Nutraceuticals in the Treatment of Pulmonary Arterial Hypertension


 , ,
, ,  , and
, and 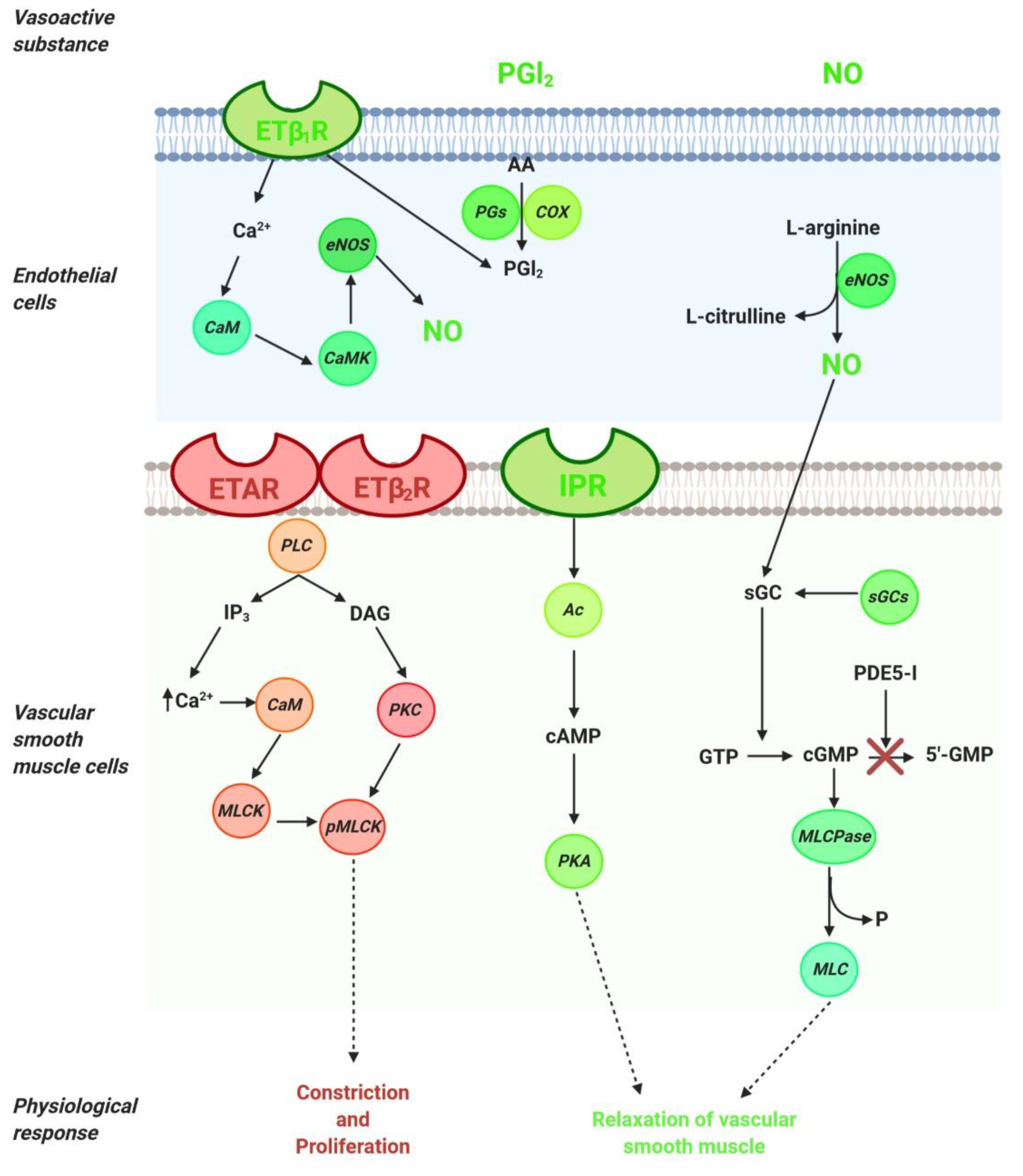{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. An Overview of Molecular Pathways in the Treatment of PAH
2.1. NO Pathway
2.2. ET-1 Pathway
2.3. PGI2 Pathway
3. Nutraceutical in the Treatment of Cardiovascular Disease
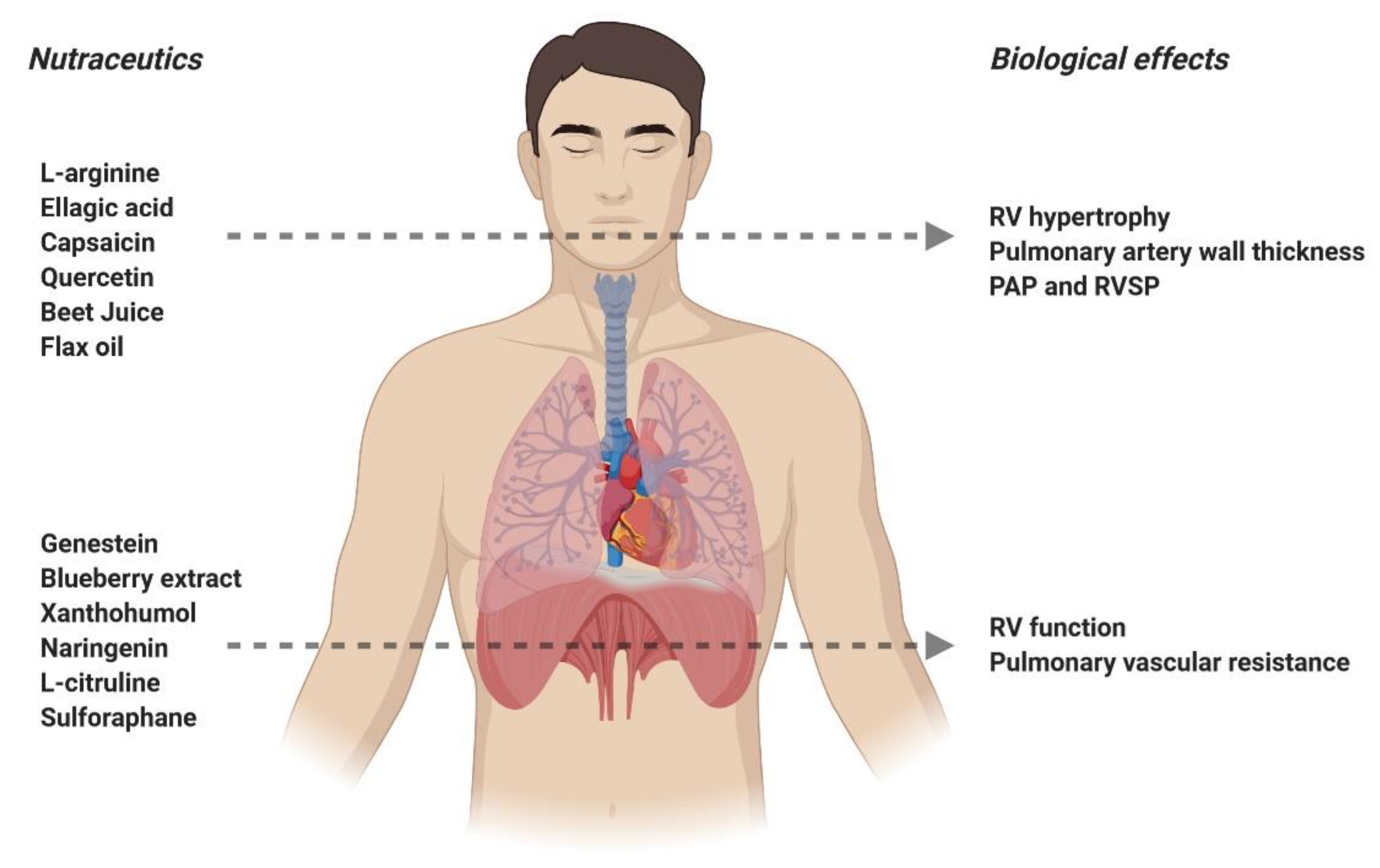
4. Nutraceuticals in the Treatment of Pulmonary Arterial Hypertension
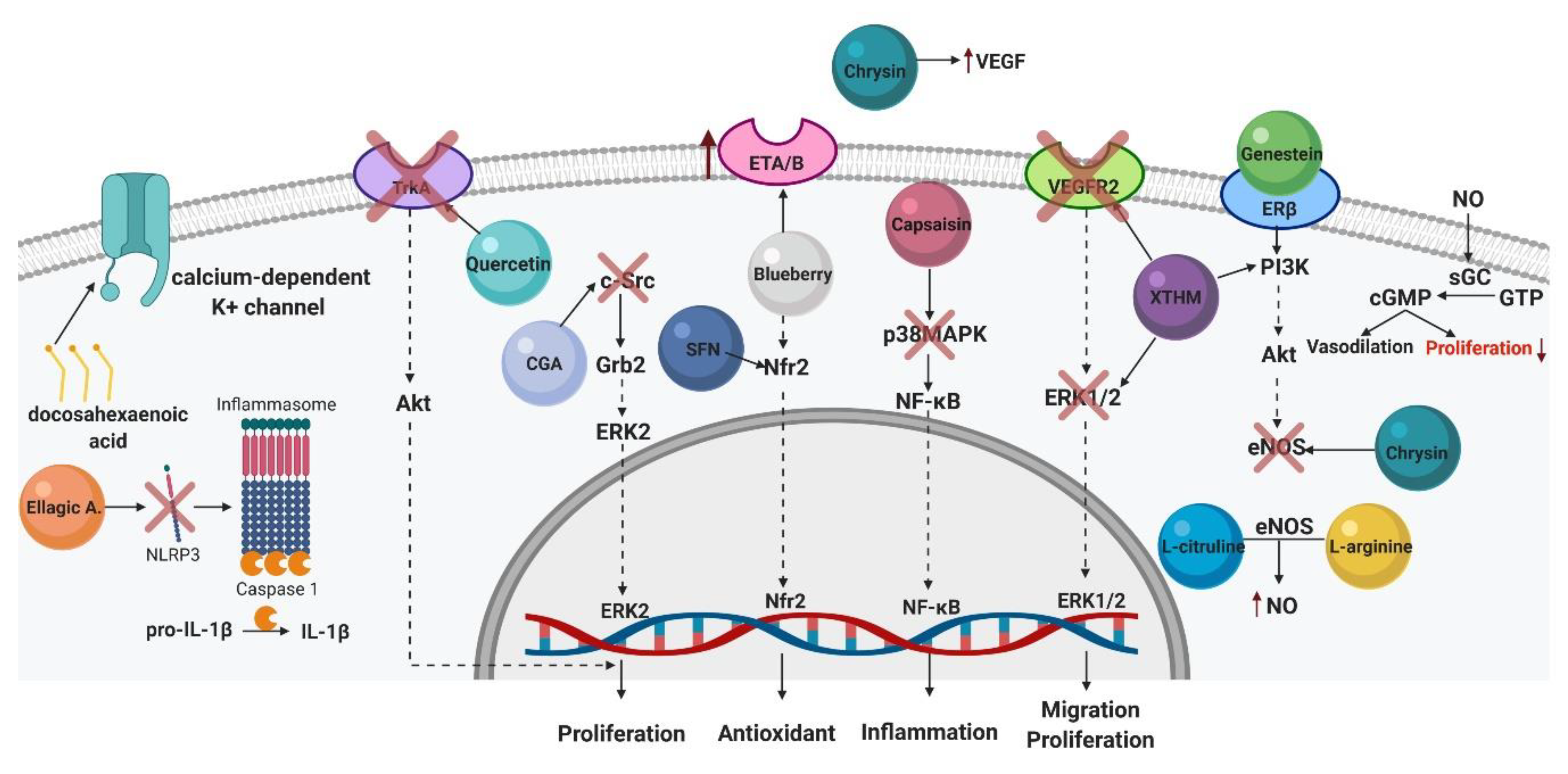
4.1. Genistein
4.2. L-arginine
4.3. Berberine
4.4. Naringenin
4.5. Ellagic Acid
4.6. L-citrulline
4.7. Capsaicin
4.8. Xanthohumol
4.9. Chrysin
4.10. Blueberry Extract
4.11. Quercetin
4.12. Beet Juice
4.13. Chlorogenic Acid
4.14. Fatty Acids
4.15. Sulforaphane
4.16. Allicin
5. Future Directions
- Studies that evaluate the effects of these nutraceuticals in patients with PAH and their intersections with current therapies.
- Determining the dose that exerts beneficial effects in animal models to calculate the amount by which people diagnosed with PAH should obtain from these nutraceuticals.
- Test the use of these nutraceuticals as part of an alternative diet for PAH patients.
- Study the involvement of the signaling pathways responsible for the development of PAH, such as BMPR-2 (Bone morphogenic protein receptor type 2) and ALK-1 (Activin A receptor type II-like kinase-1), in response to nutraceutics.
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 5-HT | Serotonin |
| Akt | Serine threonine protein kinase |
| ALK-1 | Activin A receptor type II-like kinase-1 |
| ATP | Adenosine triphosphate |
| BMPR-2 | Bone morphogenic protein receptor type 2 |
| cGMP | Cyclic guanosine monophosphate |
| c-Src | Proto-oncogene tyrosine-tyrosine-protein kinase Src |
| EC | Endothelial cell |
| eNOS | Endothelial nitric oxide synthase |
| ER | Estrogen receptor α and β |
| ERK | Mitogen-activated protein (MAP) kinase |
| ERK2 | Extracellular-regulated kinase 2 |
| ET | Endothelin |
| Grb2 | Growth factor-bound protein 2 |
| GTP | Guanosine triphosphate |
| IP3 | Inositol trisphosphate |
| MCT | Monocrotaline |
| NFkB | Nuclear factor kappa B |
| NO | Nitric oxide |
| Nrf2 | Nuclear factor erythroid 2-related factor 2 |
| Superoxide anion | |
| P38MAPK | P38 mitogen-activated protein kinases |
| PAH | Pulmonary arterial hypertension |
| PAP | Pulmonary arterial pressure |
| PASMC | Pulmonary artery smooth muscle cells |
| PDE5 | Phosphodiesterase-5 |
| PGI2 | Prostacyclin |
| PH | Pulmonary hypertension |
| PI3K | Phosphatidylinositol 3-kinase |
| PKA | Protein kinase-A |
| PVR | Pulmonary vascular resistance |
| RV | Right vesicle |
| sGC | Guanylate cyclase |
| Trka | Tropomyosin receptor kinase A |
| VEGF | Vascular endothelial growth factor |
| VSMCs | Vascular smooth muscle cells |
| WU | Wood units |
References
- Simonneau, G.; Montani, D.; Celermajer, D.S.; Denton, C.P.; Gatzoulis, M.A.; Krowka, M.; Williams, P.G.; Souza, R. Haemodynamic definitions and updated clinical classification of pulmonary hypertension. Eur. Respir. J. 2019, 53, 1801913. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, W.; Van De Veerdonk, M.C.; Trip, P.; De Man, F.; Heymans, M.W.; Marcus, J.T.; Kawut, S.M.; Bogaard, H.J.; Boonstra, A.; Vonk-Noordegraaf, A. The Right Ventricle Explains Sex Differences in Survival in Idiopathic Pulmonary Arterial Hypertension. Chest 2014, 145, 1230–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatton, N.; Ryan, J.J. Sex differences in response to pulmonary arterial hypertension therapy: Is what’s good for the goose, good for the gander? Chest 2014, 145, 1184–1186. [Google Scholar] [CrossRef] [PubMed]
- Humbert, M.; Guignabert, C.; Bonnet, S.; Dorfmüller, P.; Klinger, J.R.; Nicolls, M.R.; Olschewski, A.J.; Pullamsetti, S.S.; Schermuly, R.T.; Stenmark, K.R. Pathology and pathobiology of pulmonary hypertension: State of the art and research perspectives. Eur. Respir. J. 2019, 53. [Google Scholar] [CrossRef] [Green Version]
- Guignabert, C.; Tu, L.; Girerd, B.; Ricard, N.; Huertas, A.; Montani, D.; Humbert, M. New molecular targets of pulmonary vascular remodeling in pulmonary arterial hypertension: Importance of endothelial communication. Chest 2015, 147, 529–537. [Google Scholar] [CrossRef]
- Huertas, A.; Perros, F.; Tu, L.; Cohen-Kaminsky, S.; Montani, D.; Dorfmüller, P.; Guignabert, C.; Humbert, M. Immune dysregulation and endothelial dysfunction in pulmonary arterial hypertension: A complex interplay. Circulation 2014, 129, 1332–1340. [Google Scholar] [CrossRef]
- Ryan, U.; Ryan, J. Cell biology of pulmonary endothelium. Circulation 1984, 70, III46–III62. [Google Scholar]
- Pulmonary Endothelium: A Dynamic Interface. Available online: https://pubmed.ncbi.nlm.nih.gov/3015468/ (accessed on 12 May 2020).
- Huertas, A.; Guignabert, C.; Barberà, J.A.; Bärtsch, P.; Bhattacharya, J.; Bhattacharya, S.; Bonsignore, M.R.; Dewachter, L.; Dinh-Xuan, A.T.; Dorfmüller, P.; et al. Pulmonary vascular endothelium: The orchestra conductor in respiratory diseases. Eur. Respir. J. 2018, 51, 1700745. [Google Scholar] [CrossRef]
- Parikh, V.; Bhardwaj, A.; Nair, A. Pharmacotherapy for pulmonary arterial hypertension. J. Thorac. Dis. 2019, 11, S1767–S1781. [Google Scholar] [CrossRef]
- Kass, D.A.; Takimoto, E.; Nagayama, T.; Champion, H.C. Phosphodiesterase regulation of nitric oxide signaling. Cardiovasc. Res. 2007, 7, 303–314. Available online: https://0-academic-oup-com.brum.beds.ac.uk/cardiovascres/article-lookup/doi/10.1016/j.cardiores.2007.02.031 (accessed on 13 May 2020). [CrossRef]
- Nagendran, J.; Archer, S.L.; Soliman, D.; Gurtu, V.; Moudgil, R.; Haromy, A.; St Aubin, C.; Webster, L.; Rebeyka, I.M.; Ross, D.B. Phosphodiesterase type 5 is highly expressed in the hypertrophied human right ventricle, and acute inhibition of phosphodiesterase type 5 improves contractility. Circulation 2007, 116, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Corbin, J.D.; Francis, S.H. Molecular Biology and Pharmacology of PDE-5-Inhibitor Therapy for Erectile Dysfunction. J. Androl. 2003, 24, S38–S41. [Google Scholar] [CrossRef] [PubMed]
- Stasch, J.P.; Pacher, P.; Evgenov, O.V. Soluble guanylate cyclase as an emerging therapeutic target in cardiopulmonary disease. Circulation 2011, 123, 2263–2273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stasch, J.-P.; Evgenov, O.V. Soluble Guanylate Cyclase Stimulators in Pulmonary Hypertension. Handb. Exp. Pharmacol. 2013, 218, 279–313. [Google Scholar] [PubMed]
- Galiè, N.; Müller, K.; Scalise, A.V.; Grünig, E. PATENT PLUS: A blinded, randomised and extension study of riociguat plus sildenafil in pulmonary arterial hypertension. Eur. Respir. J. 2015, 45, 1314–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tovar, J.M.; Gums, J.G. Tezosentan in the Treatment of Acute Heart Failure. Ann. Pharmacother. 2003, 37, 1877–1883. [Google Scholar] [CrossRef]
- Rubin, L.J. Endothelin receptor antagonists for the treatment of pulmonary artery hypertension. Life Sci. 2012, 91, 517–521. [Google Scholar] [CrossRef] [Green Version]
- Frumkin, L.R. The pharmacological treatment of pulmonary arterial hypertension. Pharmacol. Rev. 2012, 64, 583–620. [Google Scholar] [CrossRef] [Green Version]
- Hirata, Y.; Emori, T.; Eguchi, S.; Kanno, K.; Imai, T.; Ohta, K.; Marumo, F. Endothelin receptor subtype B mediates synthesis of nitric oxide by cultured bovine endothelial cells. J. Clin. Investig. 1993, 91, 1367–1373. [Google Scholar] [CrossRef] [Green Version]
- Eguchi, S.; Hirata, Y.; Marumo, F. Endothelin subtype B receptors are coupled to adenylate cyclase via inhibitory G protein in cultured bovine endothelial cells. J. Cardiovasc. Pharmacol. 1993, 22, S161–S163. [Google Scholar] [CrossRef]
- Enevoldsen, F.C.; Sahana, J.; Wehland, M.; Grimm, D.; Infanger, M.; Krüger, M. Endothelin Receptor Antagonists: Status Quo and Future Perspectives for Targeted Therapy. J. Clin. Med. 2020, 9, 824. [Google Scholar] [CrossRef] [Green Version]
- Giaid, A.; Yanagisawa, M.; Langleben, D.; Michel, R.P.; Levy, R.; Shennib, H.; Kimura, S.; Masaki, T.; Duguid, W.P.; Stewart, D.J. Expression of Endothelin-1 in the Lungs of Patients with Pulmonary Hypertension. N. Engl. J. Med. 1993, 328, 1732–1739. [Google Scholar] [CrossRef] [PubMed]
- Davie, N.; Haleen, S.J.; Upton, P.D.; Polak, J.M.; Yacoub, M.H.; Morrell, N.W.; Wharton, J. ETA and ETB receptors modulate the proliferation of human pulmonary artery smooth muscle cells. Am. J. Respir. Crit. Care Med. 2002, 165, 398–405. [Google Scholar] [CrossRef] [PubMed]
- Rubin, L.J.; Badesch, D.B.; Barst, R.J.; Galiè, N.; Black, C.M.; Keogh, A.; Pulido, T.; Frost, A.; Roux, S.; Leconte, I.; et al. Bosentan therapy for pulmonary arterial hypertension. N. Engl. J. Med. 2002, 346, 896–903. [Google Scholar] [CrossRef] [PubMed]
- Galiè, N.; Rubin, L.; Hoeper, M.; Jansa, P.; Al-Hiti, H.; Meyer, G.; Chiossi, E.; Kusic-Pajic, A.; Simonneau, G. Treatment of patients with mildly symptomatic pulmonary arterial hypertension with bosentan (EARLY study): A double-blind, randomised controlled trial. Lancet 2008, 371, 2093–2100. [Google Scholar] [CrossRef]
- Pulido, T.; Adzerikho, I.; Channick, R.N.; Delcroix, M.; Galiè, N.; Ghofrani, H.A.; Jansa, P.; Jing, Z.-C.; Le Brun, F.-O.; Mehta, S.; et al. Macitentan and morbidity and mortality in pulmonary arterial hypertension. N. Engl. J. Med. 2013, 369, 809–818. [Google Scholar] [CrossRef] [Green Version]
- Macchia, A.; Mariani, J.; Tognoni, G. Macitentan and pulmonary arterial hypertension. N. Engl. J. Med. 2014, 370, 82. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/24382074 (accessed on 13 May 2020).
- Chaumais, M.C.; Guignabert, C.; Savale, L.; Jaïs, X.; Boucly, A.; Montani, D.; Simonneau, G.; Humbert, M.; Sitbon, O. Clinical Pharmacology of Endothelin Receptor Antagonists Used in the Treatment of Pulmonary Arterial Hypertension. In American Journal of Cardiovascular Drugs; Springer International Publishing: Cham, Switzerland, 2015; Volume 15, pp. 13–26. [Google Scholar]
- Vane, J.; Corin, R.E. Prostacyclin: A vascular mediator. Eur. J. Vasc. Endovasc. Surg. 2003, 26, 571–578. [Google Scholar] [CrossRef] [Green Version]
- Consigny, P.M. Vascular Smooth Muscle Contraction and Relaxation: Pathways and Chemical Modulation. J. Vasc. Interv. Radiol. 1991, 2, 309–317. [Google Scholar] [CrossRef]
- Blindt, R.; Bosserhoff, A.K.; vom Dahl, J.; Hanrath, P.; Schrör, K.; Hohlfeld, T.; Meyer-Kirchrath, J. Activation of IP and EP3 receptors alters cAMP-dependent cell migration. Eur. J. Pharmacol. 2002, 444, 31–37. [Google Scholar] [CrossRef]
- Wharton, J.; Davie, N.; Upton, P.D.; Yacoub, M.H.; Polak, J.M.; Morrell, N.W. Prostacyclin analogues differentially inhibit growth of distal and proximal human pulmonary artery smooth muscle cells. Circulation 2000, 102, 3130–3136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christman, B.W.; Mcpherson, C.D.; Newman, J.H.; King, G.A.; Bernard, G.R.; Groves, B.M.; Loyd, J.E. An Imbalance between the Excretion of Thromboxane and Prostacyclin Metabolites in Pulmonary Hypertension. N. Engl. J. Med. 1992, 327, 70–75. [Google Scholar] [CrossRef]
- Tuder, R.M.; Cool, C.D.; Geraci, M.W.; Wang, J.; Abman, S.H.; Wright, L.; Badesch, D.; Voelkel, N.F. Prostacyclin synthase expression is decreased in lungs from patients with severe pulmonary hypertension. Am. J. Respir. Crit. Care Med. 1999, 159, 1925–1932. [Google Scholar] [CrossRef] [PubMed]
- Barst, R.J.; Rubin, L.J.; Long, W.A.; Mcgoon, M.D.; Rich, S.; Badesch, D.B.; Groves, B.M.; Tapson, V.F.; Bourge, R.C.; Brundage, B.H.; et al. A comparison of continuous intravenous epoprostenol (prostacyclin) with conventional therapy for primary pulmonary hypertension. N. Engl. J. Med. 1996, 334, 296–300. [Google Scholar] [CrossRef]
- Simonneau, G.; Barst, R.J.; Galie, N.; Naeije, R.; Rich, S.; Bourge, R.C.; Keogh, A.; Oudiz, R.; Frost, A.; Blackburn, S.D.; et al. Continuous subcutaneous infusion of treprostinil, a prostacyclin analogue, in patients with pulmonary arterial hypertension: A double-blind, randomized, placebo-controlled trial. Am. J. Respir. Crit. Care Med. 2002, 165, 800–804. [Google Scholar] [CrossRef]
- McLaughlin, V.V.; Benza, R.L.; Rubin, L.J.; Channick, R.N.; Voswinckel, R.; Tapson, V.F.; Robbins, I.M.; Olschewski, H.; Rubenfire, M.; Seeger, W. Addition of inhaled treprostinil to oral therapy for pulmonary arterial hypertension: A randomized controlled clinical trial. J. Am. Coll. Cardiol. 2010, 55, 1915–1922. [Google Scholar] [CrossRef] [Green Version]
- Olschewski, H.; Simonneau, G.; Galiè, N.; Higenbottam, T.; Naeije, R.; Rubin, L.J.; Nikkho, S.; Speich, R.; Hoeper, M.M.; Behr, J.; et al. Inhaled iloprost for severe pulmonary hypertension. N. Engl. J. Med. 2002, 347, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Tapson, V.F.; Jing, Z.C.; Xu, K.F.; Pan, L.; Feldman, J.; Kiely, D.G.; Kotlyar, E.; McSwain, C.S.; Laliberte, K.; Arneson, C.; et al. Oral treprostinil for the treatment of pulmonary arterial hypertension in patients receiving background endothelin receptor antagonist and phosphodiesterase type 5 inhibitor therapy (The FREEDOM-C2 Study): A randomized controlled trial. Chest 2013, 144, 952–958. [Google Scholar] [CrossRef] [PubMed]
- Tapson, V.F.; Sanchez Diaz, C.J.; Bohns Meyer, G.M.; Pulido, T.; Sepulveda, P.; Wang, K.Y.; Deng, C.Q.; Grover, R.; Solum, D.; Ousmanou, A.; et al. Treatment with Oral Treprostinil Delays Time to Clinical Worsening in Patients with Pulmonary Arterial Hypertension—Results from FREEDOM-EV. J. Heart Lung Transplant. 2019, 38, S94–S95. [Google Scholar] [CrossRef]
- Simonneau, G.; Torbicki, A.; Hoeper, M.M.; Delcroix, M.; Karlócai, K.; Galièe, N.; Degano, B.; Bonderman, D.; Kurzyna, M.; Efficace, M.; et al. Selexipag: An oral, selective prostacyclin receptor agonist for the treatment of pulmonary arterial hypertension. Eur. Respir. J. 2012, 40, 874–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klinger, J.R.; Elliott, C.G.; Levine, D.J.; Bossone, E.; Duvall, L.; Fagan, K.; Frantsve-Hawley, J.; Kawut, S.M.; Ryan, J.J.; Rosenzweig, E.B.; et al. Therapy for Pulmonary Arterial Hypertension in Adults: Update of the CHEST Guideline and Expert Panel Report. Chest 2019, 155, 565–586. [Google Scholar] [CrossRef] [PubMed]
- Prior, D.L.; Adams, H.; Williams, T.J. Update on pharmacotherapy for pulmonary hypertension. Med. J. Aust. 2016, 205, 271–276. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, V.V.; Shah, S.J.; Souza, R.; Humbert, M. Management of pulmonary arterial hypertension. J. Am. Coll. Cardiol. 2015, 65, 1976–1997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palthur, M.P.; Sajala Palthur, S.S.; Chitta, S.K. Nutraceuticals: A Conceptual Definition. Int. J. Pharm. Pharm. Sci. 2010, 2, 19–27. [Google Scholar]
- Nasri, H.; Baradaran, A.; Shirzad, H.; Kopaei, M.R. New concepts in nutraceuticals as alternative for pharmaceuticals. Int. J. Prev. Med. 2014, 5, 1487–1499. [Google Scholar]
- Hertog MG, L.; Feskens EJ, M.; Kromhout, D.; Hollman PC, H.; Katan, M.B. Dietary antioxidant flavonoids and risk of coronary heart disease: The Zutphen Elderly Study. Lancet 1993, 342, 1007–1011. [Google Scholar] [CrossRef]
- Khosravi-Boroujeni, H.; Sarrafzadegan, N.; Mohammadifard, N.; Sajjadi, F.; Maghroun, M.; Asgari, S.; Rafieian-kopaei, M.; Azadbakht, L. White rice consumption and CVD risk factors among Iranian population. J. Health Popul. Nutr. 2013, 31, 252–261. [Google Scholar] [CrossRef] [Green Version]
- Khosravi-Boroujeni, H.; Mohammadifard, N.; Sarrafzadegan, N.; Sajjadi, F.; Maghroun, M.; Khosravi, A.; Rafieian-kopaei, M.; Azadbakht, L. Potato consumption and cardiovascular disease risk factors among Iranian population. Int. J. Food Sci. Nutr. 2012, 63, 913–920. [Google Scholar] [CrossRef]
- Galiè, N.; Humbert, M.; Vachiery, J.L.; Gibbs, S.; Lang, I.; Torbicki, A.; Simonneau, G.; Peacock, A.; Noordegraaf, A.V.; Beghetti, M.; et al. 2015 ESC/ERS Guidelines for the diagnosis and treatment of pulmonary hypertension. In European Respiratory Journal; European Respiratory Society: Sheffield, UK, 2015; pp. 903–975. [Google Scholar]
- Henrohn, D.; Björkstrand, K.; Lundberg, J.O.; Granstam, S.O.; Baron, T.; Ingimarsdóttir, I.J.; Hedenström, H.; Malinovschi, A.; Wernroth, M.L.; Jansson, M.; et al. Effects of Oral Supplementation With Nitrate-Rich Beetroot Juice in Patients With Pulmonary Arterial Hypertension—Results From BEET-PAH, an Exploratory Randomized, Double-Blind, Placebo-Controlled, Crossover Study. J. Card. Fail. 2018, 24, 640–653. [Google Scholar] [CrossRef]
- Kupari, M.; Rapola, J. Reversible pulmonary hypertension associated with vitamin C deficiency. Chest 2012, 142, 225–227. [Google Scholar] [CrossRef]
- Ahn, B.H.; Park, H.K.; Cho, H.G.; Lee, H.A.; Lee, Y.M.; Yang, E.K.; Lee, W.J. Estrogen and Enalapril attenuate the Development of Right Ventricular Hypertrophy induced by Monocrotaline in Ovariectomized Rats. J. Korean Med. Sci. 2003, 18, 641–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tofovic, S.P. Estrogens and development of pulmonary hypertension: Lnteraction of estradiol metabolism and pulmonary vascular disease. J. Cardiovasc. Pharmacol. 2010, 56, 696–708. [Google Scholar] [CrossRef] [Green Version]
- Lahm, T.; Albrecht, M.; Fisher, A.J.; Selej, M.; Patel, N.G.; Brown, J.A.; Justice, M.J.; Brown, M.B.; van Demark, M.; Trulock, K.M.; et al. 17β-Estradiol attenuates hypoxic pulmonary hypertension via estrogen receptor-mediated effects. Am. J. Respir. Crit. Care Med. 2012, 185, 965–980. [Google Scholar] [CrossRef] [PubMed]
- Bal, E.; Ilgin, S.; Atli, O.; Ergun, B.; Sirmagul, B. The effects of gender difference on monocrotaline-induced pulmonary hypertension in rats. Hum. Exp. Toxicol. 2013, 32, 766–774. [Google Scholar] [CrossRef] [PubMed]
- Rafikova, O.; Rafikov, R.; Meadows, M.L.; Kangath, A.; Jonigk, D.; Black, S.M. The sexual dimorphism associated with pulmonary hypertension corresponds to a fibrotic phenotype. Pulm. Circ. 2015, 5, 184–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Challa, H.J.; Tadi, P.; Uppaluri, K.R. DASH Diet (Dietary Approaches to Stop Hypertension); StatPearls Publishing: Treasure Island, FL, USA, 2019; pp. 6–9. [Google Scholar]
- Turner, P.V.; Brabb, T.; Pekow, C.; Vasbinder, M.A. Administration of substances to laboratory animals: Routes of administration and factors to consider. J. Am. Assoc. Lab. Anim. Sci. 2011, 50, 600–613. [Google Scholar] [PubMed]
- Babbar, N.; Oberoi, H.S.; Sandhu, S.K. Therapeutic and Nutraceutical Potential of Bioactive Compounds Extracted from Fruit Residues. Crit. Rev. Food Sci. Nutr. 2015, 55, 319–337. [Google Scholar] [CrossRef]
- Anadón, A.; Martínez-Larrañaga, M.R.; Ares, I.; Martínez, M.A. Interactions between Nutraceuticals/Nutrients and Therapeutic Drugs. In Nutraceuticals: Efficacy, Safety and Toxicity; Elsevier Inc.: Amsterdam, The Netherlands, 2016; pp. 855–874. [Google Scholar]
- Xiang, L.; Li, Y.; Deng, X.; Kosanovic, D.; Schermuly, R.T.; Li, X. Natural plant products in treatment of pulmonary arterial hypertension. In Pulmonary Circulation; SAGE Publications Ltd.: Thousand Oaks, VA, USA, 2018; Volume 8. [Google Scholar]
- Jasemi, S.V.; Khazaei, H.; Aneva, I.Y.; Farzaei, M.H.; Echeverría, J. Medicinal Plants and Phytochemicals for the Treatment of Pulmonary Hypertension. Front. Pharmacol. 2020, 11, 145. [Google Scholar] [CrossRef]
- Boots, A.W.; Drent, M.; de Boer, V.C.J.; Bast, A.; Haenen, G.R.M.M. Quercetin reduces markers of oxidative stress and inflammation in sarcoidosis. Clin. Nutr. 2011, 30, 506–512. [Google Scholar] [CrossRef] [Green Version]
- Sawicka, B.H.; Ziarati, P.; Krochmal-Marczak, B.; Skiba, D. Nutraceuticals in food and pharmacy. A Review. Agron. Sci. 2020, 74, 7–31. [Google Scholar] [CrossRef]
- Tresserra-Rimbau, A.; Rimm, E.B.; Medina-Remón, A.; Martínez-González, M.A.; de la Torre, R.; Corella, D.; Salas-Salvadó, J.; Gómez-Gracia, E.; Lapetra, J.; Arós, F.; et al. Inverse association between habitual polyphenol intake and incidence of cardiovascular events in the PREDIMED study. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Hua, C.; Zhao, J.; Wang, H.; Chen, F.; Meng, H.; Chen, L.; Zhang, Q.; Yan, J.; Yuan, L. Apple polyphenol relieves hypoxia-induced pulmonary arterial hypertension via pulmonary endothelium protection and smooth muscle relaxation: In vivo and in vitro studies. Biomed. Pharmacother. 2018, 107, 937–944. [Google Scholar] [CrossRef] [PubMed]
- Hsu, W.L.; Lin, Y.C.; Jeng, J.R.; Chang, H.Y.; Chou, T.C. Baicalein Ameliorates Pulmonary Arterial Hypertension Caused by Monocrotaline through Downregulation of ET-1 and ETAR in Pneumonectomized Rats. Am. J. Chin. Med. 2018, 46, 769–783. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Gu, Y.; Liu, P.; Jiang, X.; Yu, W.; Ye, P.; Chao, Y.; Yang, H.; Zhu, L.; Zhou, L.; et al. Berberine attenuates pulmonary arterial hypertension via protein phosphatase 2A signaling pathway both in vivo and in vitro. J. Cell. Physiol. 2018, 233, 9750–9762. [Google Scholar] [CrossRef]
- Li, T.; Luo, X.J.; Wang, E.L.; Li, N.S.; Zhang, X.J.; Song, F.L.; Yang, J.F.; Liu, B.; Peng, J. Magnesium lithospermate B prevents phenotypic transformation of pulmonary arteries in rats with hypoxic pulmonary hypertension through suppression of NADPH oxidase. Eur. J. Pharmacol. 2019, 847, 32–41. [Google Scholar] [CrossRef]
- Gao, H.; Chen, C.; Huang, S.; Li, B. Quercetin attenuates the progression of monocrotaline-induced pulmonary hypertension in rats. J. Biomed. Res. 2012, 26, 98–102. [Google Scholar] [CrossRef] [Green Version]
- Guan, Z.; Shen, L.; Liang, H.; Yu, H.; Hei, B.; Meng, X.; Yang, L. Resveratrol inhibits hypoxia-induced proliferation and migration of pulmonary artery vascular smooth muscle cells by inhibiting the phosphoinositide 3-kinase/protein kinase B signaling pathway. Mol. Med. Rep. 2017, 16, 1653–1660. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, N.; Akhtar, J.; Singh, S.P.; Ahsan, F. An Overview on Genistein and its Various Formulations. Drug Res. 2019, 69, 305–313. [Google Scholar] [CrossRef]
- Liggins, J.; Mulligan, A.; Runswick, S.; Bingham, S.A. Daidzein and genistein content of cereals. Eur. J. Clin. Nutr. 2002, 56, 961–966. [Google Scholar] [CrossRef]
- Si, H.; Liu, D. Isoflavone genistein protects human vascular endothelial cells against tumor necrosis factor-a-induced apoptosis through the p38β mitogen-activated protein kinase. Apoptosis 2009, 14, 66–76. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.W.; Lee, J.H. Effects of oven-drying, roasting, and explosive puffing process on isoflavone distributions in soybeans. Food Chem. 2009, 112, 316–320. [Google Scholar] [CrossRef]
- Chung, I.M.; Seo, S.H.; Ahn, J.K.; Kim, S.H. Effect of processing, fermentation, and aging treatment to content and profile of phenolic compounds in soybean seed, soy curd and soy paste. Food Chem. 2011, 127, 960–967. [Google Scholar] [CrossRef]
- Aguiar, C.L.; Haddad, R.; Eberlin, M.N.; Carrão-Panizzi, M.C.; Tsai, S.M.; Park, Y.K. Thermal behavior of malonylglucoside isoflavones in soybean flour analyzed by RPHPLC/DAD and eletrospray ionization mass spectrometry. LWT Food Sci. Technol. 2012, 48, 114–119. [Google Scholar] [CrossRef]
- Jackson, C.J.C.; Dini, J.P.; Lavandier, C.; Rupasinghe, H.P.V.; Faulkner, H.; Poysa, V.; Buzzell, D.; DeGrandis, S. Effects of processing on the content and composition of isoflavones during manufacturing of soy beverage and tofu. Process Biochem. 2002, 37, 1117–1123. [Google Scholar] [CrossRef]
- Pelbagai, K.; Memasak, K.; Isoflavon, K.; Berasaskan, H.; Di Malaysia, S.; Haron, H.; Shaari, T.; Keng, C.B. Effects of Different Cooking Methods on Isoflavone Content in Malaysian Soy-Based Dishes. Sains Malays. 2016, 45, 1329–1335. [Google Scholar]
- Day, A.J.; Cañada, F.J.; Díaz, J.C.; Kroon, P.A.; McLauchlan, R.; Faulds, C.B.; Plumb, G.W.; Morgan, M.R.; Williamson, G. Dietary flavonoid and isoflavone glycosides are hydrolysed by the lactase site of lactase phlorizin hydrolase. FEBS Lett. 2000, 468, 166–170. [Google Scholar] [CrossRef] [Green Version]
- Kwon, S.H.; Kang, M.J.; Huh, J.S.; Ha, K.W.; Lee, J.R.; Lee, S.K.; Lee, B.S.; Han, I.H.; Lee, M.S.; Lee, M.W.; et al. Comparison of oral bioavailability of genistein and genistin in rats. Int. J. Pharm. 2007, 337, 148–154. [Google Scholar] [CrossRef]
- Nielsen, I.L.F.; Williamson, G. Review of the factors affecting bioavailability of soy isoflavones in humans. Nutr. Cancer 2007, 57, 1–10. [Google Scholar] [CrossRef]
- Setchell, K.D.R.; Brown, N.M.; Desai, P.; Zimmer-Nechemias, L.; Wolfe, B.E.; Brashear, W.T.; Kirschner, A.S.; Cassidy, A.; Heubi, J.E. Bioavailability of Pure Isoflavones in Healthy Humans and Analysis of Commercial Soy Isoflavone Supplements. J. Nutr. 2001, 131, 1362S–1375S. [Google Scholar] [CrossRef] [Green Version]
- Setchell, K.D.R.; Brown, N.M.; Desai, P.B.; Zimmer-Nechimias, L.; Wolfe, B.; Jakate, A.S.; Creutzinger, V.; Heubi, J.E. Bioavailability, Disposition, and Dose-Response Effects of Soy Isoflavones When Consumed by Healthy Women at Physiologically Typical Dietary Intakes. J. Nutr. 2003, 133, 1027–1035. [Google Scholar] [CrossRef] [Green Version]
- Rassu, G.; Porcu, E.P.; Fancello, S.; Obinu, A.; Senes, N.; Galleri, G.; Migheli, R.; Gavini, E.; Giunchedi, P. Intranasal delivery of genistein-loaded nanoparticles as a potential preventive system against neurodegenerative disorders. Pharmaceutics 2019, 11, 8. [Google Scholar] [CrossRef] [Green Version]
- Deodato, B.; Altavilla, D.; Squadrito, G.; Campo, G.M.; Arlotta, M.; Minutoli, L.; Saitta, A.; Cucinotta, D.; Calapai, G.; Caputi, A.P.; et al. Cardioprotection by the phytoestrogen genistein in experimental myocardial ischaemia-reperfusion injury. Br. J. Pharmacol. 1999, 128, 1683–1690. [Google Scholar] [CrossRef] [Green Version]
- Tissier, R.; Waintraub, X.; Couvreur, N.; Gervais, M.; Bruneval, P.; Mandet, C.; Zini, R.; Enriquez, B.; Berdeaux, A.; Ghaleh, B. Pharmacological postconditioning with the phytoestrogen genistein. J. Mol. Cell. Cardiol. 2007, 42, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Ferreira, D. Genistein. Phytochemistry 2002, 60, 205–211. [Google Scholar] [CrossRef]
- Wang, T.T.Y.; Sathyamoorthy, N.; Phang, J.M. Molecular effects of genistein on estrogen receptor mediated pathways. Carcinogenesis 1996, 17, 271–275. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.; Song, J.; Mark, C.; Eyster, K. Understanding the Cardiovascular Actions of Soy Isoflavones: Potential Novel Targets for Antihypertensive Drug Development. Cardiovasc. Hematol. Disord. Targets 2008, 8, 297–312. [Google Scholar] [CrossRef] [PubMed]
- Siow, R.C.M.; Li, F.Y.L.; Rowlands, D.J.; de Winter, P.; Mann, G.E. Cardiovascular targets for estrogens and phytoestrogens: Transcriptional regulation of nitric oxide synthase and antioxidant defense genes. Free Radic. Biol. Med. 2007, 42, 909–925. [Google Scholar] [CrossRef]
- Matori, H.; Umar, S.; Nadadur, R.D.; Sharma, S.; Partow-Navid, R.; Afkhami, M.; Amjedi, M.; Eghbali, M. Genistein, a soy phytoestrogen, reverses severe pulmonary hypertension and prevents right heart failure in rats. Hypertension 2012, 60, 425–430. [Google Scholar] [CrossRef]
- Zheng, Z.; Yu, S.; Zhang, W.; Peng, Y.; Pu, M.; Kang, T.; Zeng, J.; Yu, Y.; Li, G. Genistein attenuates monocrotaline-induced pulmonary arterial hypertension in rats by activating PI3K/Akt/eNOS signaling. Histol. Histopathol. 2017, 32, 35–41. [Google Scholar]
- Yang, Y.; Gao, M.; Wu, Z.; Guo, Y. Genistein attenuates low temperature induced pulmonary hypertension in broiler chicks by modulating endothelial function. Eur. J. Pharmacol. 2010, 649, 242–248. [Google Scholar] [CrossRef]
- Zhang, M.; Wu, Y.; Wang, M.; Wang, Y.; Tausif, R.; Yang, Y. Genistein rescues hypoxia-induced pulmonary arterial hypertension through estrogen receptor and β-adrenoceptor signaling. J. Nutr. Biochem. 2018, 58, 110–118. [Google Scholar] [CrossRef]
- Kim, H.; Do, H.W.; Chung, H. A comparison of the essential amino acid content and the retention rate by chicken part according to different cooking methods. Korean J. Food Sci. Anim. Resour. 2017, 37, 626–634. [Google Scholar] [CrossRef] [Green Version]
- Castillo, L.; DeRojas, T.C.; Chapman, T.E.; Vogt, J.; Burke, J.F.; Tannenbaum, S.R.; Young, V.R. Splanchnic metabolism of dietary arginine in relation to nitric oxide synthesis in normal adult man. Proc. Natl. Acad. Sci. USA 1993, 90, 193–197. [Google Scholar] [CrossRef] [Green Version]
- Bode-Böger, S.M.; Böger, R.H.; Galland, A.; Tsikas, D.; Frölich, J.C. L-arginine-induced vasodilation in healthy humans: Pharmacokinetic-pharmacodynamic relationship. Br. J. Clin. Pharmacol. 1998, 46, 489–497. [Google Scholar] [CrossRef] [Green Version]
- Böger, R.H.; Tsikas, D.; Bode-Böger, S.M.; Phivthong-ngam, L.; Schwedhelm, E.; Frölich, J.C. Hypercholesterolemia impairs basal nitric oxide synthase turnover rate: A study investigating the conversion of L-[guanidino-15N2]-arginine to 15N-labeled nitrate by gas chromatography-mass spectrometry. Nitric Oxide Biol. Chem. 2004, 11, 1–8. [Google Scholar] [CrossRef]
- Closs, E.I.; Simon, A.; Vékony, N.; Rotmann, A. Plasma Membrane Transporters for Arginine. J. Nutr. 2004, 134, 2752S–2759S. [Google Scholar] [CrossRef]
- Li, C.; Huang, W.; Harris, M.B.; Goolsby, J.M.; Venema, R.C. Interaction of the endothelial nitric oxide synthase with the CAT-1 arginine transporter enhances NO by a mechanism not involving arginine transport. Biochem. J. 2005, 386, 567–574. [Google Scholar] [CrossRef]
- Meyerhoff, M.E.; Howland, J.L. Methods in nitric oxide research. Biochem. Educ. 1997, 25, 147–160. [Google Scholar] [CrossRef]
- Thiemermann, C. The Role of the L-Arginine: Nitric Oxide Pathway in Circulatory Shock. Adv. Pharmacol. 1994, 28, 45–79. [Google Scholar]
- Li, H.; Förstermann, U. Nitric Oxide in the Pathogenesis of Vascular Disease. J. Pathol. 2000, 190, 244–254. [Google Scholar] [CrossRef]
- Böger, R.H. The pharmacodynamics of L-arginine. In Alternative Therapies in Health and Medicine; InnoVision Communications: Houston, TX, USA, 2014; Volume 20, pp. 48–54. [Google Scholar]
- Bode-Boger, S.M.; Boger, R.H.; Creutzig, A.; Tsikas, D.; Gutzki, F.M.; Alexander, K.; Frölich, J.C. L-Arginine infusion decreases peripheral arterial resistance and inhibits platelet aggregation in healthy subjects. Clin. Sci. 1994, 87, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Böger, R.H.; Mügge, A.; Bode-Böger, S.M.; Heinzel, D.; Höper, M.M.; Frölich, J.C. Differential systemic and pulmonary hemodynamic effects of L-arginine in patients with coronary artery disease or primary pulmonary hypertension. Int. J. Clin. Pharmacol. Ther. 1996, 34, 323–328. [Google Scholar] [PubMed]
- Souza-Silva, A.R.; Dias-Junior, C.A.; Uzuelli, J.A.; Moreno, H.; Evora, P.R.; Tanus-Santos, J.E. Hemodynamic effects of combined sildenafil and L-arginine during acute pulmonary embolism-induced pulmonary hypertension. Eur. J. Pharmacol. 2005, 524, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.B.; Kempf, A.; Collins, C.M.; Long, G.M.; Owens, M.; Gupta, S.; Hellman, Y.; Wong, V.; Farber, M.; Lahm, T. A prescribed walking regimen plus arginine supplementation improves function and quality of life for patients with pulmonary arterial hypertension: A pilot study. Pulm. Circ. 2018, 8. [Google Scholar] [CrossRef] [Green Version]
- Ou, Z.J.; Wei, W.; Huang, D.D.; Luo, W.; Luo, D.; Wang, Z.P.; Zhang, X.; Ou, J.-S. L-Arginine restores endothelial nitric oxide synthase-coupled activity and attenuates monocrotaline-induced pulmonary artery hypertension in rats. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E1131–E1139. [Google Scholar] [CrossRef]
- Kasai, K.; Hattori, Y.; Banba, N.; Hattori, S.; Motohashi, S.; Shimoda, S.I.; Nakanishi, N.; Gross, S.S. Induction of tetrahydrobiopterin synthesis in rat cardiac myocytes: Impact on cytokine-induced NO generation. Am. J. Physiol. Heart Circ. Physiol. 1997, 273, H665–H672. [Google Scholar] [CrossRef]
- Caliceti, C.; Rizzo, P.; Cicero, A.F.G. Potential benefits of berberine in the management of perimenopausal syndrome. Oxid. Med. Cell. Longev. 2015, 2015, 723093. [Google Scholar] [CrossRef]
- Barbosa, V.A.; Luciano, T.F.; Marques, S.O.; Vitto, M.F.; Souza, D.R.; Silva, L.A.; Santos, J.P.; Moreira, J.C.; Dal-Pizzol, F.; Lira, F.S.; et al. Acute exercise induce endothelial nitric oxide synthase phosphorylation via Akt and AMP-activated protein kinase in aorta of rats: Role of reactive oxygen species. Int. J. Cardiol. 2013, 167, 2983–2988. [Google Scholar] [CrossRef] [Green Version]
- Morrow, V.A.; Foufelle, F.; Connell, J.M.C.; Petrie, J.R.; Gould, G.W.; Salt, I.P. Direct activation of AMP-activated protein kinase stimulates nitric-oxide synthesis in human aortic endothelial cells. J. Biol. Chem. 2003, 278, 31629–31639. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Wang, C.M.; Li, J.; Meng, Z.J.; Wei, S.N.; Bucala, R.; Li, J.; Li, Y.L.; Chen, L. Berberine protects against palmitate-induced endothelial dysfunction: Involvements of upregulation of AMPK and eNOS and downregulation of NOX4. Mediat. Inflamm. 2013, 2013, 1–8. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, Y.; Lam, K.S.L.; Li, Y.; Wong, W.T.; Ye, H.; Lau, C.-W.; Vanhoutte, P.M.; Xu, A. Berberine prevents hyperglycemia-induced endothelial injury and enhances vasodilatation via adenosine monophosphate-activated protein kinase and endothelial nitric oxide synthase. Cardiovasc. Res. 2009, 82, 484–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.T.; Cui, B.; Dai, D.Z.; Su, W. CPU 86017, p-chlorobenzyltetrahydroberberine chloride, attenuates monocrotaline-induced pulmonary hypertension by suppressing endothelin pathway. Acta Pharmacol. Sin. 2005, 26, 1309–1316. [Google Scholar] [CrossRef]
- Yu, W.; Luo, J.; Zhang, A.; Zheng, Y.; Zhu, L.; Gu, Y.; Zhang, H. Berberine alleviates pulmonary hypertension through Trx1 and β-catenin signaling pathways in pulmonary artery smooth muscle cells. Exp. Cell Res. 2020, 390, 111910. [Google Scholar]
- Patel, K.; Singh, G.K.; Patel, D.K. A Review on Pharmacological and Analytical Aspects of Naringenin. Chin. J. Integr. Med. 2018, 24, 551–560. [Google Scholar] [CrossRef]
- Yao, L.H.; Jiang, Y.M.; Shi, J.; Tomás-Barberán, F.A.; Datta, N.; Singanusong, R.; Chen, S.S. Flavonoids in food and their health benefits. Plant Foods Hum. Nutr. 2004, 59, 113–122. [Google Scholar] [CrossRef]
- Garg, A.; Garg, S.; Zaneveld, L.J.D.; Singla, A.K. Chemistry and pharmacology of the Citrus bioflavonoid hesperidin. Phytother. Res. 2001, 15, 655–669. [Google Scholar] [CrossRef]
- Zhang, L.; Song, L.; Zhang, P.; Liu, T.; Zhou, L.; Yang, G.; Lin, R.; Zhang, J. Solubilities of naringin and naringenin in different solvents and dissociation constants of naringenin. J. Chem. Eng. Data 2015, 60, 932–940. [Google Scholar] [CrossRef]
- Orrego-Lagarón, N.; Martínez-Huélamo, M.; Vallverdú-Queralt, A.; Lamuela-Raventos, R.M.; Escribano-Ferrer, E. High gastrointestinal permeability and local metabolism of naringenin: Influence of antibiotic treatment on absorption and metabolism. Br. J. Nutr. 2015, 114, 169–180. [Google Scholar] [CrossRef] [Green Version]
- Day, A.J.; Dupont, M.S.; Ridley, S.; Rhodes, M.; Rhodes, M.J.C.; Morgan, M.R.A.; Williamson, G. Deglycosylation of flavonoid and isoflavonoid glycosides by human small intestine and liver β-glucosidase activity. FEBS Lett. 1998, 436, 71–75. [Google Scholar] [CrossRef] [Green Version]
- Justesen, U.; Knuthsen, P.; Leth, T. Quantitative analysis of flavonols, flavones, and flavanones in fruits, vegetables and beverages by high-performance liquid chromatography with photo-diode array and mass spectrometric detection. J. Chromatogr. A 1998, 799, 101–110. [Google Scholar] [CrossRef]
- Wang, N.; Li, D.; Lu, N.H.; Yi, L.; Huang, X.W.; Gao, Z.H. Peroxynitrite and hemoglobin-mediated nitrative/oxidative modification of human plasma protein: Effects of some flavonoids. J. Asian Nat. Prod. Res. 2010, 12, 257–264. [Google Scholar] [CrossRef]
- Pinho-Ribeiro, F.A.; Zarpelon, A.C.; Mizokami, S.S.; Borghi, S.M.; Bordignon, J.; Silva, R.L.; Cunha, T.M.; Alves-Filho, J.C.; Cunha, F.Q.; Casagrande, R.; et al. The citrus flavonone naringenin reduces lipopolysaccharide-induced inflammatory pain and leukocyte recruitment by inhibiting NF-κB activation. J. Nutr. Biochem. 2016, 33, 8–14. [Google Scholar] [CrossRef]
- Han, X.; Ren, D.; Fan, P.; Shen, T.; Lou, H. Protective effects of naringenin-7-O-glucoside on doxorubicin-induced apoptosis in H9C2 cells. Eur. J. Pharmacol. 2008, 581, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Scholz, E.P.; Zitron, E.; Kiesecker, C.; Lück, S.; Thomas, D.; Kathöfer, S.; Kreye, V.A.W.; Katus, H.A.; Kiehn, J.; Schoels, W.; et al. Inhibition of cardiac HERG channels by grapefruit flavonoid naringenin: Implications for the influence of dietary compounds on cardiac repolarisation. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2005, 371, 516–525. [Google Scholar] [CrossRef]
- Bugianesi, R.; Salucci, M.; Leonardi, C.; Ferracane, R.; Catasta, G.; Azzini, E.; Maiani, G. Effect of domestic cooking on human bioavailability of naringenin, chlorogenic acid, lycopene and β-carotene in cherry tomatoes. Eur. J. Nutr. 2004, 43, 360–366. [Google Scholar] [CrossRef]
- Manach, C.; Morand, C.; Gil-Izquierdo, A.; Bouteloup-Demange, C.; Rémésy, C. Bioavailability in humans of the flavanones hesperidin and narirutin after the ingestion of two dose of orange juice. Eur. J. Clin. Nutr. 2003, 57, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Lucas-Abellán, C.; Pérez-Abril, M.; Castillo, J.; Serrano, A.; Mercader, M.T.; Fortea, M.I.; Gabaldón, J.A.; Núñez-Delicado, E. Effect of temperature, pH, β- and HP-β-cds on the solubility and stability of flavanones: Naringenin and hesperetin. LWT 2019, 108, 233–239. [Google Scholar] [CrossRef]
- Maatouk, M.; Elgueder, D.; Mustapha, N.; Chaaban, H.; Bzéouich, I.M.; Loannou, I.; Kilani, S.; Ghoul, M.; Ghedira, K.; Chekir-Ghedira, L. Effect of heated naringenin on immunomodulatory properties and cellular antioxidant activity. Cell Stress Chaperones 2016, 21, 1101–1109. [Google Scholar] [CrossRef] [Green Version]
- Tundis, R.; Loizzo, M.R.; Menichini, F.; Bonesi, M.; Colica, C.; Menichini, F. In vitro cytotoxic activity of extracts and isolated constituents of Salvia leriifolia benth. against a panel of human cancer cell lines. Chem. Biodivers. 2011, 8, 1152–1162. [Google Scholar] [CrossRef]
- Qin, L.; Jin, L.; Lu, L.; Lu, X.; Zhang, C.; Zhang, F.; Liang, W. Naringenin reduces lung metastasis in a breast cancer resection model. Protein Cell 2011, 2, 507–516. [Google Scholar] [CrossRef]
- Ahmed, L.A.; Obaid, A.A.Z.; Zaki, H.F.; Agha, A.M. Naringenin adds to the protective effect of l-arginine in monocrotaline-induced pulmonary hypertension in rats: Favorable modulation of oxidative stress, inflammation and nitric oxide. Eur. J. Pharm. Sci. 2014, 62, 161–170. [Google Scholar] [CrossRef]
- Masamune, A.; Satoh, M.; Kikuta, K.; Suzuki, N.; Satoh, K.; Shimosegawa, T. Ellagic acid blocks activation of pancreatic stellate cells. Biochem. Pharmacol. 2005, 70, 869–878. [Google Scholar] [CrossRef]
- Kang, E.H.; Kown, T.Y.; Oh, G.T.; Park, W.F.; Park, S.I.; Park, S.K.; Lee, Y.I. The flavonoid ellagic acid from a medicinal herb inhibits host immune tolerance induced by the hepatitis B virus-e antigen. Antiviral Res. 2006, 72, 100–106. [Google Scholar] [CrossRef]
- Bala, I.; Bhardwaj, V.; Hariharan, S.; Kumar, M.N.V.R. Analytical methods for assay of ellagic acid and its solubility studies. J. Pharm. Biomed. Anal. 2006, 40, 206–210. [Google Scholar] [CrossRef] [PubMed]
- Marić, B.; Bodroža-Solarov, M.; Ilić, N.; Kojić, J.; Krulj, J. Influence of different extrusion temperatures on the stability of ellagic acid from raspberry seeds. Food Feed Res. 2018, 45, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Talcott, S.T.; Lee, J.H. Ellagic acid and flavonoid antioxidant content of muscadine wine and juice. J. Agric. Food Chem. 2002, 50, 3186–3192. [Google Scholar] [CrossRef] [PubMed]
- Whitley, A.C.; Stoner, G.D.; Darby, M.V.; Walle, T. Intestinal epithelial cell accumulation of the cancer preventive polyphenol ellagic acid—Extensive binding to protein and DNA. Biochem. Pharmacol. 2003, 66, 907–915. [Google Scholar] [CrossRef]
- Murugan, V.; Mukherjee, K.; Maiti, K.; Mukherjee, P.K. Enhanced oral bioavailability and antioxidant profile of ellagic acid by phospholipids. J. Agric. Food Chem. 2009, 57, 4559–4565. [Google Scholar] [CrossRef] [PubMed]
- Seeram, N.P.; Lee, R.; Heber, D. Bioavailability of ellagic acid in human plasma after consumption of ellagitannins from pomegranate (Punica granatum L.) juice. Clin. Chim. Acta 2004, 348, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Saxena, M.; Mir, A.H.; Sharma, M.; Malla, M.Y.; Qureshi, S.; Mir, M.I.; Chaturvedi, Y. Phytochemical Screening and In-vitro antioxidant activity isolated bioactive compounds from tridax procumbens linn. Pak. J. Biol. Sci. 2013, 16, 1971–1977. [Google Scholar]
- Sun, Y.Q.; Tao, X.; Men, X.M.; Xu, Z.W.; Wang, T. In vitro and in vivo antioxidant activities of three major polyphenolic compounds in pomegranate peel: Ellagic acid, punicalin, and punicalagin. J. Integr. Agric. 2017, 16, 1808–1818. [Google Scholar] [CrossRef] [Green Version]
- Kilic, I.; Yeşiloǧlu, Y.; Bayrak, Y. Spectroscopic studies on the antioxidant activity of ellagic acid. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2014, 130, 447–452. [Google Scholar] [CrossRef]
- Papoutsi, Z.; Kassi, E.; Tsiapara, A.; Fokialakis, N.; Chrousos, G.P.; Moutsatsou, P. Evaluation of estrogenic/antiestrogenic activity of ellagic acid via the estrogen receptor subtypes ERα and ERβ. J. Agric. Food Chem. 2005, 53, 7715–7720. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Chen, G.X.; Liang, M.Y.; Yao, J.P.; Wu, Z.K. Ellagic acid prevents monocrotaline-induced pulmonary artery hypertension via inhibiting NLRP3 inflammasome activation in rats. Int. J. Cardiol. 2015, 180, 134–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curis, E.; Nicolis, I.; Moinard, C.; Osowska, S.; Zerrouk, N.; Bénazeth, S.; Cynober, L. Almost all about citrulline in mammals. Amino Acids 2005, 29, 177–205. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.R.; Webber, C.L.; Fish, W.W.; Wehner, T.C.; King, S.; Perkins-Veazie, P. L-citrulline levels in watermelon cultigens tested in two environments. HortScience 2011, 46, 1572–1575. [Google Scholar] [CrossRef]
- Moinard, C.; Nicolis, I.; Neveux, N.; Darquy, S.; Bénazeth, S.; Cynober, L. Dose-ranging effects of citrulline administration on plasma amino acids and hormonal patterns in healthy subjects: The Citrudose pharmacokinetic study. Br. J. Nutr. 2008, 99, 855–862. [Google Scholar] [CrossRef]
- Vadgama, J.V.; Evered, D.F. Characteristics of L-citrulline transport across rat small intestine in vitro. Pediatr. Res. 1992, 32, 472–478. [Google Scholar] [CrossRef] [Green Version]
- Bahri, S.; Curis, E.; El Wafi, F.Z.; Aussel, C.; Chaumeil, J.C.; Cynober, L.; Zerrouk, N. Mechanisms and kinetics of citrulline uptake in a model of human intestinal epithelial cells. Clin. Nutr. 2008, 27, 872–880. [Google Scholar] [CrossRef]
- Hayashi, T.; Juliet, P.A.R.; Matsui-Hirai, H.; Miyasaki, A.; Fukatsu, A.; Funami, J.; Iguchi, A.; Ignarro, L.J. L-citrulline and L-arginine supplementation retards the progression of high-cholesterol-diet-induced atherosclerosis in rabbits. Proc. Natl. Acad. Sci. USA 2005, 102, 13681–13686. [Google Scholar] [CrossRef] [Green Version]
- El-Kirsh, A.A.A.; Abd El-Wahab, H.M.F.; Abd-Ellah Sayed, H.F. The effect of L-arginine or L-citrulline supplementation on biochemical parameters and the vascular aortic wall in high-fat and high-cholesterol-fed rats. Cell Biochem. Funct. 2011, 29, 414–428. [Google Scholar] [CrossRef]
- Nagy, I.Z.; Floyd, R.A. Hydroxyl free radical reactions with amino acids and proteins studied by electron spin resonance spectroscopy and spin-trapping. Biochim. Biophys. Acta (BBA)/Protein Struct. Mol. 1984, 790, 238–250. [Google Scholar]
- Morris, S.M. Arginine: Beyond protein. Am. J. Clin. Nutr. 2006, 83, 508–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Q.; Joshi, M.; DiPiazza, J.; Joshi, V. Functional Relevance of Citrulline in the Vegetative Tissues of Watermelon during Abiotic Stresses. Front. Plant Sci. 2020, 11, 512. [Google Scholar] [CrossRef] [PubMed]
- Allerton, T.D.; Proctor, D.N.; Stephens, J.M.; Dugas, T.R.; Spielmann, G.; Irving, B.A. L-citrulline supplementation: Impact on cardiometabolic health. Nutrients 2018, 10, 921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarazona-Díaz, M.P.; Viegas, J.; Moldao-Martins, M.; Aguayo, E. Bioactive compounds from flesh and by-product of fresh-cut watermelon cultivars. J. Sci. Food Agric. 2011, 91, 805–812. [Google Scholar] [CrossRef]
- Fike, C.D.; Dikalova, A.; Kaplowitz, M.R.; Cunningham, G.; Summar, M.; Aschner, J.L. Rescue treatment with L-citrulline inhibits hypoxia-induced pulmonary hypertension in newborn pigs. Am. J. Respir. Cell Mol. Biol. 2015, 53, 255–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lourdes Reyes-Escogido, M.; Gonzalez-Mondragon, E.G.; Vazquez-Tzompantzi, E. Chemical and pharmacological aspects of capsaicin. Molecules 2011, 16, 1253–1270. [Google Scholar] [CrossRef] [Green Version]
- Hwang, I.G.; Shin, Y.J.; Lee, S.; Lee, J.; Yoo, S.M. Effects of different cooking methods on the antioxidant properties of red pepper (Capsicum annuum L.). Prev. Nutr. Food Sci. 2012, 17, 286–292. [Google Scholar] [CrossRef] [Green Version]
- Kawada, T.; Suzuki, T.; Takahashi, M.; Iwai, K. Gastrointestinal absorption and metabolism of capsaicin and dihydrocapsaicin in rats. Toxicol. Appl. Pharmacol. 1984, 72, 449–456. [Google Scholar] [CrossRef]
- Rollyson, W.D.; Stover, C.A.; Brown, K.C.; Perry, H.E.; Stevenson, C.D.; McNees, C.A.; Ball, J.G.; Valentovic, M.A.; Dasgupta, P. Bioavailability of capsaicin and its implications for drug delivery. J. Control. Release 2014, 196, 96–105. [Google Scholar] [CrossRef] [Green Version]
- Reilly, C.A.; Ehlhardt, W.J.; Jackson, D.A.; Kulanthaivel, P.; Mutlib, A.E.; Espina, R.J.; Moody, D.E.; Crouch, D.J.; Yost, G.S. Metabolism of capsaicin by cytochrome P450 produces novel dehydrogenated metabolites and decreases cytotoxicity to lung and liver cells. Chem. Res. Toxicol. 2003, 16, 336–349. [Google Scholar] [CrossRef]
- Chanda, S.; Bashir, M.; Babbar, S.; Koganti, A.; Bley, K. In vitro hepatic and skin metabolism of capsaicin. Drug Metab. Dispos. 2008, 36, 670–675. [Google Scholar] [CrossRef] [PubMed]
- Sancho, R.; Lucena, C.; Macho, A.; Calzado, M.A.; Blanco-Molina, M.; Minassi, A.; Appendino, G.; Muñoz, E. Immunosuppressive Activity of Capsaicinoids: Capsiate Derived From Sweet Peppers Inhibits NF-kappaB Activation and Is a Potent Antiinflammatory Compound in Vivo. Eur. J. Immunol. 2002, 32, 1753–1763. [Google Scholar] [CrossRef]
- Materska, M.; Perucka, I. Antioxidant activity of the main phenolic compounds isolated from hot pepper fruit (Capsicum annuum L.). J. Agric. Food Chem. 2005, 53, 1750–1756. [Google Scholar] [CrossRef] [PubMed]
- Cortright, D.W.; Szallasi, A. Biochemical pharmacology of the vanilloid receptor TRPV1: An update. Eur. J. Biochem. 2004, 271, 1814–1819. [Google Scholar] [CrossRef] [PubMed]
- Zholos, A. TRP Channels in Respiratory Pathophysiology: The Role of Oxidative, Chemical Irritant and Temperature Stimuli. Curr. Neuropharmacol. 2015, 13, 279–291. [Google Scholar] [CrossRef]
- Xu, Y.; Gu, Q.; Qu, C. Capsaicin pretreatment reversed pulmonary arterial hypertension by alleviating inflammation via p38MAPK pathway. Exp. Lung Res. 2017, 43, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Hansen, P.E.; Wang, G.; Qiu, L.; Dong, J.; Yin, H.; Qian, Z.; Yang, M.; Miao, J. Pharmacological profile of xanthohumol, a prenylated flavonoid from hops (Humulus lupulus). Molecules 2015, 20, 754–779. [Google Scholar] [CrossRef] [PubMed]
- Kondo, K. Beer and health: Preventive effects of beer components on lifestyle-related diseases. BioFactors 2004, 22, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.H.; Fu, M.L.; Chen, M.M.; Liu, J.; Liu, X.J.; He, G.Q.; Pu, S.C. Preparative isolation and purification of xanthohumol from hops (Humulus lupulus L.) by high-speed counter-current chromatography. Food Chem. 2012, 132, 619–623. [Google Scholar] [CrossRef] [PubMed]
- Wunderlich, S.; Zürcher, A.; Back, W. Enrichment of xanthohumol in the brewing process. Mol. Nutr. Food Res. 2005, 49, 874–881. [Google Scholar] [CrossRef] [PubMed]
- Bolton, J.L.; Dunlap, T.L.; Hajirahimkhan, A.; Mbachu, O.; Chen, S.N.; Chadwick, L.; Nikolic, D.; van Breemen, R.B.; Pauli, G.F.; Dietz, B.M. The Multiple Biological Targets of Hops and Bioactive Compounds. Chem. Res. Toxicol. 2019, 32, 222–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanoli, P.; Zavatti, M. Pharmacognostic and pharmacological profile of Humulus lupulus L. J. Ethnopharmacol. 2008, 116, 383–396. [Google Scholar] [CrossRef]
- Negrão, R.; Costa, R.; Duarte, D.; Taveira Gomes, T.; Mendanha, M.; Moura, L.; Vasques, L.; Azevedo, I.; Soares, R. Angiogenesis and inflammation signaling are targets of beer polyphenols on vascular cells. J. Cell. Biochem. 2010, 111, 1270–1279. [Google Scholar] [CrossRef]
- Silva, A.F.; Faria-Costa, G.; Sousa-Nunes, F.; Santos, M.F.; Ferreira-Pinto, M.J.; Duarte, D.; Rodrigues, I.; Guimarães, J.T.; Leite-Moreira, A.; Moreira-Gonçalves, D. Anti-remodeling effects of xanthohumol-fortified beer in pulmonary arterial hypertension mediated by ERK and AKT inhibition. Nutrients 2019, 11, 583. [Google Scholar] [CrossRef] [Green Version]
- Naz, S.; Imran, M.; Rauf, A.; Orhan, I.E.; Shariati, M.A.; Iahtisham-Ul-Haq; IqraYasming; Shahbazh, M.; Qaisranih, T.B.; Shah, Z.A. Chrysin: Pharmacological and therapeutic properties. Life Sci. 2019, 235, 116797. [Google Scholar] [CrossRef]
- Babangida, S.; Ibrahim, S.; Muhammad, A.; Arthur, D.E.; Uzairu, A.; Garba, A. The role of molecular modelling strategies in validating the effects of chrysin on sodium arsenite-induced chromosomal and DNA damage. Hum. Exp. Toxicol. 2018, 37, 1037–1047. [Google Scholar] [CrossRef]
- Mani, R.; Natesan, V. Chrysin: Sources, beneficial pharmacological activities, and molecular mechanism of action. Phytochemistry 2018, 145, 187–196. [Google Scholar] [CrossRef]
- Zhang, L.; Zuo, Z.; Lin, G. Intestinal and hepatic glucuronidation of flavonoids. Mol. Pharm. 2007, 4, 833–845. [Google Scholar] [CrossRef]
- Mantawy, E.M.; El-Bakly, W.M.; Esmat, A.; Badr, A.M.; El-Demerdash, E. Chrysin alleviates acute doxorubicin cardiotoxicity in rats via suppression of oxidative stress, inflammation and apoptosis. Eur. J. Pharmacol. 2014, 728, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Siddiq, M.; Dolan, K.D. Characterization and heat inactivation kinetics of polyphenol oxidase from blueberry (Vaccinium corymbosum L.). Food Chem. 2017, 218, 216–220. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.N.; Meng, X.; Li, Y.; Li, S.; Liu, Q.; Tang, G.Y.; Li, H.B. Fruits for prevention and treatment of cardiovascular diseases. Nutrients 2017, 9, 598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, R.R.; Renfroe, M.H.; Brevard, P.B.; Lee, R.E.; Gloeckner, J.W. Cooking does not decrease hydrophilic antioxidant capacity of wild blueberries. Int. J. Food Sci. Nutr. 2009, 60 (Suppl. 2), 88–98. [Google Scholar] [CrossRef] [PubMed]
- Smeriglio, A.; Barreca, D.; Bellocco, E.; Trombetta, D. Chemistry, Pharmacology and Health Benefits of Anthocyanins. Phytother. Res. 2016, 30, 1265–1286. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.A.; Figueroa, A.; Navaei, N.; Wong, A.; Kalfon, R.; Ormsbee, L.T.; Feresin, R.G.; Elam, M.L.; Hooshmand, S.; Payton, M.E.; et al. Daily blueberry consumption improves blood pressure and arterial stiffness in postmenopausal women with pre- and stage 1-hypertension: A randomized, double-blind, placebo-controlled clinical trial. J. Acad. Nutr. Diet. 2015, 115, 369–377. [Google Scholar] [CrossRef]
- Cutler, B.R.; Petersen, C.; Anandh Babu, P.V. Mechanistic insights into the vascular effects of blueberries: Evidence from recent studies. Mol. Nutr. Food Res. 2017, 61, 1600271. [Google Scholar] [CrossRef]
- Türck, P.; Fraga, S.; Salvador, I.; Campos-Carraro, C.; Lacerda, D.; Bahr, A.; Ortiz, V.; Hickmann, A.; Koetz, M.; Belló-Klein, A.; et al. Blueberry extract decreases oxidative stress and improves functional parameters in lungs from rats with pulmonary arterial hypertension. Nutrition 2020, 70, 110579. [Google Scholar] [CrossRef]
- D’Andrea, G. Quercetin: A flavonol with multifaceted therapeutic applications? Fitoterapia 2015, 106, 256–271. [Google Scholar] [CrossRef]
- Andres, S.; Pevny, S.; Ziegenhagen, R.; Bakhiya, N.; Schäfer, B.; Hirsch-Ernst, K.I.; Lampen, A. Safety Aspects of the Use of Quercetin as a Dietary Supplement. Mol. Nutr. Food Res. 2018, 62, 1700447. [Google Scholar] [CrossRef]
- Ioku, K.; Aoyama, Y.; Tokuno, A.; Terao, J.; Nakatani, N.; Takei, Y. Various cooking methods and the flavonoid content in onion. J. Nutr. Sci. Vitaminol. 2001, 47, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Németh, K.; Takácsova, M.; Piskula, M.K. Effect of cooking on yellow onion. Pol. J. Food Nutr. Sci. 2003, 53, 170–174. [Google Scholar]
- Awad, M.A.; De Jager, A.; Van Der Plas, L.H.W.; Van Der Krol, A.R. Flavonoid and chlorogenic acid changes in skin of “Elstar” and “Jonagold” apples during development and ripening. Sci. Hortic. 2001, 90, 69–83. [Google Scholar] [CrossRef]
- Awad, M.A.; De Jager, A.; Van Westing, L.M. Flavonoid and chlorogenic acid levels in apple fruit: Characterisation of variation. Sci. Hortic. 2000, 83, 249–263. [Google Scholar] [CrossRef]
- Day, A.J.; Gee, J.M.; DuPont, M.S.; Johnson, I.T.; Williamson, G. Absorption of quercetin-3-glucoside and quercetin-4′-glucoside in the rat small intestine: The role of lactase phlorizin hydrolase and the sodium-dependent glucose transporter. Biochem. Pharmacol. 2003, 65, 1199–1206. [Google Scholar] [CrossRef]
- Ferry, D.R.; Smith, A.; Malkhandi, J.; Fyfe, D.W.; DeTakats, P.G.; Anderson, D.; Baker, J.; Kerr, D.J. Phase I clinical trial of the flavonoid quercetin: Pharmacokinetics and evidence for in vivo tyrosine kinase inhibition. Clin. Cancer Res. 1996, 2, 659–668. [Google Scholar]
- Duarte, J.; Pérez-Vizcaíno, F.; Zarzuelo, A.; Jiménez, J.; Tamargo, J. Vasodilator effects of quercetin in isolated rat vascular smooth muscle. Eur. J. Pharmacol. 1993, 239, 1–7. [Google Scholar] [CrossRef]
- Gryglewski, R.J.; Korbut, R.; Robak, J.; Świȩs, J. On the mechanism of antithrombotic action of flavonoids. Biochem. Pharmacol. 1987, 36, 317–322. [Google Scholar] [CrossRef]
- Perez-Vizcaino, F.; Duarte, J.; Jimenez, R.; Santos-Buelga, C.; Osuna, A. Antihypertensive effects of the flavonoid quercetin. In Pharmacological Reports; Polish Academy of Sciences: Warsaw, Poland, 2009; pp. 67–75. [Google Scholar]
- Kasikci, M.B.; Bagdatlioglu, N. Bioavailability of quercetin. Curr. Res. Nutr. Food Sci. 2016, 4, 146–151. [Google Scholar] [CrossRef]
- Morales-Cano, D.; Menendez, C.; Moreno, E.; Moral-Sanz, J.; Barreira, B.; Galindo, P.; Pandolfi, R.; Jimenez, R.; Moreno, L.; Cogolludo, A.; et al. The flavonoid quercetin reverses pulmonary hypertension in rats. PLoS ONE 2014, 9, e114492. [Google Scholar] [CrossRef]
- He, Y.; Cao, X.; Liu, X.; Li, X.; Xu, Y.; Liu, J.; Shi, J. Quercetin reverses experimental pulmonary arterial hypertension by modulating the TrkA pathway. Exp. Cell Res. 2015, 339, 122–134. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.; Eliot, K.; Heuertz, R.M.; Weiss, E. Whole Beetroot Consumption Acutely Improves Running Performance. J. Acad. Nutr. Diet. 2012, 112, 548–552. [Google Scholar] [CrossRef] [PubMed]
- Salloum, F.N.; Sturz, G.R.; Yin, C.; Rehman, S.; Hoke, N.N.; Kukreja, R.C.; Xi, L. Beetroot juice reduces infarct size and improves cardiac function following ischemia–reperfusion injury: Possible involvement of endogenous H2S. Exp. Biol. Med. 2015, 240, 669–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhaswant, M.; Brown, L.; McAinch, A.J.; Mathai, M.L. Beetroot and Sodium Nitrate Ameliorate Cardiometabolic Changes in Diet-Induced Obese Hypertensive Rats. Mol. Nutr. Food Res. 2017, 61, 478. [Google Scholar] [CrossRef]
- Tamme, T.; Reinik, M.; Püssa, T.; Roasto, M.; Meremäe, K.; Kiis, A. Dynamics of nitrate and nitrite content during storage of home-made and small-scale industrially produced raw vegetable juices and their dietary intake. Food Addit. Contam.-Part A Chem. Anal. Control Exp. Risk Assess. 2010, 27, 487–495. [Google Scholar] [CrossRef] [Green Version]
- Saikia, S.; Mahanta, C.L. Effect of steaming, boiling and microwave cooking on the total phenolics, flavonoids and antioxidant properties of different vegetables of Assam, India. Int. J. Food Nutr. Sci. 2013, 2, 47. [Google Scholar]
- Gilchrist, M.; Winyard, P.G.; Fulford, J.; Anning, C.; Shore, A.C.; Benjamin, N. Dietary nitrate supplementation improves reaction time in type 2 diabetes: Development and application of a novel nitrate-depleted beetroot juice placebo. Nitric Oxide Biol. Chem. 2014, 40, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Stanaway, L.; Rutherfurd-Markwick, K.; Page, R.; Ali, A. Performance and Health Benefits of Dietary Nitrate Supplementation in Older Adults: A Systematic Review. Nutrients 2017, 9, 1171. [Google Scholar] [CrossRef]
- Duncan, C.; Dougall, H.; Johnston, P.; Green, S.; Brogan, R.; Leifert, C.; Smith, L.; Golden, M.H.N.; Benjamin, N. Chemical generation of nitric oxide in the mouth from the enterosalivary circulation of dietary nitrate. Nat. Med. 1995, 1, 546–551. [Google Scholar] [CrossRef]
- Lundberg, J.O.; Govoni, M. Inorganic nitrate is a possible source for systemic generation of nitric oxide. Free Radic. Biol. Med. 2004, 37, 395–400. [Google Scholar] [CrossRef]
- Baliga, R.S.; Milsom, A.B.; Ghosh, S.M.; Trinder, S.L.; MacAllister, R.J.; Ahluwalia, A.; Hobbs, A.J. Dietary nitrate ameliorates pulmonary hypertension: Cytoprotective role for endothelial nitric oxide synthase and xanthine oxidoreductase. Circulation 2012, 125, 2922–2932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, Y.C.; Tabima, D.M.; Dube, J.J.; Hughan, K.S.; Vanderpool, R.R.; Goncharov, D.A.; Hobbs, A.J. SIRT3-AMP-Activated Protein Kinase Activation by Nitrite and Metformin Improves Hyperglycemia and Normalizes Pulmonary Hypertension Associated with Heart Failure with Preserved Ejection Fraction. Circulation 2016, 133, 717–731. [Google Scholar] [CrossRef] [PubMed]
- Pankey, E.A.; Badejo, A.M.; Casey, D.B.; Lasker, G.F.; Riehl, R.A.; Murthy, S.N.; Nossaman, B.D.; Kadowitz, P.J. Effect of chronic sodium nitrite therapy on monocrotaline-induced pulmonary hypertension. Nitric Oxide Biol. Chem. 2012, 27, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuckerbraun, B.S.; Shiva, S.; Ifedigbo, E.; Mathier, M.A.; Mollen, K.P.; Rao, J.; Bauer, P.M.; Choi, J.J.W.; Curtis, E.; Choi, A.M.K.; et al. Nitrite potently inhibits hypoxic and inflammatory pulmonary arterial hypertension and smooth muscle proliferation via xanthine oxidoreductase- dependent nitric oxide generation. Circulation 2010, 121, 98–109. [Google Scholar] [CrossRef]
- Tawa, M.; Furukawa, T.; Tongu, H.; Sugihara, M.; Taguwa, S.; Yamanaka, M.; Yano, Y.; Matsumori, H.; Kitada, R.; Sawano, T.; et al. Stimulation of nitric oxide-sensitive soluble guanylate cyclase in monocrotaline-induced pulmonary hypertensive rats. Life Sci. 2018, 203, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Tawa, M.; Yano, Y.; Yamanaka, M.; Sawano, T.; Iesaki, K.; Murata, Y.; Tanaka, R.; Nakagawa, K.; Ohkita, M.; Matsumura, Y. Effects of Beet Juice Supplementation on Monocrotaline-Induced Pulmonary Hypertension in Rats. Am. J. Hypertens. 2019, 32, 216–222. [Google Scholar] [CrossRef] [Green Version]
- Upadhyay, R.; Mohan Rao, L.J. An Outlook on Chlorogenic Acids-Occurrence, Chemistry, Technology, and Biological Activities. Crit. Rev. Food Sci. Nutr. 2013, 53, 968–984. [Google Scholar] [CrossRef]
- Liang, N.; Kitts, D.D. Role of chlorogenic acids in controlling oxidative and inflammatory stress conditions. Nutrients 2015, 8, 16. [Google Scholar] [CrossRef] [Green Version]
- Niggeweg, R.; Michael, A.J.; Martin, C. Engineering plants with increased levels of the antioxidant chlorogenic acid. Nat. Biotechnol. 2004, 22, 746–754. [Google Scholar] [CrossRef]
- Zhao, M.; Wang, H.; Yang, B.; Tao, H. Identification of cyclodextrin inclusion complex of chlorogenic acid and its antimicrobial activity. Food Chem. 2010, 120, 1138–1142. [Google Scholar] [CrossRef]
- Luo, H.J.; Wang, J.Z.; Chen, J.F.; Zou, K. Docking study on chlorogenic acid as a potential H5N1 influenza A virus neuraminidase inhibitor. Med. Chem. Res. 2011, 20, 554–557. [Google Scholar] [CrossRef]
- Li, G.; Wang, X.; Xu, Y.; Zhang, B.; Xia, X. Antimicrobial effect and mode of action of chlorogenic acid on Staphylococcus aureus. Eur. Food Res. Technol. 2014, 238, 589–596. [Google Scholar] [CrossRef]
- Vereshchagina, Y.V.; Bulgakov, V.P.; Grigorchuk, V.P.; Rybin, V.G.; Veremeichik, G.N.; Tchernoded, G.K.; Gorpenchenko, T.Y.; Koren, O.G.; Phan, N.H.; Minh, N.T.; et al. The rolC gene increases caffeoylquinic acid production in transformed artichoke cells. Appl. Microbiol. Biotechnol. 2014, 98, 7773–7780. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, S.; Majhi, S.; Saha, B.P.; Mukherjee, P.K. Chlorogenic acid-phospholipid complex improve protection against UVA induced oxidative stress. J. Photochem. Photobiol. B Biol. 2014, 130, 293–298. [Google Scholar] [CrossRef]
- Jin, S.; Chang, C.; Zhang, L.; Liu, Y.; Huang, X.; Chen, Z. Chlorogenic acid improves late diabetes through adiponectin receptor signaling pathways in db/db mice. PLoS ONE 2015, 10, e0120842. [Google Scholar] [CrossRef] [Green Version]
- Šilarová, P.; Boulekbache-Makhlouf, L.; Pellati, F.; Česlová, L. Monitoring of chlorogenic acid and antioxidant capacity of Solanum melongena L. (eggplant) under different heat and storage treatments. Antioxidants 2019, 8, 234. [Google Scholar] [CrossRef] [Green Version]
- Mills, C.E.; Oruna-Concha, M.J.; Mottram, D.S.; Gibson, G.R.; Spencer, J.P.E. The effect of processing on chlorogenic acid content of commercially available coffee. Food Chem. 2013, 141, 3335–3340. [Google Scholar] [CrossRef]
- Mubarak, A.; Bondonno, C.P.; Liu, A.H.; Considine, M.J.; Rich, L.; Mas, E.; Croft, K.D.; Hodgson, J.M. Acute effects of chlorogenic acid on nitric oxide status, endothelial function, and blood pressure in healthy volunteers: A randomized trial. J. Agric. Food Chem. 2012, 60, 9130–9136. [Google Scholar] [CrossRef]
- Gonthier, M.P.; Remesy, C.; Scalbert, A.; Cheynier, V.; Souquet, J.M.; Poutanen, K.; Aura, A.M. Microbial metabolism of caffeic acid and its esters chlorogenic and caftaric acids by human faecal microbiota in vitro. Biomed. Pharmacother. 2006, 60, 536–540. [Google Scholar] [CrossRef]
- Lafay, S.; Morand, C.; Manach, C.; Besson, C.; Scalbert, A. Absorption and metabolism of caffeic acid and chlorogenic acid in the small intestine of rats. Br. J. Nutr. 2006, 96, 39–46. [Google Scholar] [CrossRef]
- Kozuma, K.; Tsuchiya, S.; Kohori, J.; Hase, T.; Tokimitsu, I. Antihypertensive effect of green coffee bean extract on mildly hypertensive subjects. Hypertens. Res. 2005, 28, 711–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.Y.; Zhu, Y.F.; Zhang, M.; Chen, L.; Zhang, Z.; Du, Y.L.; Ren, G.-Q.; Tang, J.-M.; Zhong, M.-K.; Shi, X.-J. Chlorogenic acid inhibits hypoxia-induced pulmonary artery smooth muscle cells proliferation via c-Src and Shc/Grb2/ERK2 signaling pathway. Eur. J. Pharmacol. 2015, 751, 81–88. [Google Scholar] [CrossRef]
- Sayeski, P.P.; Ali, M.S. The critical role of c-Src and the Shc/Grb2/ERK2 signaling pathway in angiotensin II-dependent VSMC proliferation. Exp. Cell Res. 2003, 287, 339–349. [Google Scholar] [CrossRef]
- Cholewski, M.; Tomczykowa, M.; Tomczyk, M. A comprehensive review of chemistry, sources and bioavailability of omega-3 fatty acids. Nutrients 2018, 10, 1662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelhamid, A.S.; Martin, N.; Bridges, C.; Brainard, J.S.; Wang, X.; Brown, T.J.; Hanson, S.; Jimoh, O.F.; Ajabnoor, S.M.; Deane, K.H.O.; et al. Polyunsaturated fatty acids for the primary and secondary prevention of cardiovascular disease. Cochrane Database Syst. Rev. 2018, 2018, CD012345. [Google Scholar]
- Alfaia, C.M.M.; Alves, S.P.; Lopes, A.F.; Fernandes, M.J.E.; Costa, A.S.H.; Fontes, C.M.G.A.; Castro, M.L.F.; Bessa, R.J.B.; Prates, J.A.M. Effect of cooking methods on fatty acids, conjugated isomers of linoleic acid and nutritional quality of beef intramuscular fat. Meat Sci. 2010, 84, 769–777. [Google Scholar] [CrossRef]
- Domínguez, R.; Borrajo, P.; Lorenzo, J.M. The effect of cooking methods on nutritional value of foal meat. J. Food Compos. Anal. 2015, 43, 61–67. [Google Scholar] [CrossRef]
- Childs, C.E.; Romeu-Nadal, M.; Burdge, G.C.; Calder, P.C. Gender differences in the n-3 fatty acid content of tissues. Proc. Nutr. Soc. 2008, 67, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Decsi, T.; Kennedy, K. Sex-specific differences in essential fatty acid metabolism. Am. J. Clin. Nutr. 2011, 94 (Suppl. 6), 1914S–1919S. [Google Scholar] [CrossRef]
- Von Schacky, C. Omega-3 fatty acids and cardiovascular disease. Curr. Opin. Clin. Nutr. Metab. Care 2007, 10, 129–135. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Human Requirement for N-3 Polyunsaturated Fatty Acids. Poult. Sci. 2000, 79, 961–970. [Google Scholar] [CrossRef] [PubMed]
- Wideman, R.F.; Hamal, K.R.; Bayona, M.T.; Lorenzoni, A.G.; Cross, D.; Khajali, F.; Rhoads, D.D.; Erf, G.F.; Anthony, N.B. Plexiform Lesions in the Lungs of Domestic Fowl Selected for Susceptibility to Pulmonary Arterial Hypertension: Incidence and Histology. Anat. Rec. 2011, 294, 739–755. [Google Scholar] [CrossRef] [PubMed]
- Rostami, A.; Zamani Moghaddam, A.K.; Hassanpour, H.; Khajali, F. Pulmonary hypertension and right ventricular failure in broiler chickens reared at high altitude is affected by dietary source of n-6 and n-3 fatty acids. J. Anim. Physiol. Anim. Nutr. 2016, 100, 701–706. [Google Scholar] [CrossRef] [PubMed]
- Yamagata, K. Docosahexaenoic acid regulates vascular endothelial cell function and prevents cardiovascular disease. Lipids Health Dis. 2017, 16, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, L.H.; Wang, R.X.; Jiang, W.P.; Yang, X.J.; Song, J.P.; Li, X.R.; Tao, G. Effects of docosahexaenoic acid on large-conductance Ca2+-activated K+ channels and voltage-dependent K+ channels in rat coronary artery smooth muscle cells. Acta Pharmacol. Sin. 2009, 30, 314–320. [Google Scholar] [CrossRef] [Green Version]
- Nagaraj, C.; Tang, B.; Nagy, B.M.; Papp, R.; Jain, P.P.; Marsh, L.M.; Meredith, A.L.; Ghanim, B.; Klepetko, W.; Kwapiszewska, G.; et al. Docosahexaenoic acid causes rapid pulmonary arterial relaxation via KCa channel-mediated hyperpolarisation in pulmonary hypertension. Eur. Respir. J. 2016, 48, 1127–1136. [Google Scholar] [CrossRef] [Green Version]
- Conaway, C.; Yang, Y.; Chung, F. Isothiocyanates as Cancer Chemopreventive Agents: Their Biological Activities and Metabolism in Rodents and Humans. Curr. Drug Metab. 2005, 3, 233–255. [Google Scholar] [CrossRef]
- Wu, X.; Zhao, Y.; Haytowitz, D.B.; Chen, P.; Pehrsson, P.R. Effects of domestic cooking on flavonoids in broccoli and calculation of retention factors. Heliyon 2019, 5, e01310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, R.B.; Frisina, C.L.; Winkler, S.; Imsic, M.; Tomkins, R.B. Cooking method significantly effects glucosinolate content and sulforaphane production in broccoli florets. Food Chem. 2010, 123, 237–242. [Google Scholar] [CrossRef]
- Li, F.; Hullar, M.A.J.; Beresford, S.A.A.; Lampe, J.W. Variation of glucoraphanin metabolism in vivo and ex vivo by human gut bacteria. Br. J. Nutr. 2011, 106, 408–416. [Google Scholar] [CrossRef] [Green Version]
- Clarke, J.D.; Hsu, A.; Riedl, K.; Bella, D.; Schwartz, S.J.; Stevens, J.F.; Ho, E. Bioavailability and inter-conversion of sulforaphane and erucin in human subjects consuming broccoli sprouts or broccoli supplement in a cross-over study design. Pharmacol. Res. 2011, 64, 456–463. [Google Scholar] [CrossRef] [Green Version]
- Vermeulen, M.; Klöpping-Ketelaars, I.W.A.A.; Van Den Berg, R.; Vaes, W.H.J. Bioavailability and kinetics of sulforaphane in humans after consumption of cooked versus raw broccoli. J. Agric. Food Chem. 2008, 56, 10505–10509. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Dinkova-Kostova, A.T.; Wade, K.L.; Zhang, Y.; Shapiro, T.A.; Talalay, P. Quantitative determination of dithiocarbamates in human plasma, serum, erythrocytes and urine: Pharmacokinetics of broccoli sprout isothiocyanates in humans. Clin. Chim. Acta 2002, 316, 43–53. [Google Scholar] [CrossRef]
- Callaway, E.C.; Zhang, Y.; Chew, W.; Chow, H.H.S. Cellular accumulation of dietary anticarcinogenic isothiocyanates is followed by transporter-mediated export as dithiocarbamates. Cancer Lett. 2004, 204, 23–31. [Google Scholar] [CrossRef]
- Ji, Y.; Kuo, Y.; Morris, M.E. Pharmacokinetics of dietary phenethyl isothiocyanate in rats. Pharm. Res. 2005, 22, 1658–1666. [Google Scholar] [CrossRef]
- Barba, F.J.; Nikmaram, N.; Roohinejad, S.; Khelfa, A.; Zhu, Z.; Koubaa, M. Bioavailability of Glucosinolates and Their Breakdown Products: Impact of Processing. Front. Nutr. 2016, 3, 24. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Khor, T.O.; Yang, Q.; Huang, Y.; Wu, T.Y.; Saw, C.L.L.; Lin, W.; Androulakis, I.P.; Kong, A.-N.T. Pharmacokinetics and pharmacodynamics of phase II drug metabolizing/ antioxidant enzymes gene response by anticancer agent sulforaphane in rat lymphocytes. Mol. Pharm. 2012, 9, 2819–2827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, S.M.U.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef]
- Bai, Y.; Cui, W.; Xin, Y.; Miao, X.; Barati, M.T.; Zhang, C.; Chen, Q.; Tan, Y.; Cui, T.; Zheng, Y.; et al. Prevention by sulforaphane of diabetic cardiomyopathy is associated with up-regulation of Nrf2 expression and transcription activation. J. Mol. Cell. Cardiol. 2013, 57, 82–95. [Google Scholar] [CrossRef]
- Kyung, S.Y.; Kim, D.Y.; Yoon, J.Y.; Son, E.S.; Kim, Y.J.; Park, J.W.; Jeong, S.H. Sulforaphane attenuates pulmonary fibrosis by inhibiting the epithelial-mesenchymal transition. BMC Pharmacol. Toxicol. 2018, 19, 13. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Wang, S.; Zhou, S.; Yan, X.; Wang, Y.; Chen, J.; Mellen, N.; Kong, M.; Gu, J.; Tan, Y.; et al. Sulforaphane prevents the development of cardiomyopathy in type 2 diabetic mice probably by reversing oxidative stress-induced inhibition of LKB1/AMPK pathway. J. Mol. Cell. Cardiol. 2014, 77, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Zhang, G.; Huang, E.C.; Huang, J.; Cai, J.; Cai, L.; Wang, S.; Keller, B.B. Sulforaphane prevents right ventricular injury and reduces pulmonary vascular remodeling in pulmonary arterial hypertension. Am. J. Physiol. Heart Circ. Physiol. 2020, 318, H853–H866. [Google Scholar] [CrossRef]
- Slusarenko, A.J.; Patel, A.; Portz, D. Control of plant diseases by natural products: Allicin from garlic as a case study. In European Journal of Plant Pathology; Springer: Dordrecht, The Netherlands, 2008; pp. 313–322. [Google Scholar]
- Song, K.; Milner, J.A. Influence of Heating on the Anticancer Properties of Garlic. J. Nutr. 2001, 131, 1054S–1057S. [Google Scholar] [CrossRef] [PubMed]
- Varga-Visi, É.; Jócsák, I.; Ferenc, B.; Végvári, G. Effect of crushing and heating on the formation of volatile organosulfur compounds in garlic. CyTA J. Food 2019, 17, 796–803. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.; Jiang, X.; Wang, H.; Zhao, Z.; Wang, W. Drug Metabolism and Pharmacokinetics of Organosulfur Compounds from Garlic. J. Drug Metab. Toxicol. 2013, 4, 10. [Google Scholar]
- Elkayam, A.; Peleg, E.; Grossman, E.; Shabtay, Z.; Sharabi, Y. Effects of allicin on cardiovascular risk factors in spontaneously hypertensive rats. Isr. Med. Assoc. J. 2013, 15, 170–173. [Google Scholar]
- Liu, C.; Cao, F.; Tang, Q.Z.; Yan, L.; Dong, Y.G.; Zhu, L.H.; Wang, L.; Bian, Z.-Y.; Li, H. Allicin protects against cardiac hypertrophy and fibrosis via attenuating reactive oxygen species-dependent signaling pathways. J. Nutr. Biochem. 2010, 21, 1238–1250. [Google Scholar] [CrossRef]
- García-Trejo, E.M.A.; Arellano-Buendía, A.S.; Argüello-García, R.; Loredo-Mendoza, M.L.; García-Arroyo, F.E.; Arellano-Mendoza, M.G.; Castillo-Hernández, M.C.; Guevara-Balcázar, G.; Tapia, E.; Sánchez-Lozada, L.G.; et al. Effects of Allicin on Hypertension and Cardiac Function in Chronic Kidney Disease. Oxid. Med. Cell. Longev. 2016, 2016. [Google Scholar] [CrossRef] [Green Version]
- Szmitko, P.E.; Wang, C.H.; Weisel, R.D.; De Almeida, J.R.; Anderson, T.J.; Verma, S. New Markers of Inflammation and Endothelial Cell Activation Part I. Circulation 2003, 108, 1917–1923. [Google Scholar] [CrossRef]
- Zhang, C. The role of inflammatory cytokines in endothelial dysfunction. Basic Res. Cardiol. 2008, 103, 398–406. [Google Scholar] [CrossRef] [Green Version]
- Park, B.M.; Chun, H.; Chae, S.W.; Kim, S.H. Fermented garlic extract ameliorates monocrotaline-induced pulmonary hypertension in rats. J. Funct. Foods 2017, 30, 247–253. [Google Scholar] [CrossRef]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Gloria, J.L.; Osorio-Alonso, H.; Arellano-Buendía, A.S.; Carbó, R.; Hernández-Díazcouder, A.; Guzmán-Martín, C.A.; Rubio-Gayosso, I.; Sánchez-Muñoz, F. Nutraceuticals in the Treatment of Pulmonary Arterial Hypertension. Int. J. Mol. Sci. 2020, 21, 4827. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21144827
Sánchez-Gloria JL, Osorio-Alonso H, Arellano-Buendía AS, Carbó R, Hernández-Díazcouder A, Guzmán-Martín CA, Rubio-Gayosso I, Sánchez-Muñoz F. Nutraceuticals in the Treatment of Pulmonary Arterial Hypertension. International Journal of Molecular Sciences. 2020; 21(14):4827. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21144827
Chicago/Turabian StyleSánchez-Gloria, José L., Horacio Osorio-Alonso, Abraham S. Arellano-Buendía, Roxana Carbó, Adrián Hernández-Díazcouder, Carlos A. Guzmán-Martín, Ivan Rubio-Gayosso, and Fausto Sánchez-Muñoz. 2020. "Nutraceuticals in the Treatment of Pulmonary Arterial Hypertension" International Journal of Molecular Sciences 21, no. 14: 4827. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21144827