Neuroprotective Effect of Bean Phosphatidylserine on TMT-Induced Memory Deficits in a Rat Model


Abstract
:1. Introduction
2. Results
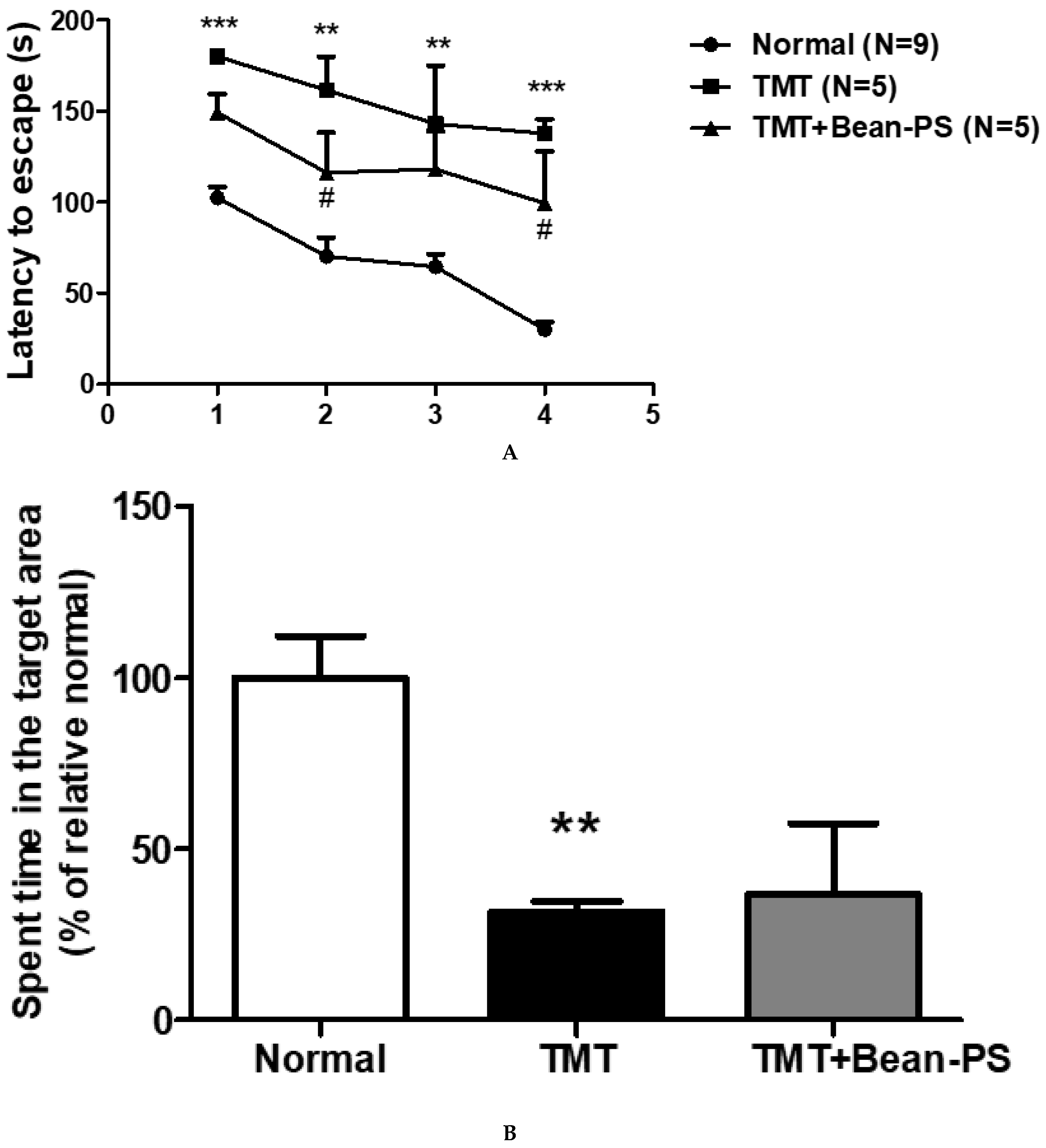
2.1. Morris Water Maze Test
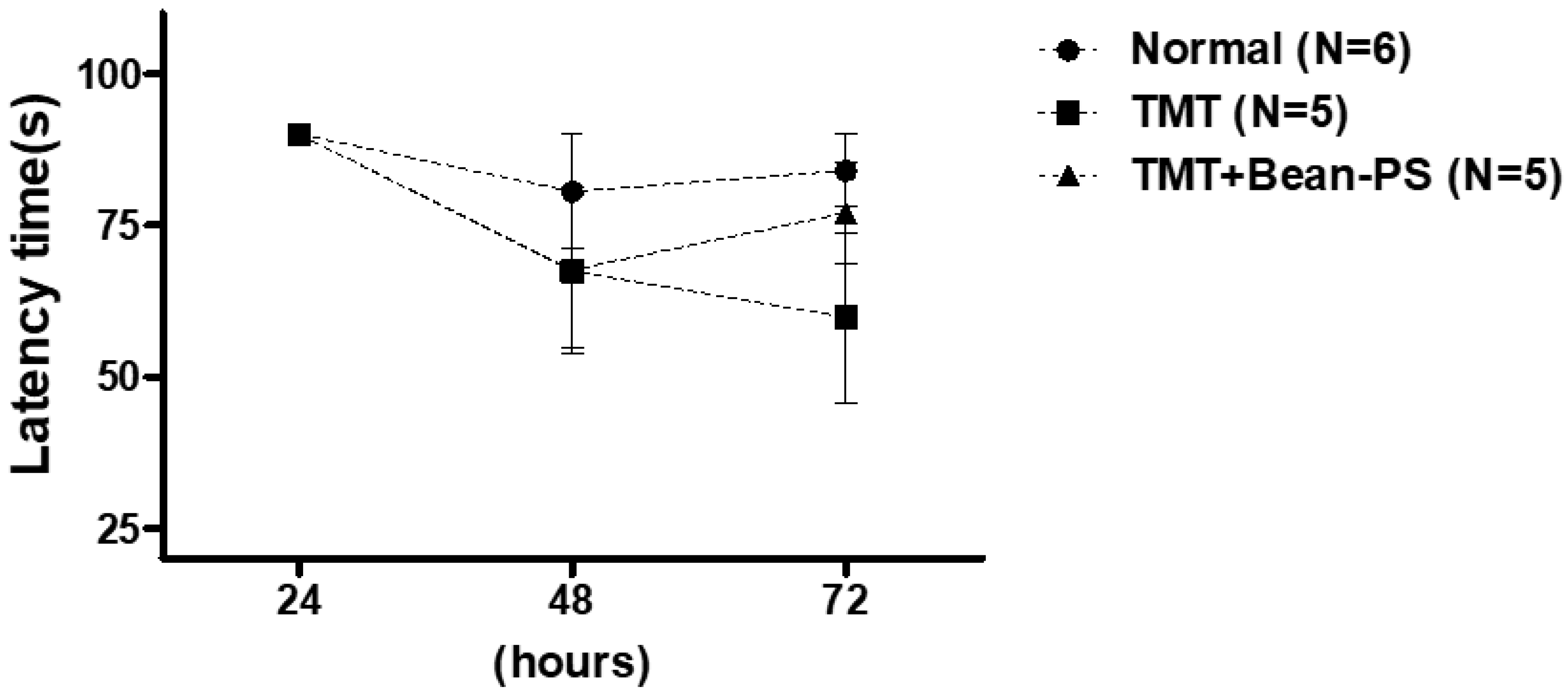
2.2. Passive Avoidance Test
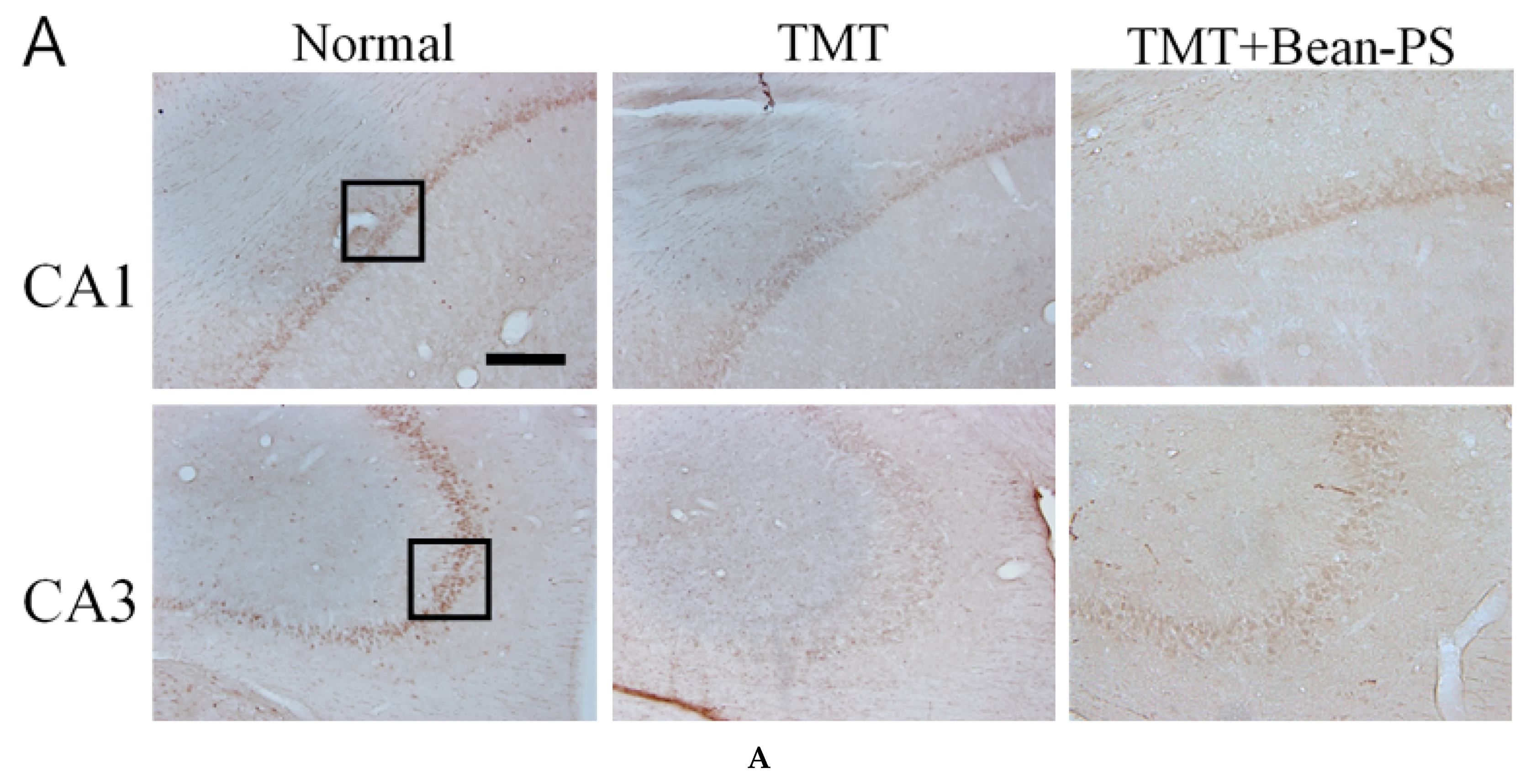
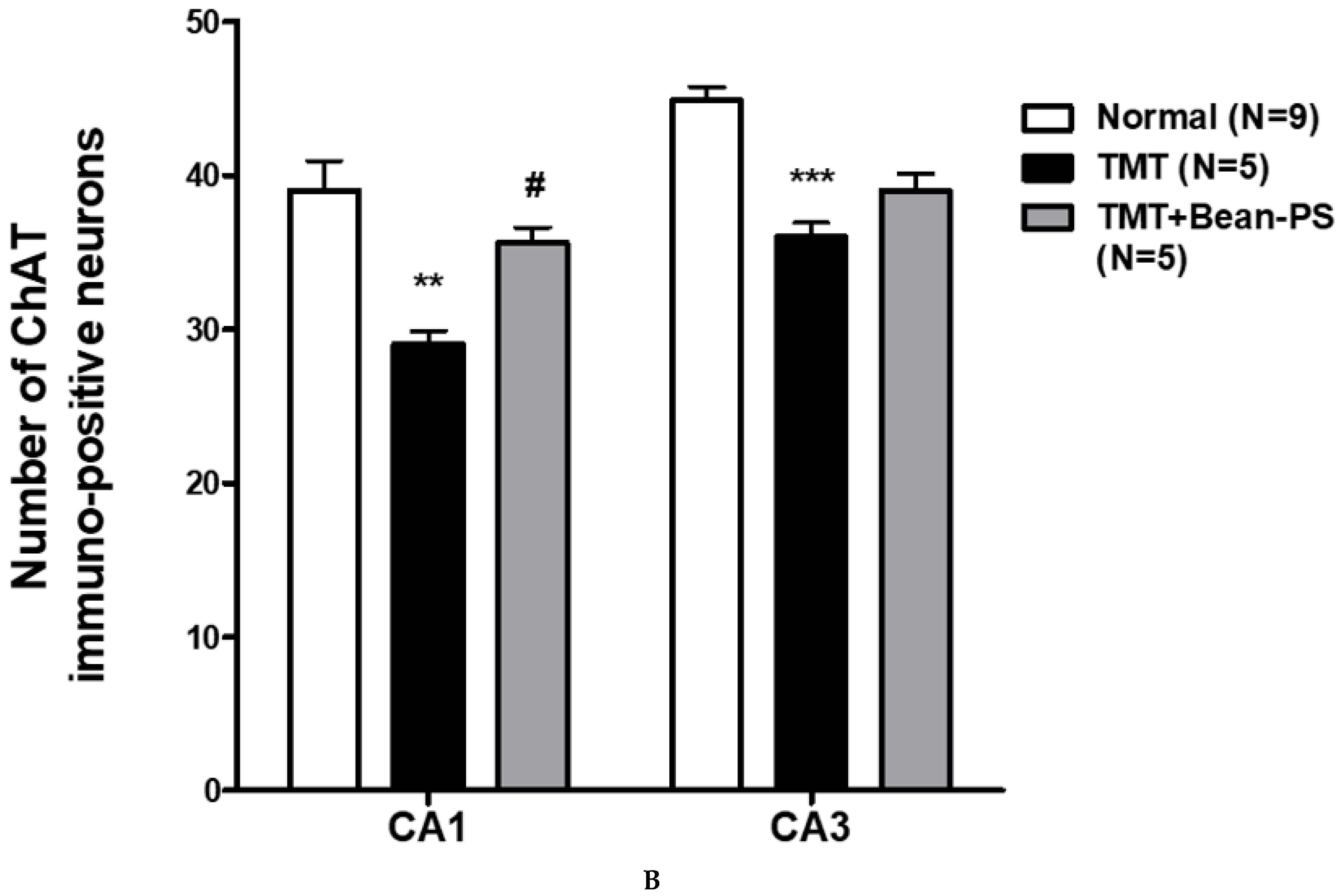
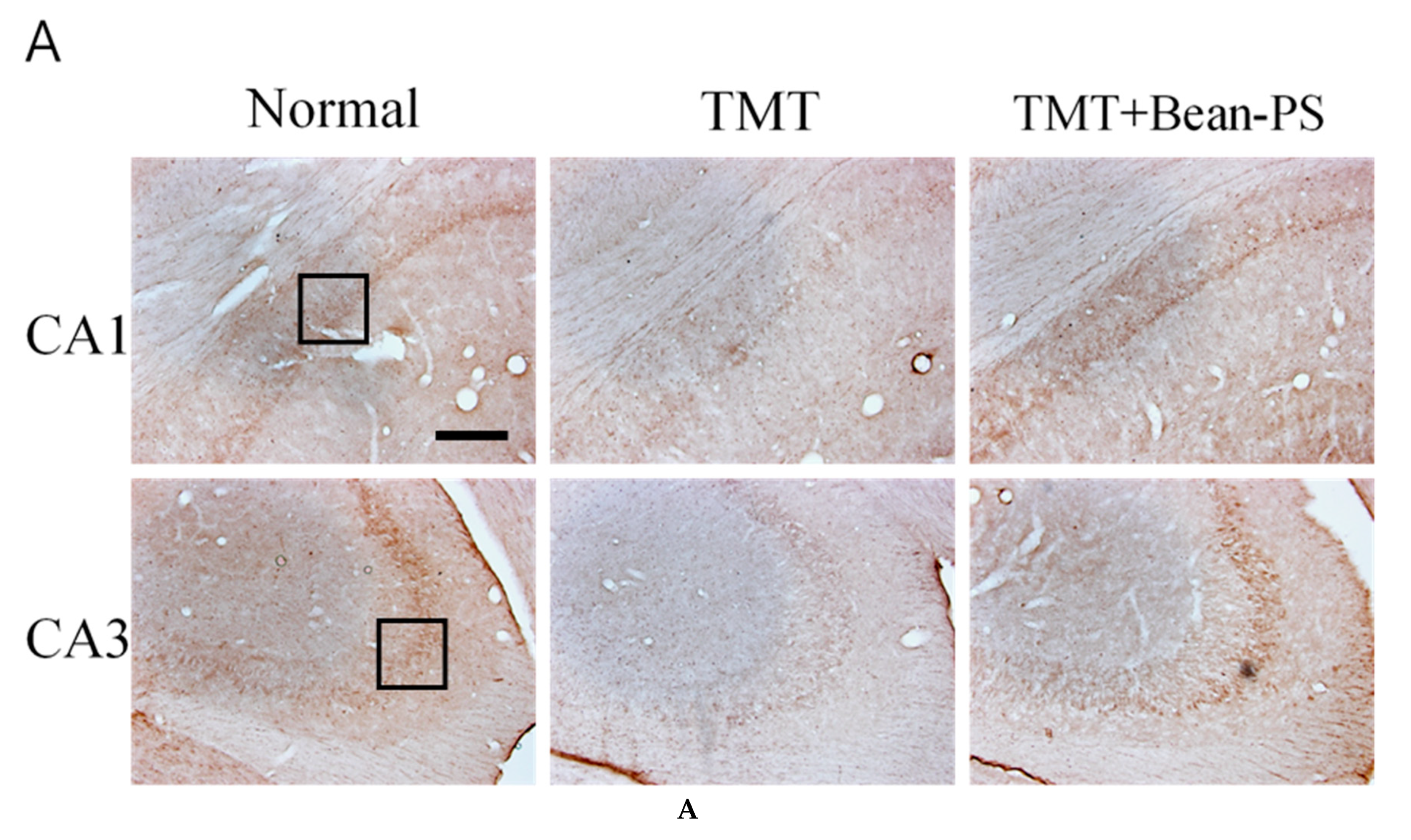
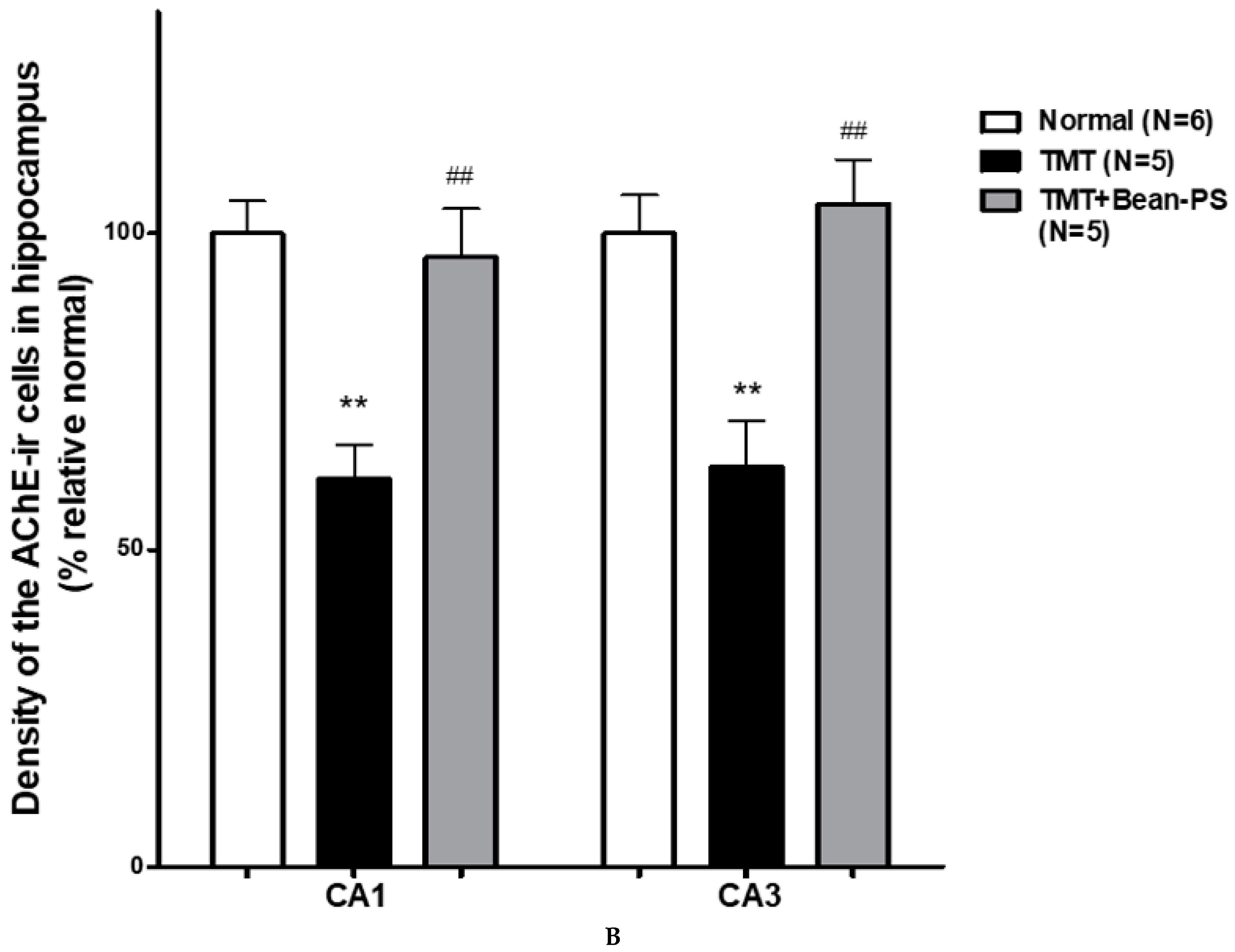
2.3. ChAT and AchE Immunoreactivity
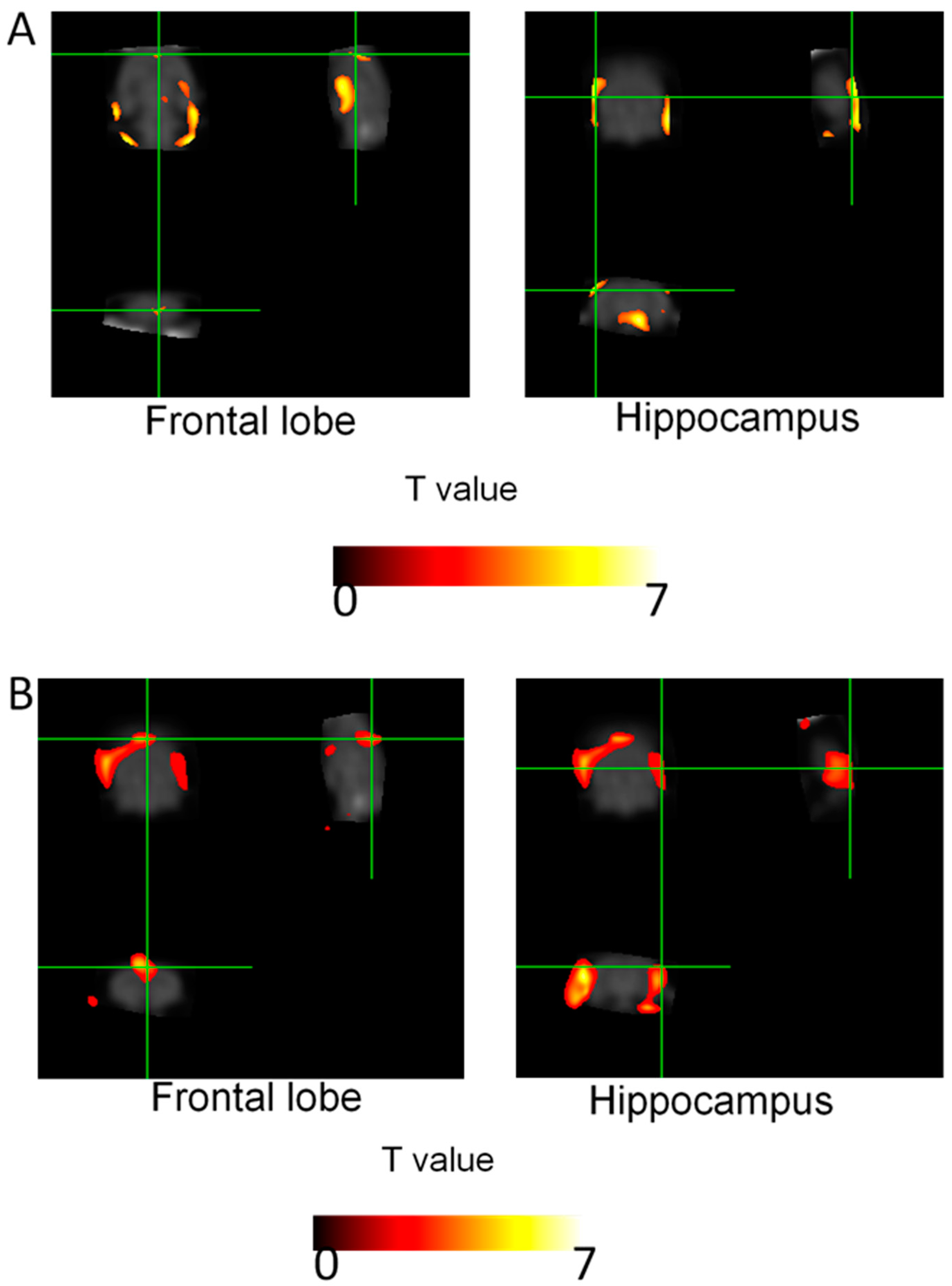
2.4. Brain Glucose Metabolism
3. Discussion
4. Material and Methods
4.1. Animals and Experimental Design
4.2. Drug Treatment
4.3. Morris Water Maze (MWM)
4.4. Passive Avoidance Task (PAT)
4.5. Immunohistochemical Staining
4.6. Image Processing and Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Jung, E.-Y.; Lee, M.-S.; Ahn, C.J.; Cho, S.-H.; Bae, H.; Shim, I. The Neuroprotective Effect of Gugijihwang-Tang on Trimethyltin-Induced Memory Dysfunction in the Rat. Evid. Based Complement. Altern. Med. 2013, 2013, 1–6. [Google Scholar] [CrossRef]
- Kang, J.Y.; Park, S.K.; Guo, T.J.; Ha, J.S.; Lee, D.S.; Kim, J.M.; Lee, U.; Kim, D.; Heo, H.J. Reversal of Trimethyltin-Induced Learning and Memory Deficits by 3,5-Dicaffeoylquinic Acid. Oxidative Med. Cell. Longev. 2016, 2016, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Huang, C.; Zheng, L.; Simonich, M.; Bai, C.; Tanguay, R.L.; Dong, Q. Trimethyltin chloride (TMT) neurobehavioral toxicity in embryonic zebrafish. Neurotoxicol. Teratol. 2011, 33, 721–726. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Jiang, C.; Li, R.; Zhao, S.; Li, W.; Tang, X. The neuroprotective effect of lithium chloride on cognitive impairment through glycogen synthase kinase-3beta inhibition in intracerebral hemorrhage rats. Eur. J. Pharmacol. 2018, 840, 50–59. [Google Scholar] [CrossRef]
- Corvino, V.; Marchese, E.; Giannetti, S.; Lattanzi, W.; Bonvissuto, D.; Biamonte, F.; Mongiovì, A.M.; Michetti, F.; Geloso, M.C. The neuroprotective and neurogenic effects of neuropeptide Y administration in an animal model of hippocampal neurodegeneration and temporal lobe epilepsy induced by trimethyltin. J. Neurochem. 2012, 122, 415–426. [Google Scholar] [CrossRef]
- Dyer, R.S.; Boyes, W.K. Trimethyltin reduces recurrent inhibition in rats. Neurobehav. Toxicol. Teratol. 1984, 6, 367–371. [Google Scholar]
- Naalsund, L.U.; Allen, C.N.; Fonnum, F. Changes in neurobiological parameters in the hippocampus after exposure to trimethyltin. NeuroToxicology 1985, 6, 145–158. [Google Scholar]
- Valdes, J.J.; Mactutus, C.F.; Santos-Anderson, R.M.; Dawson, R.; Annau, Z. Selective neurochemical and histological lesions in rat hippocampus following chronic trimethyltin exposure. Neurobehav. Toxicol. Teratol. 1983, 5, 357–361. [Google Scholar]
- Chang, L.W.; Dyer, R.S. Early effects of trimethyltin on the dentate gyrus basket cells: A morphological study. J. Toxicol. Environ. Heal. Part A 1985, 16, 641–653. [Google Scholar] [CrossRef]
- Oderfeld-Nowak, B.; Zaremba, M. GM1 ganglioside potentiates trimethyltin-induced expression of interleukin-1 beta and the nerve growth factor in reactive astrocytes in the rat hippocampus: An immunocytochemical study. Neurochem. Res. 1998, 23, 443–453. [Google Scholar] [CrossRef]
- Koczyk, D.; Oderfeld-Nowak, B. Long-term microglial and astroglial activation in the hippocampus of trimethyltin-intoxicated rat: Stimulation of NGF and TrkA immunoreactivities in astroglia but not in microglia. Int. J. Dev. Neurosci. 2000, 18, 591–606. [Google Scholar] [CrossRef]
- Halladay, A.; Wilson, D.; Wagner, G.; Reuhl, K. Trimethyltin-induced alterations in behavior are linked to changes in PSA-NCAM expression. NeuroToxicol. 2006, 27, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.R.; Choi, S.J.; Kwon, Y.K.; Kim, J.K.; Kim, Y.-J.; Park, G.G.; Shin, N.-H. Cinnamomum loureirii Extract Inhibits Acetylcholinesterase Activity and Ameliorates Trimethyltin-Induced Cognitive Dysfunction in Mice. Boil. Pharm. Bull. 2016, 39, 1130–1136. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, I.F.; Lima, L.C.F.; Graceli, J.B.; Rodrigues, L.C.D.M. Organotins in Neuronal Damage, Brain Function, and Behavior: A Short Review. Front. Endocrinol. 2018, 8, 366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodruff, M.L.; Baisden, R.H.; Cannon, R.L.; Kalbfleisch, J.; Freeman, J.N. Effects of trimethyltin on acquisition and reversal of a light-dark discrimination by rats. Physiol. Behav. 1994, 55, 1055–1061. [Google Scholar] [CrossRef]
- Kobayashi, H.; Saito, F.; Yuyama, A. Effects of organotins on the cholinergic system in the chicken brain in vitro. Toxicol. Vitr. 1992, 6, 337–343. [Google Scholar] [CrossRef]
- Francis, P.T.; Palmer, A.M.; Snape, M.; Wilcock, G.K. The cholinergic hypothesis of Alzheimer’s disease: A review of progress. J. Neurol. Neurosurg. Psychiatry 1999, 66, 137–147. [Google Scholar] [CrossRef]
- Van Marum, R. Current and future therapy in Alzheimer’s disease. Fundam. Clin. Pharmacol. 2008, 22, 265–274. [Google Scholar] [CrossRef]
- Lee, B.; Sur, B.-J.; Han, J.-J.; Shim, I.; Her, S.; Lee, H.-J.; Hahm, D.-H. Krill phosphatidylserine improves learning and memory in Morris water maze in aged rats. Prog. Neuro-Psychopharm. Boil. Psychiatry 2010, 34, 1085–1093. [Google Scholar] [CrossRef]
- Amaducci, L.; Crook, T.H.; Lippi, A.; Bracco, L.; Baldereschi, M.; Latorraca, S.; Piersanti, P.; Tesco, G.; Sorbi, S. Use of Phosphatidylserine in Alzheimer’s Disease. Ann. N. Y. Acad. Sci. 1991, 640, 245–249. [Google Scholar] [CrossRef]
- Friedland, R.P.; Petersen, R.B.; Rubenstein, R. Bovine Spongiform Encephalopathy and Aquaculture. J. Alzheimer’s Dis. 2008, 17, 277–279. [Google Scholar] [CrossRef]
- Kato-Kataoka, A.; Sakai, M.; Ebina, R.; Nonaka, C.; Asano, T.; Miyamori, T. Soybean-Derived Phosphatidylserine Improves Memory Function of the Elderly Japanese Subjects with Memory Complaints. J. Clin. Biochem. Nutr. 2010, 47, 246–255. [Google Scholar] [CrossRef] [Green Version]
- Jorissen, B.; Brouns, F.; Van Boxtel, M.; Ponds, R.; Verhey, F.; Jolles, J.; Riedel, W. The influence of soy-derived phosphatidylserine on cognition in age-associated memory impairment. Nutr. Neurosci. 2001, 4, 121–134. [Google Scholar] [CrossRef] [Green Version]
- Wimo, A.; Winblad, B. Economical Aspects of Dementia. Hendbook Clin. Neurol. 2008, 89, 137–146. [Google Scholar] [CrossRef]
- Fortemps, E.; Amand, G.; Bomboir, A.; Lauwerys, R.; Laterre, E.C. Trimethyltin poisoning report of two cases. Int. Arch. Occup. Environ. Health 1978, 41, 1–6. [Google Scholar] [CrossRef]
- Dyer, R.S. Physiological methods for assessment of Trimethyltin exposure. Neurobehav. Toxicol. Teratol. 1982, 4, 659–664. [Google Scholar]
- Brown, A.W.; Aldridge, W.N.; Street, B.W.; Verschoyle, R.D. The behavioral and neuropathologic sequelae of intoxication by trimethyltin compounds in the rat. Am. J. Pathol. 1979, 97, 59–82. [Google Scholar]
- Chang, L.W.; Tiemeyer, T.M.; Wenger, G.R.; McMillan, D.E. Neuropathology of trimethyltin intoxication. III. Changes in the brain stem neurons. Environ. Res. 1983, 30, 399–411. [Google Scholar] [CrossRef]
- Balaban, C.; Callaghan, J.; Billingsle, M. Trimethyltin-induced neuronal damage in the rat brain: Comparative studies using silver degeneration stains, immunocytochemistry and immunoassay for neuronotypic and gliotypic proteins. Neuroscience 1988, 26, 337–361. [Google Scholar] [CrossRef]
- Chang, L.W.; Dyer, R.S. A time-course study of trimethyltin induced neuropathology in rats. Neurobehav. Toxicol. Teratol. 1983, 5, 443–459. [Google Scholar]
- Chang, L.W.; Dyer, R.S. Trimethyltin induced pathology in sensory neurons. Neurobehav. Toxicol. Teratol. 1983, 5, 673–696. [Google Scholar]
- Chang, L.W.; Wenger, G.R.; McMillan, D.E.; Dyer, R.S. Species and strain comparison of acute neurotoxic effects of trimethyltin in mice and rats. Neurobehav. Toxicol. Teratol. 1983, 5, 337–350. [Google Scholar]
- Chang, L.W.; Dyer, R.S. Septotemporal gradients of trimethyltin-induced hippocampal lesions. Neurobehav. Toxicol. Teratol. 1985, 7, 43–49. [Google Scholar] [PubMed]
- Noraberg, J.; Gramsbergen, J.B.P.; Fonnum, F.; Zimmer, J. Trimethyltin (TMT) neurotoxicity in organotypic rat hippocampal slice cultures. Brain Res. 1998, 783, 305–315. [Google Scholar] [CrossRef]
- Swartzwelder, H.S.; Hepler, J.; Holahan, W.; King, S.E.; Leverenz, H.A.; A Miller, P.; Myers, R.D. Imparied maze performance in the rat caused by trimethyltin treatment: Problem-solving deficits and perseveration. Neurobehav. Toxicol. Teratol. 1982, 4, 169–176. [Google Scholar]
- Andersson, H.; Luthman, J.; Lindqvist, E.; Olson, L. Time-course of trimethyltin effects on the monoaminergic systems of the rat brain. NeuroToxicology 1995, 16, 201–210. [Google Scholar] [PubMed]
- Koczyk, D.; Skup, M.; Zaremba, M.; Oderfeld-Nowak, B. Trimethyltin-induced plastic neuronal changes in rat hippocampus are accompanied by astrocytic trophic activity. Acta Neurobiol. Exp. 1996, 56, 237–241. [Google Scholar]
- Zhao, T.F.; Xu, C.X.; Li, Z.W.; Xie, F.; Zhao, Y.T.; Wang, S.Q.; Luo, C.H.; Lu, R.S.; Ni, G.L.; Ku, Z.Q.; et al. (Effect of Tremella fuciformis Berk on acute radiation sickness in dogs (author’s transl)). Zhongguo Yi Xue Ke Xue Yuan Xue Bao 1982, 4, 20–23. [Google Scholar]
- Walsh, T.J.; Gallagher, M.; Bostock, E.; Dyer, R.S. Trimethyltin impairs retention of a passive avoidance task. Neurobehav. Toxicol. Teratol. 1982, 4, 163–167. [Google Scholar]
- Squire, L.R.; Davis, H.P. The Pharmacology of Memory: A Neurobiological Perspective. Annu. Rev. Pharmacol. Toxicol. 1981, 21, 323–356. [Google Scholar] [CrossRef]
- Lanier, L.P.; Isaacson, R.L. Activity changes related to the location of lesions in the hippocampus. Behav. Boil. 1975, 13, 59–69. [Google Scholar] [CrossRef]
- Agrawal, A.K.; Roy, A.; Seth, K.; Raghubir, R.; Seth, P. Restorative potential of cholinergic rich transplants in cholchicine induced lesioned rats: A comparative study of single and multiple micro-transplantation approach. Int. J. Dev. Neurosci. 2003, 21, 191–198. [Google Scholar] [CrossRef]
- Muir, J.L. Acetylcholine, Aging, and Alzheimer’s Disease. Pharmacol. Biochem. Behav. 1997, 56, 687–696. [Google Scholar] [CrossRef]
- Shinjo, H.; Ueki, A.; Miwa, C.; Morita, Y. Effect of entorhinal cortex lesion on hippocampal cholinergic system in rat in operant learning task as studied by in vivo brain microdialysis. J. Neurol. Sci. 1998, 157, 13–18. [Google Scholar] [CrossRef]
- Giacobini, E. Long-term stabilizing effect of cholinesterase inhibitors in the therapy of Alzheimer’ disease. J. Neural. Transm. Suppl. 2002, 62, 181–187. [Google Scholar] [CrossRef]
- Lorenzini, C.A.; Baldi, E.; Bucherelli, C.; Sacchetti, B.; Tassoni, G. Role of dorsal hippocampus in acquisition, consolidation and retrieval of rat’s passive avoidance response: A tetrodotoxin functional inactivation study. Brain Res. 1996, 730, 32–39. [Google Scholar] [CrossRef]
- Ajanal, M.; Gundkalle, M.B.; Nayak, S.U. Estimation of total alkaloid in Chitrakadivati by UV-Spectrophotometer. Anc. Sci. Life 2012, 31, 198–201. [Google Scholar] [CrossRef]
- Bao, Q.; Newport, D.; Chen, M.; Stout, D.B.; Chatziioannou, A.F. Performance evaluation of the inveon dedicated PET preclinical tomograph based on the NEMA NU-4 standards. J. Nucl. Med. 2009, 50, 401–408. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A | ||
| Brain Area | Coordinates (x, y, z) | Z Value |
| Frontal lobe | (8, 1, 10) | 1.95 |
| Hippocampus (Right) | (9, −1, 11) | 1.69 |
| Hippocampus (Left) | (−9, −1, 10) | 1.66 |
| B | ||
| Brain Area | Coordinates (x, y, z) | Z Value |
| Frontal lobe | (2, 4, 8) | 3.24 |
| Hippocampus (Right) | (7, −3, 3) | 4.57 |
| Hippocampus (Left) | (−5, −5, 1) | 3.3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, M.; Han, B.H.; Kim, J.S.; Kim, K.; Shim, I. Neuroprotective Effect of Bean Phosphatidylserine on TMT-Induced Memory Deficits in a Rat Model. Int. J. Mol. Sci. 2020, 21, 4901. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21144901
Ye M, Han BH, Kim JS, Kim K, Shim I. Neuroprotective Effect of Bean Phosphatidylserine on TMT-Induced Memory Deficits in a Rat Model. International Journal of Molecular Sciences. 2020; 21(14):4901. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21144901
Chicago/Turabian StyleYe, Minsook, Bong Hee Han, Jin Su Kim, Kyungsoo Kim, and Insop Shim. 2020. "Neuroprotective Effect of Bean Phosphatidylserine on TMT-Induced Memory Deficits in a Rat Model" International Journal of Molecular Sciences 21, no. 14: 4901. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21144901