ATG4 Mediated Psm ES4326/AvrRpt2-Induced Autophagy Dependent on Salicylic Acid in Arabidopsis Thaliana


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
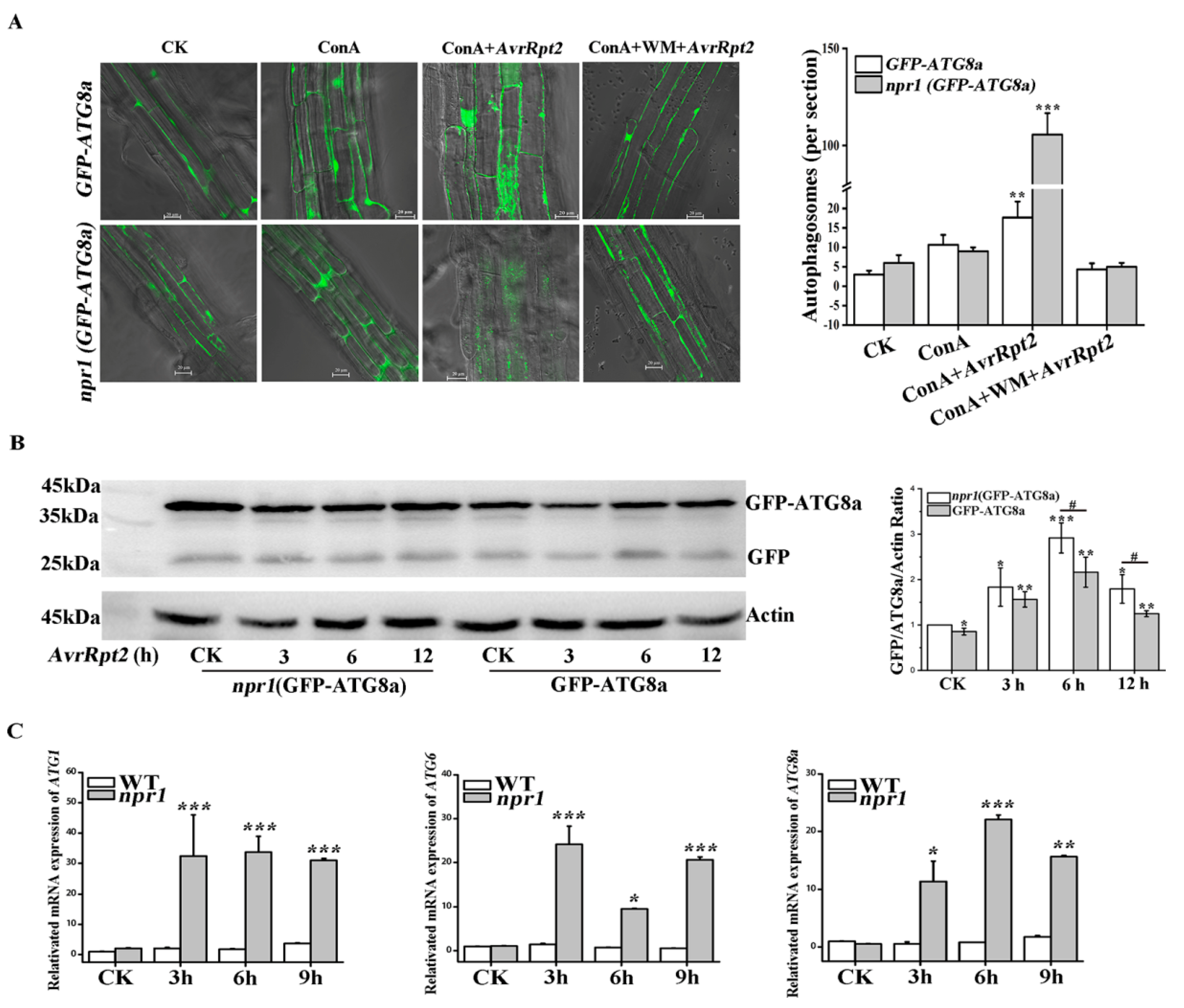
2.1. NPR1 Inhibited AvrRpt2-Induced Autophagy
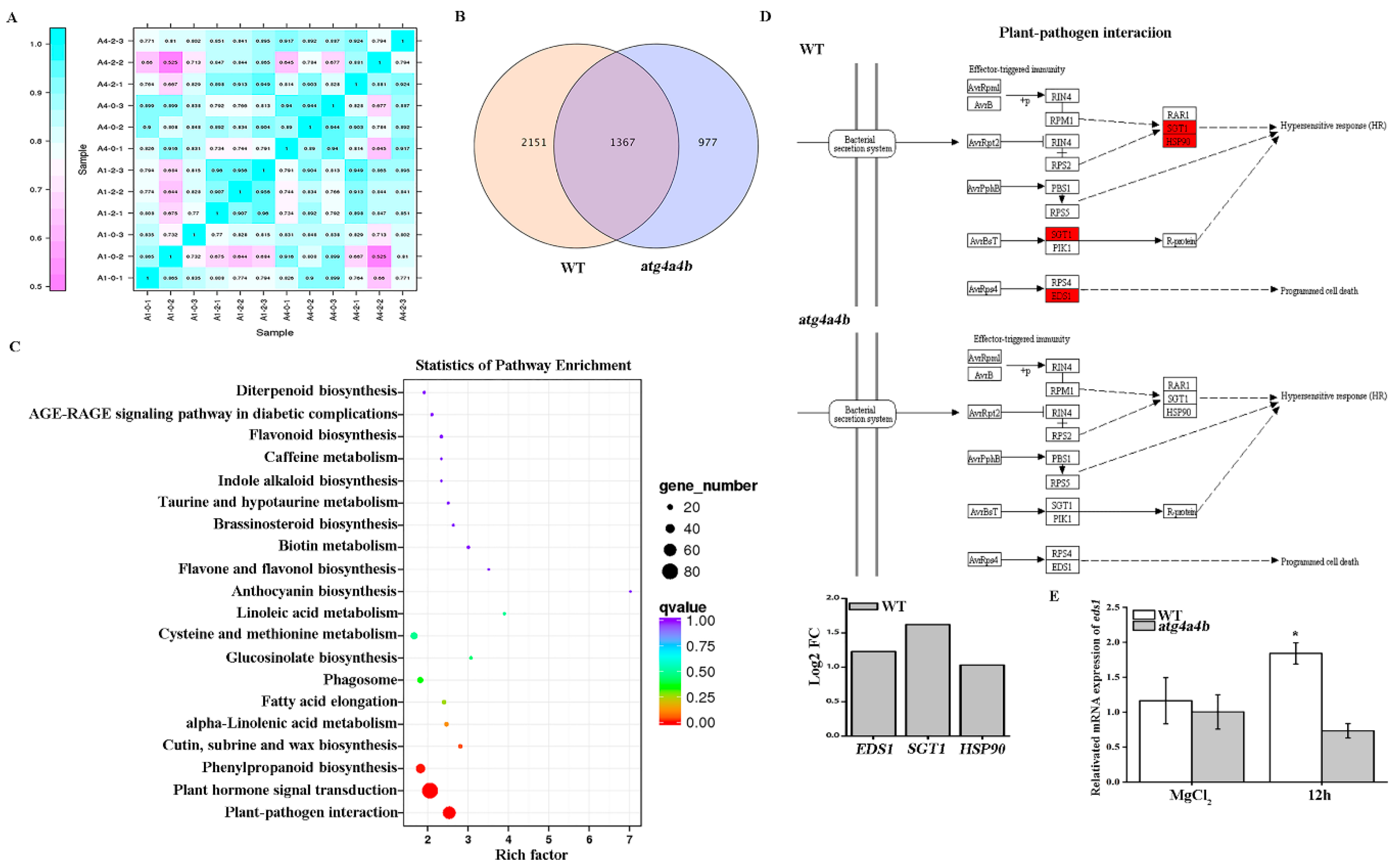
2.2. EDS1 Was Up-Regulated Under AvrRpt2 Infection
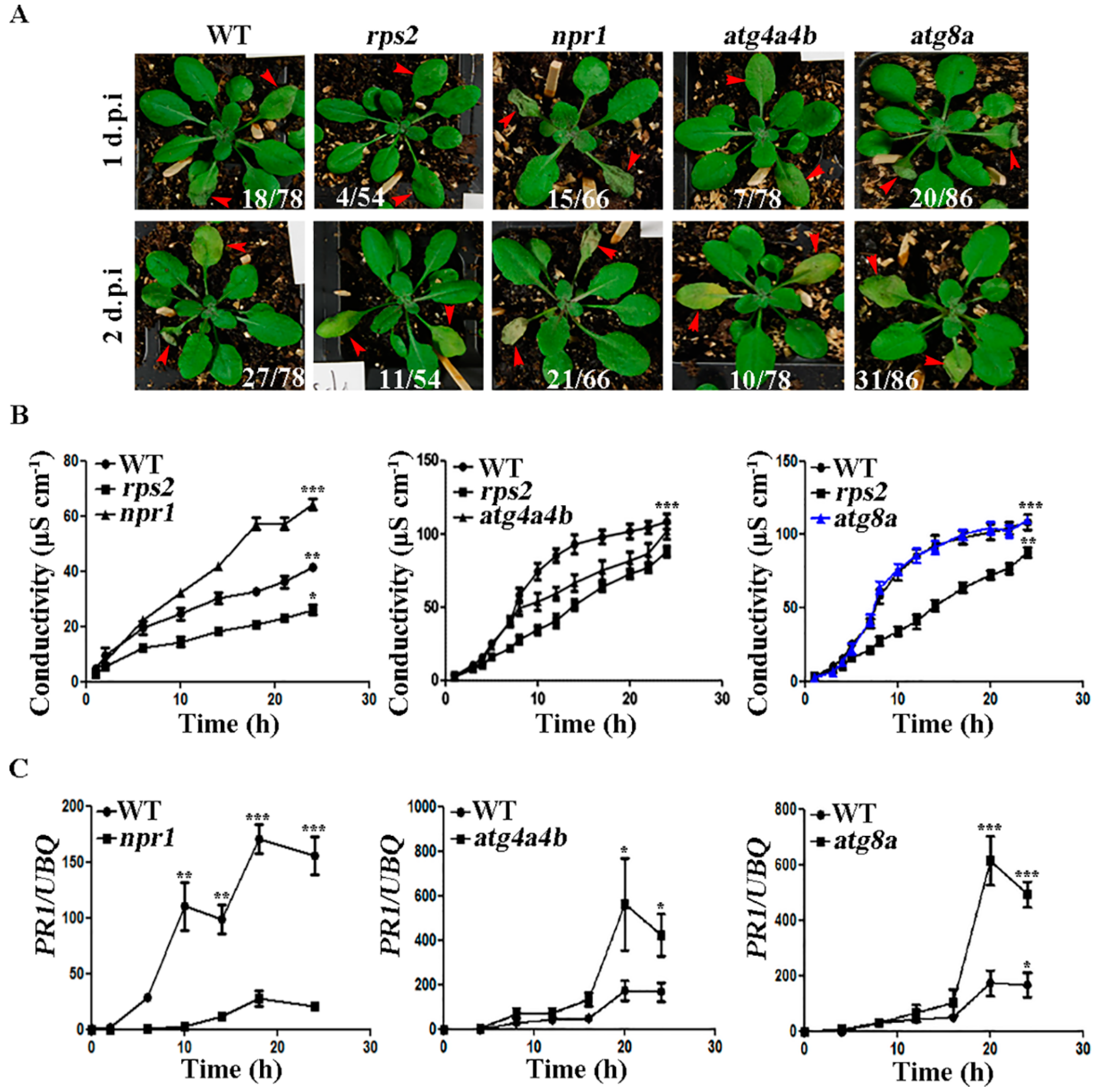
2.3. ATG4 Inhibited the Occurrence of HR during AvrRpt2 Infection
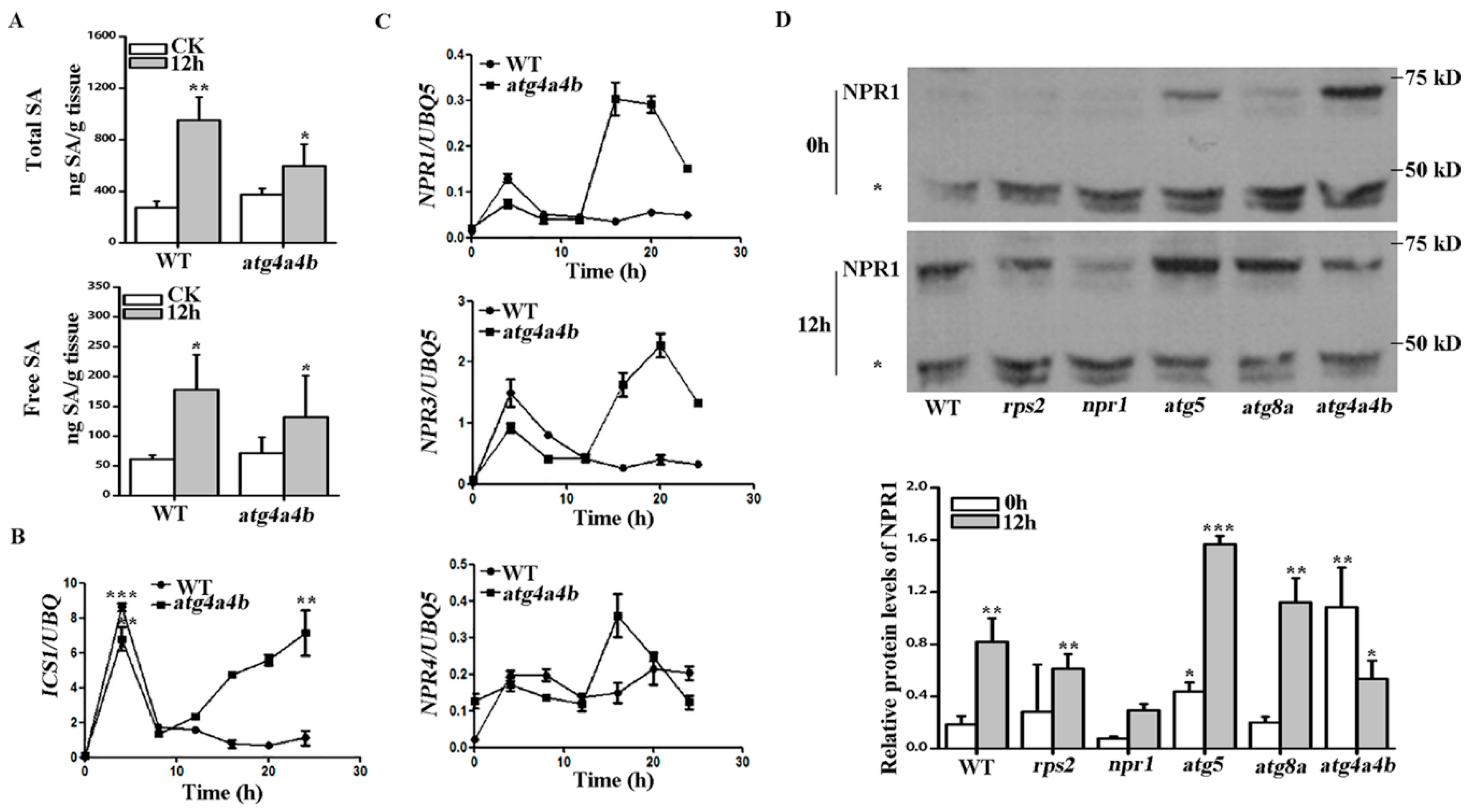
2.4. ATG4 Inhibited SA Consumption during AvrRpt2 Induced Autophagy-Dependent HR
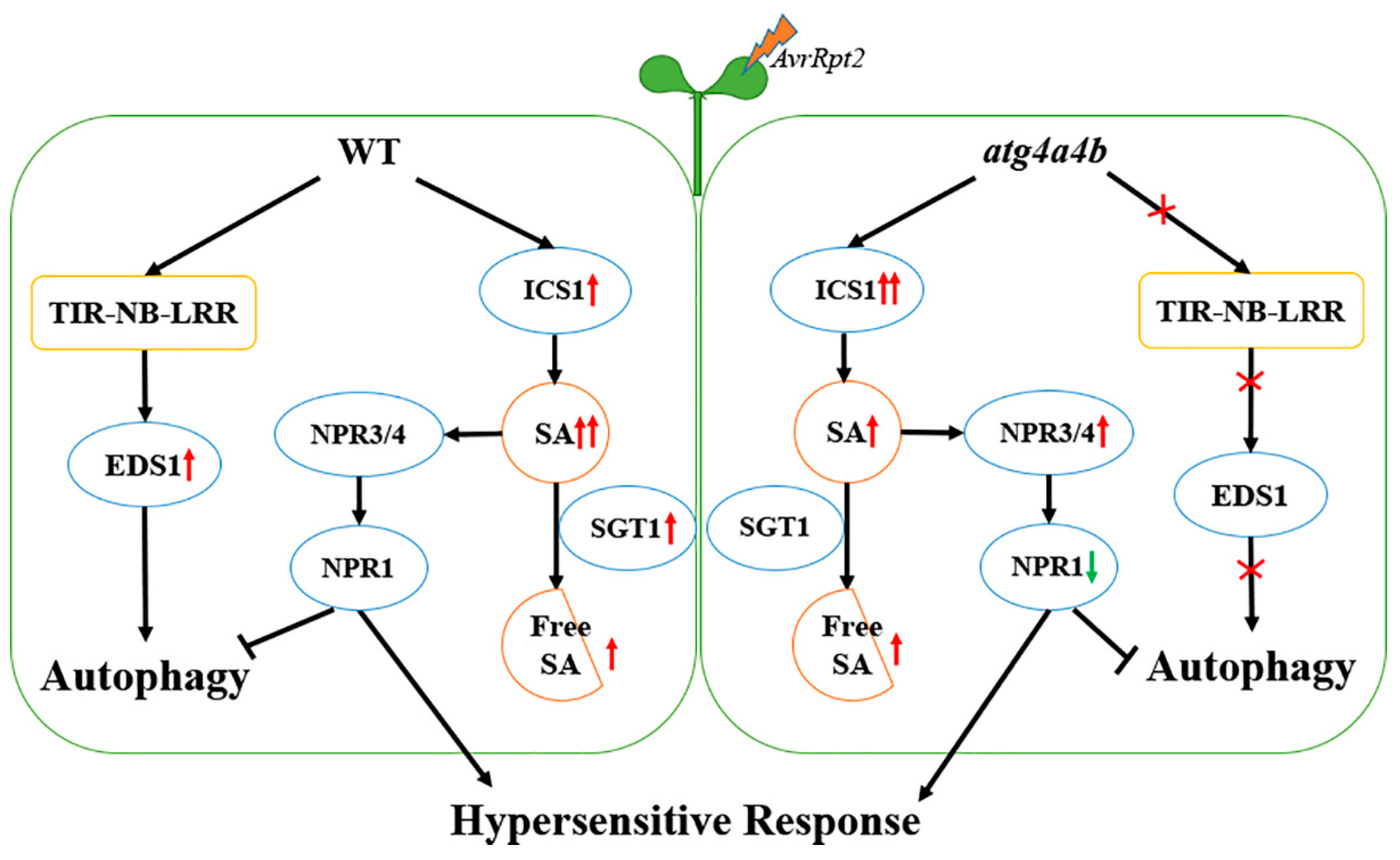
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Chemical Treatment
4.2. Pathogen Growth and Inoculation
4.3. SA Measurement
4.4. Total RNA Extraction and Quantitative Reverse Transcription-PCR (qRT-PCR)
4.5. Protein Extraction
4.6. Western Blotting
4.7. Confocal Microscopy
4.8. Ion Leakage
4.9. Transcriptome Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| NPR1 | nonexpressor of PR genes 1 |
| EDS1 | enhanced disease susceptibility 1 |
| SGT1 | salicylic acid glucosyltransferase 1 |
| SAG | salicylic acid 2-O-β-D-glucose |
| ICS1 | isochorismate synthase 1 |
| NDR1 | non-race specific disease resistance |
| PAL | phenylalanine ammonia lyase |
| PR | pathogenesis-related |
| SA | salicylic acid |
| JA | jasmonic acid |
| TIR-NB-LRR | Toll/Interleukin-1 receptor (TIR)-nucleotide binding site (NB)-leucine rich repeat (LRR) |
| CC-NB-LRR | coiled-coil (CC)-nucleotide binding site (NB)-leucine rich repeat (LRR) |
| PCD | programmed cell death |
| HR | hypersensitive response |
| PAMP | pathogen-associated molecular patterns |
| ETI | effector-triggered immunity |
| PTI | PAMP-triggered immunity |
| PRS2 | resistant to P. syringae 2 |
| PRRs | pattern recognition receptors |
| KEGG | Kyoto encyclopedia of genes and genomes |
| WM | wortmannin |
| ConA | concanamycin A |
References
- Mizushima, N.; Komatsu, M. Autophagy: Renovation of Cells and Tissues. Cell 2011, 147, 728–741. [Google Scholar] [CrossRef] [Green Version]
- Hofius, D.; Lie, L.; Hafren, A.; Coll, N.S. Autophagy as an emerging arena for plant-pathogen interactions. Curr. Opin. Plant Biol. 2017, 38, 117–123. [Google Scholar] [CrossRef]
- Il Kwon, S.; Park, O.K. Autophagy in Plants. J. Plant Biol. 2008, 51, 313–320. [Google Scholar] [CrossRef]
- Bisgrove, S.R.; Simonich, M.T.; Smith, N.M.; Sattler, A.; Innes, R.W. A disease resistance gene in Arabidopsis with specificity for two different pathogen avirulence genes. Plant. Cell 1994, 6, 927–933. [Google Scholar] [CrossRef] [Green Version]
- Hatsugai, N.; Igarashi, D.; Mase, K.; Lu, Y.; Tsuda, Y.; Chakravarthy, S.; Wei, H.L.; Foley, J.W.; Collmer, A.; Glazebrook, J.; et al. A plant effector-triggered immunity signaling sector is inhibited by pattern-triggered immunity. Embo J. 2017, 36, 2758–2769. [Google Scholar] [CrossRef] [PubMed]
- Hofius, D.; Schultz-Larsen, T.; Joensen, J.; Tsitsigiannis, D.I.; Petersen, N.H.T.; Mattsson, O.; Jorgensen, L.B.; Jones, J.D.G.; Mundy, J.; Petersen, M. Autophagic Components Contribute to Hypersensitive Cell Death in Arabidopsis. Cell 2009, 137, 773–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.Y.; Wang, X.D.; Ma, C.; Chen, W.L. EDS1-mediated activation of autophagy regulates Pst DC3000 (AvrRps4)-induced programmed cell death in Arabidopsis. Acta Physiol. Plant. 2016, 38. [Google Scholar] [CrossRef]
- Betsuyaku, S.; Katou, S.; Takebayashi, Y.; Sakakibara, H.; Nomura, N.; Fukuda, H. Salicylic Acid and Jasmonic Acid Pathways are Activated in Spatially Different Domains Around the Infection Site During Effector-Triggered Immunity in Arabidopsis thaliana. Plant Cell Physiol. 2018, 59, 8–16. [Google Scholar] [CrossRef]
- Wildermuth, M.C.; Dewdney, J.; Wu, G.; Ausubel, F.M. Isochorismate synthase is required to synthesize salicylic acid for plant defence. Nature 2001, 414, 562–565. [Google Scholar] [CrossRef]
- Sendon, P.M.; Seo, H.S.; Song, J.T. Salicylic Acid Signaling: Biosynthesis, Metabolism, and Crosstalk with Jasmonic Acid. J. Korean Soc. Appl. Biol. Chem. 2011, 54, 501–506. [Google Scholar] [CrossRef]
- Song, J.T.; Koo, Y.J.; Seo, H.S.; Kim, M.C.; Choi, Y.D.; Kim, J.H. Overexpression of AtSGT1, an Arabidopsis salicylic acid glucosyltransferase, leads to increased susceptibility to Pseudomonas syringae. Phytochemistry 2008, 69, 1128–1134. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.T.; Abas, M.; Verstraeten, I.; Glanc, M.S.; Molnar, G.; Hajny, J.; Lasak, P.; Petrik, I.; Russinova, E.; Petrasek, J.; et al. Salicylic Acid Targets Protein Phosphatase 2A to Attenuate Growth in Plants. Curr. Biol. 2020, 30, 381–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Z.Q.; Dong, X.N. Systemic Acquired Resistance: Turning Local Infection into Global Defense. Annu. Rev. Plant Biol. 2013, 64, 839–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.W.; Zhao, D.; Dong, J.W.; Kong, X.L.; Zhang, Q.; Li, T.T.; Meng, Y.L.; Shan, W.X. AtRTP5 negatively regulates plant resistance to Phytophthora pathogens by modulating the biosynthesis of endogenous jasmonic acid and salicylic acid. Mol. Plant Pathol. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimoto, K.; Jikumaru, Y.; Kamiya, Y.; Kusano, M.; Consonni, C.; Panstruga, R.; Ohsumi, Y.; Shirasu, K. Autophagy Negatively Regulates Cell Death by Controlling NPR1-Dependent Salicylic Acid Signaling during Senescence and the Innate Immune Response in Arabidopsis. Plant Cell 2009, 21, 2914–2927. [Google Scholar] [CrossRef] [Green Version]
- Fu, Z.Q.; Yan, S.; Saleh, A.; Wang, W.; Ruble, J.; Oka, N.; Mohan, R.; Spoel, S.H.; Tada, Y.; Zheng, N.; et al. NPR3 and NPR4 are receptors for the immune signal salicylic acid in plants. Nature 2012, 486, 228–232. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, X.H.; Wang, H.; Lam, S.K.; Gao, C.J.; Wang, X.F.; Cai, Y.; Jiang, L.W. A BAR-Domain Protein SH3P2, Which Binds to Phosphatidylinositol 3-Phosphate and ATG8, Regulates Autophagosome Formation in Arabidopsis. Plant Cell 2013, 25, 4596–4615. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Yu, L.J.; Zhang, X.; Fan, B.; Wang, F.Z.; Dai, Y.S.; Qi, H.; Zhou, Y.; Xie, L.J.; Xiao, S. Autophagy regulates glucose-mediated root meristem activity by modulating ROS production in Arabidopsis. Autophagy 2019, 15, 407–422. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.Y.; Xu, M.Y.; Wang, G.P.; Galili, G. Autophagy: An Important Biological Process That Protects Plants from Stressful Environments. Front. Plant Sci. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Yoo, H.; Greene, G.H.; Yuan, M.; Xu, G.Y.; Burton, D.; Liu, L.J.; Marques, J.; Dong, X.N. Translational Regulation of Metabolic Dynamics during Effector-Triggered Immunity. Mol. Plant 2020, 13, 88–98. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, M.; Zeier, J. N-hydroxypipecolic acid and salicylic acid: A metabolic duo for systemic acquired resistance. Curr. Opin. Plant Biol. 2019, 50, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Romera, F.J.; Garcia, M.J.; Lucena, C.; Martinez-Medina, A.; Aparicio, M.A.; Ramos, J.; Alcantara, E.; Angulo, M.; Perez-Vicente, R. Induced Systemic Resistance (ISR) and Fe Deficiency Responses in Dicot Plants. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Mackey, D.; Belkhadir, Y.; Alonso, J.M.; Ecker, J.R.; Dangl, J.L. Arabidopsis RIN4 is a target of the type III virulence effector AvrRpt2 and modulates RPS2-mediated resistance. Cell 2003, 112, 379–389. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.J.; Sonbol, F.M.; Huot, B.; Gu, Y.N.; Withers, J.; Mwimba, M.; Yao, J.; He, S.Y.; Dong, X.N. Salicylic acid receptors activate jasmonic acid signalling through a non-canonical pathway to promote effector-triggered immunity. Nat. Commun. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Kirisako, T.; Ichimura, Y.; Okada, H.; Kabeya, Y.; Mizushima, N.; Yoshimori, T.; Ohsumi, M.; Takao, T.; Noda, T.; Ohsumi, Y. The reversible modification regulates the membrane-binding state of Apg8/Aut7 essential for autophagy and the cytoplasm to vacuole targeting pathway. J. Cell Biol. 2000, 151, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Ichimura, Y.; Kirisako, T.; Takao, T.; Satomi, Y.; Shimonishi, Y.; Ishihara, N.; Mizushima, N.; Tanida, I.; Kominami, E.; Ohsumi, M.; et al. A ubiquitin-like system mediates protein lipidation. Nature 2000, 408, 488–492. [Google Scholar] [CrossRef]
- Wang, K.; Mysore, K.S. SGT1 is required for plant disease resistance and symptom development during disease susceptibility. Phytopathology 2006, 96, S120. [Google Scholar]
- Qian, L.C.; Zhao, J.P.; Du, Y.M.; Zhao, X.J.; Han, M.; Liu, Y.L. Hsp90 Interacts With Tm-2(2) and Is Essential for Tm-2(2)-Mediated Resistance to Tobacco mosaic virus. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.K.; Kwon, S.Y.; Choi, D. Rpi-blb2-Mediated Hypersensitive Cell Death Caused by Phytophthora infestans AVRblb2 Requires SGT1, but not EDS1, NDR1, Salicylic Acid-, Jasmonic Acid-, or Ethylene-Mediated Signaling. Plant. Pathol. J. 2014, 30, 254–260. [Google Scholar] [CrossRef]
- Wang, X.D.; Gao, Y.Y.; Yan, Q.Q.; Chen, W.L. Salicylic acid promotes autophagy via NPR3 and NPR4 in Arabidopsis senescence and innate immune response. Acta Physiol. Plant. 2016, 38. [Google Scholar] [CrossRef]
- Lv, F.; Zhou, J.; Zeng, L.; Xing, D. beta-cyclocitral upregulates salicylic acid signalling to enhance excess light acclimation in Arabidopsis. J. Exp. Bot. 2015, 66, 4719–4732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karppinen, K.; Taulavuori, E.; Hohtola, A. Optimization of protein extraction from Hypericum perforatum tissues and immunoblotting detection of Hyp-1 at different stages of leaf development. Mol. Biotechnol. 2010, 46, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.J.; Chen, W.L. The Role of Autophagy in Chloroplast Degradation and Chlorophagy in Immune Defenses during Pst DC3000 (AvrRps4) Infection. PLoS ONE 2013, 8, e0073091. [Google Scholar] [CrossRef] [PubMed] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, W.; Li, B.; Zhang, B.; Chen, W. ATG4 Mediated Psm ES4326/AvrRpt2-Induced Autophagy Dependent on Salicylic Acid in Arabidopsis Thaliana. Int. J. Mol. Sci. 2020, 21, 5147. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21145147
Gong W, Li B, Zhang B, Chen W. ATG4 Mediated Psm ES4326/AvrRpt2-Induced Autophagy Dependent on Salicylic Acid in Arabidopsis Thaliana. International Journal of Molecular Sciences. 2020; 21(14):5147. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21145147
Chicago/Turabian StyleGong, Wenjun, Bingcong Li, Baihong Zhang, and Wenli Chen. 2020. "ATG4 Mediated Psm ES4326/AvrRpt2-Induced Autophagy Dependent on Salicylic Acid in Arabidopsis Thaliana" International Journal of Molecular Sciences 21, no. 14: 5147. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21145147