Genome-Wide Association Study Unravels LRK1 as a Dark Respiration Regulator in Rice (Oryza sativa L.)

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
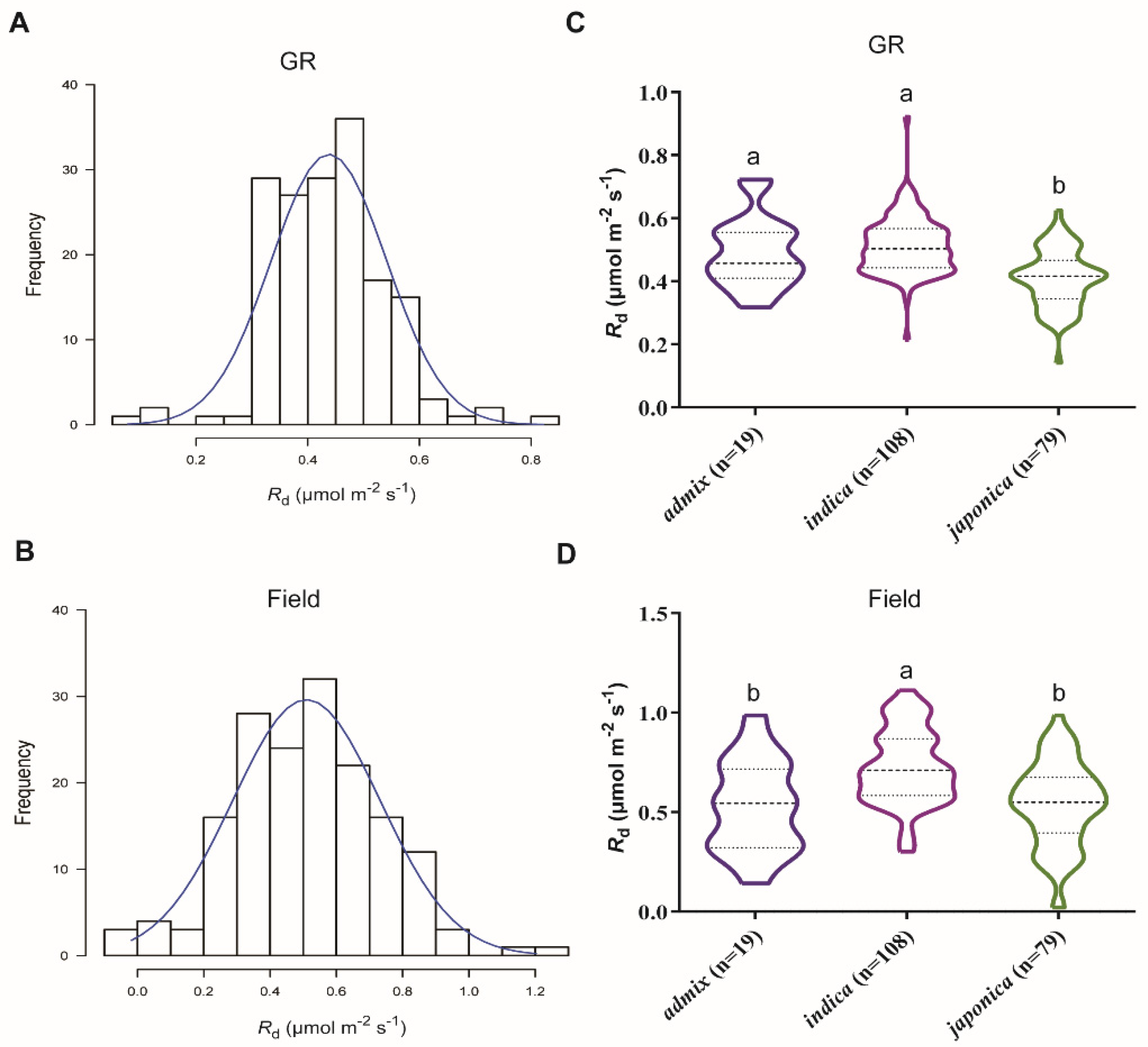
2.1. Genotypic Variation in Rd
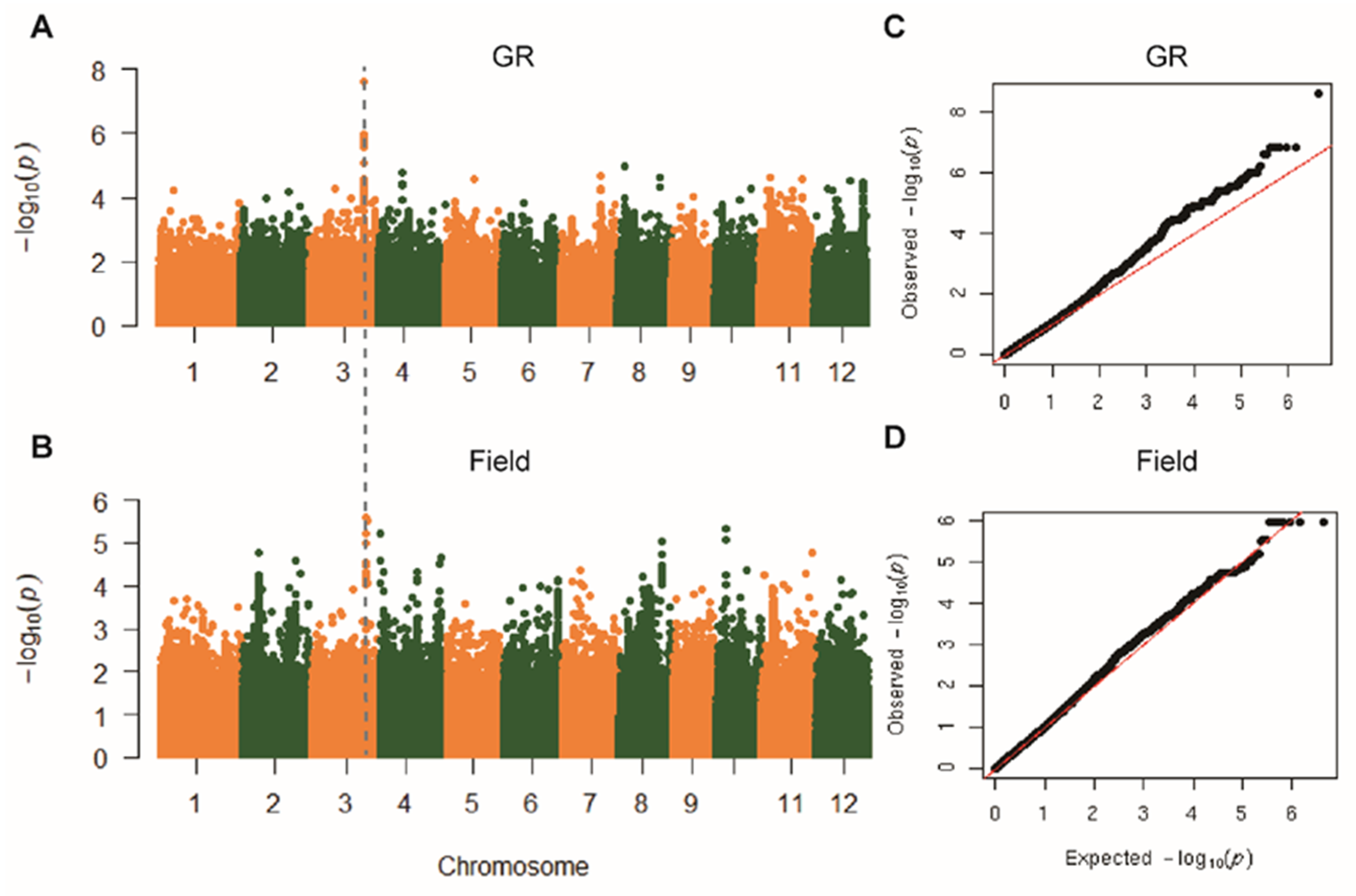
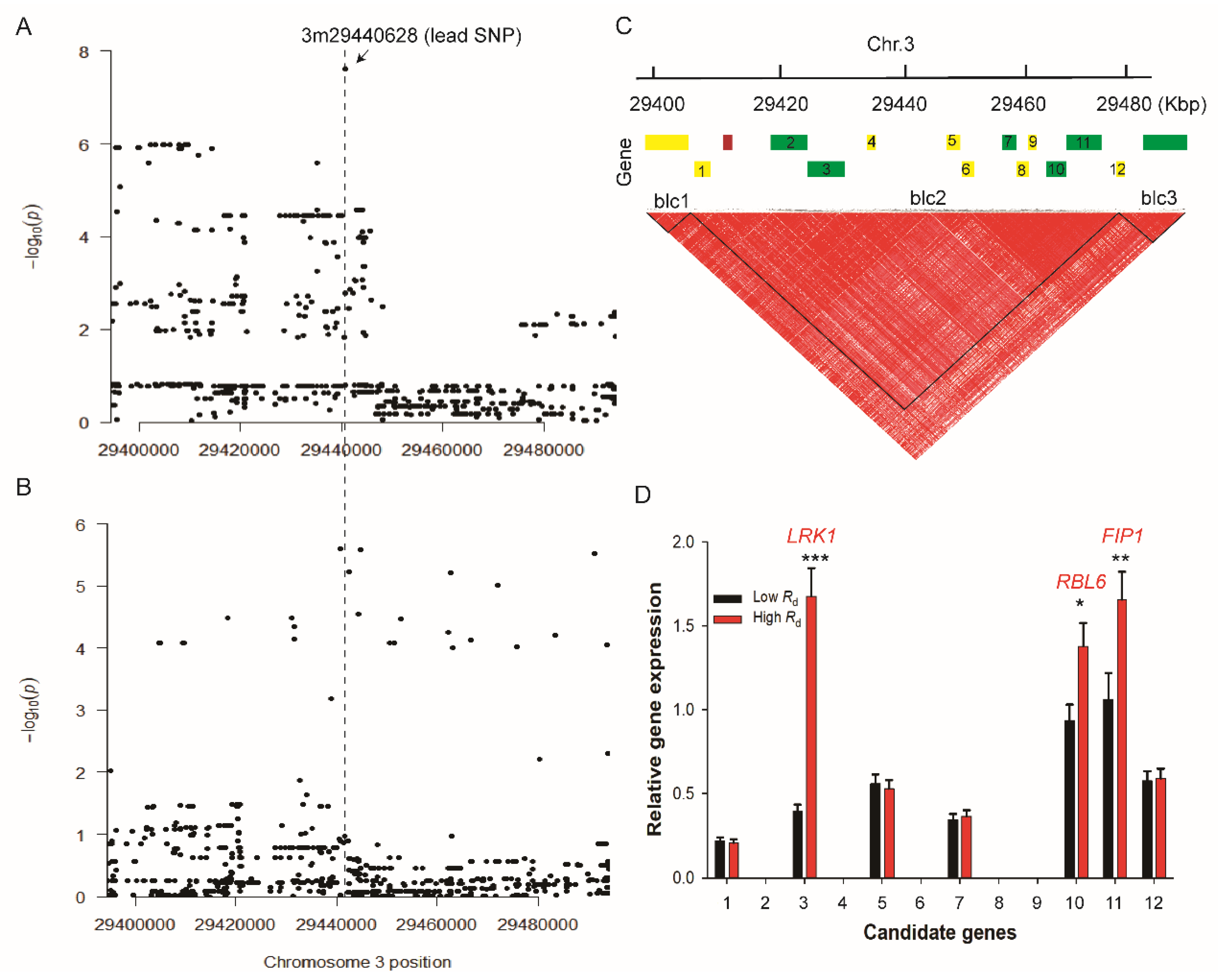
2.2. Genome-Wide Association Study on Rd
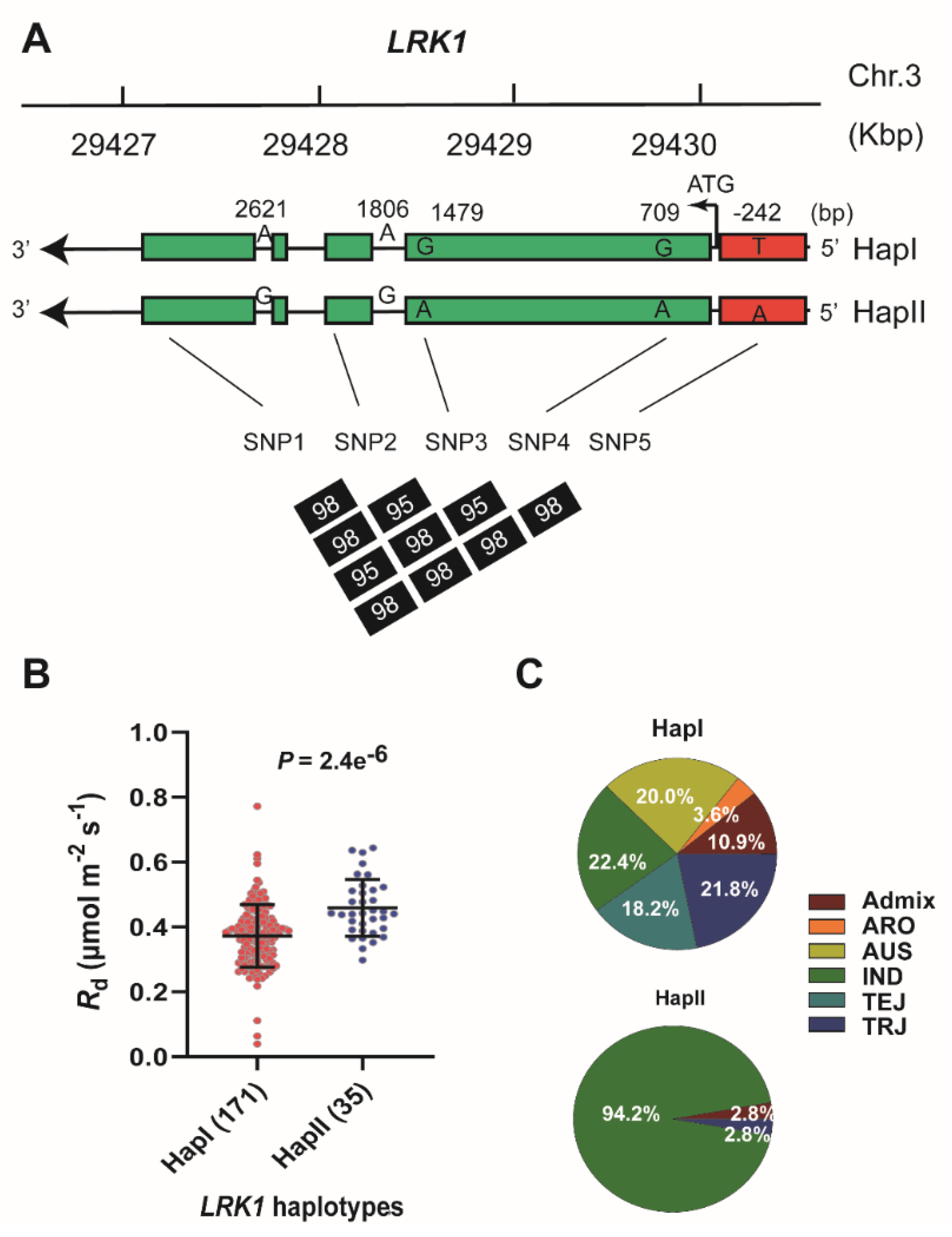
2.3. Haplotype Analysis of Candidate Genes Underlying Rd
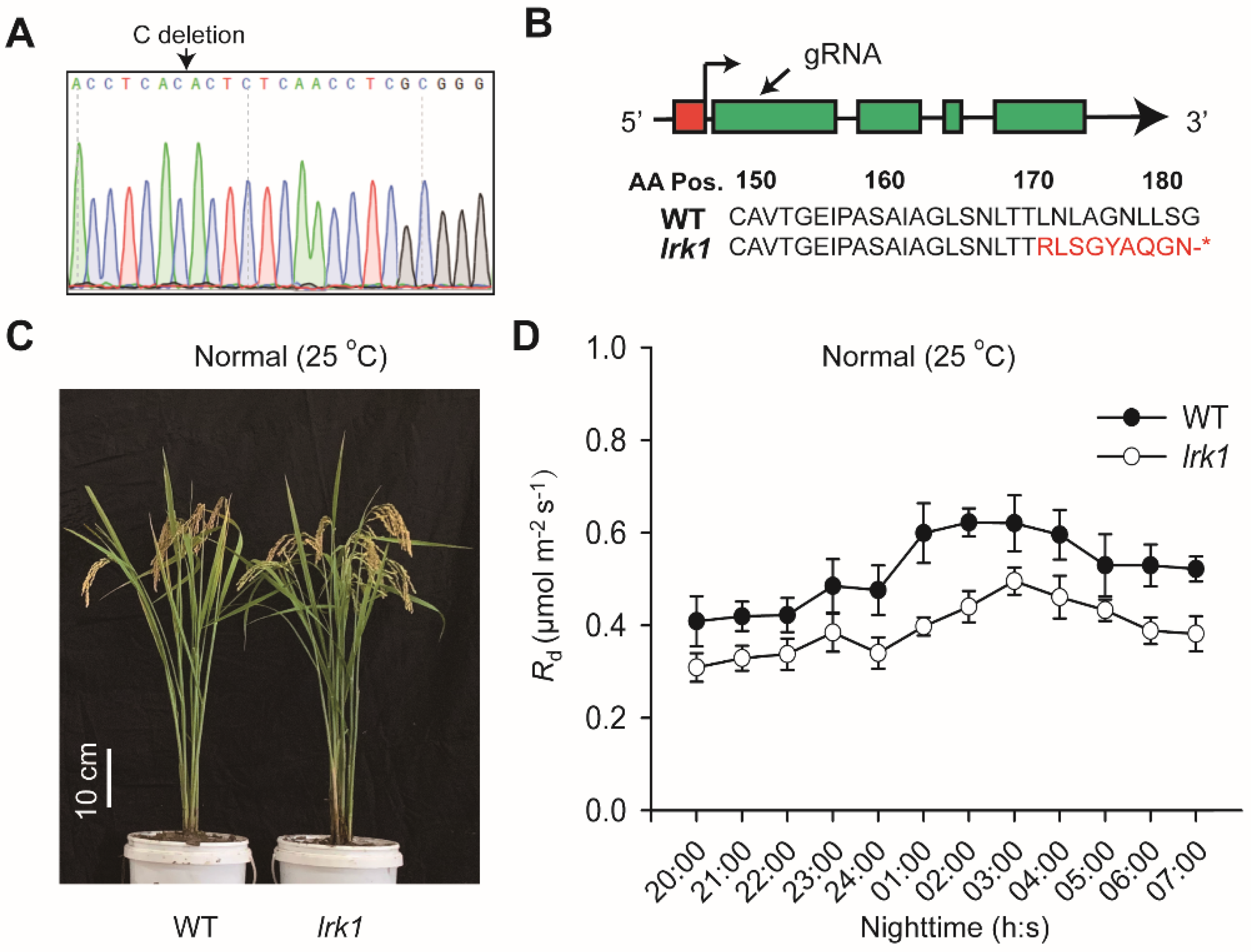
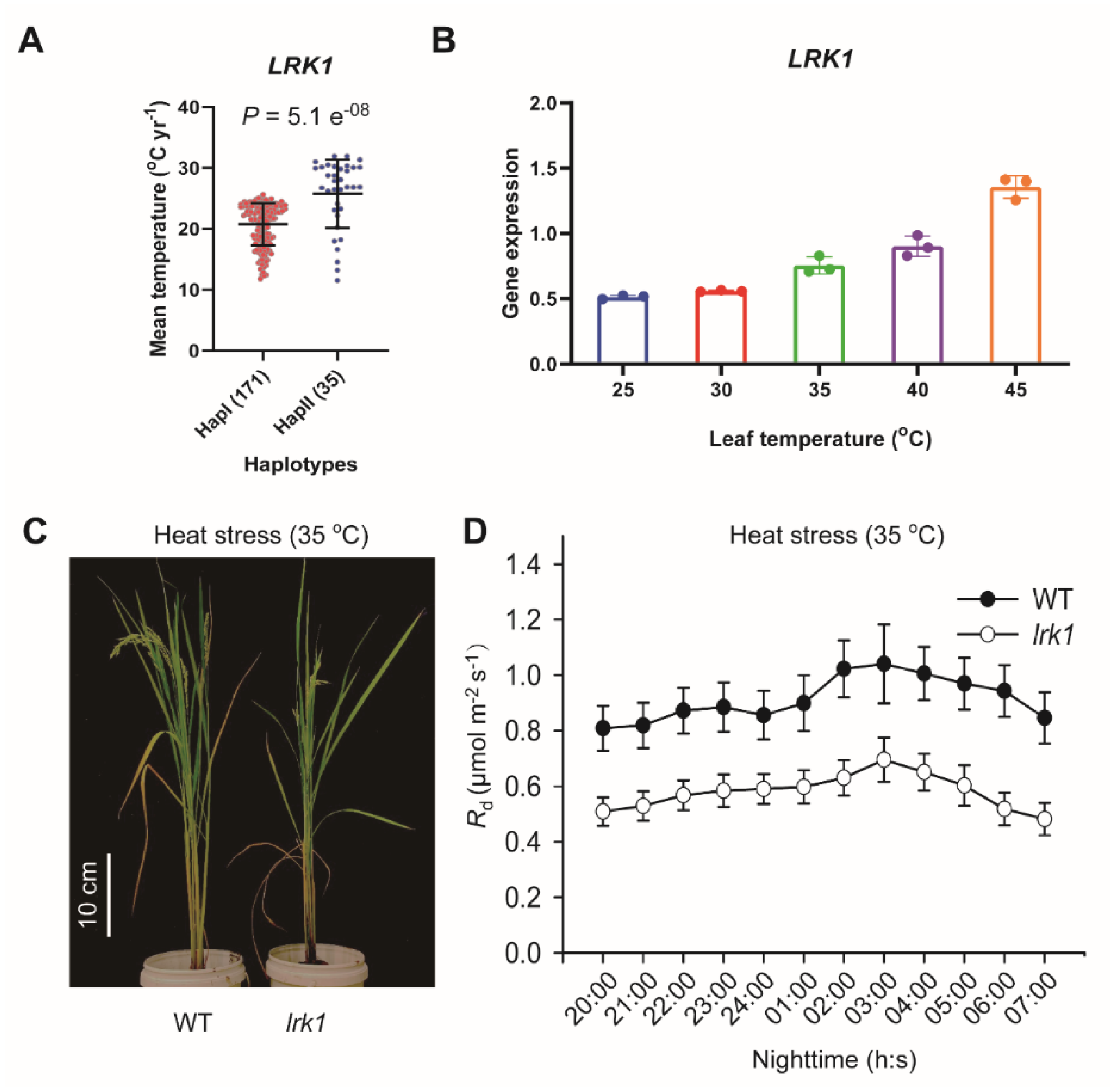
2.4. Biological Function of LRK1 Gene
3. Discussion
4. Materials and Methods
4.1. Rice Minicore Population
4.2. Measurements of Rd and Leaf Physiological Traits
4.3. Heat Stress Treatments
4.4. Genome-Wide Association Analysis
4.5. Transcript Abundance Analysis
4.6. Development of CRISPR-edited LRK1 Lines
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Xu, M.; DeBiase, T.; Qi, Y.; Goldstein, A.; Liu, Z.G. Ecosystem respiration in a young ponderosa pine plantation in the Sierra Nevada Mountains, California. Tree Physiol. 2001, 21, 309–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raich, J.; Schlesinger, W. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus B 1992, 44, 81–99. [Google Scholar] [CrossRef] [Green Version]
- Field, C.B. Plant physiology of the “missing” carbon sink. Plant Physiol. 2001, 125, 25–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Meler, M.A.; Taneva, L.; Trueman, R.J. Plant respiration and elevated atmospheric CO2 concentration: Cellular responses and global significance. Ann. Bot. 2004, 94, 647–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schimel, D.S. Terrestrial ecosystems and the carbon cycle. Glob. Chang. Biol. 1995, 1, 77–91. [Google Scholar] [CrossRef]
- Poorter, H.; Remkes, C.; Lambers, H. Carbon and Nitrogen Economy of 24 Wild Species Differing in Relative Growth Rate. Plant Physiol. 1990, 94, 621–627. [Google Scholar] [CrossRef] [Green Version]
- Atkin, O.K.; Botman, B.; Lambers, H. The Causes of Inherently Slow Growth in Alpine Plants: An Analysis Based on the Underlying Carbon Economies of Alpine and Lowland Poa Species. Funct. Ecol. 1996, 10, 698–707. [Google Scholar] [CrossRef]
- Loveys, B.R.; Scheurwater, I.; Pons, T.L.; Fitter, A.H.; Atkin, O.K. Growth temperature influences the underlying components of relative growth rate: An investigation using inherently fast- and slow-growing plant species. Plant Cell Environ. 2002, 25, 975–988. [Google Scholar] [CrossRef]
- Hoefnagel, M.H.N.; Atkin, O.K.; Wiskich, J.T. Interdependence between chloroplasts and mitochondria in the light and the dark. Biochim. Biophys. Acta Bioenerg. 1998, 1366, 235–255. [Google Scholar] [CrossRef] [Green Version]
- Atkin, O.K.; Edwards, E.J.; Loveys, B.R. Response of root respiration to changes in temperature and its relevance to global warming. New Phytol. 2000, 147, 141–154. [Google Scholar] [CrossRef]
- Qu, M.; Zheng, G.; Hamdani, S.; Essemine, J.; Song, Q.; Wang, H.; Chu, C.; Sirault, X.; Zhu, X.G. Leaf photosynthetic parameters related to biomass accumulation in a global rice diversity survey. Plant Physiol. 2017, 175, 248–258. [Google Scholar] [CrossRef] [Green Version]
- Huber, S.C. Biochemical Mechanism for Regulation of Sucrose Accumulation in Leaves during Photosynthesis. Plant Physiol. 1989, 91, 656–662. [Google Scholar] [CrossRef] [Green Version]
- Noguchi, K. Effects of Light Intensity and Carbohydrate Status on Leaf and Root Respiration. In Plant Respiration; Lambers, H., Ribas-Carbo, M., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 63–83. ISBN 978-1-4020-3589-0. [Google Scholar]
- Lee, K.; Akita, S. Factors Causing the Variation in the Temperature Coefficient of Dark Respiration in Rice (Oryza sativa L.). Plant Prod. Sci. 2000, 3, 38–42. [Google Scholar] [CrossRef]
- Azcon-Bieto, J.; Day, D.A.; Lambers, H. The regulation of respiration in the dark in wheat leaf slices. Plant Sci. Lett. 1983, 32, 313–320. [Google Scholar] [CrossRef]
- Noguchi, K.; Nakajima, N.; Terashima, I. Acclimation of leaf respiratory properties in Alocasia odora following reciprocal transfers of plants between high- and low-light environments. Plant. Cell Environ. 2001, 24, 831–839. [Google Scholar] [CrossRef]
- Amthor, J.S. The McCree–de Wit–Penning de Vries–Thornley Respiration Paradigms: 30 Years Later. Ann. Bot. 2000, 86, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Amthor, J.S. Crop Growth and Maintenance Respiration. In Respiration and Crop Productivity; Amthor, J.S., Ed.; Springer: New York, NY, USA, 1989; pp. 69–104. ISBN 978-1-4615-9667-7. [Google Scholar]
- Zagdanska, B. Respiratory energy demand for protein turnover and ion transport in wheat leaves upon water deficit. Physiol. Plant. 2006, 95, 428–436. [Google Scholar] [CrossRef]
- Atkin, O.K.; Scheurwater, I.; Pons, T.L. Respiration as a percentage of daily photosynthesis in whole plants is homeostatic at moderate, but not high, growth temperatures. New Phytol. 2007, 174, 367–380. [Google Scholar] [CrossRef]
- Barneix, A.; Cooper, H.; Stulen, I.; Lambers, H. Metabolism and translocation of nitrogen in two Lolium population with contrasting rates mature leaf respiraiion and yield. Physiol. Plant. 2007, 72, 631–636. [Google Scholar] [CrossRef]
- Leary, B.M.; Lee, C.P.; Atkin, O.K.; Cheng, R.; Brown, T.B.; Millar, A.H. Variation in leaf respiration rates at night correlates with carbohydrate and amino acid supply. Plant Physiol. 2017, 174, 2261–2273. [Google Scholar]
- Huang, X.; Wei, X.; Sang, T.; Zhao, Q.; Feng, Q.; Zhao, Y.; Li, C.; Zhu, C.; Lu, T.; Zhang, Z.; et al. Genome-wide asociation studies of 14 agronomic traits in rice landraces. Nat. Genet. 2010, 42, 961. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Han, B. Natural variations and genome-wide association studies in crop plants. Annu. Rev. Plant Biol. 2014, 65, 531–551. [Google Scholar] [CrossRef]
- Hamdani, S.; Wang, H.; Zheng, G.; Perveen, S.; Qu, M.; Khan, N.; Khan, W.; Jiang, J.; Li, M.; Liu, X.; et al. Genome-wide association study identifies variation of glucosidase being linked to natural variation of the maximal quantum yield of photosystem II. Physiol. Plant. 2019, 166, 105–119. [Google Scholar] [CrossRef] [PubMed]
- Agrama, H.A.; Yan, W.G.; Lee, F.; Robert, F.; Chen, M.H.; Jia, M.; McClung, A. Genetic assessment of a mini-core subset developed from the USDA rice genebank. Crop Sci. 2009, 49, 1336–1346. [Google Scholar] [CrossRef]
- Wang, H.; Xu, X.; Vieira, F.G.; Xiao, Y.; Li, Z.; Wang, J.; Nielsen, R.; Chu, C. The Power of Inbreeding: NGS-Based GWAS of Rice Reveals Convergent Evolution during Rice Domestication. Mol. Plant 2016, 9, 975–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Stephens, M. Genome-wide efficient mixed-model analysis for association studies. Nat. Genet. 2012, 44, 821–824. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Guo, Z.; Huang, C.; Duan, L.; Chen, G.; Jiang, N.; Fang, W.; Feng, H.; Xie, W.; Lian, X.; et al. Combining high-throughput phenotyping and genome-wide association studies to reveal natural genetic variation in rice. Nat. Commun. 2014, 5, 5087. [Google Scholar] [CrossRef]
- Berry, J.A.; Raison, J.K. Responses of Macrophytes to Temperature. In Physiological Plant Ecology I; Lange, O.L., Nobel, P.S., Osmond, C.B., Ziegler, H., Eds.; Springer: Berlin, Germany, 1981; pp. 277–338. ISBN 978-3-642-68090-8. [Google Scholar]
- Atkin, O.K.; Bloomfield, K.J.; Reich, P.B.; Tjoelker, M.G.; Asner, G.P.; Bonal, D.; Bönisch, G.; Bradford, M.G.; Cernusak, L.A.; Cosio, E.G.; et al. Global variability in leaf respiration in relation to climate, plant functional types and leaf traits. New Phytol. 2015, 206, 614–636. [Google Scholar] [CrossRef] [Green Version]
- Wright, I.J.; Reich, P.B.; Atkin, O.K.; Lusk, C.H.; Tjoelker, M.G.; Westoby, M. Irradiance, temperature and rainfall influence leaf dark respiration in woody plants: Evidence from comparisons across 20 sites. New Phytol. 2006, 169, 309–319. [Google Scholar] [CrossRef]
- Reich, P.B.; Tjoelker, M.G.; Pregitzer, K.S.; Wright, I.J.; Oleksyn, J.; Machado, J.-L. Scaling of respiration to nitrogen in leaves, stems and roots of higher land plants. Ecol. Lett. 2008, 11, 793–801. [Google Scholar] [CrossRef]
- Zhu, X.G.; Long, S.P.; Ort, D.R. What is the maximum efficiency with which photosynthesis can convert solar energy into biomass? Curr. Opin. Biotechnol. 2008, 19, 153–159. [Google Scholar] [CrossRef]
- Paul, V.; Pandey, R.; Anand, A. Measurement of plant respiration by infrared gas analyzer (IRGA). In Manual of ICAR Sponsored Training Programme for Technical Staff of ICAR Institutes on Physiological Techniques to Analyze the Impact of Climate Change on Crop Plants; ICAR-Indian Agricultural Research Institute (IARI): New Delhi, India, 2017. [Google Scholar]
- Poorter, H.; Van Rijn, C.P.E.; Vanhala, T.K.; Verhoeven, K.J.F.; De Jong, Y.E.M.; Stam, P.; Lambers, H. A genetic analysis of relative growth rate and underlying components in Hordeum spontaneum. Oecologia 2005, 142, 360–377. [Google Scholar] [CrossRef] [Green Version]
- Shiu, S.-H.; Karlowski, W.M.; Pan, R.; Tzeng, Y.-H.; Mayer, K.F.X.; Li, W.-H. Comparative analysis of the receptor-like kinase family in Arabidopsis and rice. Plant Cell 2004, 16, 1220–1234. [Google Scholar] [CrossRef] [Green Version]
- Osakabe, Y.; Arinaga, N.; Umezawa, T.; Katsura, S.; Nagamachi, K.; Tanaka, H.; Ohiraki, H.; Yamada, K.; Seo, S.-U.; Abo, M.; et al. Osmotic Stress Responses and Plant Growth Controlled by Potassium Transporters in Arabidopsis. Plant Cell 2013, 25, 609–624. [Google Scholar] [CrossRef] [Green Version]
- Diévart, A.; Clark, S.E. LRR-containing receptors regulating plant development and defense. Development 2004, 131, 251–261. [Google Scholar] [CrossRef] [Green Version]
- Zan, Y.; Ji, Y.; Zhang, Y.; Yang, S.; Song, Y.; Wang, J. Genome-wide identification, characterization and expression analysis of populus leucine-rich repeat receptor-like protein kinase genes. BMC Genomics 2013, 14, 318. [Google Scholar] [CrossRef] [Green Version]
- Song, W.Y.; Wang, G.L.; Chen, L.L.; Kim, H.S.; Pi, L.Y.; Holsten, T.; Gardner, J.; Wang, B.; Zhai, W.X.; Zhu, L.H.; et al. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science 1995, 270, 1804–1806. [Google Scholar] [CrossRef] [Green Version]
- Zha, X.; Luo, X.; Qian, X.; He, G.; Yang, M.; Li, Y.; Yang, J. Over-expression of the rice LRK1 gene improves quantitative yield components. Plant Biotechnol. J. 2009, 7, 611–620. [Google Scholar] [CrossRef]
- Ehleringer, J.; Björkman, O. Quantum yiel for CO2 uptake in C3 and C4 plants. Dependence on temperature, CO2 and O2 concentration. Plant Physiol. 1977, 59, 86–90. [Google Scholar] [CrossRef] [Green Version]
- Lusk, C.H.; Reich, P.B. Relationships of leaf dark respiration with light environment and tissue nitrogen content in juveniles of 11 cold-temperate tree species. Oecologia 2000, 123, 318–329. [Google Scholar] [CrossRef]
- Atkin, O.K.; Tjoelker, M.G. Thermal acclimation and the dynamic response of plant respiration to temperature. Trends Plant Sci. 2003, 8, 343–351. [Google Scholar] [CrossRef]
- Slot, M.; Rey-Sánchez, C.; Gerber, S.; Lichstein, J.W.; Winter, K.; Kitajima, K. Thermal acclimation of leaf respiration of tropical trees and lianas: Response to experimental canopy warming, and consequences for tropical forest carbon balance. Glob. Chang. Biol. 2014, 20, 2915–2926. [Google Scholar] [CrossRef]
- Agrama, H.A.; Yan, W.; Jia, M.; Fjellstrom, R.; McClung, A.M. Genetic structure associated with diversity and geographic distribution in the USDA rice world collection. Nat. Sci. 2010, 2, 247–291. [Google Scholar] [CrossRef] [Green Version]
- Qu, M.; Hamdani, S.; Li, W.; Wang, S.; Tang, J.; Chen, Z.; Song, Q.; Li, M.; Zhao, H.; Chang, T.; et al. Rapid stomatal response to fluctuating light: An under-explored mechanism to improve drought tolerance in rice. Funct. Plant Biol. 2016, 43, 727–738. [Google Scholar] [CrossRef] [Green Version]
- Korneliussen, T.S.; Albrechtsen, A.; Nielsen, R. ANGSD: Analysis of Next Generation Sequencing Data. BMC Bioinform. 2014, 15, 356. [Google Scholar] [CrossRef] [Green Version]
- Browning, S.R.; Browning, B.L. Rapid and accurate haplotype phasing and missing-data inference for whole-genome association studies by use of localized haplotype clustering. Am. J. Hum. Genet. 2007, 81, 1084–1097. [Google Scholar] [CrossRef] [Green Version]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; De Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Turner, S.D. qqman: An R package for visualizing GWAS results using Q-Q and manhattan plots. BioRxiv 2014. [Google Scholar] [CrossRef]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2004, 21, 263–265. [Google Scholar] [CrossRef] [Green Version]
- Gabriel, S.B.; Schaffner, S.F.; Nguyen, H.; Moore, J.M.; Roy, J.; Blumenstiel, B.; Higgins, J.; DeFelice, M.; Lochner, A.; Faggart, M.; et al. The structure of haplotype blocks in the human genome. Science 2002, 296, 2225–2229. [Google Scholar] [CrossRef] [Green Version]
- Devlin, B.; Risch, N. A comparison of linkage disequilibrium measures for fine-scale mapping. Genomics 1995, 29, 311–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C (T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Lee, S.H.; Goddard, M.E.; Visscher, P.M. GCTA: A tool for genome-wide complex trait analysis. Am. J. Hum. Genet. 2011, 88, 76–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qu, M.; Essemine, J.; Li, M.; Chang, S.; Chang, T.; Chen, G.-Y.; Zhu, X.-G. Genome-Wide Association Study Unravels LRK1 as a Dark Respiration Regulator in Rice (Oryza sativa L.). Int. J. Mol. Sci. 2020, 21, 4930. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21144930
Qu M, Essemine J, Li M, Chang S, Chang T, Chen G-Y, Zhu X-G. Genome-Wide Association Study Unravels LRK1 as a Dark Respiration Regulator in Rice (Oryza sativa L.). International Journal of Molecular Sciences. 2020; 21(14):4930. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21144930
Chicago/Turabian StyleQu, Mingnan, Jemaa Essemine, Ming Li, Shuoqi Chang, Tiangen Chang, Gen-Yun Chen, and Xin-Guang Zhu. 2020. "Genome-Wide Association Study Unravels LRK1 as a Dark Respiration Regulator in Rice (Oryza sativa L.)" International Journal of Molecular Sciences 21, no. 14: 4930. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21144930