Transcriptomic Profile of Primary Culture of Skeletal Muscle Cells Isolated from Semitendinosus Muscle of Beef and Dairy Bulls



Abstract
:1. Introduction
2. Results
2.1. Course of In Vitro Primary Culture of Muscle Cells
2.1.1. Primary Skeletal Muscle Cell Culture
2.1.2. Differentiation Culture
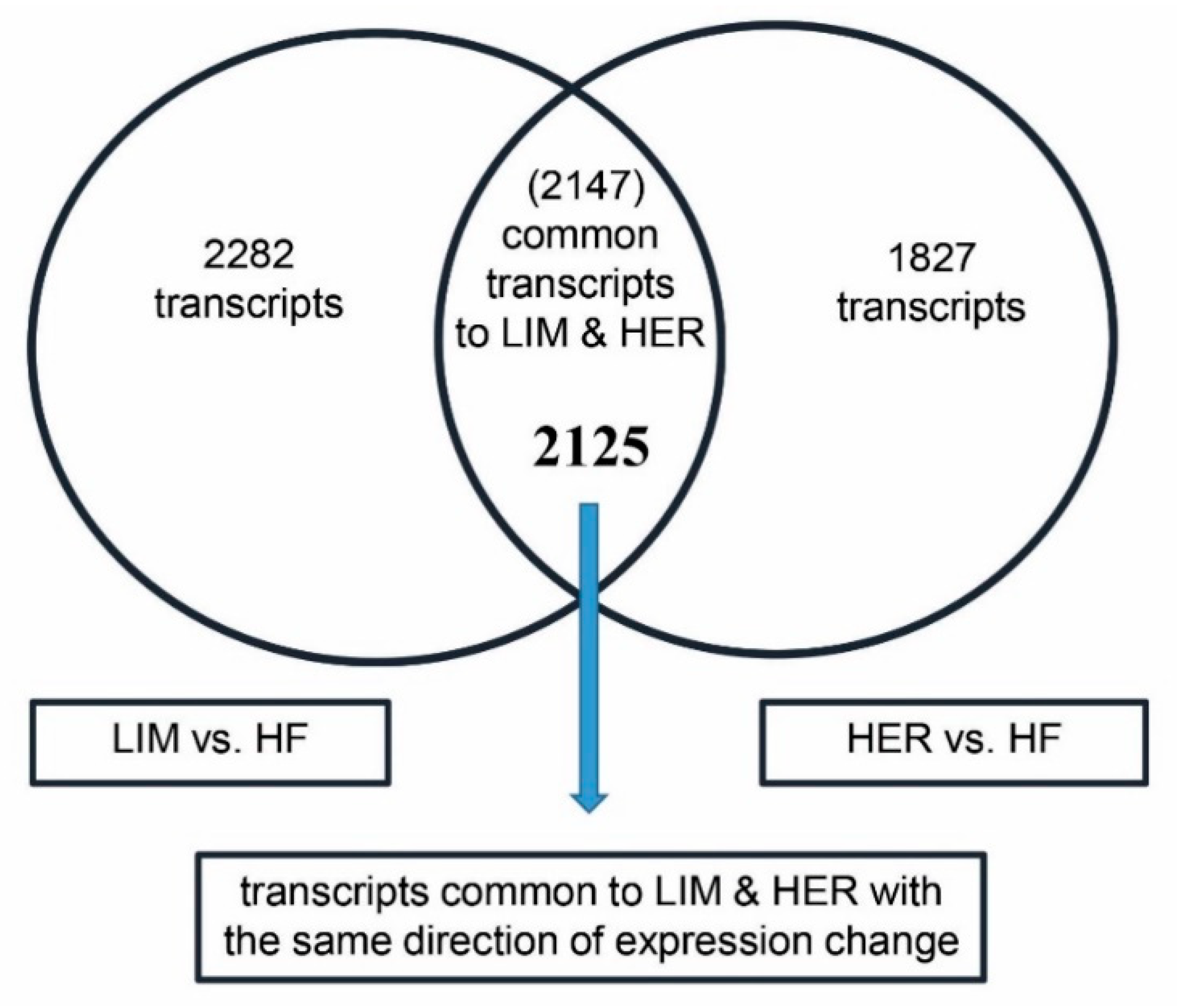
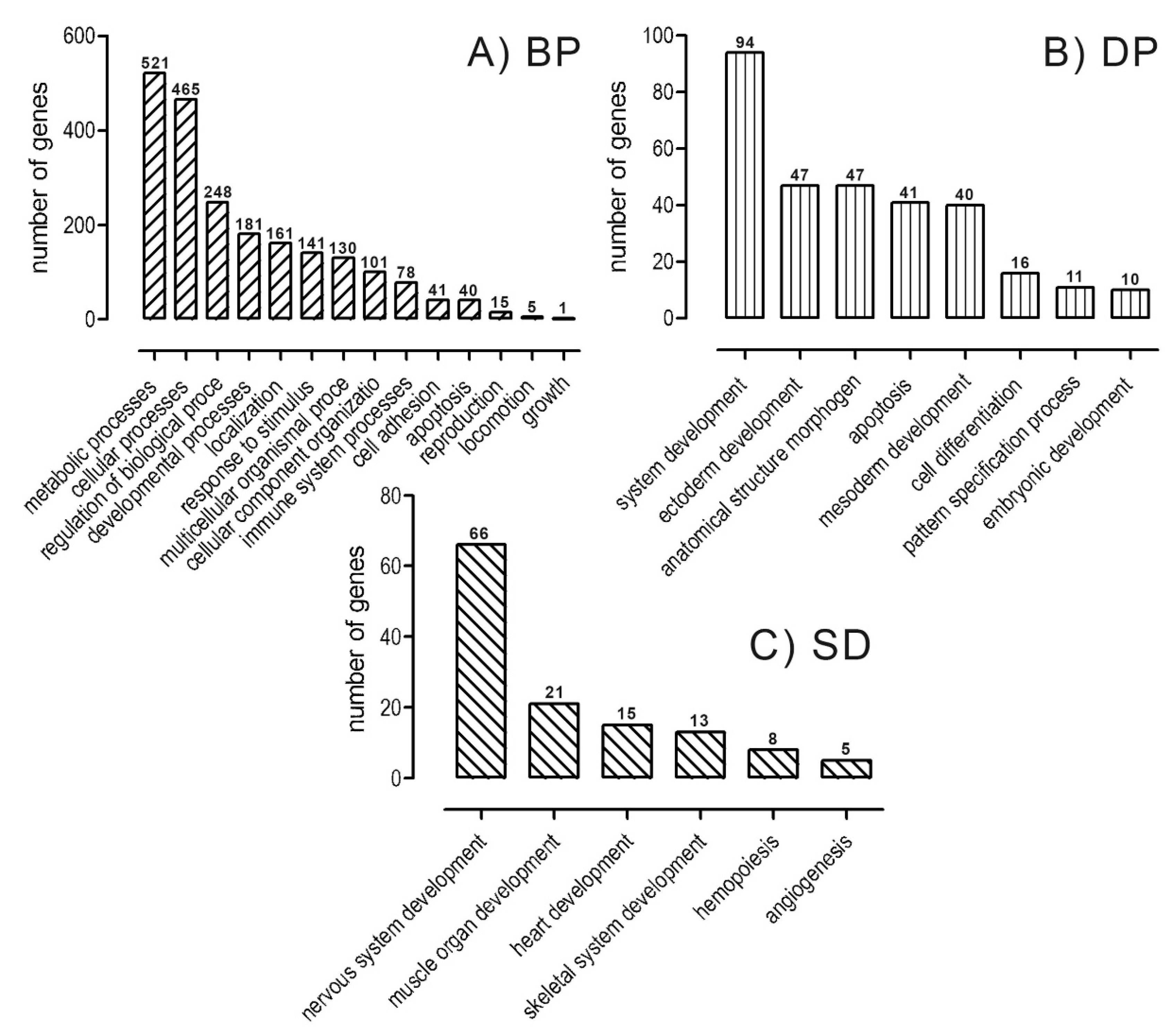
2.2. Transcriptomic and Ontological Analysis
2.3. Result Verification with the Use of qPCR Method
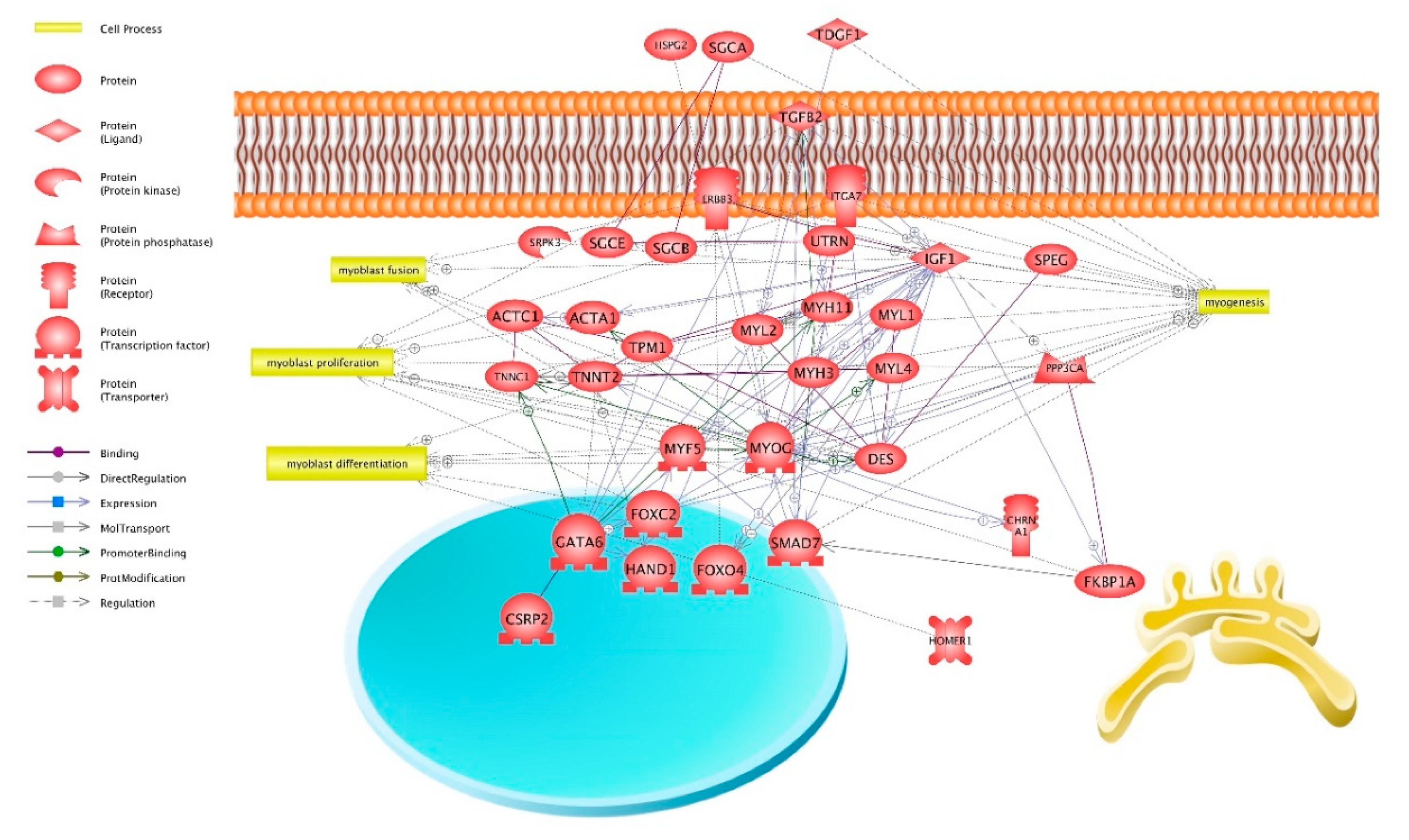
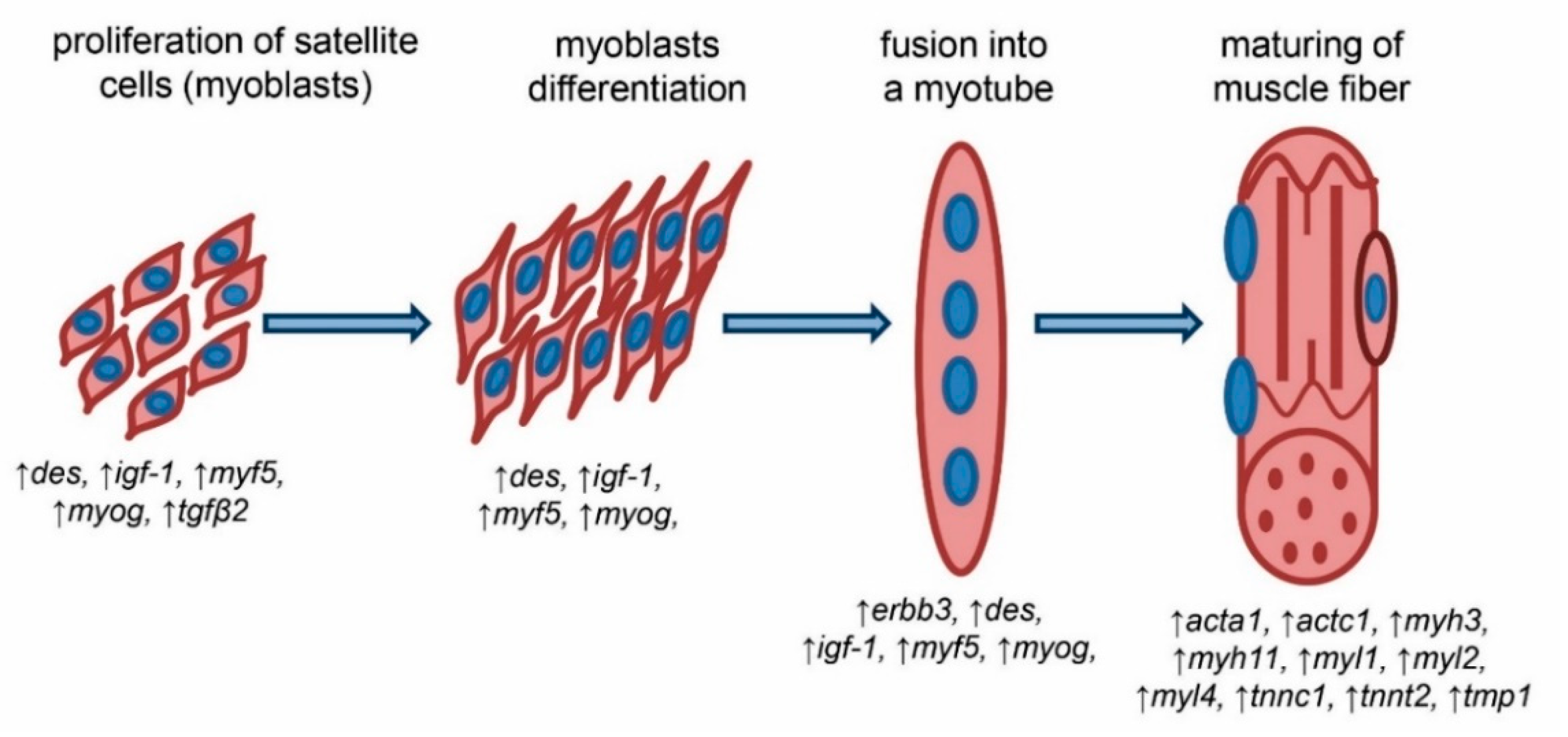
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Animals and Cell Samples
4.3. Skeletal Muscle Cells Isolation, Proliferation, and Differentiation
4.4. Immunocytochemical Analysis
4.5. RNA Isolation and Validation
4.6. Gene Expression Analysis
4.7. qPCR Validation
4.8. Statistical Analysis
4.9. Functional Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 7AAD | 7-aminoactinomycin D |
| DAVID | Database for Annotation, Visualization, and Integrated Discovery |
| DMEM | Dulbecco’s Modified Eagle Medium |
| DMSO | Dimethyl sulfoxide |
| FBS | Fetal bovine serum |
| FC | Fold change |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| HER | Hereford |
| HF | Holstein-Friesian |
| HS | Horse serum |
| LIM | Limousin |
| NCBI GEO | National Center for Biotechnology Information Gene Expression Omnibus database |
| PBS | Phosphate-buffered saline |
| qPCR | Real-time polymerase chain reaction |
| RIN | RNA Integrity Number |
References
- Motyl, T.; Sadkowski, T.; Jank, M.; Wicik, Z. Miogeneza-rozwój mięśni szkieletowych. In Genomika Bydła i Świń-Wybrane Zagadnienia; Zwierzchowski, L., Świtoński, M., Eds.; Instytut Genetyki i Hodowli Zwierząt, Polska Akademia Nauk: Jastrzębiec, Poland, 2009; Volume 9.1, pp. 250–261. [Google Scholar]
- Sadkowski, T.; Jank, M.; Oprzadek, J.; Motyl, T. Age-dependent changes in bovine skeletal muscle transcriptomic profile. J. Physiol. Pharmacol. 2006, 57, S95–S110. [Google Scholar]
- Robelin, J.; Tulloh, N.M. Paterns of growth of cattle. In Beef Cattle Production; Jarrige, R., Beranger, C., Eds.; Elsevier: Paris, France, 1992; pp. 111–129. [Google Scholar]
- Dhawan, J.; Rando, T.A. Stem cells in postnatal myogenesis: Molecular mechanisms of satellite cell quiescence, activation and replenishment. Trends Cell Biol. 2005, 15, 666–673. [Google Scholar] [CrossRef] [PubMed]
- Relaix, F.; Zammit, P.S. Satellite cells are essential for skeletal muscle regeneration: The cell on the edge returns centre stage. Development 2012, 139, 2845–2856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steyn, P.J.; Dzobo, K.; Smith, R.I.; Myburgh, K.H. Interleukin-6 Induces Myogenic Differentiation via JAK2-STAT3 Signaling in Mouse C2C12 Myoblast Cell Line and Primary Human Myoblasts. Int. J. Mol. Sci. 2019, 20, 5273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuang, S.; Rudnicki, M.A. The emerging biology of satellite cells and their therapeutic potential. Trends Mol. Med. 2008, 14, 82–91. [Google Scholar] [CrossRef]
- Bazgir, B.; Fathi, R.; Rezazadeh Valojerdi, M.; Mozdziak, P.; Asgari, A. Satellite Cells Contribution to Exercise Mediated Muscle Hypertrophy and Repair. Cell J. 2017, 18, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Yamakawa, H.; Kusumoto, D.; Hashimoto, H.; Yuasa, S. Stem Cell Aging in Skeletal Muscle Regeneration and Disease. Int. J. Mol. Sci. 2020, 21, 1830. [Google Scholar] [CrossRef] [Green Version]
- Ciecierska, A.; Sadkowski, T.; Motyl, T. Role of satellite cells in growth and regeneration of skeletal muscles. Med. Weter. 2019, 75, 707–712. [Google Scholar] [CrossRef]
- Goh, Q.; Millay, D.P. Requirement of myomaker-mediated stem cell fusion for skeletal muscle hypertrophy. Elife 2017, 6, 20007. [Google Scholar] [CrossRef]
- Chen, J.F.; Tao, Y.; Li, J.; Deng, Z.; Yan, Z.; Xiao, X.; Wang, D.Z. MicroRNA-1 and microRNA-206 regulate skeletal muscle satellite cell proliferation and differentiation by repressing Pax7. J. Cell Biol. 2010, 190, 867–879. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.J.; Pokharel, S.; Jan, A.T.; Huh, S.; Galope, R.; Lim, J.H.; Lee, D.M.; Choi, S.W.; Nahm, S.S.; Kim, Y.W.; et al. Transthyretin: A Transporter Protein Essential for Proliferation of Myoblast in the Myogenic Program. Int. J. Mol. Sci. 2017, 18, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dey, B.; Gagan, J.; Dutta, A. MiR-206 and -486 induce myoblast differentiation by downregulating Pax7. Mol. Cell Biol. 2011, 31, 203–214. [Google Scholar] [CrossRef] [Green Version]
- Berkes, C.A.; Tapscott, S.J. MyoD and the transcriptional control of myogenesis. Semin. Cell Dev. Biol. 2005, 16, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Sadkowski, T.; Ciecierska, A.; Oprządek, J.; Balcerek, E. Breed-dependent microRNA expression in the primary culture of skeletal muscle cells subjected to myogenic differentiation. BMC Genom. 2018, 19, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laker, R.C.; Ryall, J.G. DNA Methylation in Skeletal Muscle Stem Cell Specification, Proliferation, and Differentiation. Stem Cells Int. 2016, 2016, 5725927. [Google Scholar] [CrossRef] [Green Version]
- Zammit, P.S. Function of the myogenic regulatory factors Myf5, MyoD, Myogenin and MRF4 in skeletal muscle, satellite cells and regenerative myogenesis. Semin. Cell Dev. Biol. 2017, 72, 19–32. [Google Scholar] [CrossRef]
- Yu, M.; Wang, H.; Xu, Y.; Yu, D.; Li, D.; Liu, X.; Du, W. Insulin-like growth factor-1 (IGF-1) promotes myoblast proliferation and skeletal muscle growth of embryonic chickens via the PI3K/Akt signalling pathway. Cell Biol. Int. 2015, 39, 910–922. [Google Scholar] [CrossRef]
- Lindon, C.; Montarras, D.; Pinset, C. Cell cycle-regulated expression of the muscle determination factor Myf5 in proliferating myoblasts. J. Cell Biol. 1998, 140, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Panda, A.C.; Abdelmohsen, K.; Martindale, J.L.; Di Germanio, C.; Yang, X.; Grammatikakis, I.; Noh, J.H.; Zhang, Y.; Lehrmann, E.; Dudekula, D.B.; et al. Novel RNA-binding activity of MYF5 enhances Ccnd1/Cyclin D1 mRNA translation during myogenesis. Nucleic Acids Res. 2016, 44, 2393–2408. [Google Scholar] [CrossRef] [Green Version]
- Ustanina, S.; Carvajal, J.; Rigby, P.; Braun, T. The myogenic factor Myf5 supports efficient skeletal muscle regeneration by enabling transient myoblast amplification. Stem Cells 2007, 25, 2006–2016. [Google Scholar] [CrossRef]
- Coles, C.A.; Wadeson, J.; Leyton, C.P.; Siddell, J.P.; Greenwood, P.L.; White, J.D.; McDonagh, M.B. Proliferation rates of bovine primary muscle cells relate to liveweight and carcase weight in cattle. PLoS ONE 2015, 10, e0124468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadkowski, T.; Jank, M.; Zwierzchowski, L.; Oprzadek, J.; Motyl, T. Transcriptomic index of skeletal muscle of beef breeds bulls. J. Physiol. Pharmacol. 2009, 60, S15–S28. [Google Scholar]
- Iwanowska, A.; Pospiech, E.; Łyczyński, A.; Rosochacki, S.; Grześl, B.; Mikołajczak, B.; Iwańska, E.; Rzosińska, E.; Czyżak-Runowska, G. Evaluation of variations in principal indices of the culinary meat quality obtained from young bulls of various breeds. Acta Sci. Pol. Technol. Aliment. 2010, 9, 133–149. [Google Scholar]
- De Arcangelis, V.; Coletti, D.; Conti, M.; Lagarde, M.; Molinaro, M.; Adamo, S.; Nemoz, G.; Naro, F. IGF-I-induced differentiation of L6 myogenic cells requires the activity of cAMP-phosphodiesterase. Mol. Biol. Cell 2003, 14, 1392–1404. [Google Scholar] [CrossRef] [Green Version]
- Lawson-Smith, M.J.; McGeachie, J.K. The identification of myogenic cells in skeletal muscle, with emphasis on the use of tritiated thymidine autoradiography and desmin antibodies. J. Anat. 1998, 192, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Aswad, H.; Jalabert, A.; Rome, S. Depleting extracellular vesicles from fetal bovine serum alters proliferation and differentiation of skeletal muscle cells in vitro. BMC Biotechnol. 2016, 16, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malik, A.; Lee, E.J.; Jan, A.T.; Ahmad, S.; Cho, K.H.; Kim, J.; Choi, I. Network Analysis for the Identification of Differentially Expressed Hub Genes Using Myogenin Knock-down Muscle Satellite Cells. PLoS ONE 2015, 10, e0133597. [Google Scholar] [CrossRef] [Green Version]
- Oldham, J.M.; Martyn, J.A.; Sharma, M.; Jeanplong, F.; Kambadur, R.; Bass, J.J. Molecular expression of myostatin and MyoD is greater in double-muscled than normal-muscled cattle fetuses. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 280, R1488–R1493. [Google Scholar] [CrossRef]
- Wang, Y.X.; Dumont, N.A.; Rudnicki, M.A. Muscle stem cells at a glance. J. Cell Sci. 2014, 127, 4543–4548. [Google Scholar] [CrossRef] [Green Version]
- André, L.M.; Ausems, C.R.M.; Wansink, D.G.; Wieringa, B. Abnormalities in Skeletal Muscle Myogenesis, Growth, and Regeneration in Myotonic Dystrophy. Front. Neurol. 2018, 9, 368. [Google Scholar] [CrossRef]
- Lee, E.J.; Malik, A.; Pokharel, S.; Ahmad, S.; Mir, B.A.; Cho, K.H.; Kim, J.; Kong, J.C.; Lee, D.M.; Chung, K.Y.; et al. Identification of genes differentially expressed in myogenin knock-down bovine muscle satellite cells during differentiation through RNA sequencing analysis. PLoS ONE 2014, 9, e92447. [Google Scholar] [CrossRef] [PubMed]
- Rønning, S.B.; Pedersen, M.E.; Andersen, P.V.; Hollung, K. The combination of glycosaminoglycans and fibrous proteins improves cell proliferation and early differentiation of bovine primary skeletal muscle cells. Differentiation 2013, 86, 13–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.; Li, S.; Tong, H.; Li, S.; Yan, Y. WDR13 promotes the differentiation of bovine skeletal muscle-derived satellite cells by affecting PI3K/AKT signaling. Cell Biol. Int. 2019, 43, 799–808. [Google Scholar] [CrossRef] [PubMed]
- De Las Heras-Saldana, S.; Chung, K.Y.; Lee, S.H.; Gondro, C. Gene expression of Hanwoo satellite cell differentiation in longissimus dorsi and semimembranosus. BMC Genom. 2019, 20, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Younis, S.; Naboulsi, R.; Wang, X.; Cao, X.; Larsson, M.; Sargsyan, E.; Bergsten, P.; Welsh, N.; Andersson, L. Dissection of the cellular function of the ZBED6 transcription factor in mouse myoblast cells using gene editing, RNAseq and proteomics. bioRxiv 2019. [Google Scholar] [CrossRef]
- Park, J.W.; Lee, J.H.; Kim, S.W.; Han, J.S.; Kang, K.S.; Kim, S.J.; Park, T.S. Muscle differentiation induced up-regulation of calcium-related gene expression in quail myoblasts. Asian-Australas. J. Anim. Sci. 2018, 31, 1507–1515. [Google Scholar] [CrossRef]
- Kumar, A.; Yamauchi, J.; Girgenrath, T.; Girgenrath, M. Muscle-specific expression of insulin-like growth factor 1 improves outcome in Lama2Dy-w mice, a model for congenital muscular dystrophy type 1A. Hum. Mol. Genet. 2011, 20, 2333–2343. [Google Scholar] [CrossRef] [Green Version]
- Adams, G.R.; Haddad, F.; Baldwin, K.M. Time course of changes in markers of myogenesis in overloaded rat skeletal muscles. J. Appl. Physiol. 1999, 87, 1705–1712. [Google Scholar] [CrossRef]
- McKay, B.R.; O’Reilly, C.E.; Phillips, S.M.; Tarnopolsky, M.A.; Parise, G. Co-expression of IGF-1 family members with myogenic regulatory factors following acute damaging muscle-lengthening contractions in humans. J. Physiol. 2008, 586, 5549–5560. [Google Scholar] [CrossRef]
- Chal, J.; Pourquié, O. Making muscle: Skeletal myogenesis in vivo and in vitro. Development 2017, 144, 2104–2122. [Google Scholar] [CrossRef] [Green Version]
- Ascenzi, F.; Barberi, L.; Dobrowolny, G.; Villa Nova Bacurau, A.; Nicoletti, C.; Rizzuto, E.; Rosenthal, N.; Scicchitano, B.M.; Musarò, A. Effects of IGF-1 isoforms on muscle growth and sarcopenia. Aging Cell 2019, 18, e12954. [Google Scholar] [CrossRef] [PubMed]
- Musarò, A.; McCullagh, K.; Paul, A.; Houghton, L.; Dobrowolny, G.; Molinaro, M.; Barton, E.R.; Sweeney, H.L.; Rosenthal, N. Localized Igf-1 transgene expression sustains hypertrophy and regeneration in senescent skeletal muscle. Nat. Genet. 2001, 27, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Pietrangelo, T.; Puglielli, C.; Mancinelli, R.; Beccafico, S.; Fanò, G.; Fulle, S. Molecular basis of the myogenic profile of aged human skeletal muscle satellite cells during differentiation. Exp. Gerontol. 2009, 44, 523–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szewczuk, M.; Sablik, P.; Kulig, H. The effect of polymorphism within exon 12 of IGF1R gene on meat production traits in Limousin and Hereford cattle. Acta Sci. Pol. Zootech. 2017, 16, 53–56. [Google Scholar] [CrossRef]
- Ge, W.; Davis, M.E.; Hines, H.C.; Irvin, K.M.; Simmen, R.C. Association of a genetic marker with blood serum insulin-like growth factor-I concentration and growth traits in Angus cattle. J. Anim. Sci. 2001, 79, 1757–1762. [Google Scholar] [CrossRef] [Green Version]
- Szewczuk, M.; Zych, S.; Wójcik, J.; Czerniawska-Piątkowska, E. Association of two SNPs in the coding region of the insulin-like growth factor 1 receptor (IGF1R) gene with growth-related traits in Angus cattle. J. Appl. Genet. 2013, 54, 305–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szewczuk, M. Analysis of the relationship between insulin-like growth factor 1 receptor gene polymorphisms in Montbeliarde cows and the birth weight of their calves. Acta Vet. Brno. 2016, 85, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Chung, E.; Kim, W. Association of SNP Marker in IGF-I and MYF5 Candidate Genes with Growth Traits in Korean Cattle. Asian-Aust. J. Anim. Sci. 2005, 18, 1061–1065. [Google Scholar] [CrossRef]
- Ouyang, H.; Wang, Z.; Chen, X.; Yu, J.; Li, Z.; Nie, Q. Proteomic Analysis of Chicken Skeletal Muscle during Embryonic Development. Front. Physiol. 2017, 8, 281. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.J.; Bajracharya, P.; Lee, D.M.; Kang, S.W.; Lee, Y.S.; Lee, H.J.; Hong, S.K.; Chang, J.; Kim, J.W.; Schnabel, R.D.; et al. Gene expression profiles during differentiation and transdifferentiation of bovine myogenic satellite cells. Genes Genom. 2012, 34, 133–148. [Google Scholar] [CrossRef]
- Moran, J.L.; Li, Y.; Hill, A.A.; Mounts, W.M.; Miller, C.P. Gene expression changes during mouse skeletal myoblast differentiation revealed by transcriptional profiling. Physiol. Genom. 2002, 10, 103–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sterrenburg, E.; Turk, R.; Hoen, P.A.; van Deutekom, J.C.; Boer, J.M.; van Ommen, G.J.; den Dunnen, J.T. Large-scale gene expression analysis of human skeletal myoblast differentiation. Neuromuscul. Disord. 2004, 14, 507–518. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xu, X.; Liu, Y.; Zhang, L.; Odle, J.; Lin, X.; Zhu, H.; Wang, X.; Liu, Y. EPA and DHA Inhibit Myogenesis and Downregulate the Expression of Muscle-related Genes in C2C12 Myoblasts. Genes (Basel) 2019, 10, 64. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Shi, T.; Cai, H.; Zhou, Y.; Lan, X.; Zhang, C.; Lei, C.; Qi, X.; Chen, H. Associations of MYH3 gene copy number variations with transcriptional expression and growth traits in Chinese cattle. Gene 2014, 535, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, X.; Niu, F.; Wang, H.; He, H.; Gu, Y. Single nucleotide polymorphisms, haplotypes and combined genotypes in MYH3 gene and their associations with growth and carcass traits in Qinchuan cattle. Mol. Biol. Rep. 2013, 40, 417–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matheny, R.W., Jr.; Nindl, B.C. Loss of IGF-IEa or IGF-IEb impairs myogenic differentiation. Endocrinology 2011, 152, 1923–1934. [Google Scholar] [CrossRef] [Green Version]
- Cronin, E.M.; Thurmond, F.A.; Bassel-Duby, R.; Williams, R.S.; Wright, W.E.; Nelson, K.D.; Garner, H.R. Protein-coated poly(L-lactic acid) fibers provide a substrate for differentiation of human skeletal muscle cells. J. Biomed. Mater. Res. A 2004, 69, 373–381. [Google Scholar] [CrossRef]
- Li, X.J.; Zhou, J.; Liu, L.Q.; Qian, K.; Wang, C.L. Identification of genes in longissimus dorsi muscle differentially expressed between Wannanhua and Yorkshire pigs using RNA-sequencing. Anim. Genet. 2016, 47, 324–333. [Google Scholar] [CrossRef]
- Yan, Z.; Choi, S.; Liu, X.; Zhang, M.; Schageman, J.J.; Lee, S.Y.; Hart, R.; Lin, L.; Thurmond, F.A.; Williams, R.S. Highly coordinated gene regulation in mouse skeletal muscle regeneration. J. Biol. Chem. 2003, 278, 8826–8836. [Google Scholar] [CrossRef] [Green Version]
- Jaeger, M.A.; Sonnemann, K.J.; Fitzsimons, D.P.; Prins, K.W.; Ervasti, J.M. Context-dependent functional substitution of alpha-skeletal actin by gamma-cytoplasmic actin. FASEB J. 2009, 23, 2205–2214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lolis, A.A.; Londhe, P.; Beggs, B.C.; Byrum, S.D.; Tackett, A.J.; Davie, J.K. Myogenin recruits the histone chaperone facilitates chromatin transcription (FACT) to promote nucleosome disassembly at muscle-specific genes. J. Biol. Chem. 2013, 288, 7676–7687. [Google Scholar] [CrossRef] [Green Version]
- Bisping, E.; Ikeda, S.; Sedej, M.; Wakula, P.; McMullen, J.R.; Tarnavski, O.; Sedej, S.; Izumo, S.; Pu, W.T.; Pieske, B. Transcription factor GATA4 is activated but not required for insulin-like growth factor 1 (IGF1)-induced cardiac hypertrophy. J. Biol. Chem. 2012, 287, 9827–9834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chua, M.J.; Yildirim, E.D.; Tan, J.E.; Chua, Y.B.; Low, S.C.; Ding, S.L.S.; Li, C.W.; Jiang, Z.; Teh, B.T.; Yu, K.; et al. Assessment of different strategies for scalable production and proliferation of human myoblasts. Cell Prolif. 2019, 52, e12602. [Google Scholar] [CrossRef] [Green Version]
- De Mello, F.; Streit, D.P., Jr.; Sabin, N.; Gabillard, J.C. Dynamic expression of tgf-β2, tgf-β3 and inhibin βA during muscle growth resumption and satellite cell differentiation in rainbow trout (Oncorhynchus mykiss). Gen. Comp. Endocrinol. 2015, 210, 23–29. [Google Scholar] [CrossRef]
- Rudolf, A.; Schirwis, E.; Giordani, L.; Parisi, A.; Lepper, C.; Taketo, M.M.; Le Grand, F. β-Catenin Activation in Muscle Progenitor Cells Regulates Tissue Repair. Cell Rep. 2016, 15, 1277–1290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szcześniak, K.A.; Ciecierska, A.; Ostaszewski, P.; Sadkowski, T. Characterisation of equine satellite cell transcriptomic profile response to β-hydroxy-β-methylbutyrate (HMB). Br. J. Nutr. 2016, 116, 1315–1325. [Google Scholar] [CrossRef] [Green Version]
- Pawlak, A.; Gil, R.J.; Grajkowska, W.; Nasierowska-Guttmejer, A.M.; Rzezak, J.; Kulawik, T. Significance of low desmin expression in cardiomyocytes in patients with idiopathic dilated cardiomyopathy. Am. J. Cardiol. 2013, 111, 393–399. [Google Scholar] [CrossRef]
- Kelc, R.; Trapecar, M.; Gradisnik, L.; Rupnik, M.S.; Vogrin, M. Platelet-rich plasma, especially when combined with a TGF-β inhibitor promotes proliferation, viability and myogenic differentiation of myoblasts in vitro. PLoS ONE 2015, 10, e0117302. [Google Scholar] [CrossRef] [PubMed]
- Portilho, D.M.; Soares, C.P.; Morrot, A.; Thiago, L.S.; Butler-Browne, G.; Savino, W.; Costa, M.L.; Mermelstein, C. Cholesterol depletion by methyl-β-cyclodextrin enhances cell proliferation and increases the number of desmin-positive cells in myoblast cultures. Eur. J. Pharmacol. 2012, 694, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, F.; Yamamoto, M.; Kitamura, K.; Asuka, K.; Kinoshita, H.; Matsunaga, S.; Abe, S. Desmin and Vimentin Expression during Embryonic Development of Tensor Veli Palatini Muscle in Mice. J. Hard Tissue Biol. 2015, 24, 134–142. [Google Scholar] [CrossRef] [Green Version]
- Pawlak, A.; Gil, R.J. Desmin—An important structural protein of a cardiac myocyte. Kardiol. Pol. 2007, 65, 303–309. [Google Scholar] [PubMed]
- Juretić, N.; Díaz, J.; Romero, F.; González, G.; Jaimovich, E.; Riveros, N. Interleukin-6 and neuregulin-1 as regulators of utrophin expression via the activation of NRG-1/ErbB signaling pathway in mdx cells. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 770–780. [Google Scholar] [CrossRef] [PubMed]
- Moscoso, L.M.; Chu, G.C.; Gautam, M.; Noakes, P.G.; Merlie, J.P.; Sanes, J.R. Synapse-associated expression of an acetylcholine receptor-inducing protein, ARIA/heregulin, and its putative receptors, ErbB2 and ErbB3, in developing mammalian muscle. Dev. Biol. 1995, 172, 158–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zorzano, A.; Kaliman, P.; Gumà, A.; Palacín, M. Intracellular signals involved in the effects of insulin-like growth factors and neuregulins on myofibre formation. Cell Signal. 2003, 15, 141–149. [Google Scholar] [CrossRef]
- Kim, D.; Chi, S.; Lee, K.H.; Rhee, S.; Kwon, Y.K.; Chung, C.H.; Kwon, H.; Kang, M.S. Neuregulin stimulates myogenic differentiation in an autocrine manner. J. Biol. Chem. 1999, 274, 15395–15400. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, C.; Duggan, D.; Fischbach, G. Neuregulin induces the expression of transcription factors and myosin heavy chains typical of muscle spindles in cultured human muscle. Proc. Natl. Acad. Sci. USA 2004, 101, 12218–12223. [Google Scholar] [CrossRef] [Green Version]
- Gumà, A.; Martínez-Redondo, V.; López-Soldado, I.; Cantó, C.; Zorzano, A. Emerging role of neuregulin as a modulator of muscle metabolism. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E742–E750. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Liu, X.; Bai, Y.; Tang, N.; Li, J.; Zhang, Y.; Wu, J.; Wang, X.; Wei, J. Neuregulin-1β modulates myogenesis in septic mouse serum-treated C2C12 myotubes in vitro through PPARγ/NF-κB signaling. Mol. Biol. Rep. 2018, 45, 1611–1619. [Google Scholar] [CrossRef]
- Hellyer, N.J.; Mantilla, C.B.; Park, E.W.; Zhan, W.Z.; Sieck, G.C. Neuregulin-dependent protein synthesis in C2C12 myotubes and rat diaphragm muscle. Am. J. Physiol. Cell Physiol. 2006, 291, C1056–C1061. [Google Scholar] [CrossRef] [Green Version]
- Sadkowski, T.; Jank, M.; Zwierzchowski, L.; Oprzadek, J.; Motyl, T. Comparison of skeletal muscle transcriptional profiles in dairy and beef breeds bulls. J. Appl. Genet. 2009, 50, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Wicik, Z.; Sadkowski, T.; Jank, M.; Motyl, T. The transcriptomic signature of myostatin inhibitory influence on the differentiation of mouse C2C12 myoblasts. Pol. J. Vet. Sci. 2011, 14, 643–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szcześniak, K.A.; Ciecierska, A.; Ostaszewski, P.; Sadkowski, T. Transcriptomic profile adaptations following exposure of equine satellite cells to nutriactive phytochemical gamma-oryzanol. Genes Nutr. 2016, 11, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez, R.; Tupac-Yupanqui, I.; Dunner, S. Evaluation of suitable reference genes for gene expression studies in bovine muscular tissue. BMC Mol. Biol. 2008, 9, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadkowski, T.; Ciecierska, A.; Majewska, A.; Oprządek, J.; Dasiewicz, K.; Ollik, M.; Wicik, Z.; Motyl, T. Transcriptional background of beef marbling - novel genes implicated in intramuscular fat deposition. Meat Sci. 2014, 97, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Gene symbol | Gene name (GenBank Accession Number) | LIM vs. HF FC | HER vs. HF FC |
|---|---|---|---|---|
| 1 | acta1 | Bos taurus actin, alpha 1, skeletal muscle (ACTA1), mRNA [NM_174225] | 26.71 | 25.31 |
| 2 | unc45b | unc-45 homolog B (C. elegans) [Source:HGNC Symbol;Acc:14304] [ENSBTAT00000003766] | 22.15 | 10.91 |
| 3 | myh3 | Bos taurus myosin, heavy chain 3, skeletal muscle, embryonic (MYH3), mRNA [NM_001101835] | 19.69 | 16.58 |
| 4 | myl2 | Bos taurus myosin, light chain 2, regulatory, cardiac, slow (MYL2), mRNA [NM_001035025] | 15.10 | 8.60 |
| 5 | col11a1 | Bos taurus collagen, type XI, alpha 1 (COL11A1), mRNA [NM_001166509] | 14.60 | 8.95 |
| 6 | actc1 | Bos taurus actin, alpha, cardiac muscle 1 (ACTC1), mRNA [NM_001034585] | 12.39 | 8.16 |
| 7 | eln | elastin [Source:HGNC Symbol;Acc:3327] [ENSBTAT00000057593] | 12.10 | 8.66 |
| 8 | tnnt2 | Bos taurus troponin T type 2 (cardiac) (TNNT2), mRNA [NM_174771] | 11.90 | 12.98 |
| 9 | myl1 | Bos taurus myosin, light chain 1, alkali; skeletal, fast (MYL1), mRNA [NM_001079578] | 11.56 | 8.76 |
| 10 | igf1 | PREDICTED: Bos taurus insulin like growth factor 1 (IGF1), transcript variant X8, mRNA [XM_005206500] | 11.10 | 13.75 |
| 11 | ttn | titin [Source:HGNC Symbol;Acc:12403] [ENSBTAT00000061449] | 9.10 | 4.27 |
| 12 | tnnc1 | Bos taurus troponin C type 1 (slow) (TNNC1), mRNA [NM_001034351] | 8.07 | 5.69 |
| 13 | des | Bos taurus desmin (DES), mRNA [NM_001081575] | 7.51 | 5.54 |
| 14 | hspg2 | PREDICTED: Bos taurus heparan sulfate proteoglycan 2 (HSPG2), mRNA [XM_582024] | 7.07 | 4.37 |
| 15 | myf5 | Bos taurus myogenic factor 5 (MYF5), mRNA [NM_174116] | 5.32 | 2.61 |
| 16 | sgca | PREDICTED: Bos taurus sarcoglycan alpha (SGCA), transcript variant X1, mRNA [XM_005220623] | 4.90 | 4.57 |
| 17 | tpm1 | Bos taurus tropomyosin 1 (alpha) (TPM1), mRNA [NM_001013590] | 4.87 | 4.38 |
| 18 | myl4 | Bos taurus myosin, light chain 4, alkali; atrial, embryonic (MYL4), mRNA [NM_001075149] | 4.26 | 3.43 |
| 19 | myog | Bos taurus myogenin (myogenic factor 4) (MYOG), mRNA [NM_001111325] | 4.11 | 4.19 |
| 20 | tagln3 | Bos taurus transgelin 3 (TAGLN3), mRNA [NM_001034499] | 3.38 | 2.97 |
| 21 | speg | Bos taurus SPEG complex locus, mRNA (cDNA clone IMAGE:8085922), partial cds, [BC113258] | 3.36 | 3.57 |
| 22 | itga7 | Bos taurus integrin, alpha 7 (ITGA7), mRNA [NM_001191305] | 3.10 | 2.64 |
| 23 | erbb3 | Bos taurus v-erb-b2 erythroblastic leukemia viral oncogene homolog 3 (avian) (ERBB3), mRNA [NM_001103105] | 2.91 | 1.57 |
| 24 | myl6b | Bos taurus myosin, light chain 6B, alkali, smooth muscle and non-muscle (MYL6B), mRNA [NM_001075713] | 2.91 | 2.33 |
| 25 | homer1 | PREDICTED: Bos taurus homer scaffolding protein 1 (HOMER1), transcript variant X2, mRNA [XM_015473042] | 2.83 | 3.15 |
| 26 | sgce | Bos taurus sarcoglycan, epsilon (SGCE), mRNA [NM_001075145] | 2.59 | 2.11 |
| 27 | tgfb2 | Bos taurus transforming growth factor, beta 2 (TGFB2), mRNA [NM_001113252] | 2.57 | 2.39 |
| 28 | chrna1 | Bos taurus cholinergic receptor, nicotinic, alpha 1 (muscle) (CHRNA1), mRNA [NM_176664] | 2.46 | 2.82 |
| 29 | foxo4 | Bos taurus forkhead box O4 (FOXO4), mRNA [NM_001101277] | 2.44 | 2.28 |
| 30 | myh11 | Bos taurus myosin, heavy chain 11, smooth muscle (MYH11), mRNA [NM_001102127] | 2.24 | 3.88 |
| 31 | myl6 | Bos taurus myosin, light chain 6, alkali, smooth muscle and non-muscle (MYL6), mRNA [NM_175780] | 2.18 | 1.62 |
| 32 | sgcb | Bos taurus sarcoglycan, beta (43kDa dystrophin-associated glycoprotein) (SGCB), mRNA [NM_001102188] | 2.13 | 1.83 |
| 33 | fkbp1a | Bos taurus FK506 binding protein 1A, 12kDa, mRNA (cDNA clone IMAGE:7951983), partial cds. [BC102338] | 2.05 | 1.80 |
| 34 | utrn | Bos taurus utrophin (UTRN), mRNA [NM_001278561] | 1.95 | 2.26 |
| 35 | hand1 | Bos taurus heart and neural crest derivatives expressed 1 (HAND1), mRNA [NM_001075761] | −2.84 | −1.80 |
| 36 | csrp2 | Bos taurus cysteine and glycine-rich protein 2 (CSRP2), mRNA [NM_001038183] | −2.59 | −2.34 |
| 37 | gata6 | GATA binding protein 6 [Source:HGNC Symbol;Acc:4174] [ENSBTAT00000007537] | −2.51 | −1.75 |
| 38 | srpk3 | Bos taurus SRSF protein kinase 3 (SRPK3), mRNA [NM_001083390] | −2.27 | −1.69 |
| 39 | smad7 | PREDICTED: Bos taurus SMAD family member 7 (SMAD7), transcript variant X1, mRNA [XM_005224231] | −2.27 | −2.11 |
| 40 | ppp3ca | Bos taurus protein phosphatase 3, catalytic subunit, alpha isozyme (PPP3CA), mRNA [NM_174787] | −1.73 | −1.79 |
| 41 | tdgf1 | Bos taurus teratocarcinoma-derived growth factor 1 (TDGF1), mRNA [NM_001080358] | −1.72 | −1.58 |
| 42 | foxc2 | Bos taurus forkhead box C2 (MFH-1, mesenchyme forkhead 1) (FOXC2), mRNA [NM_001193072] | −1.61 | −1.49 |
| 43 | nf1 | Bos taurus neurofibromin 1 (NF1), mRNA, [Source:RefSeq mRNA;Acc:NM_001122728] [ENSBTAT00000015699] | −1.33 | −1.49 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciecierska, A.; Motyl, T.; Sadkowski, T. Transcriptomic Profile of Primary Culture of Skeletal Muscle Cells Isolated from Semitendinosus Muscle of Beef and Dairy Bulls. Int. J. Mol. Sci. 2020, 21, 4794. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134794
Ciecierska A, Motyl T, Sadkowski T. Transcriptomic Profile of Primary Culture of Skeletal Muscle Cells Isolated from Semitendinosus Muscle of Beef and Dairy Bulls. International Journal of Molecular Sciences. 2020; 21(13):4794. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134794
Chicago/Turabian StyleCiecierska, Anna, Tomasz Motyl, and Tomasz Sadkowski. 2020. "Transcriptomic Profile of Primary Culture of Skeletal Muscle Cells Isolated from Semitendinosus Muscle of Beef and Dairy Bulls" International Journal of Molecular Sciences 21, no. 13: 4794. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134794