
- Nadeem, M.A.; Nawaz, M.A.; Shahid, M.Q.; Dogan, Y.; Comertpay, G.; Yildiz, M.; Hatipoglu, R.; Ahmad, F.; Alsaleh, A.; Labhane, N.; et al. DNA molecular markers in plant breeding: Current status and recent advancements in genomic selection and genome editing. Biotechnol. Biotechnol. Equip. 2018, 32, 261–285. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Zhang, Q. The Rice Genome: Implications for Breeding Rice and Other Cereals. In New Directions for a Diverse Planet, Proceedings of the 4th International Crop Science Congress, Brisbane, Australia, 26 September–1 October 2014; Fischer, R.A., Ed.; Crop Science Society of America: Madison WI, USA, 2014. [Google Scholar]
- Jamali, S.H.; Cockram, J.; Hickey, L.T. Insights into deployment of DNA markers in plant variety protection and registration. Theor. Appl. Genet. 2018, 132, 1911–1929. [Google Scholar] [CrossRef]
- Hodgkin, T.; Roviglioni, R.; De Vicente, M.C.; Dudnik, N. Molecular methods in the conservation and use of plant genetic resources. Acta Hort. 2001, 546, 107–118. [Google Scholar] [CrossRef]
- Kaisoon, O.; Siriamornpun, S.; Meeso, N. Distinction between cereal genotypes based on the protein and DNA composition of the grain by capillary electrophoresis. World Appl. Sci. J. 2008, 4, 384–395. [Google Scholar]
- Salgotra, R.K.; Gupta, B.B.; Monika, S. Biotechnological interventions and their role in sustainable hill agriculture. J. Plant Sci. Res. 2015, 2, 1–8. [Google Scholar]
- Jiang, G.L. Molecular markers and marker-assisted breeding in plants. In Plant Breeding from Laboratories to Fields; Andersen, S.B., Ed.; InTech: Rijeka, Croatia, 2013; pp. 45–83. [Google Scholar]
- Singh, K.; Kumar, S.; Kumar, S.R.; Singh, M.; Gupta, K. Plant genetic resources management and pre-breeding in genomics era. Indian J. Genet. 2019, 79, 117–130. [Google Scholar] [CrossRef]
- Rai, V.P.; Singh, A.K.; Jaiswal, H.K. Evaluation of molecular markers linked to fragrance and genetic diversity in Indian aromatic rice. Turk. J. Bot. 2015, 39, 209–217. [Google Scholar] [CrossRef]
- Beaumont, M.A.; Balding, D.J. Identifying adaptive genetic divergence among populations from genome scans. Mol. Ecol. 2004, 13, 969–980. [Google Scholar] [CrossRef] [PubMed]
- Murphy, K.; Lammer, D.; Lyon, S.; Carter, B.; Jones, S.S. Breeding for organic and low-input farming systems: An evolutionary-participatory breeding method for inbred cereal grains. Renew. Agric. Food Syst. 2005, 20, 48–55. [Google Scholar] [CrossRef]
- Henry, R.J. (Ed.) Plant Genotyping—The DNA Fingerprinting of Plants; CABI Publishing: Wallingford, UK, 2001. [Google Scholar]
- Abdel-Mawgood, A.L. DNA Based techniques for studying genetic diversity. In Genetic Diversity in Microorganisms; IntechOpen Limited: London, UK, 2012; pp. 95–122. [Google Scholar]
- Loera-Sanchez, M.; Studer, B.; Kolliker, R. DNA-based assessment of genetic diversity in grassland plant species: Challenges, approaches, and applications. Agronomy 2019, 9, 881. [Google Scholar] [CrossRef] [Green Version]
- Tabor, H.K.; Risch, N.J.; Myers, R.M. Candidate-gene approaches for studying complex genetic traits: Practical considerations. Nat. Rev. Genet. 2002, 3, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, A.; Srivastava, S. Plant Breeding. In Participatory Plant Breeding: Concept and Applications; Springer: Singapore, 2019; pp. 29–68. [Google Scholar]
- Lubberstedt, T.; Zein, I.; Andersen, J.; Wenzel, G.; Krutzfeldt, B.; Eder, J.; Ouzunova, M.; Chun, S. Development and application of functional markers in maize. Euphytica 2005, 146, 101–108. [Google Scholar] [CrossRef]
- Lema, M. Marker assisted selection in comparison to conventional plant breeding. Agric. Res. Technol. 2018, 14, 555914. [Google Scholar] [CrossRef] [Green Version]
- Semagn, K.; Bjornstad, A.; Ndjiondjop, M.N. An overview of molecular marker methods for plants. Afr. J. Biotechnol. 2006, 2540, 25–68. [Google Scholar]
- Archak, S.; Lakshminarayanareddy, V.; Nagaraju, J. High-throughput multiplex microsatellite marker assay for detection and quantification of adulteration in basmati rice (Oryza sativa). Electrophoresis 2007, 28, 2396–2405. [Google Scholar] [CrossRef] [PubMed]
- Salgotra, R.K.; Millwood, R.J.; Agarwal, S.; Stewart, N.C. High-throughput functional marker assay for detection of Xa/xa and fgr genes in rice (Oryza sativa L.). Electrophoresis 2011, 32, 2216–2222. [Google Scholar] [CrossRef] [PubMed]
- Abdulmalik, R.O.; Menkir, A.; Meseka, S.; Unachukwu, N.; Ado, S.G.; Olarewaju, J.D.; Aba, D.A.; Hearne, S.; Crossa, J.; Gedil, M. Genetic gains in grain yield of a maize population improved through marker assisted recurrent selection under stress and non-stress conditions in West Africa. Front. Plant Sci. 2017, 8, 841. [Google Scholar] [CrossRef] [Green Version]
- Mullis, K.; Faloona, F.; Scharf, S.; Saiki, R.; Horn, G.; Erlich, H. Specific enzymatic amplification of DNA in vitro: The polymerase chain reaction. Cold Spring Harb. Symp. Quant. Biol. 1986, 51, 263–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iyer, A.S.; McCouch, S.R. Functional markers for xa5 mediated resistance in rice (Oryza sativa L.). Mol. Breed. 2007, 19, 291–296. [Google Scholar] [CrossRef]
- Chukwu, S.C.; Rafii, M.Y.; Ramlee, S.I.; Ismail, S.I.; Oladosu, Y.; Okporie, E.E.; Onyashi, G.; Utobo, E.; Ekwu, L.; Swaray, S.; et al. Marker-assisted selection and gene pyramiding for resistance to bacterial leaf blight disease of rice (Oryza sativa L.). Biotechnol. Biotechnol. Equip. 2019, 33, 440–455. [Google Scholar] [CrossRef] [Green Version]
- Azhaguvel, P.; Saraswathi, D.; Sharma, A.; Varshney, R.K. Methodological advancement in molecular markers to delimit the gene(s) for crop improvement. In Advances in Molecular Markers for Crop Improvement; Azhaguvel, P., Ed.; Global Science Books: Isleworth, UK, 2006; pp. 460–469. [Google Scholar]
- Meuwissen, T.H.E.; Hayes, B.J.; Goddard, M.E. Prediction of total genetic value using genome-wide dense marker maps. Genetics 2001, 157, 1819–1829. [Google Scholar] [PubMed]
- Saint Pierre, C.; Burgueno, J.; Crossa, J.; Davila, G.F.; Lopez, P.F.; Moya, E.S.; Moreno, I.I.; Muela, V.M.H.; Villa, V.M.H.; Vikram, P.; et al. Genomic prediction models for grain yield of spring bread wheat in diverse agro-ecological zones. Sci. Rep. 2016, 6, 27312. [Google Scholar] [CrossRef] [PubMed]
- Bhat, J.A.; Ali, S.; Salgotra, R.K.; Mir, Z.A.; Dutta, S.; Jadon, V.; Tyagi, A.; Mushtaq, M.; Jain, N.; Singh, P.K.; et al. Genomic selection in the era of next generation sequencing for complex traits in plant breeding. Front. Genet. 2016, 7, 221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crossa, J.; Beyene, Y.; Kassa, S.; Perez, P.; Hickey, J.M.; Chen, C.; de los Campos, G.; Burgueño, J.; Windhausen, V.S.; Buckler, E.; et al. Genomic prediction in maize breeding populations with genotyping-bysequencing. G3 (Bethesda) 2013, 3, 1903–1926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newell, M.A.; Jannink, J.L. Genomic selection in plant breeding. In Crop Breeding: Methods and Protocols; Fleury, D., Whitford, R., Eds.; Humana Press: New York, NY, USA, 2014; pp. 117–130. [Google Scholar]
- Lorenz, A.J.; Chao, S.; Asoro, F.G.; Heffner, E.L.; Hayashi, T.; Iwata, H.; Smith, K.P.; Sorrells, M.E.; LucJannink, J. Genomic selection in plant breeding: Knowledge and prospects. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: San Diego, CA, USA, 2011; Volume 110, pp. 77–123. [Google Scholar]
- Poland, J.A.; Brown, P.J.; Sorrells, M.E.; Jannink, J.L. Development of high-density genetic maps for barley and wheat using a novel two-enzyme genotyping-by-sequencing approach. PLoS ONE 2012, 7, e32253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorjanc, G.; Jenko, J.; Hearne, S.J.; Hickey, J.M. Initiating maize pre-breeding programs using genomic selection to harness polygenic variation from landrace populations. BMC Genom. 2016, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werner, C.R.; Voss-Fels, K.P.; Miller, C.N.; Quan, W.; Hua, W.; Guan, C.Y.; Snowdon, R.J.; Qian, L. Effective genomic selection in a narrow-genepool crop with low-density markers: Asian rapeseed as an example. Plant Genome 2018, 11, 170084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.B.; Yang, M.H.; Zeng, F.; Biligetu, B. Searching for an accurate marker-based prediction of an individual quantitative trait in molecular plant breeding. Front. Plant Sci. 2017, 8, 1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, T.G.; Andrew, T.; Cooper, G.M.; Margulies, E.H.; Mullikin, J.C.; Balding, D.J. Functional constraint and small insertions and deletions in the ENCODE regions of the human genome. Genome Biol. 2007, 8, R180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, E.; Chu, W.C.; Ayele, M.; Ho, J.; Bruggeman, E.; Yourstone, K.; Rafalski, A.; Smith, O.S.; McMullen, M.D.; Bezawada, C.; et al. Development of single nucleotide polymorphism (SNP) markers for use in commercial maize (Zea mays L.) germplasm. Mol. Breed. 2009, 24, 165–176. [Google Scholar] [CrossRef]
- Liu, Y.; He, Z.H.; Appels, R.; Xia, X.C. Functional markers in wheat: Current status and future prospects. Theor. Appl. Genet. 2012, 125, 1–10. [Google Scholar] [CrossRef]
- Jiang, Q.; Hou, J.; Hao, C.; Wang, L.; Ge, H.; Dong, Y.; Zhang, X. The wheat (T. aestivum) sucrose synthase 2 gene (TaSus2) active in endosperm development is associated with yield traits. Funct. Integr. Genom. 2011, 11, 49–61. [Google Scholar] [CrossRef]
- Yang, Z.; Bai, Z.; Li, X.; Wang, P.; Wu, Q.; Yang, L.; Li, L.; Li, X. SNP identification and allelic-specific PCR markers development for TaGW2, a gene linked to wheat kernel weight. Theor. Appl. Genet. 2012, 125, 1057–1068. [Google Scholar] [CrossRef]
- Zhang, Y.; Miao, X.; Xia, X.; He, Z. Cloning of seed dormancy gene (TaSdr) associated with tolerance to pre-harvest sprouting in common wheat and development of a functional marker. Theor. Appl. Genet. 2014, 127, 855–866. [Google Scholar] [CrossRef]
- Ellis, M.; Spielmeyer, W.; Gale, K.; Rebetzke, J.; Richards, A. “Perfect” markers for the Rht-B1b and Rht-D1b dwarfing genes in wheat. Theor. Appl. Genet. 2002, 105, 1038–1042. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Nakazaki, T.; Chen, S.; Chen, W.; Saito, H.; Tsukiyama, T.; Okumoto, Y.; Xu, Z.; Tanisaka, T. Identification and characterization of the erect-pose panicle gene EP conferring high grain yield in rice (Oryza sativa L.). Theor. Appl. Genet. 2009, 119, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Spielmeyer, W.; Ellis, M.; Robertson, M.; Ali, S.; Lenton, J.R.; Chandler, P.M. Isolation of gibberellin metabolic pathway genes from barley and comparative mapping in barley, wheat and rice. Theor. Appl. Genet. 2004, 109, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Raina, M.; Salgotra, R.K.; Pandotra, P.; Rathour, R.; Singh, K. Genetic enhancement for semi-dwarf and bacterial blight resistance with enhanced grain quality characteristics in traditional Basmati rice through marker-assisted selection. Comptes Rendus Biol. 2019, 342, 142–153. [Google Scholar] [CrossRef]
- Qi, Y.; Wang, L.; Gui, J.; Zhang, L.; Liu, Q.; Wang, J. Development and validation of a functional co-dominant SNP marker for the photoperiod thermo-sensitive genic male sterility pms3 (p/tms12-1) gene in rice. Breed. Sci. 2017, 67, 535–539. [Google Scholar] [CrossRef] [Green Version]
- Fu, D.; Szucs, P.; Yan, L.; Helguera, M.; Skinner, J.S.; von Zitzewitz, J.; Hayes, P.M.; Dubcovsky, J. Large deletions within the first intron in VRN-1 are associated with spring growth habit in barley and wheat. Mol. Genet. Genom. 2005, 273, 54–65. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Mirlohi, S.; Li, X.; Hea, Y. Identification of functional single-nucleotide polymorphisms affecting leaf hair number in Brassica rapa. Plant Physiol. 2018, 177, 490–503. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Yang, S.; Zhou, Y.; He, Z.; Xia, X. Distribution of the Rht-B1b, Rht-D1b and Rht8 reduced height genes in autumn-sown Chinese wheats detected by molecular markers. Euphytica 2006, 152, 109–116. [Google Scholar] [CrossRef]
- Su, Z.; Hao, C.; Wang, L.; Dong, Y.; Zhang, X. Identification and development of a functional marker of TaGW2 associated with grain weight in bread wheat (T. aestivum L.). Theor. Appl. Genet. 2011, 122, 211–223. [Google Scholar] [CrossRef]
- Andeden, E.; Yediay, F.; Baloch, F.; Shaaf, S.; Kilian, B.; Nachit, M.; Ozkan, H. Distribution of vernalization and photoperiod genes (Vrn-A1, Vrn-B1, Vrn-D1, Vrn-B3, Ppd-D1) in Turkish bread wheat cultivars and landraces. Cereal Res. Commun. 2011, 39, 352–364. [Google Scholar] [CrossRef]
- Feuillet, C.; Stein, N.; Rossini, L.; Praud, S.; Mayer, K.; Schulman, A.; Eversole, K.; Appels, R. Integrating cereal genomics to support innovation in the Triticeae. Funct. Integr. Genom. 2012, 12, 573–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, Y.Y.; Hong, L.Y.; Fei, T.J.; Qasim, S.M.; Zhi-Xiong, C.; Lan, W.; Jin-Quan, L.; Xiang-Dong, L.; Yong-Gen, L. Wide-compatibility gene S5n exploited by functional molecular markers and its effect on fertility of intersubspecific rice hybrids. Crop Sci. 2012. [Google Scholar] [CrossRef]
- Doebley, J.; Stec, A.; Gustus, C. Teosinte branched1 and the origin of maize: Evidence for epistasis and the evolution of dominance. Genetics 1995, 141, 333–346. [Google Scholar] [PubMed]
- Dunford, R.P.; Yano, M.; Kurata, N.; Sasaki, T.; Huestis, G.; Rocheford, T.; Laurie, D.A. Comparative mapping of the barley Phd-H1 photoperiod response gene region, which lies close to a junction between two rice linkage segments. Genetics 2002, 161, 825–834. [Google Scholar] [PubMed]
- Kole, C.; Quijada, P.; Michaels, S.D.; Amasino, R.M.; Osborn, T.C. Evidence for homology of flowering-time genes VFR2 from Brassica rapa and FLC from Arabidopsis thaliana. Theor. Appl. Genet. 2001, 102, 425–430. [Google Scholar] [CrossRef]
- Fileppi, M.; Gallsso, I.; Tagliabue, G.; Campion, B.; Doria, E.; Sparvoli, F. Characterisation of structural genes involved in phytic acid biosynthesis in common bean (Phaseolus vulgaris L.). Mol. Breed. 2010, 25, 453–470. [Google Scholar] [CrossRef]
- Hecht, V.; Foucher, F.; Ferrandiz, C.; Macknight, R.; Navarro, C.; Morin, J.; Vardy, M.E.; Ellis, N.; Beltran, J.P.; Rameau, C.; et al. Conservation of Arabidopsis flowering genes in model legumes. Plant Physiol. 2005, 137, 1420–1434. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Tang, F.; Gao, M.; Krishnan, H.B.; Zhu, H. R gene-controlled host specificity in the legume–rhizobia symbiosis. Proc. Natl. Acad. Sci. USA 2010, 107, 18735–18740. [Google Scholar] [CrossRef] [Green Version]
- Nesbitt, T.C.; Tanksley, S.D. Comparative sequencing in the genus Lycopersicon. Implications for the evolution of fruit size in the domestication of cultivated tomatoes. Genetics 2002, 162, 365–379. [Google Scholar]
- Fukino, N.; Ohara, T.; Sugiyama, M.; Kubo, N.; Hirai, M.; Sakata, Y.; Matsumoto, S. Mapping of a gene that confers short lateral branching (slb) in melon (Cucumis melo L.). Euphytica 2012, 187, 133–143. [Google Scholar] [CrossRef]
- Diaz, A.; Fergany, M.; Formisano, G.; Ziarsolo, P.; Blanca, J.; Fei, Z.; Staub, J.E.; Zalapa, J.E.; Cuevas, H.E.; Dace, G.; et al. A consensus linkage map for molecular markers and quantitative trait loci associated with economically important traits in melon (Cucumis melo L.). BMC Plant Biol. 2011, 11, 111. [Google Scholar] [CrossRef] [Green Version]
- Naresh, P.; Lin, S.; Lin, C.; Wang, Y.; Schafleitner, R.; Kilian, A.; Kumar, S. Molecular markers associated to two non-allelic genic male sterility genes in peppers (Capsicum annuum L.). Front. Plant Sci. 2018, 9, 1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nogales, A.; Nobre, T.; Gardoso, H.G.; Munoz-Sanhueza, L.; Valadas, V.; Campos, M.D.; Arnholdt-Schmitt, B. Allelic variation on DcAOX1 gene in carrot (Daucus carota L.): An interesting simple sequence repeat in a highly variable intron. Plant Gene 2016, 5, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Sheng, X.; Yu, H.; Wang, J.S.; Shen, Y.S.; Gu, H.H. Identification of QTLs associated with curd architecture in cauliflower. BMC Plant Biol. 2020, 20, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abuyusuf, M.; Nath, U.K.; Kim, H.; Islam, R.M.; Park, J.I.; Nou, I.S. Molecular markers based on sequence variation in BoFLC1.C9 for characterizing early- and late-flowering cabbage genotypes. BMC Genet. 2019, 20, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Zhuang, M.; Fang, Z.; Wang, Q.; Zhang, Y.; Liu, Y.; Yang, L.; Cheng, F. A co-dominant marker BoE332 applied to marker-assisted selection of homozygous male-sterile plants in cabbage (Brassica oleracea var. capitata L.). J. Integr. Agric. 2013, 12, 596–602. [Google Scholar] [CrossRef]
- Kim, S.; Jung, Y.; Seong, K.; Kim, S.C.; Jung, Y.H.; Kim, M.; Jang, K.C.; Song, E.Y.; Seong, K.C.; Um, Y.C. Development of a SCAR marker for sex identification in asparagus. Korean J. Plant Res. 2014, 27, 236–241. [Google Scholar] [CrossRef]
- Zhao, X.L.; Ma, E.W.; Gale, E.K.R.; Lei, Z.S.; He, Z.H.; Sun, Q.X.; Xia, X.C. Identification of SNPs and development of functional markers for LMW-GS genes at Glu-D3 and Glu-B3 loci in bread wheat (Triticum aestivum L.). Mol. Breed. 2007, 20, 223–231. [Google Scholar] [CrossRef]
- He, X.; Zhang, Y.; He, Z.; Wu, Y.P.; Xiao, Y.G.; Ma, C.X.; Xia, X.C. Characterization of phytoene synthase 1 gene (Psy1) located on common wheat chromosome 7A and development of a functional marker. Theor. Appl. Genet. 2008, 116, 213–221. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, X.; He, Z.; Ma, W.; Appels, R.; Pena, R.J.; Xia, X.C. Characterization of low molecular-weight glutenin subunit Glu-B3 genes and development of STS markers in common wheat (T. aestivum L.). Theor. Appl. Genet. 2009, 118, 525–539. [Google Scholar] [CrossRef]
- Zhang, C.; Dong, C.; He, X.; Zhang, L.; Xia, X.; Zhonghu, H. Allelic variants at the TaZds-D1 locus on wheat chromosome 2DL and their association with yellow pigment content. Crop Sci. 2011, 51, 1580–1590. [Google Scholar] [CrossRef]
- Geng, H.; Xia, X.; Zhang, L.; Qu, Y.; He, Z. Development of functional markers for Lipoxygenase gene Talox-B1 on chromosome 4 BS in common wheat. Crop Sci. 2012, 52, 568–576. [Google Scholar] [CrossRef]
- Shi, W.; Yang, Y.; Chen, S.; Xu, M. Discovery of a new fragrance allele and the development of functional markers for the breeding of fragrant rice varieties. Mol. Breed. 2008, 22, 185–192. [Google Scholar] [CrossRef]
- Amarawathi, Y.; Singh, R.; Singh, A.K.; Singh, V.P.; Mohapatra, T.; Sharma, T.R. Mapping of quantitative trait loci for basmati quality traits in rice (Oryza sativa L.). Mol. Breed. 2008, 21, 49–65. [Google Scholar] [CrossRef]
- Sakthivel, K.; Sundaram, R.M.; Rani, N.S.; Balachandran, S.M.; Neereja, C.N. Genetic and molecular basis of fragrance in rice. Biotechnol. Adv. 2009, 27, 468–473. [Google Scholar] [CrossRef]
- Chen, T.; Meng-xiang, T.; Zhang, Y.; Zhu, Z.; Zhao, L.; Zhao, Q.; Lin, J.; Zhou, L.; Wang, C. Development of simple functional markers for low glutelin content gene 1 (Lgc1) in rice (Oryza sativa). Rice Sci. 2010, 17, 173–178. [Google Scholar] [CrossRef]
- Anuradha, K.; Agarwal, S.; Rao, Y.V.; Rao, K.V.; Viraktamath, B.C.; Sarla, N. Mapping QTLs and candidate genes for iron and zinc concentrations in unpolished rice of Madhukar × Swarna RILs. Gene 2012, 508, 233–240. [Google Scholar] [CrossRef]
- Zhou, L.; Chen, S.; Yang, G.; Zha, W.; Cai, H.; Li, S.; Chen, Z.; Liu, K.; Xu, H.; You, A. A perfect functional marker for the gene of intermediate amylose content Wx-in in rice (Oryza sativa L.). Crop Breed. Appl. Biotechnol. 2018, 18, 103–109. [Google Scholar] [CrossRef] [Green Version]
- Chai, Y.; Hao, X.; Yang, X.; Allen, W.B.; Li, J.; Yan, J.; Shen, B.; Li, J. Validation of DGAT1-2 polymorphisms associated with oil content and development of functional markers for molecular breeding of high-oil maize. Mol. Breed. 2012, 29, 939–949. [Google Scholar] [CrossRef]
- Zhou, Y.; Han, Y.; Li, Z.; Fu, Y.; Fu, Z.; Xu, S.; Li, J.; Yan, J.; Jang, X. ZmcrtRB3 encodes a carotenoid hydroxylase that affects the accumulation of a-carotene in maize kernel. J. Integr. Plant Biol. 2012, 54, 260–269. [Google Scholar] [CrossRef]
- Chhabra, R.; Hossain, F.; Muthusamy, V.; Baveja, A.; Mehta, B.K.; Zunjare, R.U. Development and validation of breeder-friendly functional markers of sugary1 gene encoding starch-debranching enzyme affecting kernel sweetness in maize (Zea mays). Crop Pasture Sci. 2019, 70, 868–875. [Google Scholar] [CrossRef]
- Obeng-Bio, E.; Badu-Apraku, B.; Elorhor Ifie, B.; Danquah, A.; Blay, E.T.; Dadzie, M.A. Phenotypic characterization and validation of provitamin A functional genes in early maturing provitamin A-quality protein maize (Zea mays) inbred lines. Plant Breed. 2019. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Nie, Y.D.; Han, F.X.; Zhao, X.N.; Dun, B.Q.; Lu, M.; Li, G. Allelic variation of a soluble acid invertase gene (SAI-1) and development of a functional marker in sweet sorghum (Sorghum bicolor (L.). Mol. Breed. 2014, 33, 721–730. [Google Scholar] [CrossRef]
- Zanan, R.; Khandagale, K.; Hinge, V.; Elangovan, M.; Henry, R.J.; Nadaf, A. Characterization of fragrance in sorghum (Sorghum bicolor L.) Moench) grain and development of a gene-based marker for selection in breeding. Mol. Breed. 2016, 36. [Google Scholar] [CrossRef]
- Juwattanasomran, R.; Somta, P.; Kaga, A.; Chankaew, S.; Shimizu, T.; Sorajjapinun, W.; Srinives, P. Identification of a new fragrance allele in soybean and development of its functional marker. Mol. Breed. 2010. [Google Scholar] [CrossRef]
- Cuevas, H.E.; Staub, J.E.; Simon, P.W.; Zalapa, J.E. A consensus linkage map identifies genomic regions controlling fruit maturity and beta-carotene-associated flesh color in melon (Cucumis melo L.). Theor. Appl. Genet. 2009, 119, 741–756. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; He, L.; Xu, S.; Wan, Y.; Wang, H.; Wang, Y.; Yu, L.; Zhu, W. Expression analysis, functional marker development and verification of AgFNSI in celery. Sci. Rep. 2020, 10, 531. [Google Scholar] [CrossRef]
- Cheng, T.; Zheng, X.; Xie, K.; Liu, J.; Zheng, X.; Jin, S.; Diao, Y.; Hu, Z.; Wang, J. Development and identification of three functional markers associated with starch content in lotus (Nelumbo nucifera). Sci. Rep. 2020, 10, 4242. [Google Scholar] [CrossRef]
- Saritnum, O.; Minami, M.; Matsushima, K.; Minamiyama, Y.; Hirai, M.; Baba, T.; Bansho, H.; Nemoto, K. Inheritance of few-pungent trait in chilli pepper ‘S3212’ (Capsicum frutescens). J. Jpn. Soc. Hort. Sci. 2008, 77, 265–269. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, N.; Torres, A.M. Characterization and diagnostic marker for TTG1 regulating tannin and anthocyanin biosynthesis in faba bean. Sci. Rep. 2019, 9, 16174. [Google Scholar] [CrossRef] [Green Version]
- Gemenet, D.C.; Kitavi, M.N.; David, M. Development of diagnostic SNP markers for quality assurance and control in sweetpotato [Ipomoea batatas (L.) Lam.] breeding programs. PLoS ONE 2020, 15, e0232173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arizio, C.M.; Costa Tártara, S.M.; Manifesto, M.M. Carotenoids gene markers for sweetpotato (Ipomoea batatas L. Lam): Applications in genetic mapping, diversity evaluation and cross-species transference. Mol. Genet. Genom. 2014. [Google Scholar] [CrossRef]
- Tommasini, L.; Yahiaoui, N.; Srichumpa, P.; Keller, B. Development of functional markers specific for seven Pm3 resistance alleles and their validation in the bread wheat gene pool. Theor. Appl. Genet. 2006, 114, 165–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Periyannan, S.; Bansal, U.; Bariana, H.; Deal, K.; Luo, M.; Dvorak, J.; Lagudah, E. Identification of a robust molecular marker for the detection of the stem rust resistance gene Sr45 in common wheat. Theor. Appl. Genet. 2014, 127, 947–955. [Google Scholar] [CrossRef]
- Hur, Y.J.; Jeung, J.; Kim, S.Y.; Park, H.; Cho, J.; Lee, J.Y.; Sohan, Y.; Song, Y.C.; Park, D.; Lee, C.; et al. Functional markers for bacterial blight resistance gene Xa3 in rice. Mol. Breed. 2013, 31, 981–985. [Google Scholar] [CrossRef]
- Hajira, S.K.; Sundaram, R.M.; Laha, G.S.; Yugander, A.; Balachandran, S.M.; Viraktamath, B.C.; Sujatha, K.; Balachiranjeevi, C.H.; Pranathi, K.; Anila, M.; et al. A single-tube, functional marker-based multiplex PCR assay for simultaneous detection of major bacterial blight resistance genes Xa21, xa13 and xa5 in rice. Rice Sci. 2016, 23, 144–151. [Google Scholar] [CrossRef] [Green Version]
- Sundaram, R.M.; Vishnupriya, M.R.; Biradar, S.K.; Laha, G.S.; Reddy, G.A.; Shobha, R.N.; Sarma, N.P.; Sonti, R.V. Marker-assisted introgression of bacterial blight resistance in Samba Mahsuri, an elite indica rice variety. Euphytica 2008, 80, 411–422. [Google Scholar] [CrossRef]
- Song, W.Y.; Pi, L.Y.; Wang, G.L.; Gardner, J.; Holsten, T.; Ronald, P.C. Evolution of the rice Xa21 disease resistance gene family. Plant Cell 1997, 9, 1279–1287. [Google Scholar]
- Ellur, R.K.; Khanna, A.; Gopala Krishnan, S.; Bhowmick, P.K.; Vinod, K.K.; Nagarajan, M.; Mondal, K.K.; Singh, N.K.; Singh, K.; Prabhu, K.B.; et al. Marker-aided incorporation of Xa38, a novel bacterial blight resistance gene, in PB1121 and comparison of its resistance spectrum with xa13 + Xa21. Sci. Rep. 2016, 6, 29188. [Google Scholar] [CrossRef]
- Zhou, L.; Chen, Z.; Lang, X.; Du, B.; Liu, K.; Yang, G.; Hu, G.; Li, S.; He, G.; You, A. Development and validation of a PCR-based functional marker system for the brown planthopper resistance gene Bph14 in rice. Breed. Sci. 2013, 63, 347–352. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, K.; Yasuda, N.; Fujita, Y.; Koizumi, S.; Yoshida, H. Identification of the blast resistance gene Pit in rice cultivars using functional markers. Theor. Appl. Genet. 2010, 121, 1357–1367. [Google Scholar] [CrossRef] [PubMed]
- Ramkumar, G.; Srinivasarao, K.; Mohan, K.M.; Sudarshan, I.; Sivaranjani, A.K.P.; Gopalkrishna, K.; Neeraja, C.N.; Balachandran, S.M.; Sundaram, S.M.; Prasad, M.S.; et al. Development and validation of functional marker targeting an InDel in the major rice blast disease resistance gene Pi54 (Pikh). Mol. Breed. 2011, 27, 129–135. [Google Scholar]
- Ma, J.; Ma, X.D.; Zhao, Z.C.; Wang, S.; Wang, J.L.; Wang, J.; Cheng, Z.J.; Lei, C.L. Development and application of a functional marker of the blast resistance gene Pi35 in rice. Acta Agron. Sin. 2015, 41, 1779–1790. [Google Scholar]
- Quan, W.F.; Hui, C.Z.; Yang, X.U.; Wang, J.; Li, W.; Fan, F.; Chen, L.; Tao, Y.; Zhong, W.; Yang, J. Development and application of the functional marker for the broad-spectrum blast resistance gene PigmR in rice. Sci. Agric. Sin. 2019, 52, 955–967. [Google Scholar]
- Wang, Y.; Jiang, W.; Liu, H.; Zeng, Y.; Du, B.; Zhu, L.; He, G.; Chen, R. Marker assisted pyramiding of Bph6 and Bph9 into elite restorer line 93–11 and development of functional marker for Bph9. Rice 2017, 10, 51. [Google Scholar] [CrossRef] [Green Version]
- Madsen, L.H.; Collins, N.C.; Rakwalska, M.; Backes, G.; Sandal, N.; Krusell, L.; Jensen, J.; Waterman, E.H.; Jahoor, A.; Ayliffe, M. Barley disease resistance gene analogs of the NBSLRR class: Identification and mapping. Mol. Genet. Genom. 2003, 269, 150–161. [Google Scholar] [CrossRef]
- Singh, K.B.; Foley, R.C.; Sanchez, L.O. Transcription factors in plant defense and stress responses. Curr. Opin. Plant Biol. 2002, 5, 430–436. [Google Scholar] [CrossRef]
- Brunner, S.; Keller, B.; Feuillet, C. A large rearrangement involving genes and low copy DNA interrupts the micro-collinearity between rice and barley at the Rph7 locus. Genetics 2003, 164, 673–683. [Google Scholar] [PubMed]
- Nevame, A.Y.M.; Xia, L.; Nchongboh, C.G.; Hasan, M.M.; Alam, M.A.; Yongbo, L.; Wenting, Z.; Yafei, H.; Emon, R.M.; Ismail, M.R.; et al. Development of a new molecular marker for the resistance to tomato yellow leaf curl virus. BioMed Res. Int. 2018. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.; Hwang, I.S.; Lee, H.J.; Lee, J.M.; Seo, E.; Choi, D.; Oh, C. Identification of a molecular marker tightly linked to bacterial wilt resistance in tomato by genome-wide SNP analysis. Theor. Appl. Genet. 2018, 131, 1017–1030. [Google Scholar] [CrossRef]
- Devran, Z.; Kahveci, E.; Hong, Y.; Studholme, D.J.; Tör, M. Identifying molecular markers suitable for Frl selection in tomato breeding. Theor. Appl. Genet. 2018, 131, 2099–2105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oumouloud, A.; Otmani, M.E.; Alvarez, J. Molecular characterization of Fom-1 gene and development of functional markers for molecular breeding of resistance to Fusarium race 2 in melon. Euphytica 2015. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, A.F.; Hoy, J.W.; Kinbeng, C.A.; Baisakh, N. Identification of genomic regions controlling leaf scald resistance in sugarcane using a bi-parental mapping population and selective genotyping by sequencing. Front. Plant Sci. 2018, 9, 877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Ren, Y.; Guo, S.; Zhang, H.; Gong, G.; Du, Y.; Xu, Y. Application of comparative genomics in developing markers tightly linked to the Pm-2F gene for powdery mildew resistance in melon (Cucumis melo L.). Euphytica 2012, 190, 157–168. [Google Scholar] [CrossRef]
- Nieto, C.; Morales, M.; Orjeda, G.; Clepet, C.; Monfort, A.; Sturbois, B.; Puigdomenech, P.; Pitrat, M.; Caboche, M.; Dogimont, C.; et al. An eIF4E allele confers resistance to an uncapped and non-polyadenylated RNA virus in melon. Plant J. 2006, 48, 452–462. [Google Scholar] [CrossRef]
- Ling, K.S.; Harris, K.R.; Meyer, J.D.F.; Levi, A.; Guner, N.; Wehner, T.C.; Bendahmane, A.; Havey, M.J. Non-synonymous single nucleotide polymorphisms in the watermelon eIF4E gene are closely associated with resistance to Zucchini yellow mosaic virus. Theor. Appl. Genet. 2009, 120, 191–200. [Google Scholar] [CrossRef]
- Gama, R.N.C.; Santos, C.A.F.; Dias, R.C.S.; Souza, R.R.C.; Queiróz, M.A. Microsatellite markers linked to powdery mildew resistance locus in watermelon. Aust. J. Crop Sci. 2015, 9, 92–97. [Google Scholar]
- Liu, P.N.; Miao, H.; Lu, H.W.; Cui, J.Y.; Tian, G.L.; Wehner, T.C.; Gu, X.F.; Zhang, S.P. Molecular mapping and candidate gene analysis for resistance to powdery mildew in Cucumis sativus stem. Genet. Mol. Res. 2017, 16, 16039680. [Google Scholar] [CrossRef]
- Shi, L.; Yang, Y.; Xie, Q.; Miao, H.; Bo, K.; Song, Z.; Wang, Y.; Xie, B.; Zhang, S.; Gu, X. Inheritance and QTL mapping of cucumber mosaic virus resistance in cucumber. PLoS ONE 2018, 13, e0200571. [Google Scholar] [CrossRef]
- Cao, B.H.; Lei, J.J.; Wang, Y.; Chen, G.J. Inheritance and identification of SCAR marker linked to bacterial wilt-resistance in eggplant. Afr. J. Biotechnol. 2009, 8, 5201–5207. [Google Scholar]
- Karna, S.; Ahn, Y. K Development of InDel markers to identify Capsicum disease resistance using whole genome resequencing. J. Plant Biotechnol. 2018, 45, 228–235. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Lu, H.; Hu, J. Molecular mapping to bacterial leaf spot in lettuce PI358001-1. Genet. Resist. 2016. [Google Scholar] [CrossRef]
- Li, G.; Qian, W.; Zhang, S.; Zhang, S.; Li, F.; Zhang, H.; Wu, J.; Wang, X.; Sun, R. Development of gene-based markers for the Turnip mosaic virus resistance gene retr02 in Brassica rapa. Plant Breed. 2016, 135. [Google Scholar] [CrossRef]
- Sun, S.; Deng, D.; Wang, Z.; Duan, C.; Wu, X.; Wang, X.; Zong, X.; Zhu, Z. A novel er1 allele and the development and validation of its functional marker for breeding pea (Pisum sativum L.) resistance to powdery mildew. Theor. Appl. Genet. 2016, 129, 909–919. [Google Scholar] [CrossRef]
- Maiti, S.; Basak, J.; Kundagrami, S.; Kundu, A.; Pal, A. Molecular marker-assisted genotyping of mungbean yellow mosaic India virus resistant germplasms of mungbean and urdbean. Mol. Biotechnol. 2011, 47, 95–104. [Google Scholar] [CrossRef]
- Lan, N.T.N.; Thao, H.T.; Son, L.V.; Mau, C.H. Overexpression of VrPDF1 gene confers resistance to weevils in transgenic mung bean plants. PeerJ Prepr. 2017. [Google Scholar] [CrossRef]
- Garg, B.; Lata, C.; Prasad, M. A study of the role of gene TaMYB2 and an associated SNP in dehydration tolerance in common wheat. Mol. Biol. Rep. 2012, 39, 10865–10871. [Google Scholar] [CrossRef]
- Pandey, B.; Sharma, P.; Pandey, D.; Sharma, I.; Chatrath, R. Identification of new aquaporin genes and single nucleotide polymorphism in bread wheat. Evol. Bioinform. 2013, 9, 437–452. [Google Scholar] [CrossRef]
- Huseynova, I.M. Application of PCR-based functional markers for identification of DREB1 genes in Triticum aestivum L. SF Biotechnol. Bioeng. J. 2018, 1, 1. [Google Scholar]
- Hamdi, K.; Brini, F.; Kharrat, N.; Masmoudi, K.; Yakoubi, I. Abscisic acid, stress, and ripening (TtASR1) gene as a functional marker for salt tolerance in durum wheat. BioMed J. Int. 2020. [Google Scholar] [CrossRef] [Green Version]
- Neeraja, C.; Maghirang-Rodriguez, R.; Pamplona, A.; Heuer, S.; Collard, B.C.Y.; Septiningsih, E.M.; Vergara, G.; Sanchez, D.; Xu, K.; Ismail, A.M.; et al. A marker-assisted backcross approach for developing submergence tolerant rice cultivars. Theor. Appl. Genet. 2007, 115, 767–776. [Google Scholar] [CrossRef] [PubMed]
- Chin, J.H.; Gamuyao, R.; Dalid, C.; Bustamam, M.; Prasetiyono, J.; Moeljopawiro, S.; Wissuwa, M.; Heuer, S. Developing rice with high yield under phosphorus deficiency: Pup1 sequence to application. Plant Physiol. 2011, 156, 1202–1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, D.; Wang, H.; Liang, G.; Yu, D. OsSAPK2 Confers abscisic acid sensitivity and tolerance to drought stress in rice. Front. Plant Sci. 2017, 8, 993. [Google Scholar] [CrossRef] [Green Version]
- Assenov, B.; Andjelkovic, V.; Ignjatovic-Micic, D.; Pagnotta, M.A. Identification of SNP mutations in In MYBF-1 gene involved in drought stress tolerance in maize. Bulg. J. Agric. Sci. 2013, 19, 181–185. [Google Scholar]
- Too, E.J.; Onkware, A.O.; Were, B.A.; Gudu, S.; Carlsson, A.; Geleta, M. Molecular markers associated with aluminium tolerance in Sorghum bicolor. Hereditas 2018, 155, 20. [Google Scholar] [CrossRef]
- Contour-Ansel, D.; Torres-Franklin, M.L.; Cruz, D.E.; Lameta, A.; Fodil, F.Z. Glutathione reductase in leaves of cowpea: Cloning of two cDNAs, expression and enzymatic activity under progressive drought stress desiccation and abscisic acid treatment. Ann. Bot. 2006, 98, 1279–1287. [Google Scholar] [CrossRef] [Green Version]
- Lata, C.; Prasad, M. Validation of an allele-specific marker associated with dehydration stress tolerance in a core set of foxtail millet accessions. Plant Breed. 2013, 132, 496–499. [Google Scholar] [CrossRef]
- Leonforte, A.; Sudheesh, S.; Cogan, N.O.; Salisbury, P.A.; Nicolas, M.E.; Materne, M.; Forster, J.W.; Kaur, S. SNP marker discovery, linkage map construction and identification of QTLs for enhanced salinity tolerance in field pea (Pisum sativum L.). BMC Plant Biol. 2013, 13, 161. [Google Scholar] [CrossRef] [Green Version]
- Ramkumar, G.; Sivaranjani, A.; Pandey, M.K.; Sakthivel, K.; Shobha, N.R.; Sudarshan, I.; Prasad, G.S.V.; Neeraja, C.N.; Sundaram, R.M.; Viraktamath, B.C.; et al. Development of a PCR-based SNP marker system for effective selection of kernel length and kernel elongation in rice. Mol. Breed. 2010, 26, 735–740. [Google Scholar] [CrossRef]
- Randhawa, M.S.; Bains, N.S.; Sohu, V.S.; Chhuneja, P.; Trethwon, R.M.; Bariana, H.S.; Banasl, U. Marker assisted transfer of stripe rust and stem rust resistance genes into four wheat cultivars. Agronomy 2019, 9, 497. [Google Scholar] [CrossRef] [Green Version]
- Lagudah, E.S.; Krattinger, S.G.; Herrera-Foessel, S.; Singh, R.P.; Huerta-Espino, J.; Spielmeyer, W.; Brown-Guedira, G.; Selter, L.L.; Keller, B. Gene specific markers for the wheat gene Lr34/Yr18/Pm38 which confers resistance to multiple fungal pathogens. Theor. Appl. Genet. 2009, 119, 889–898. [Google Scholar] [CrossRef] [Green Version]
- Yugander, A.; Sundaram, R.M.; Singh, K.; Ladhalakshmi, D.; Subba Rao, L.V.; Madhav, M.S.; Badri, J.; Prasad, M.S.; Laha, G.S. Incorporation of the novel bacterial blight resistance gene Xa38 into the genetic background of elite rice variety ‘Improved Samba Mahsuri’. PLoS ONE 2018, 13, e0198260. [Google Scholar] [CrossRef]
- Joseph, M.; Gopalakrishnan, S.; Sharma, R.K.; Singh, V.P.; Singh, A.K.; Singh, N.K.; Mohapatra, T. Combining bacterial blight resistance and Basmati quality characteristics by phenotypic and molecular marker-assisted selection in rice. Mol. Breed. 2004, 13, 377–387. [Google Scholar] [CrossRef]
- Das, G.; Rao, G.J.N.; Varier, M.; Prakash, A.; Prasad, D. Improved Tapaswini having four BB resistance genes pyramided with six genes/QTLs, resistance/ tolerance to biotic and abiotic stresses in rice. Sci. Rep. 2018, 8, 2413. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Functional markers (FMs) are functionally linked with phenotypic traits compared to random DNA markers (RDMs) and genic molecular markers (GMMs).
Figure 1.
Functional markers (FMs) are functionally linked with phenotypic traits compared to random DNA markers (RDMs) and genic molecular markers (GMMs).

Figure 2.
Different approaches involved in development of functional markers (FMs).

Figure 3.
Genome-wide association studies (GWAS) for identification of functional single nucleotide polymorphisms (SNPs) and development of FMs.
Figure 3.
Genome-wide association studies (GWAS) for identification of functional single nucleotide polymorphisms (SNPs) and development of FMs.

Figure 4.
Use of functional markers (FMs) in marker assisted selection in precision breeding. (a) Phenotypic selection for disease resistance plants. (b) Selection of disease resistance using genic molecular markers (GMM). (c) Disease resistance plants selection using functional marker (FM).
Figure 4.
Use of functional markers (FMs) in marker assisted selection in precision breeding. (a) Phenotypic selection for disease resistance plants. (b) Selection of disease resistance using genic molecular markers (GMM). (c) Disease resistance plants selection using functional marker (FM).

Figure 5.
Pyramiding of desired genes in a cultivar using functional markers (FMs).


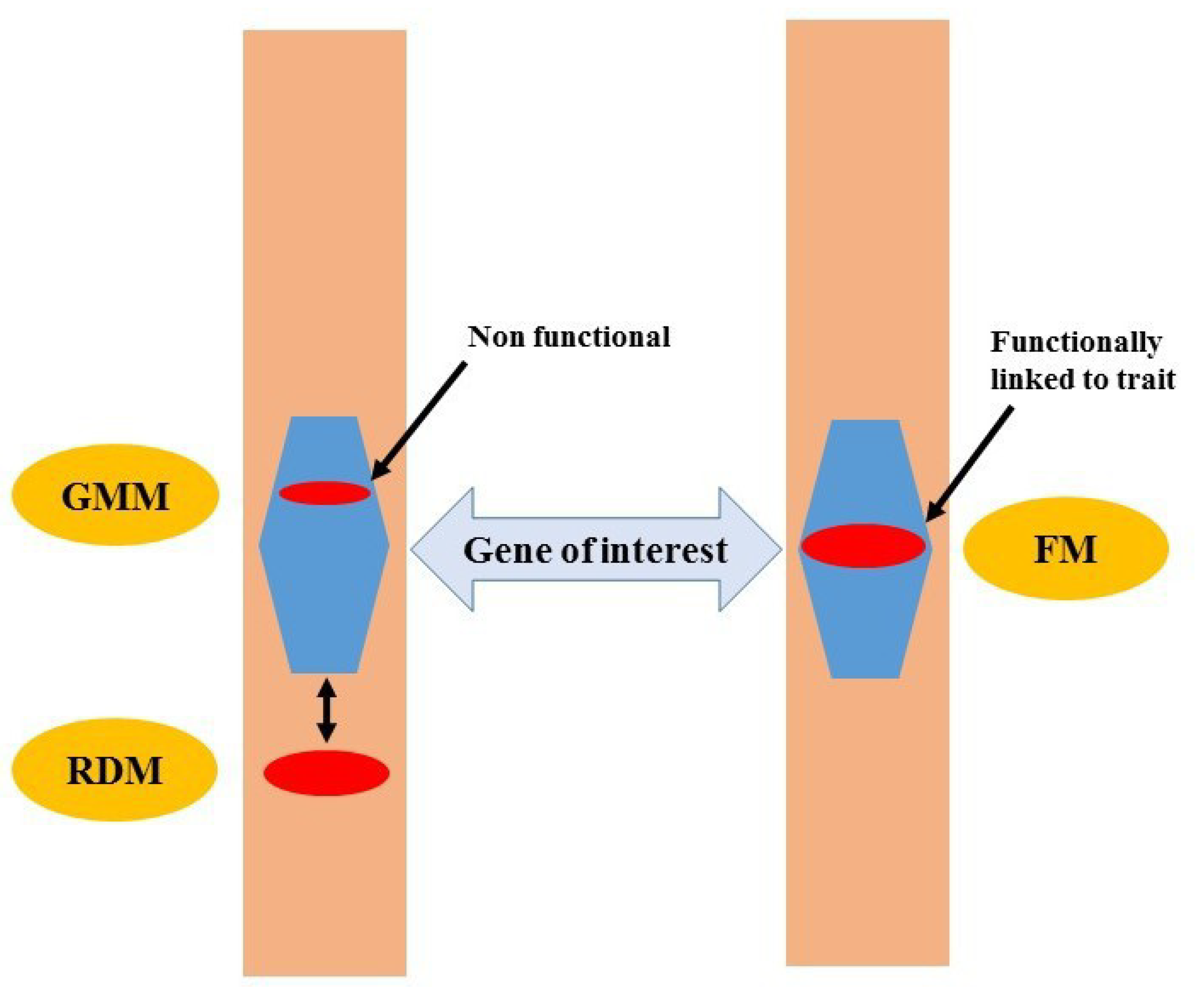{kind=link}
{kind=link}
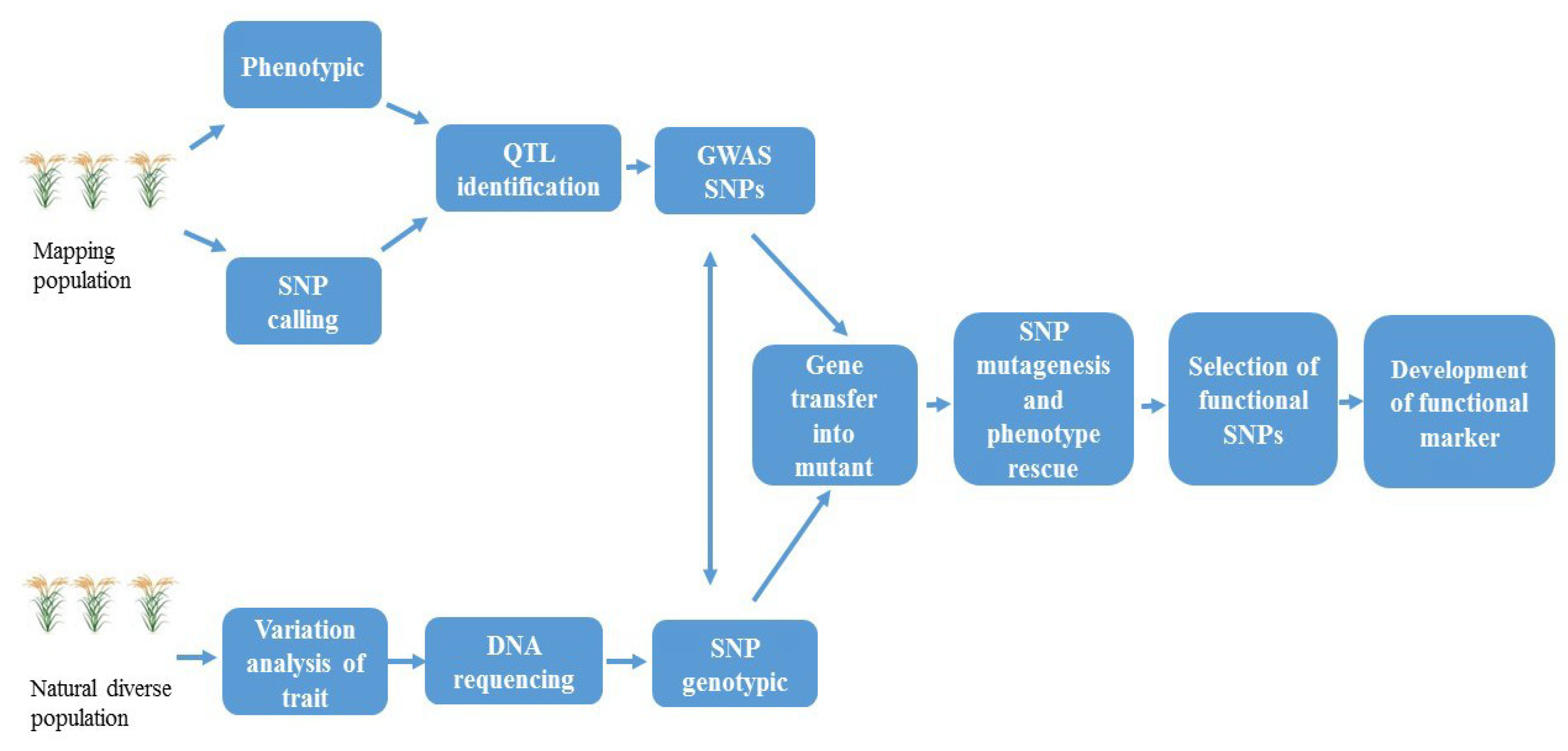{kind=link}
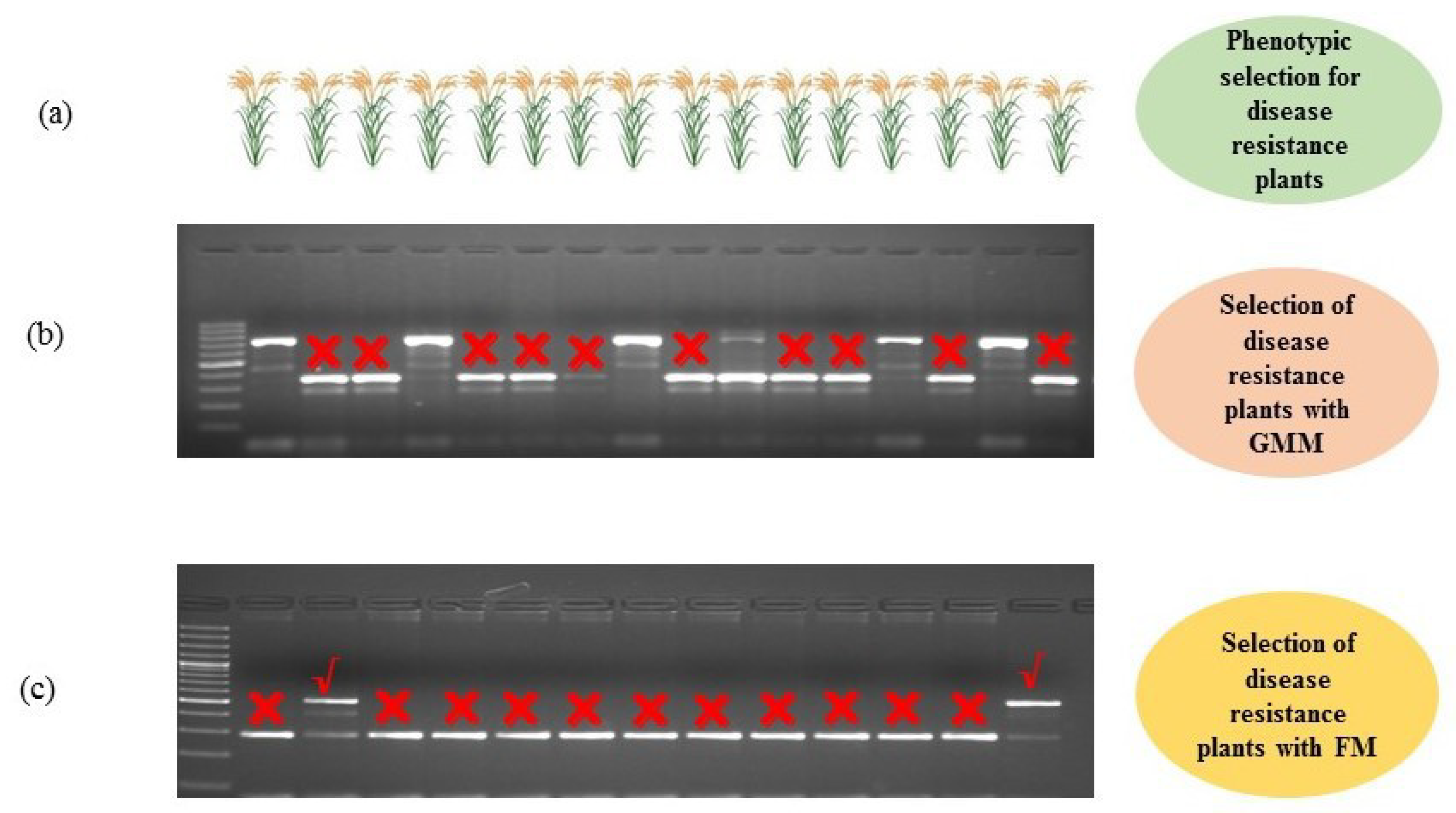{kind=link}
{kind=link}
Table 1.
Candidate genes for functional markers (FMs) development for different traits.
| Trait | Gene (s) | Chromosomal Location | Sequence | Crop | References |
|---|---|---|---|---|---|
| Agronomic traits | |||||
| Semi-dwarf stature | Rht-B1 and Rht-D1 | 4B, 4D | F-TCTCCTCCCTCCCCACCCCAAC R-CCATGGCCATCTCGAGCTGC & F-CGCGCAATTATTGGCCAGAGATAG R-CCCCATGGCCATCTCGAGCTGCTA | Wheat | [129] |
| Grain weight | TaSus2-2B | 2 | F-CGCCCTGAGCCG CATCCACA R-CGCTCGCCCGC CATTTATTTCTCT | Wheat | [118] |
| Grain weight | TaGW2 | 6 | F-ATGGGGAACAGAATAGGAGGGAGGA R-CGAGTATGCCTAGAATGGAAAGAC | Wheat | [130] |
| Photoperiod response | Phd-H1 | 2 | F-ACGCCTCCCACTACACTG R-CACTGGTGGTAGCTGAGATT | Wheat | [131] |
| Vernalization | Vrn-D4 | 5 | F-CATAATGCCAAGCCGGTGAGTAC R-ATGTCTGCCAATTAGCTAGC | Wheat | [132] |
| Semi-dwarf | sd1 | 1 | F-CACGCACGGGTTCTTCCAGGTG R-AGGAGAATAGGAGATGGTTTACC | Rice | [123] |
| Wide-compatibility gene | S5n | 6 | F-CGTCTTGCTTCTTCATTCCC R-GTAGGTAAACACAGGCAGAG | Rice | [133] |
| Photoperiod-thermo-sensitive genic male (PGMS and TGMS) sterility | pms3 (p/tms12-1) | 12 | F-GAATGCCATCTAAACACT R-ATTTTACTCTTGATGGATGGTC | Rice | [126] |
| Plant stature | tb1 | 1 | F-CACATGAGCCCATGCCTCTC R-AAAGCGGTAAGTCCATGGGG | Maize | [134] |
| Plant height | Dwarf8 | 1 | F-ACACTATCACCGCTCTATTG R-ACTCTTTCCCTGACTTCATT | Maize | [31] |
| Photoperiod response | Phd-H1 | 7 | F-CCTCTTCGCTATTAC GCCAG R –GCCCTTCCCAACAGTTGCG | Barley | [135] |
| Vernalization requirements | VRN-H 1 | 5 | F-TTCATCATGGATCGCCAGTA R-AAAGCTCCTGCCAACTACGA | Barley | [127] |
| Vernalization requirements | VFR2 | A8 | F-CTCGTAGCCCCGAGAACATC R-ACTCAAGCAACTTACCAAGTGGA | Brassica | [136] |
| Leaf hair number | BrpHL1 | A9 | F-TACTCCTCGTTCCCTCTGGG R-GGGGGAAATGCAGATTCCGA | Brassica | [128] |
| Seed development | PvMIPSs and PvMIPSv | 1 | F-TTGCCACGCACCTGCTAATA R-CCTGCAGCTGCGATTTTCAA | Common bean | [137] |
| Controlling flowering time | MADS-box, Constants and Flowering locus T/Terminal Flower1 | 16 | F-ATGCACCTAGCCCAAGTGAC R-TGTTTGCATTCATGGCGTGT & R-ATCTGTTGTGCCGGGAATGT F-AACCGAAATGCAAAACAGGTGA | Pea, soybean, and burclover | [138] |
| Nodulation formation | Rj2 and Rfg1 | 16, 3 | F-AAGTCTTAAATTGTGTTTGGATGGA R-TGAGAATTGTCACCACCGGG & F-AAGTCTTAAATTGTGTTTGGATGGA R-TGAGAATTGTCACCACCGGG | Soybean | [139] |
| Fruit size | w2.2 | 2 | F-TCTGCTCAGAAGCATGCACA R-TTGTGACCTGTACCCCAGGA | Tomato | [140] |
| Short lateral branching | slb | 11 | F-CTTGCGCTCCTTGGTATTCC R-CAAGATCGGCAAGAGACAGC | Melon | [141] |
| Sutures on the rind | s-2 | 9 | F-GCATCGGAATCTTGTTCGGC R-TCCGGTGGGAGATACCCAAT | Melon | [142] |
| Male sterility | ms3 | 5 | F-GGTACTTTGA CCCTCATAATTGG R-TTGTTTGT GGTGTACG TGCT | Capsicum | [143] |
| Alternative respiration | DcAOX1 | 1 | F-AAAATAACAATGATGATGACACG R-CTCCACTTCAGTGATATCCAA | Carrot | [144] |
| Curd architecture | qCS.C6–1 and qCS.C6–2 | 6C | F-CGGTACTGGAATGTGGACGT R-TGAATTGGTATGAACACGCCTC | Cauliflower | [145] |
| Early and late flowering | BoFLC1.C9 | Unmapped | F-GGAAAGCAACATGGTGATGA R-CATGGTGTGAACCAGAGTCC | Cabbage | [146] |
| Male sterility | CDMs399-3 | 7C | F-TCCCTTTCACATCGTCCACA R-TGCAGCCCAGAACAGTGATA | Cabbage | [147] |
| Sex identification | MYB35 | 5 | F-TTGCTTGGCGGATCATATTATG R-TTGCTTGGCGATGTCCCTTTTG | Asparagus | [148] |
| Quality traits | |||||
| Low molecular weight glutenin | Glu-D3 and Glu-B3 | 1D | F-CAGCTAAACCCATGCAAGC R-CAATGGAAGTCATCACCTCAA | Wheat | [149] |
| Yellow pigment content | Psy1 | 7A | F-ACATGCCGCTACTCCTATCC R-GTAGAGTGGCCAGACAAGGT | Wheat | [150] |
| Low molecular weight glutenin | Glu-B3 | 1B | F-ACAACAGGTTCAGGGTTCCA R-GCTATTTGGTGTGGCTGCAA | Wheat | [151] |
| Yellow pigment content | TaZds-D1 | 2D | F-ACATAGTCCTGACCGCCAAA R-AGAGTTGCTCCTTCCATGCT | Wheat | [152] |
| Lipoxygenase gene | Talox-B1 | 4B | F-ATGATACTGGGCGGGCTCGT R-TCAGATGGAGATGCTGTTGGG | Wheat | [153] |
| Fragrance | badh2 | 8 | F-AGTTATGGTCTGGCTGGTGC R-TTGTGTGCTACCCACCCTTC | Rice | [154] |
| Fragrance | nksbad2 | 4 | F-ATGGCAACATGGAAGGTAGC R-CATCAGCAAGCTCCAAACAA | Rice | [155] |
| Fragrance | BADEX7-5 | 8 | F-TTAGGTTCTGAAGCCGGTGC R-TCCCAGTAAATGCAACCTAACAGA | Rice | [156] |
| Low glutenin content | Lgc1 | 10 | F-TTCTACAATGAAGGCGATGC R-CTGGGCTTTAACGGGACT & F-ACCGTGTTATGGCAGTTT R-ATTCAAGGGCTATCGTCT | Rice | [157] |
| Fe and Zn | OsNAS3, OsNRAMP1 | 7 | F-TCCATCGCTTGCTACCTCAC R-CCCGGAGATCGATCGAGACA & F-AGCACTCCCCCATCAATCAA R-ACTACACGGGTGGCTCTTTG | Rice | [158] |
| Intermediate amylose content | Wx-in | 6 | F-CAGCGTCGACGTAAGCCTAT R-CAGGCCCCTGAAATCCATGT | Rice | [159] |
| Oil content | DGAT1-2 | 6 | F-TGGCTCTGCAATCAGGAGAA R-TGAAGCAGCAAACAACGAGC | Maize | [160] |
| Forage quality for digestibility | Bm3 | 4 | F-TTCAACAAGGCGTACGGGAT R-AGTGGTTCTTCATGCCCTCG | Maize | [96] |
| Provitamin A | ZmcrtRB3 | 2 | F-GTCGGTACTGGCAAGTGGAA R-TAGTACGTGGCCATGACGTG | Maize | [161] |
| Sweetness | sugary1 | 4 | F-TCCCGACTTCAGAACGGTTG R-ACAACAGAGCAACCCCAACA | Maize | [162] |
| Provitamin A | crtRB1 and LcyE | 10,8 | F-CACAGGTCGCTGCGTACTTA R-GGGAGACAGCTCACAGGAAC & F-CAGTGCGCTGAAGGCTACTA R-GGATGAAAGGGTCGAGCCAA | Maize | [163] |
| Soluble acid invertase | SAI-1 | 4 | F-GGATTCCACTTCCAGCCACA R-CGACGGGGTAGAAGTCGATG | Sorghum | [164] |
| Fragrance | SbBADH2 | 4 | F-CGCAGTAGTGGAGTGGTTGT R-ACTGTGGCGGTTCTTGCATA | Sorghum | [165] |
| Fragrance allele | Gmbadh2-1 and Gmbadh2-2 | 5 | F-GTGATCTGCGAGGGAGGGAG R-TGAGTTGCAGGCAGTGTCAT | Soybean | [166] |
| White flesh | wf | 9 | F-TTGGAGGTTCAATGCTTGCC R-CAAAGACCAGAGCACCATCG | Melon | [167] |
| Green flesh color | gf | 8 | F-TCTGCAAAATGGTTGCTTTGAA R-AGGTGGATGTGGCACACAAA | Melon | [141] |
| Flavonoids | AgFNSI | 4 | F-ATGGCTCCATCAAC TATAAC R-CTGCCCTGGCAATCTCCG | Celery | [168] |
| Starch content | NnHXK and NnGBSS | Unmapped | F-TCTAAATCCCAATCCGTCC R-GCACGAACTCTTGGCAATC | Lotus | [169] |
| Pungency | Pup1 | 2 | F-CCATGGATTGTTGCTCGGGCCTCC R-CCGTACCGCCCCATTGCGATTCC | Chilli | [170] |
| Anthocyanin biosynthesis | VfTTG1 | Unmapped | F-TATGAATTCATTTTTAGTTCCCACCTAAC R-GTATCCGGTTGAGGACTCTCATAGATA | Faba bean | [171] |
| β-Carotene & Flesh | QA/QC | 3 | F-AGTGCGGGACAAGATGATCA R-TCCCGAACATCTGAGCAAGT | Sweet potato | [172] |
| Carotenoids | b_CHYβ-1 | Unmapped | F-TCCAGCTTGGGAATTACGTC R-ACAACGAAGCGTGCCATAG | Sweet potato | [173] |
| Biotic stresses | |||||
| Powdery mildew | Pm3 | 1A | F-CAAGTACCAACCACAGCCAC R-CCATTGCAACCACAGGAACA | Wheat | [174] |
| Stem rust resistance | Sr45 | 1D | F-GTCCATTTTACGACGGTCCG R-CTGGTCGGTAGGGAAGCTAG | Wheat | [175] |
| Bacterial blight resistance | Xa3 | 11 | F-GAATGGGTGGGGTTGGGAAG R-CCATGCACGCTTGTCGAATC | Rice | [176] |
| Bacterial blight resistance | xa5 | 5 | F-ACGGAGTTGCAATGTTGCTG R-GGCCAGGAGTAAAGCGGATT | Rice | [177] |
| Bacterial blight resistance | xa13 | 8 | F-GGCCATGGCTCAGTGTTTAT R-GAGCTCCAGATCTCCAAATG | Rice | [178] |
| Bacterial blight resistance | Xa21 | 11 | F-AGACGCGGAAGGGTGGTTCCCGG R-AGACCGGTAATCGAAAGATGAAA | Rice | [179] |
| Bacterial blight resistance | Xa38 | 4 | F-TCTTCTATTGCTAACATTGGTG R-AGCGTAAGTAAAAGTCTC | Rice | [180] |
| Brown plant hopper resistance | Bph14 | 3 | F-CAATCCGAGCTTACGTGGTG R-GGTGGAGAAGGCAAGAGTCT | Rice | [181] |
| Blast resistance | Pit | 1 | F-GTGACGGAAGTGCATGGGTA R-ACCAGGGAACCCGACAAGAA | Rice | [182] |
| Blast resistance | Pi54 (Pikh) | 11 | F-CCTCTTGAGTTGAATTGGCACG R-CCTCGTGCAGCTGTTTTCAC | Rice | [183] |
| Blast resistance | Pi35 | 1 | F-TCCATGGCGGAGGTGGTGTTGGCTG R-AGAGCAAATCTTGGGGTGTCTGCAA | Rice | [184] |
| Blast Resistance | PigmR | 6 | F-ATGTCGGAGGAAGCAGGTC R-ATGTCACGCAGCAAAACCAT | Rice | [185] |
| Brown plant hopper resistance | Bph9 | 12 | F-CACTCGCACGGATACAATGG R-GATCGTGACACATGCATGCT | Rice | [186] |
| Powdery mildew | NBS–LRR class of resistance genes | 2 | F-CGTTTTGTATGGCGTCCGAT R-TTGTCGCTGAGGTCCATCTT | Barley | [187] |
| General stress response | ERF transcription factors | 1 | F-ACAGTGGTGGCAAGTGTGAA R-ACGGCCTCCTTCTTACTCCT | Several crops | [188] |
| Leaf rust resistance | Rph7 | 3H | F-TGGAAACCACTGTACAGCCT R-CAGGCATGGGAGTGAACCTA | Barley | [189] |
| Tomato yellow leaf curl virus | ACY | 6 | F-CCTTATGATGTCTCGTGAAAGG R-GAAGCACAGATTGAAGAAAACC | Tomato | [190] |
| Bacterial wilt | Bwr-6, Bwr-12 | 6,12 | F-TCAAGGTCCACTACCTTCATCC R-TCGGTATAGAGGGTACGTTG | Tomato | [191] |
| Fusarium wilt | frl | 9 | F-TACGATGACGTCGGT R-ATGCTACTGCGATGAAAC | Tomato | [192] |
| Fusarium wilt | Fom 1 | 7 | F-AACGAGAAGGCGGTGGAAAT R-CGATCTCCTCAAGGGAAGGTG | Melon | [193] |
| Leaf scald resistance | Rpf | Unmapped | F-TTGTTGGAACCTTTCGCTGG R-TAGACCTGTGCTGCCGTAAA | Sugarcane | [194] |
| Powdery mildew resistance | Pm-2 F | 1 | F-GCCCAACCTTCAACTCGATA R-TTGAATCTCATTTTTCTGTTGCAT | Melon | [195] |
| Melon necrotic spot virus | nsv | 4 | F-GTTTCTGATACGATGTTGTTTCCCTG R-GCCGAGATGCAGCAGGATGCTTTGCAC | Melon | [196] |
| Zucchini yellow mosaic virus (ZYMV) | eIF4E | 3 | F-TGGACITTYTGGTTYGAYAA R-GGRTCYTCCCAYTTIGGYTC | Watermelon | [197] |
| Powdery mildew | Pm | 5 | F-ATTTTCTTGCTTCAAATGGA R-ATAAGCAAAAGCATCGAAAG | Watermelon | [198] |
| Powdery mildew | Pm-s | 5 | F-CCCTATGCGTGAAAGCCACT R-CGCCTCAAACCCATACCCAA | Cucumber | [199] |
| Cauliflower mosaic virus (CMV) | cmv6.1 | 6 | F-ACAAAGCTTCTCCGCAAATG R-GGAGGGAAAGGAAGGAGAGA | Cucumber | [200] |
| Bacterial wilt-resistance | S401 | 6 | F--G ACTGCGTACCAATTCAGTT R-GATGAGTCCTGAGTAACACGATG | Eggplant | [201] |
| Powdery mildew resistance | InDel 1 | 1 | F-AACTTGGTAGCAATTTTATTGGGT R-TGGAGACAATGTGCATAAGTCTCT | Capsicum | [202] |
| Bacterial leaf spot resistance | Xcvr | 2 | F-TATCAAACGTAAAGTTGGAGCTTGT R-CCAAACACCTTGTGCATTGCT | Lettuce | [203] |
| Turnip mosaic virus resistance | retr02 | 4A | F-GGAGAAGACAAACAAACCCCC R-A TACCTTCGACACCGTCCAAGACTT | Turnip | [204] |
| Powdery mildew resistance | er1-7 | 6 | F-CGACACCGTATTCAAGCAGG R-TGTTGCCCTGTTTGATCGTT | Pea | [205] |
| Mungbean Yellow Mosaic Virus | YR4 | Unmapped | F-ACAAACATGGGCTGGAACAC R-GTGCCTGTAACTGCTCACAC | Mung bean | [206] |
| Resistance to weevils | VrPDF1 | Unmapped | F-CCAAGCTTGGTTAACAGTTTCTAGTGCACC R-GCGTCGACGATGGAGAAGAAATCACTGGCC | Mung bean | [207] |
| Abiotic stresses | |||||
| Dehydration tolerance | TaMYB2 | Unmapped | F-GAGGCCAGCTAGCAGCTGCC R-ATTGCCGGACGCGCAAGAGG | Wheat | [208] |
| Drought stress tolerance | TaAQP | Unmapped | F-ACATCAATTTTACCGTGCTTTG R-CAATCAATCTGCCGACTGTG | Wheat | [209] |
| Drought stress tolerance | DREB1 | 3D | F-GAATGGATCCCGGAAAGCAC R-GGGAATGAACCAAGCCACAG | Wheat | [210] |
| Salt tolerance | TtASR1 | Unmapped | F-ACCCCTACTTCTACATGCCG R-ATGATGGAGCTGTGGGACG | Wheat | [211] |
| Submergence tolerance | SubA1 | 9 | F-CTAGTTGGGCATACGATGGC R-ACGCTTATATGTTACGTCAAC | Rice | [212] |
| Tolerance to phosphorus (P) deficiency | Pup 1 | 12 | F-CTGGACTTGACCCCAATGTA R-TCTGATGGAGTGTTCGGAGT | Rice | [213] |
| Drought stress tolerance | OsSAPK2 | 5 | F-AAGGACATAGGGTCGGGGAA R-TGGCCAAATGTGTGGGAGTT | Rice | [214] |
| Drought tolerance | MYBE1 | 5 | F-GGTACCCTGTCAAGGTTCGG R-AATTACTGGCCCCAGGTTCG | Maize | [215] |
| Photoperiod response | Phd-H1 | 2H | F-GTTGAGATCGACAGTCCCCA R-GGGCTCCTATCTCCAACTCC | Barley | [135] |
| Aluminum stress tolerance | SbMATE | 4 | F-TAAGGCGCAATCATCATGGC R-CAACAAGATTCTGGAGCCGG | Sorghum | [216] |
| Drought and salt stress tolerance | CPRD12 | 11 | F-AAAGCATGCCCTAGTGGGAC R-ATGTCGGAAGCTACGGTTTCT | Cowpea | [217] |
| Dehydration response | SiDREB2 | Unmapped | F-CAACGGACTTGGGGCAAATG R-ATCGTTCGCTTCTGCCTTCA | Foxtail | [218] |
| Salinity tolerance | Salt index_QTL 1 | Unmapped | F-TGTACACTGTGTTTCTGTTGGT R-GTATTCGATCGTCCCTCCCG | Field pea | [219] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Salgotra, R.K.; Stewart, C.N., Jr. Functional Markers for Precision Plant Breeding. Int. J. Mol. Sci. 2020, 21, 4792. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134792
AMA Style
Salgotra RK, Stewart CN Jr. Functional Markers for Precision Plant Breeding. International Journal of Molecular Sciences. 2020; 21(13):4792. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134792
Chicago/Turabian StyleSalgotra, Romesh K., and C. Neal Stewart, Jr. 2020. "Functional Markers for Precision Plant Breeding" International Journal of Molecular Sciences 21, no. 13: 4792. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134792
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.