Gonadal Hormones E2 and P Mitigate Cerebral Ischemia-Induced Upregulation of the AIM2 and NLRC4 Inflammasomes in Rats



Abstract
:1. Introduction
2. Results
2.1. E2 and P Decrease Both Infarct Sizes and Neurological Impairments after Cerebral Ischemia in Rats
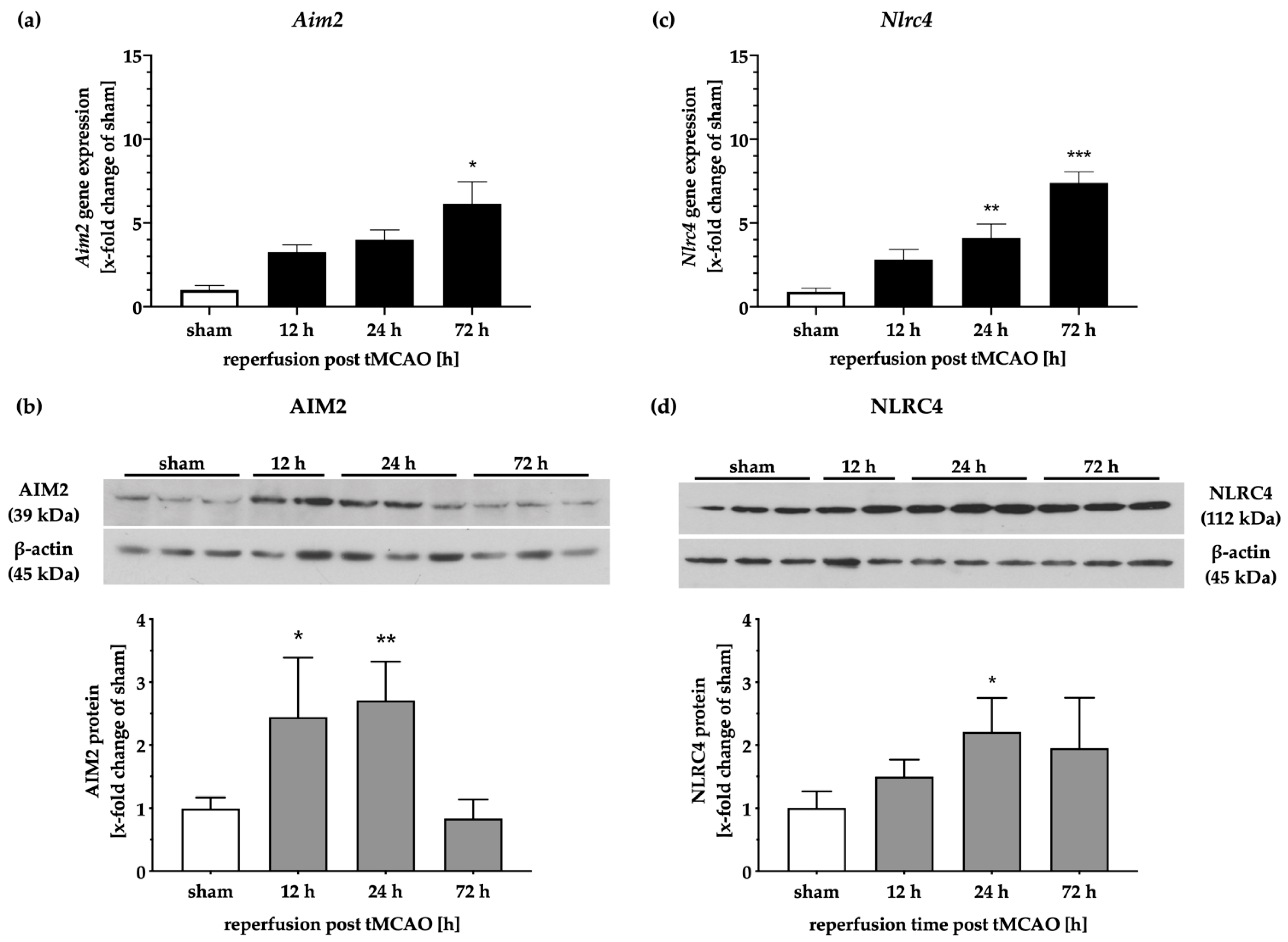
2.2. Gene and Protein Levels of the Inflammasomes AIM2 and NLRC4 Time-Dependently Increase after tMCAO
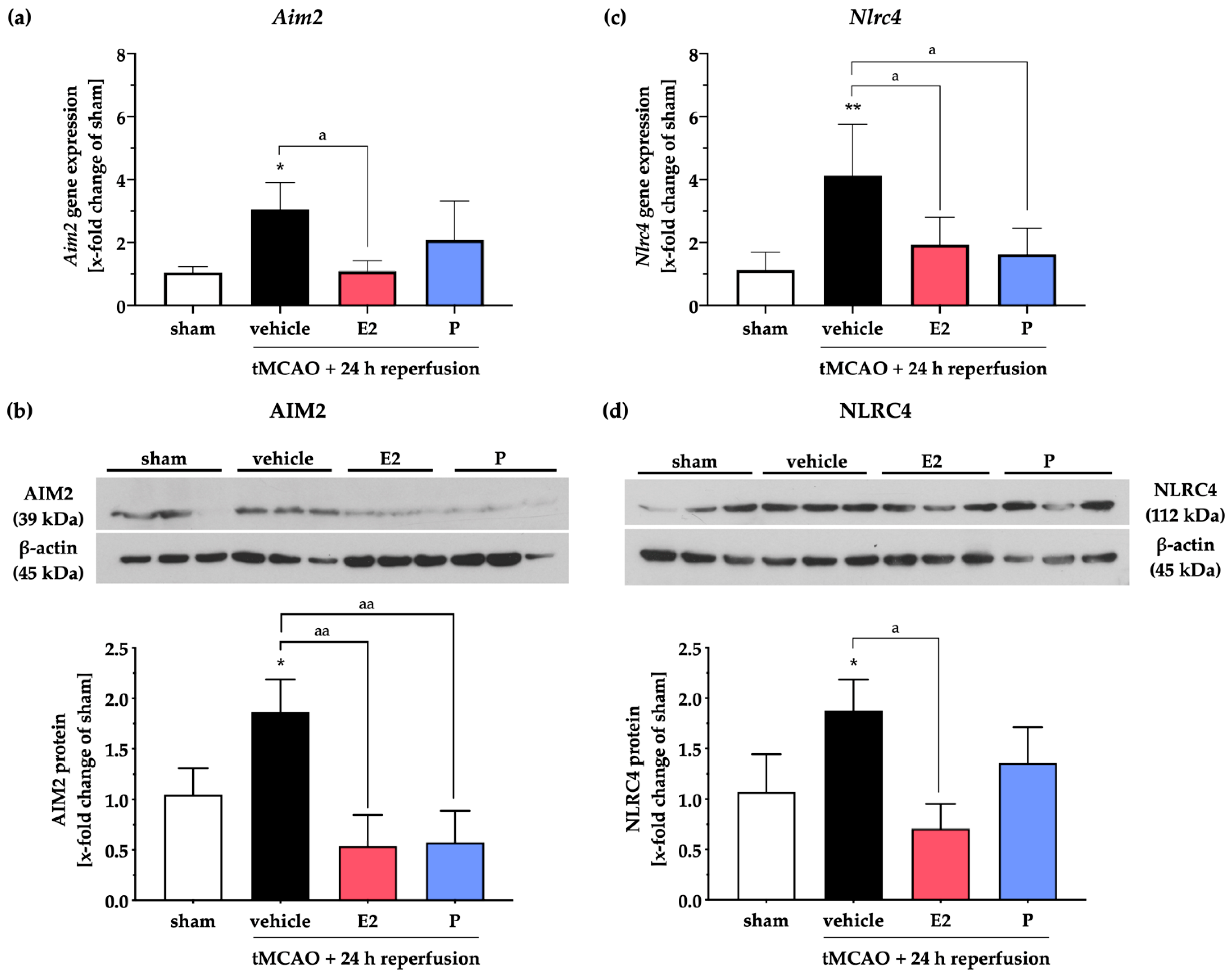
2.3. Steroid Hormones E2 or P Selectively Mitigate the Stroke-Induced Increase of AIM2 and NLRC4 Inflammasomes
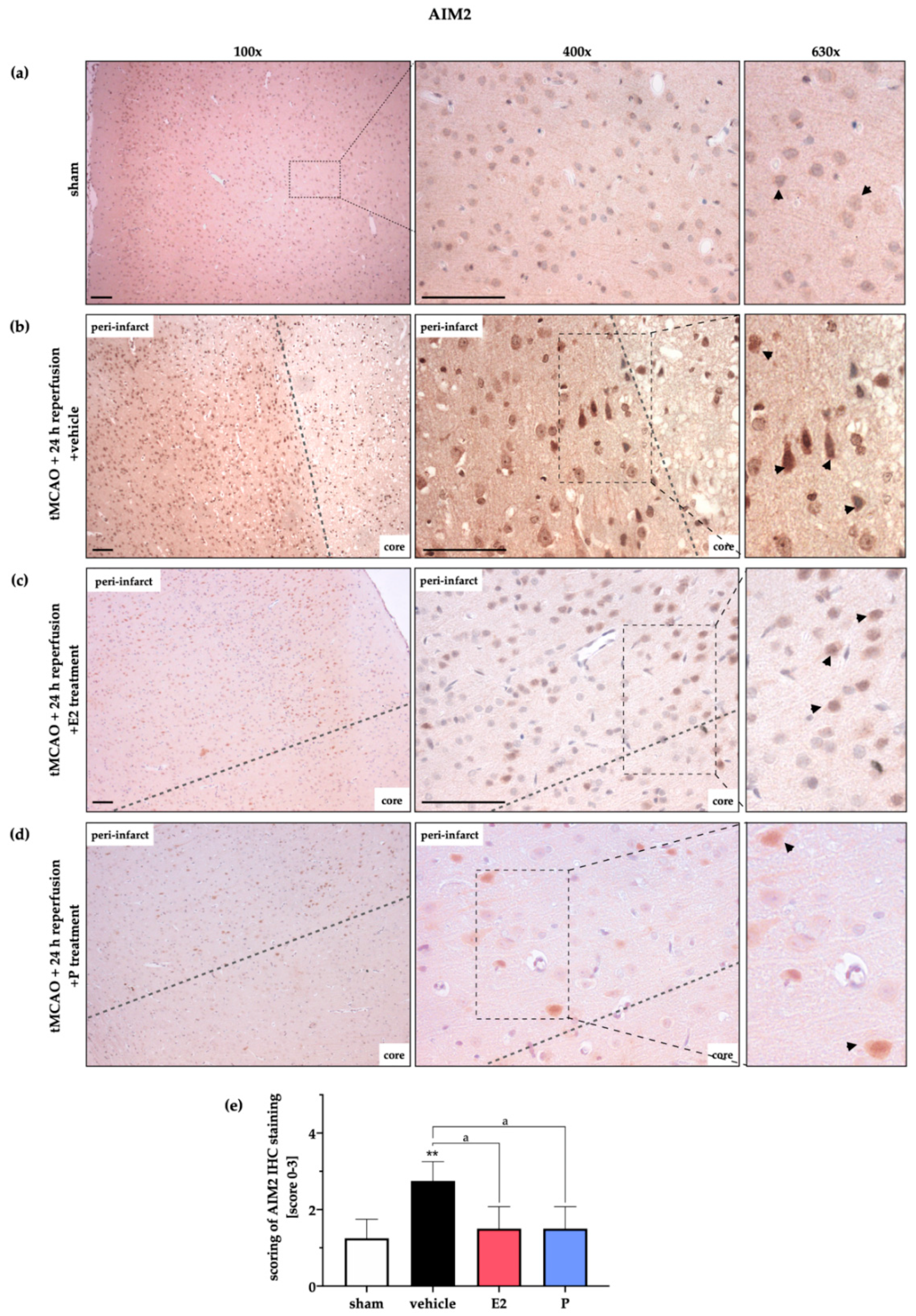
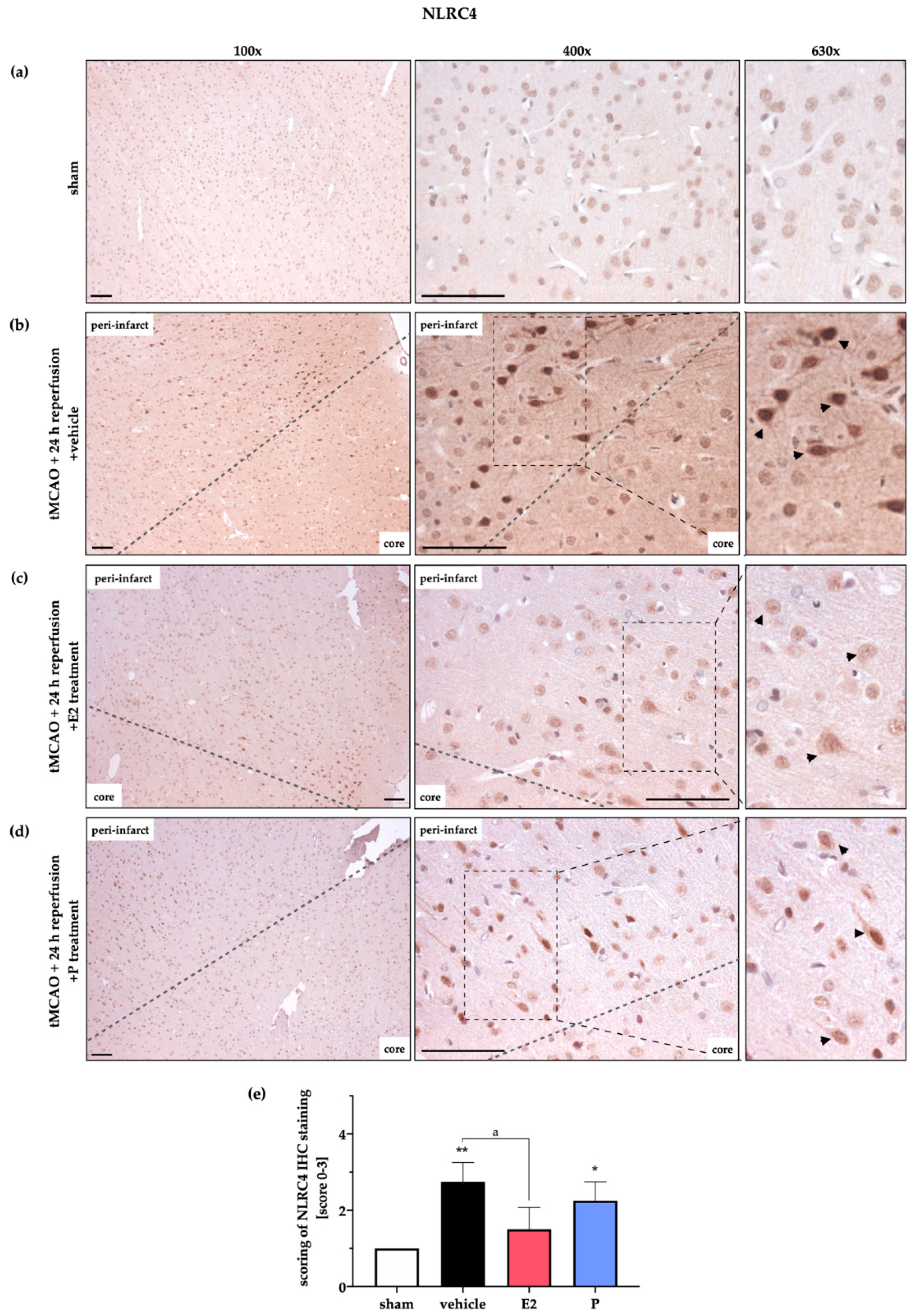
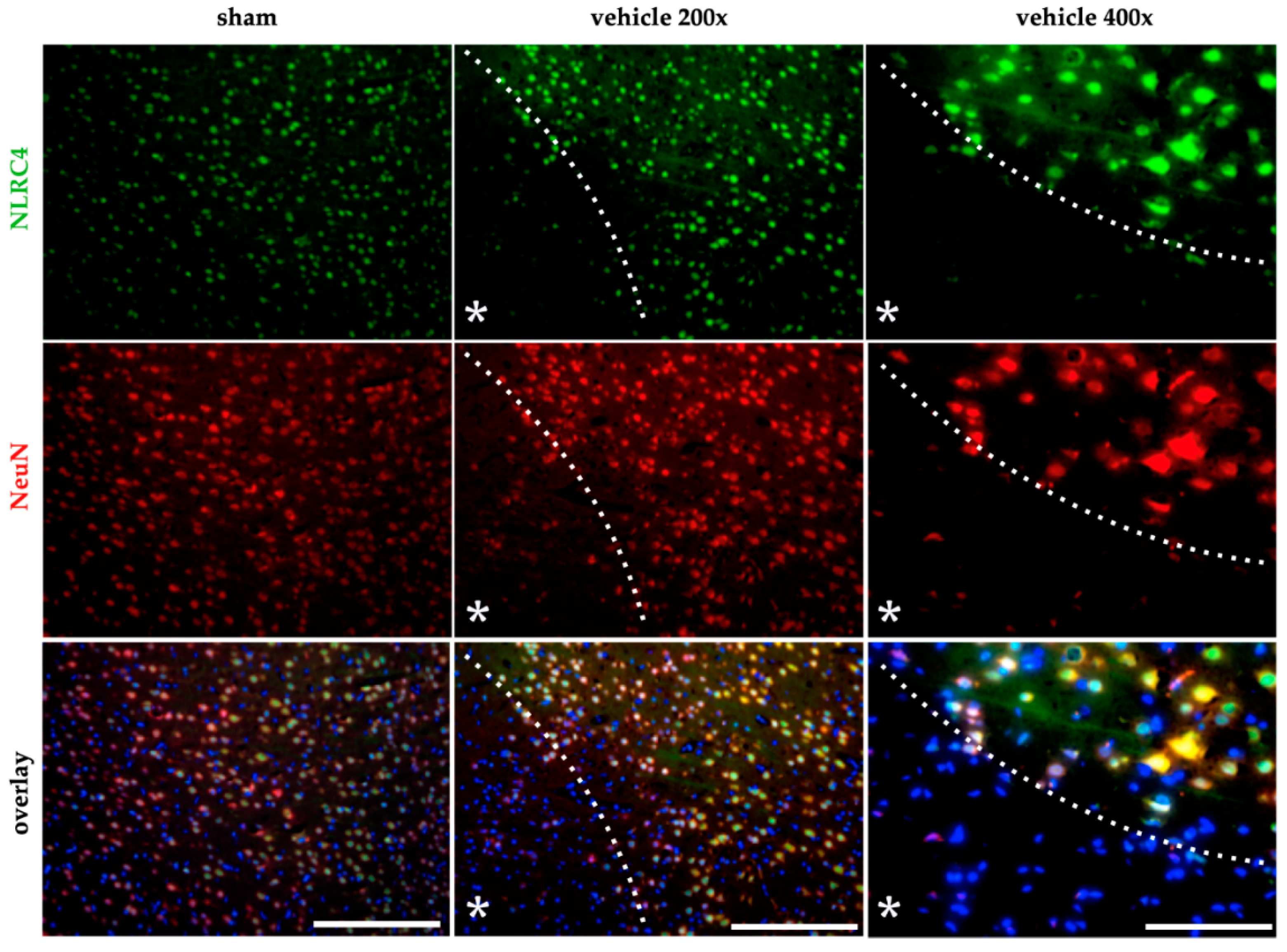
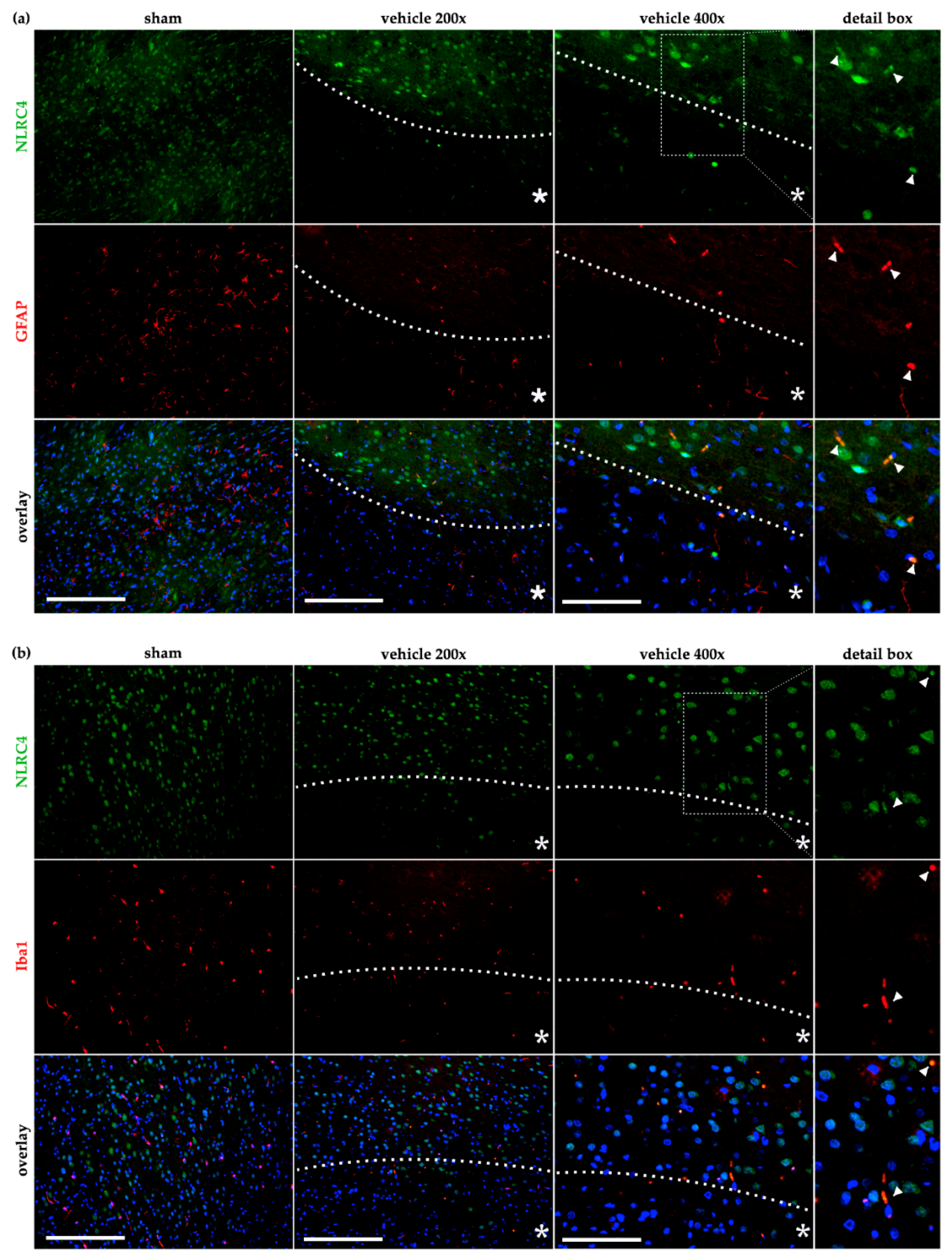
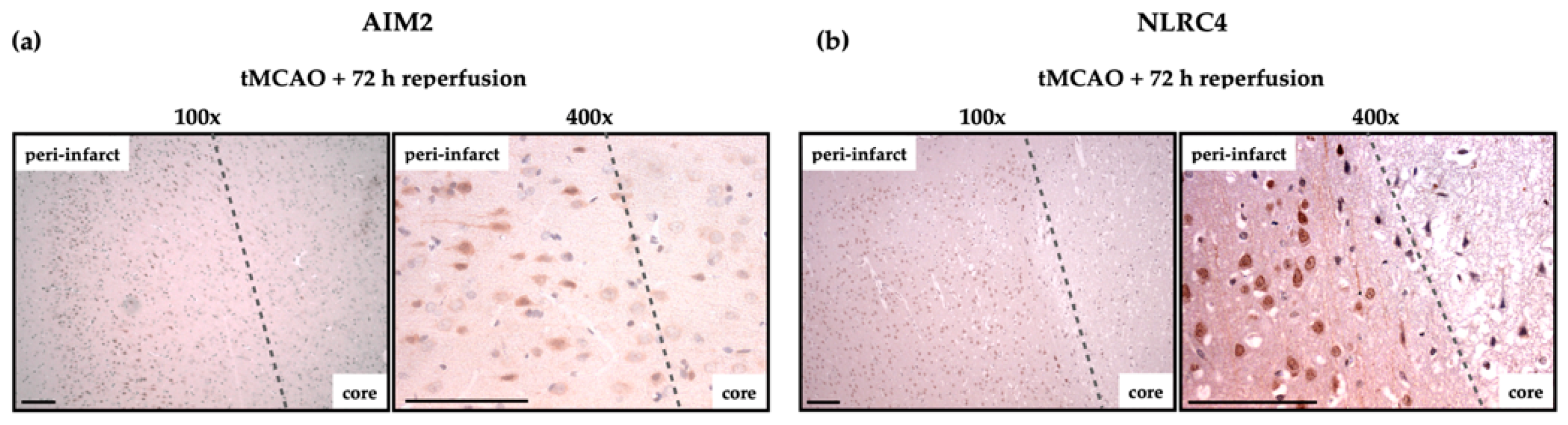
2.4. AIM2 and NLRC4 Inflammasomes Seem to Be Most Abundant in Neurons Under Physiological and Ischemic Conditions
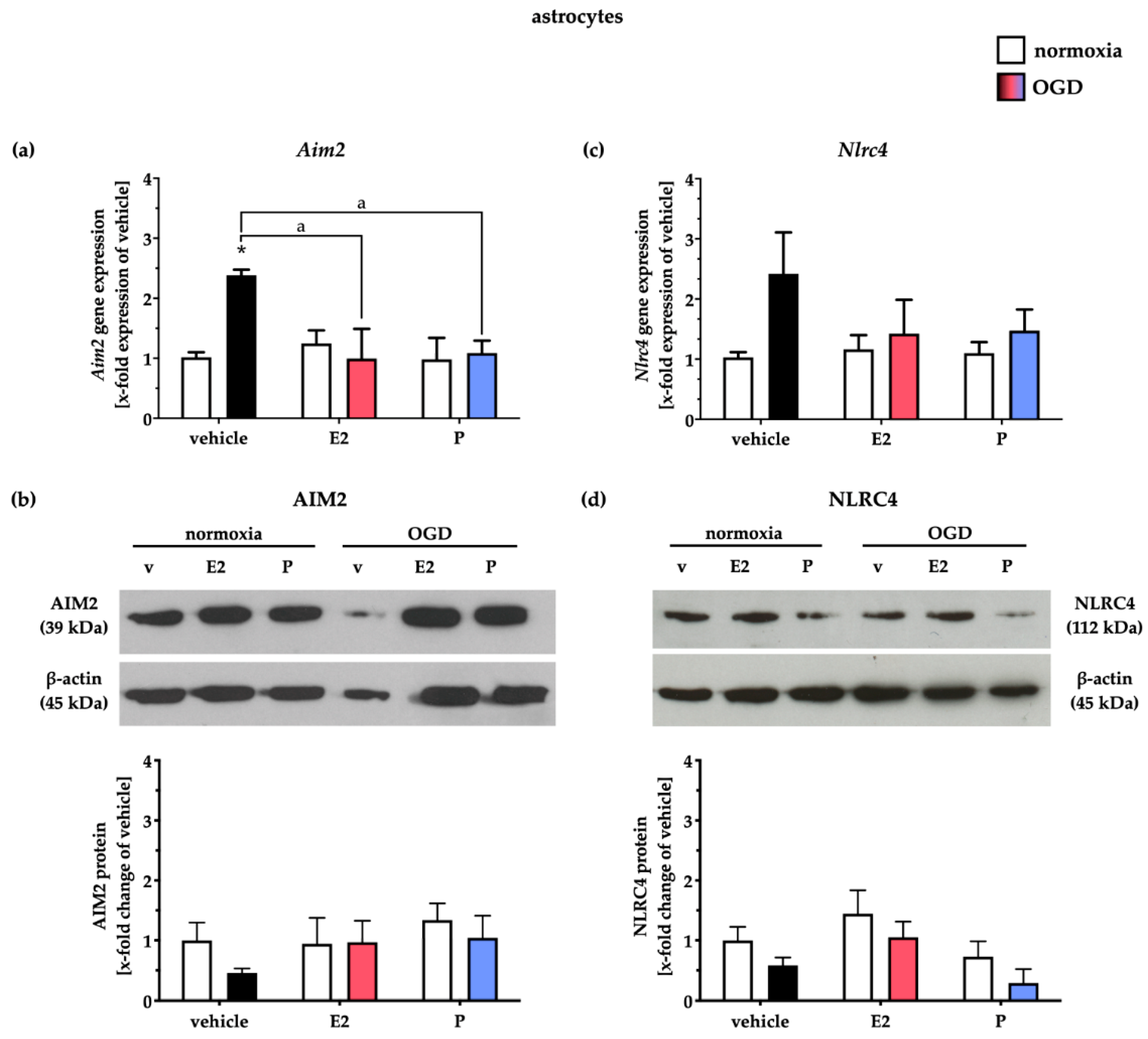
2.5. Administration of E and P Selectively Reduced AIM2 and NLRC4 in Primary Cortical Astrocytes and Microglial Cells after Oxygen–Glucose Deprivation
3. Discussion
4. Materials and Methods
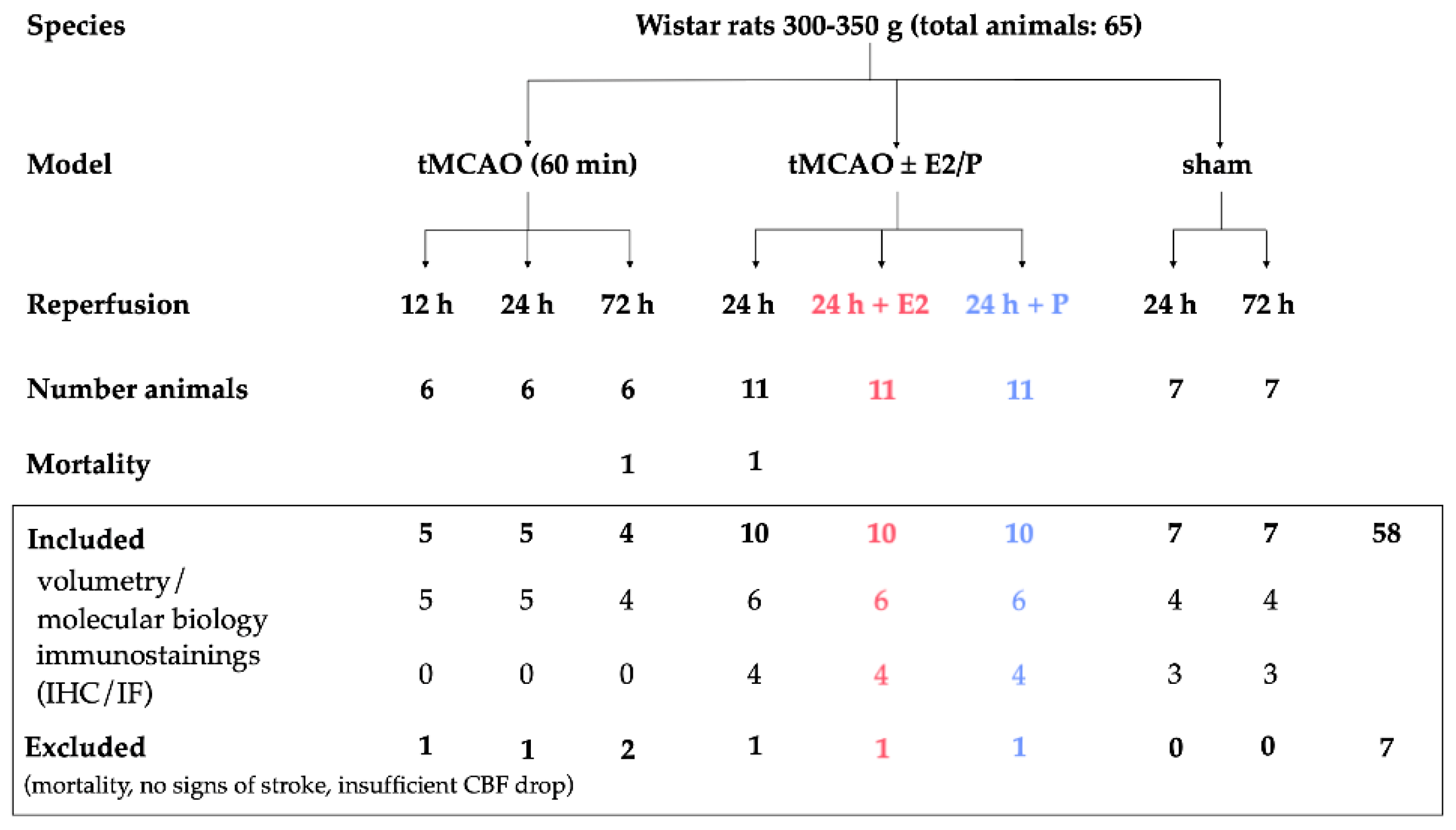
4.1. Animals and Animal Care
4.2. Stroke Surgery (Transient Middle Cerebral Artery Occlusion, tMCAO)
4.3. Behavior Testing
4.4. Hormone Application
4.5. TTC Staining and Tissue Sampling
4.6. Primary Astroglia and Microglia Culture
4.7. Oxygen–Glucose Deprivation (OGD) and Steroid Hormone Treatment
4.8. RNA Extraction
4.9. Semi-Quantitative Real-Time PCR
4.10. Western Blot Analysis
4.11. Immunohistochemistry
4.12. Immunofluorescence Stainings
4.13. Data Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AIM2 | absent in melanoma 2 |
| AIS | acute ischemic stroke |
| ASC | Apoptosis-associated speck-like protein containing a CARD |
| DAMP(s) | damage-associated molecular patterns |
| E2 | 17β-Estradiol |
| GFAP | glial fibrillary acidic protein |
| Iba1 | ionized calcium-binding adapter molecule 1 |
| NACHT | NAIP (neuronal apoptosis inhibitor protein), C2TA (class 2 transcription activator, of the MHC), HET-E (heterokaryon incompatibility) and TP1 (telomerase-associated protein 1) |
| NeuN | Fox-3, Rbfox3, or Hexaribonucleotide Binding Protein-3 |
| NLRC4 | NLR family CARD domain-containing protein 4 |
| NLRP3 | NACHT, LRR and PYD domains-containing protein 3 or cryopyrin |
| OGD | Oxygen–glucose deprivation |
| P | Progesterone |
| PAMP(s) | pathogen-associated molecular patterns |
| tMCAO | transient Middle Cerebral Artery occlusion |
| TTC | 2,3,5-triphenyltetrazolium chloride |
Appendix A


References
- Campbell, B.C.; Meretoja, A.; Donnan, G.A.; Davis, S.M. Twenty-Year History of the Evolution of Stroke Thrombolysis with Intravenous Alteplase to Reduce Long-Term Disability. Stroke 2015, 46, 2341–2346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brouns, R.; De Deyn, P.P. The complexity of neurobiological processes in acute ischemic stroke. Clin. Neurol. Neurosurg. 2009, 111, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Fann, D.Y.-W.; Lee, S.-Y.; Manzanero, S.; Chunduri, P.; Sobey, C.G.; Arumugam, T.V. Pathogenesis of acute stroke and the role of inflammasomes. Ageing Res. Rev. 2013, 12, 941–966. [Google Scholar] [CrossRef] [PubMed]
- Walsh, J.G.; Muruve, D.A.; Power, C. Inflammasomes in the CNS. Nat. Rev. Neurosci. 2014, 15, 84–97. [Google Scholar] [CrossRef]
- Yang, F.; Wang, Z.; Wei, X.; Han, H.; Meng, X.; Zhang, Y.; Shi, W.; Li, F.; Xin, T.; Pang, Q.; et al. NLRP3 deficiency ameliorates neurovascular damage in experimental ischemic stroke. Br. J. Pharm. 2014, 34, 660–667. [Google Scholar] [CrossRef] [Green Version]
- Fann, D.Y.-W.; Santro, T.; Manzanero, S.; Widiapradja, A.; Cheng, Y.-L.; Lee, S.-Y.; Chunduri, P.; Jo, D.-G.; Stranahan, A.M.; Mattson, M.P.; et al. Intermittent fasting attenuates inflammasome activity in ischemic stroke. Exp. Neurol. 2014, 257, 114–119. [Google Scholar] [CrossRef]
- Fann, D.Y.-W.; Lee, S.-Y.; Manzanero, S.; Tang, S.-C.; Gelderblom, M.; Chunduri, P.; Bernreuther, C.; Glatzel, M.; Cheng, Y.-L.; Thundyil, J.; et al. Intravenous immunoglobulin suppresses NLRP1 and NLRP3 inflammasome-mediated neuronal death in ischemic stroke. Cell Death Dis. 2013, 4, e790. [Google Scholar] [CrossRef] [Green Version]
- Lammerding, L.; Slowik, A.; Johann, S.; Beyer, C.; Zendedel, A. Poststroke Inflammasome Expression and Regulation in the Peri-Infarct Area by Gonadal Steroids after Transient Focal Ischemia in the Rat Brain. Neuroendocrinology 2015, 103, 460–475. [Google Scholar] [CrossRef] [PubMed]
- Denes, A.; Coutts, G.; Lénárt, N.; Cruickshank, S.; Pelegrin, P.; Skinner, J.; Rothwell, N.; Allan, S.M.; Brough, D. AIM2 and NLRC4 inflammasomes contribute with ASC to acute brain injury independently of NLRP3. Proc. Natl. Acad. Sci. USA 2015, 112, 4050–4055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.-J.; Zhao, Q.-C.; Xia, M.-X.; Chen, J.; Chen, Y.-T.; Cao, X.; Liu, Y.; Yuan, Z.-Q.; Wang, X.-Y.; Xu, Y. The HDAC3 inhibitor RGFP966 ameliorated ischemic brain damage by downregulating the AIM2 inflammasome. Faseb J. 2019, 34, 648–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habib, P.; Dang, J.; Slowik, A.; Victor, M.; Beyer, C. Hypoxia-induced gene expression of aquaporin-4, cyclooxygenase-2 and hypoxia-inducible factor 1alpha in rat cortical astroglia is inhibited by 17beta-estradiol and progesterone. Neuroendocrinology 2014, 99, 156–167. [Google Scholar] [CrossRef]
- Habib, P.; Dreymueller, D.; Ludwig, A.; Beyer, C.; Dang, J. Sex steroid hormone-mediated functional regulation of microglia-like BV-2 cells during hypoxia. J. Steroid Biochem. Mol. Biol. 2013, 138, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Habib, P.; Slowik, A.; Zendedel, A.; Johann, S.; Dang, J.; Beyer, C. Regulation of Hypoxia-Induced Inflammatory Responses and M1-M2 Phenotype Switch of Primary Rat Microglia by Sex Steroids. J. Mol. Neurosci. 2013, 52, 277–285. [Google Scholar] [CrossRef]
- Slowik, A.; Lammerding, L.; Zendedel, A.; Habib, P.; Beyer, C. Impact of steroid hormones E2 and P on the NLRP3/ASC/Casp1 axis in primary mouse astroglia and BV-2 cells after in vitro hypoxia. J. Steroid. Biochem. Mol. Biol. 2018, 183, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-Garcia, C.; Atif, F.; Yousuf, S.; Sayeed, I.; Neigh, G.N.; Stein, D.G. Progesterone Attenuates Stress-Induced NLRP3 Inflammasome Activation and Enhances Autophagy following Ischemic Brain Injury. Int. J. Mol. Sci. 2020, 21, 3740. [Google Scholar] [CrossRef]
- Thakkar, R.; Wang, R.; Wang, J.; Vadlamudi, R.K.; Brann, D.W. 17 beta-Estradiol Regulates Microglia Activation and Polarization in the Hippocampus Following Global Cerebral Ischemia. Oxid. Med. Cell. Longev. 2018, 2018, 4248526. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.; Wu, H.; Wang, M.; Wang, L.; Zou, H.; Li, S.; Liu, R. Estrogen ameliorates allergic airway inflammation by regulating activation of NLRP3 in mice. Biosci. Rep. 2019, 39, BSR20181117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, L.; Luo, Y.; Tai, R.; Zhang, N. Estrogen receptor beta suppresses inflammation and the progression of prostate cancer. Mol. Med. Rep. 2019, 19, 3555–3563. [Google Scholar] [PubMed]
- Espinosa-Garcia, C.; Sayeed, I.; Yousuf, S.; Atif, F.; Sergeeva, E.G.; Neigh, G.N.; Stein, D.G. Stress primes microglial polarization after global ischemia: Therapeutic potential of progesterone. Brain Behav. Immun. 2017, 66, 177–192. [Google Scholar] [CrossRef]
- Dang, J.; Mitkari, B.; Kipp, M.; Beyer, C. Gonadal steroids prevent cell damage and stimulate behavioral recovery after transient middle cerebral artery occlusion in male and female rats. Brain Behav. Immun. 2011, 25, 715–726. [Google Scholar] [CrossRef]
- Herzog, R.; Zendedel, A.; Lammerding, L.; Beyer, C.; Slowik, A. Impact of 17beta-estradiol and progesterone on inflammatory and apoptotic microRNA expression after ischemia in a rat model. J. Steroid Biochem. Mol. Biol. 2017, 167, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Ulbrich, C.; Zendedel, A.; Habib, P.; Kipp, M.; Beyer, C.; Dang, J. Long-term cerebral cortex protection and behavioral stabilization by gonadal steroid hormones after transient focal hypoxia. J. Steroid Biochem. Mol. Biol. 2012, 131, 10–16. [Google Scholar] [CrossRef]
- Sun, J.; Chi, L.; He, Z.; Gao, Y.; Gao, Y.; Huang, Y.; Nan, G.; Jing, S. NLRP3 inflammasome contributes to neurovascular unit damage in stroke. J. Drug Target. 2019, 27, 866–875. [Google Scholar] [CrossRef] [PubMed]
- Alishahi, M.; Farzaneh, M.; Ghaedrahmati, F.; Nejabatdoust, A.; Sarkaki, A.; Khoshnam, S.E. NLRP3 inflammasome in ischemic stroke: As possible therapeutic target. Int. J. Stroke 2019, 14, 574–591. [Google Scholar] [CrossRef]
- Ismael, S.; Zhao, L.; Nasoohi, S.; Ishrat, T. Inhibition of the NLRP3-inflammasome as a potential approach for neuroprotection after stroke. Sci. Rep. 2018, 8, 5971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemarchand, E.; Barrington, J.; Chenery, A.; Haley, M.; Coutts, G.; Allen, J.E.; Allan, S.M.; Brough, D. Extent of Ischemic Brain Injury After Thrombotic Stroke Is Independent of the NLRP3 (NACHT, LRR and PYD Domains-Containing Protein 3) Inflammasome. Stroke 2019, 50, 1232–1239. [Google Scholar] [CrossRef]
- Schölwer, I.; Habib, P.; Voelz, C.; Rolfes, L.; Beyer, C.; Slowik, A. NLRP3 Depletion Fails to Mitigate Inflammation but Restores Diminished Phagocytosis in BV-2 Cells After In Vitro Hypoxia. Mol. Neurobiol. 2020, 57, 2588–2599. [Google Scholar] [CrossRef]
- Kim, H.; Seo, J.S.; Lee, S.-Y.; Ha, K.-T.; Choi, B.T.; Shin, Y.-I.; Yun, Y.J.; Shin, H.K. AIM2 inflammasome contributes to brain injury and chronic post-stroke cognitive impairment in mice. Brain Behav. Immun. 2020, 87, 765–776. [Google Scholar] [CrossRef]
- Li, Q.; Cao, Y.; Dang, C.; Han, B.; Han, R.; Ma, H.; Hao, J.; Wang, L. Inhibition of double-strand DNA-sensing cGAS ameliorates brain injury after ischemic stroke. Embo Mol. Med. 2020, 12, e11002. [Google Scholar] [CrossRef]
- Liang, J.; Wang, Q.; Li, J.-Q.; Guo, T.; Yu, D. Long non-coding RNA MEG3 promotes cerebral ischemia-reperfusion injury through increasing pyroptosis by targeting miR-485/AIM2 axis. Exp. Neurol. 2020, 325, 113139. [Google Scholar] [CrossRef]
- Poh, L.; Kang, S.-W.; Baik, S.-H.; Ng, G.Y.Q.; She, D.; Balaganapathy, P.; Dheen, S.T.; Magnus, T.; Gelderblom, M.; Sobey, C.G.; et al. Evidence that NLRC4 inflammasome mediates apoptotic and pyroptotic microglial death following ischemic stroke. Brain Behav. Immun. 2019, 75, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Sui, Y.; Bian, L.; Ai, Q.; Yao, Y.; Yu, M.; Gao, H.; Zhang, A.; Fu, X.; Zhong, L.-M.; Lu, D. Gastrodin Inhibits Inflammasome Through the STAT3 Signal Pathways in TNA2 Astrocytes and Reactive Astrocytes in Experimentally Induced Cerebral Ischemia in Rats. Neuromolecular Med. 2019, 21, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Freeman, L.; Guo, H.; David, C.N.; Brickey, W.J.; Jha, S.; Ting, J.P.-Y. NLR members NLRC4 and NLRP3 mediate sterile inflammasome activation in microglia and astrocytes. J. Exp. Med. 2017, 214, 1351–1370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, C.L.; Constantin, D.; Prior, M.J.; Bath, P.M.; Murphy, S. Progesterone suppresses the inflammatory response and nitric oxide synthase-2 expression following cerebral ischemia. Exp. Neurol. 2005, 193, 522–530. [Google Scholar] [CrossRef]
- Jiang, C.; Wang, J.; Li, X.; Liu, C.; Chen, N.; Hao, Y. Progesterone exerts neuroprotective effects by inhibiting inflammatory response after stroke. Inflamm. Res. 2009, 58, 619–624. [Google Scholar] [CrossRef] [PubMed]
- Alfonso-Loeches, S.; Ureña-Peralta, J.R.; Morillo-Bargues, M.J.; La Cruz, J.O.-D.; Guerri, C. Role of mitochondria ROS generation in ethanol-induced NLRP3 inflammasome activation and cell death in astroglial cells. Front. Cell. Neurosci. 2014, 8, 216. [Google Scholar] [CrossRef] [PubMed]
- Cox, D.J.; Field, R.H.; Williams, D.G.; Baran, M.; Bowie, A.G.; Cunningham, C.; Dunne, A. DNA sensors are expressed in astrocytes and microglia in vitro and are up-regulated during gliosis in neurodegenerative disease. Glia 2015, 63, 812–825. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Chan, C. IPAF inflammasome is involved in interleukin-1beta production from astrocytes, induced by palmitate; implications for Alzheimer’s Disease. Neurobiol. Aging 2014, 35, 309–321. [Google Scholar] [CrossRef] [Green Version]
- Scholz, H.; Eder, C. Lysophosphatidylcholine activates caspase-1 in microglia via a novel pathway involving two inflammasomes. J. Neuroimmunol. 2017, 310, 107–110. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.-N.; Guo, X.-Y.; Tang, J.; Ding, S.-Q.; Shen, L.; Wang, R.; Ma, S.-F.; Hu, J.-G.; Lu, H.-Z. Expression and localization of absent in melanoma 2 in the injured spinal cord. Neural Regen. Res. 2019, 14, 542–552. [Google Scholar]
- Kovats, S. Estrogen receptors regulate innate immune cells and signaling pathways. Cell. Immunol. 2015, 294, 63–69. [Google Scholar] [CrossRef] [Green Version]
- Bai, N.; Zhang, Q.; Zhang, W.; Liu, B.; Yang, F.; Brann, D.; Wang, R. G-protein-coupled estrogen receptor activation upregulates interleukin-1 receptor antagonist in the hippocampus after global cerebral ischemia: Implications for neuronal self-defense. J. Neuroinflammation 2020, 17, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Gao, X.; Ding, C.; Lv, Y.; Shen, T.; Ma, G.; Yan, L.; Song, S. Estrogen receptors participate in carcinogenesis signaling pathways by directly regulating NOD-like receptors. Biochem. Biophys. Res. Commun. 2019, 511, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Kilkenny, C.; Browne, W.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Animal Research: Reporting in vivo Experiments—the ARRIVE Guidelines. Br. J. Pharm. 2011, 31, 991–993. [Google Scholar] [CrossRef] [Green Version]
- Garcia, J.H.; Wagner, S.; Liu, K.-F.; Hu, X.-J. Neurological Deficit and Extent of Neuronal Necrosis Attributable to Middle Cerebral Artery Occlusion in Rats. Stroke 1995, 26, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Pawlak, J.; Brito, V.; Kuppers, E.; Beyer, C. Regulation of glutamate transporter GLAST and GLT-1 expression in astrocytes by estrogen. Brain Res. Mol. Brain Res. 2005, 138, 1–7. [Google Scholar] [CrossRef]
- Zendedel, A.; Habib, P.; Dang, J.; Lammerding, L.; Hoffmann, S.; Beyer, C.; Slowik, A. Omega-3 polyunsaturated fatty acids ameliorate neuroinflammation and mitigate ischemic stroke damage through interactions with astrocytes and microglia. J. Neuroimmunol. 2015, 278, 200–211. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.G.; Madden, T. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. Bmc Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [Green Version]
- Clarner, T.; Buschmann, J.P.; Beyer, C.; Kipp, M. Glial Amyloid Precursor Protein Expression is Restricted to Astrocytes in an Experimental Toxic Model of Multiple Sclerosis. J. Mol. Neurosci. 2010, 43, 268–274. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavior Test | Points 1 |
|---|---|
| Spontaneous activity | 3 = rat moving around and exploring at least three walls 2 = rat moving around but not approaching the walls or eventually rising to the cage rim 1 = rat barely moving and no rising 0 = rat not moving at all |
| Forepaw outstretching | 3 = symmetric forepaw outstretching 2 = right side moves, but less outstretching 1 = slight movement of the right forepaw 0 = no movement of the right forepaw |
| Climbing | 3 = easy climbing and tight gripping of the wire 2 = right side is impaired or less tight gripping during climbing 1 = rat fails to climb or tends to circle |
| Body proprioception | 3 = rat reacts by turning head and being startled by the stimulus on both sides 2 = slower reaction of the rat on the right side 1 = no response to the stimulus on the right side |
| Spontaneous walking activity | 3 = rat walking straight ahead 2 = right circling 1 = rat tending to walk toward the right side 0 = rat not moving |
| Sensory function (vibrissae brushing) | 3 = rat turns head towards the stimulus 2 = slow reaction to the stimulus on the right side 1 = no respond to the stimulus on the right side |
| Primer | Sequence | bp 1 | AT 2 [°C] | |
|---|---|---|---|---|
| Aim2 | s | tgctacggagctggtgtttt | 186 | 62 |
| as | actccgtcctgtctgcaatg | |||
| CycloA | s | ggcaaatgctggaccaaacac | 196 | 65 |
| as | ttagagttgtccacagtcggagatg | |||
| Gapdh | s | aacccatcaccatcttccag | 196 | 60 |
| as | gtggttcacacccatcacaa | |||
| Nlrc4 | s | ggctgaggcccacgtataaa | 98 | 60 |
| s | as | ctcctctggctctctggact |
| Antibody | Company | Order Number | WB | IHC | IF |
|---|---|---|---|---|---|
| beta-Actin | Sigma Aldrich, Taufkirchen, Germany | 1:5000 | - | - | |
| NLRC4 | Merck–Millipore, Darmstadt, Germany | 06-1125 | 1:5000 | 1:2000 | 1:200 |
| AIM2 | Bioss antibodies, Woburn, Massachusetts, USA | bs5986R | 1:1000 | 1:300 | - |
| NeuN | Merck–Millipore, Darmstadt, Germany | MAB377 | - | - | 1:500 |
| IBA1 | Merck–Millipore, Darmstadt, Germany | MABN92 | - | - | 1:600 |
| GFAP | Abcam, Cambridge, UK | ab10062 | - | - | 1:1000 |
| Donkey-anti-mouse 594 | ThermoFisher Scientific, Waltham, Massachusetts, USA | A21203 | - | - | 1:500 |
| Donkey-anti-rabbit 488 | ThermoFisher Scientific, Waltham, Massachusetts, USA | A21206 | - | - | 1:500 |
| Donkey-anti-mouse 488 | ThermoFisher Scientific, Waltham, Massachusetts, USA | A21121 | - | - | 1:500 |
| Goat-anti-mouse 546 | ThermoFisher Scientific, Waltham, Massachusetts, USA | A21133 | - | - | 1:500 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Habib, P.; Harms, J.; Zendedel, A.; Beyer, C.; Slowik, A. Gonadal Hormones E2 and P Mitigate Cerebral Ischemia-Induced Upregulation of the AIM2 and NLRC4 Inflammasomes in Rats. Int. J. Mol. Sci. 2020, 21, 4795. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134795
Habib P, Harms J, Zendedel A, Beyer C, Slowik A. Gonadal Hormones E2 and P Mitigate Cerebral Ischemia-Induced Upregulation of the AIM2 and NLRC4 Inflammasomes in Rats. International Journal of Molecular Sciences. 2020; 21(13):4795. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134795
Chicago/Turabian StyleHabib, Pardes, Julie Harms, Adib Zendedel, Cordian Beyer, and Alexander Slowik. 2020. "Gonadal Hormones E2 and P Mitigate Cerebral Ischemia-Induced Upregulation of the AIM2 and NLRC4 Inflammasomes in Rats" International Journal of Molecular Sciences 21, no. 13: 4795. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134795