Metabolic Response of the Yeast Candida utilis During Enrichment in Selenium

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results and Discussion
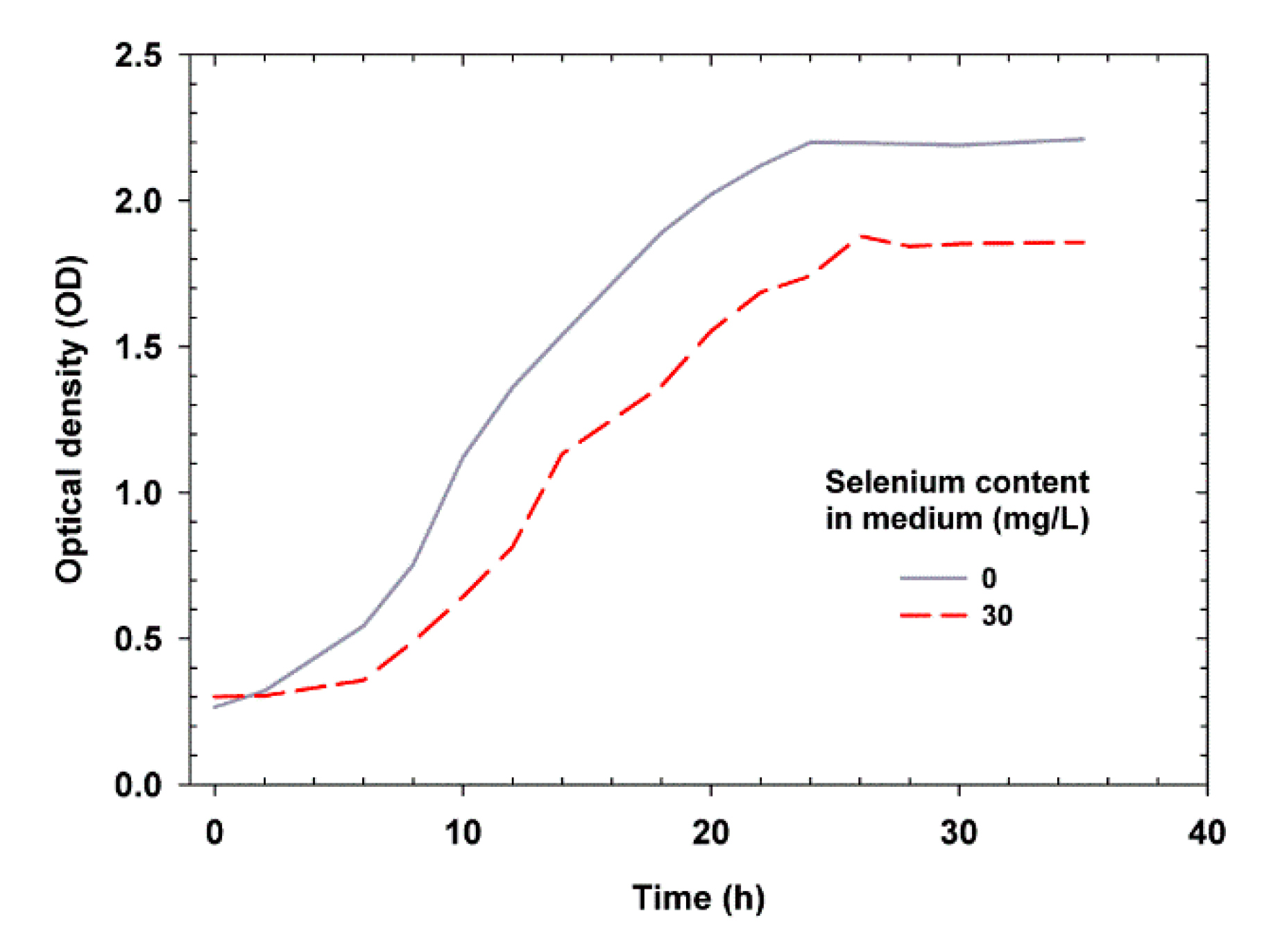
2.1. Effect of Se on Yeast Growth
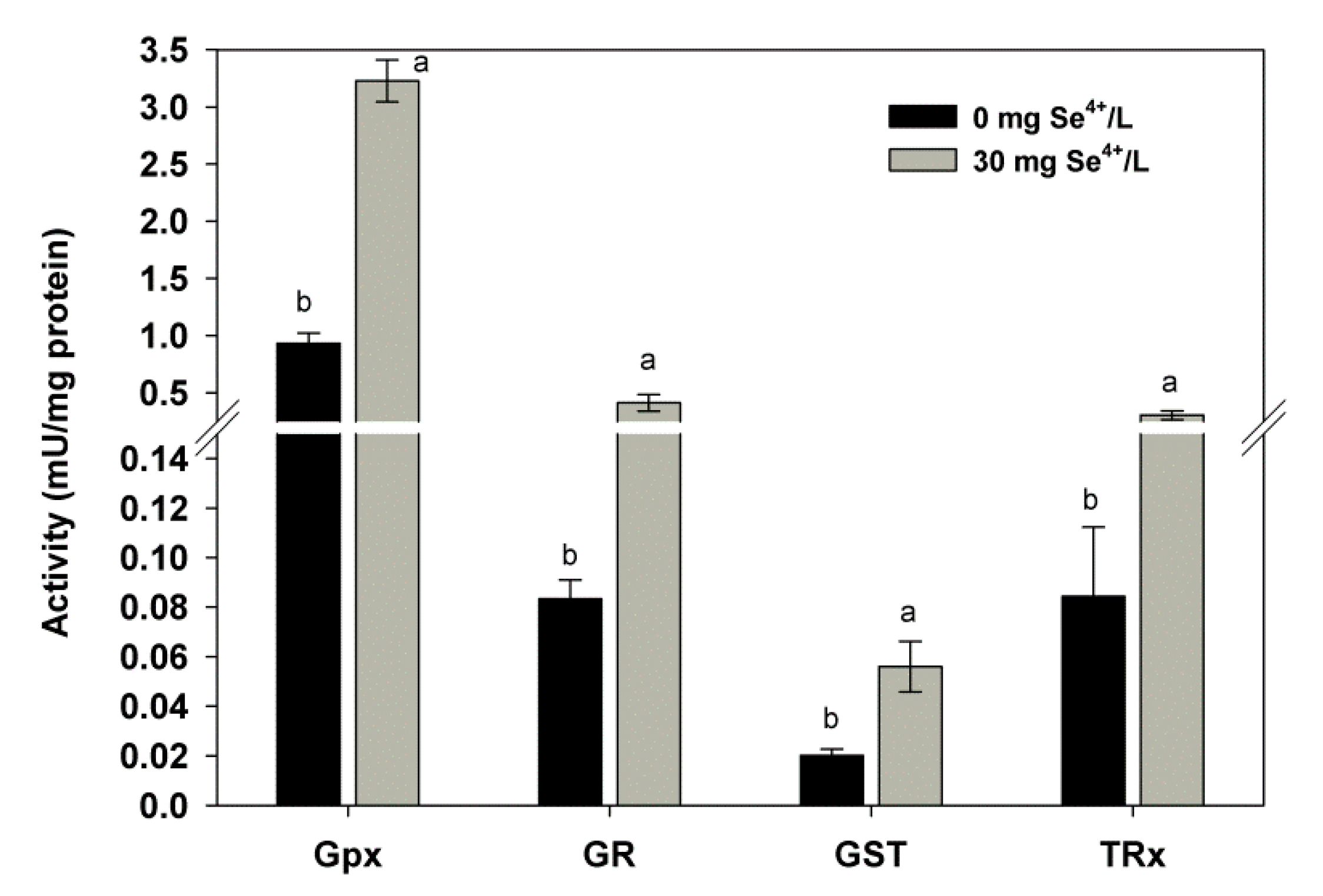
2.2. Effect of Se on the Activity of Antioxidant Enzymes
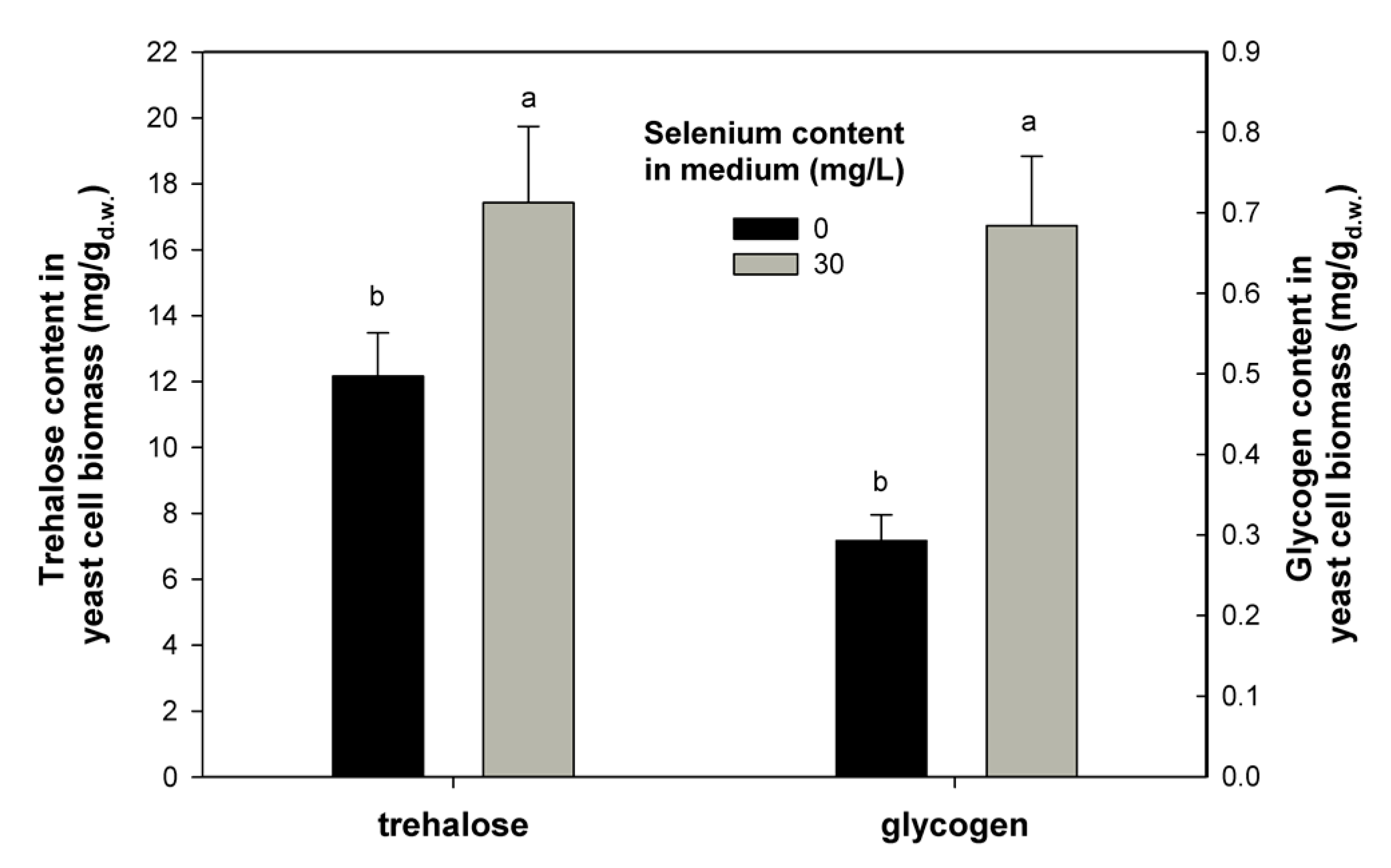
2.3. Effect of Se on Trehalose and Glycogen Content
2.4. Se Bioaccumulation
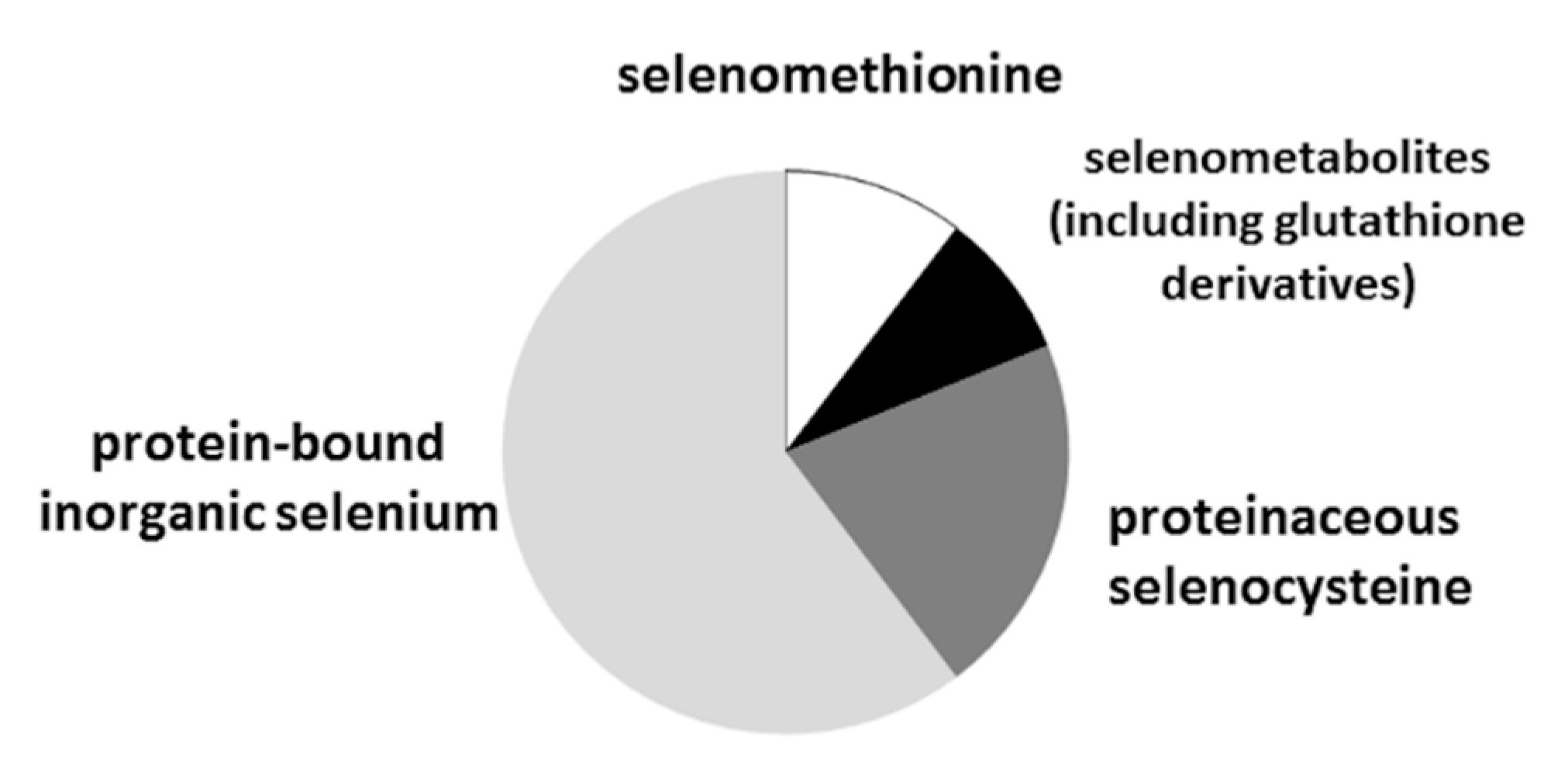
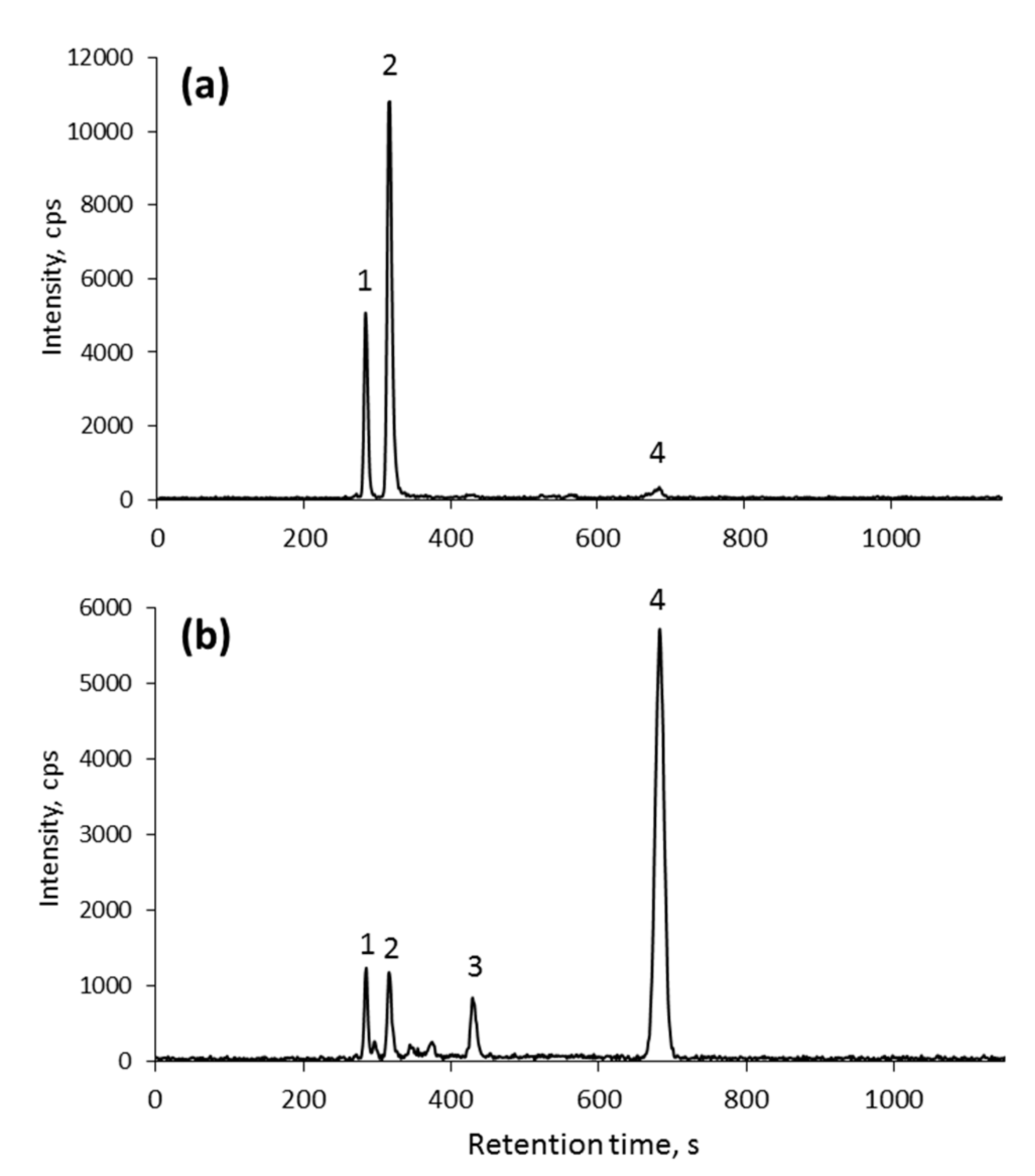
2.5. Se Speciation
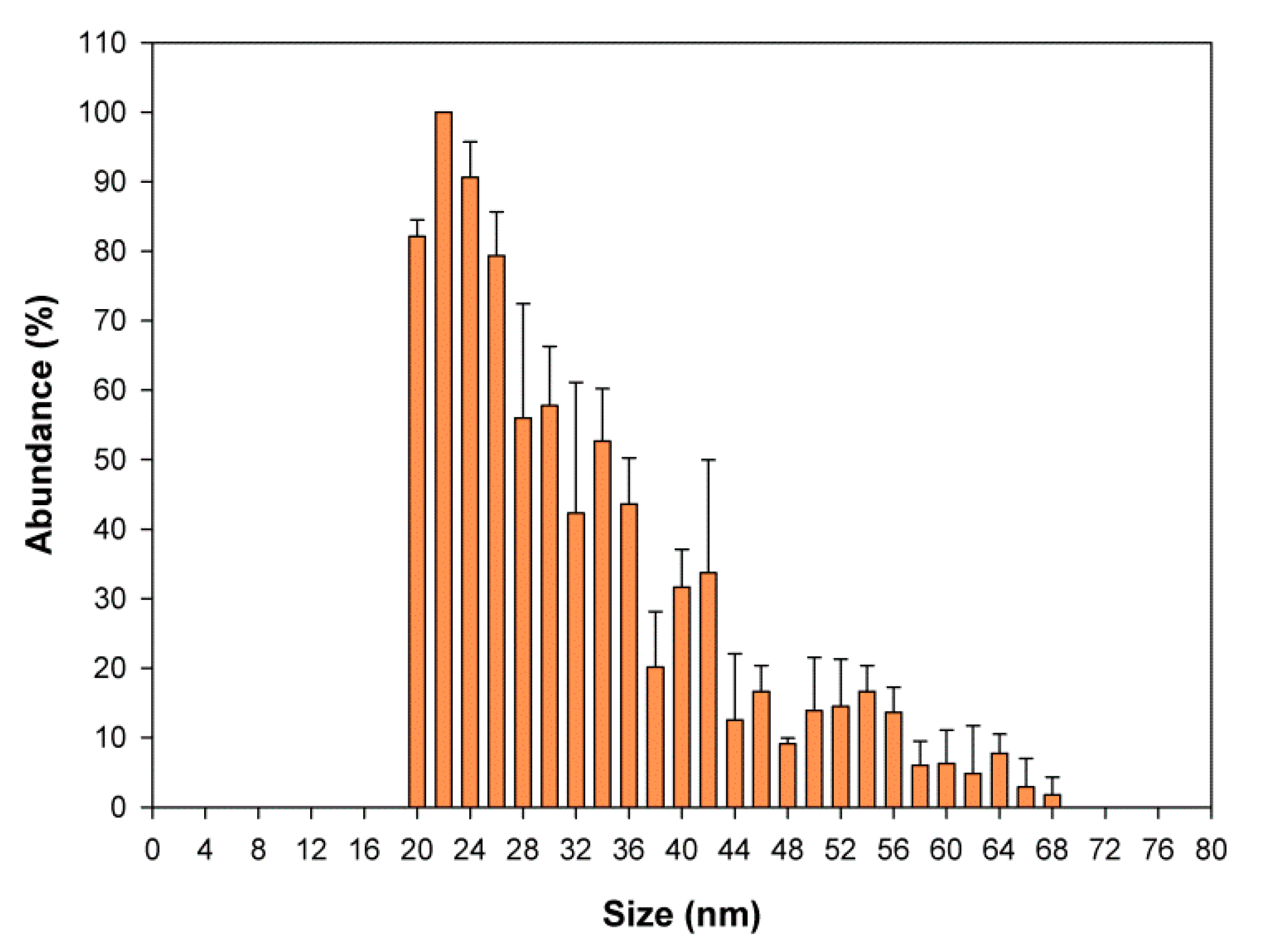
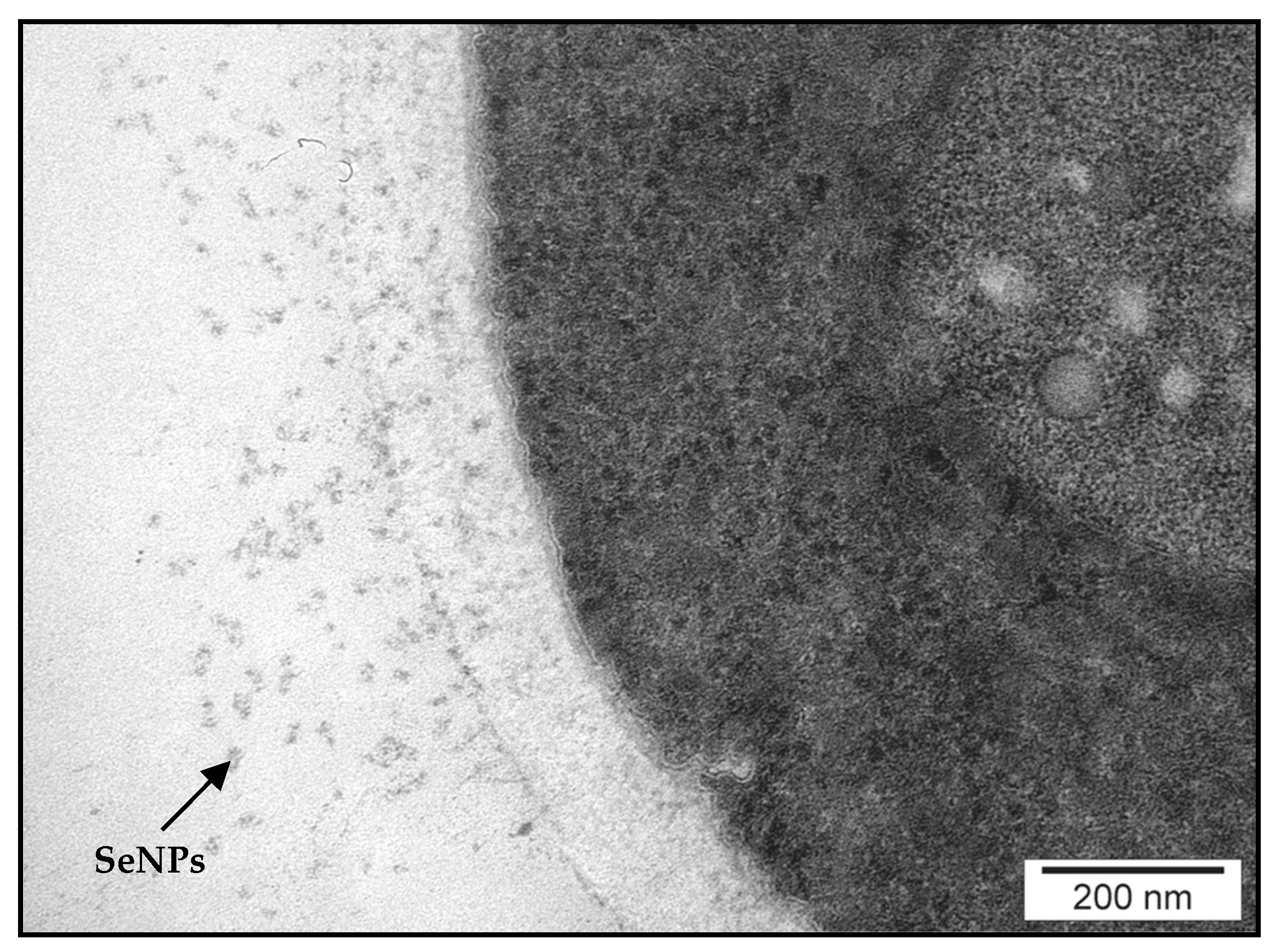
2.6. Detection and Characterization of SeNPs
3. Materials and Methods
3.1. Biological Material
3.2. Determination of Optical Density
3.3. Preparation of Cell Extract to Determine Trehalose, Glycogen and Antioxidant Enzyme Activity
3.4. Determination of Trehalose and Glycogen Content in Yeast Cell Biomass
3.5. Biochemical Determination of Antioxidant Enzymes
3.6. Determination of Protein Content
3.7. Yeast Observation Under an Electron Microscope
3.8. Total Se Determination
3.9. Determination of Total Selenomethionine (SeMet)
3.10. Determination of Protein Selenocysteine (SeCys) and Selenomethionine (SeMet)
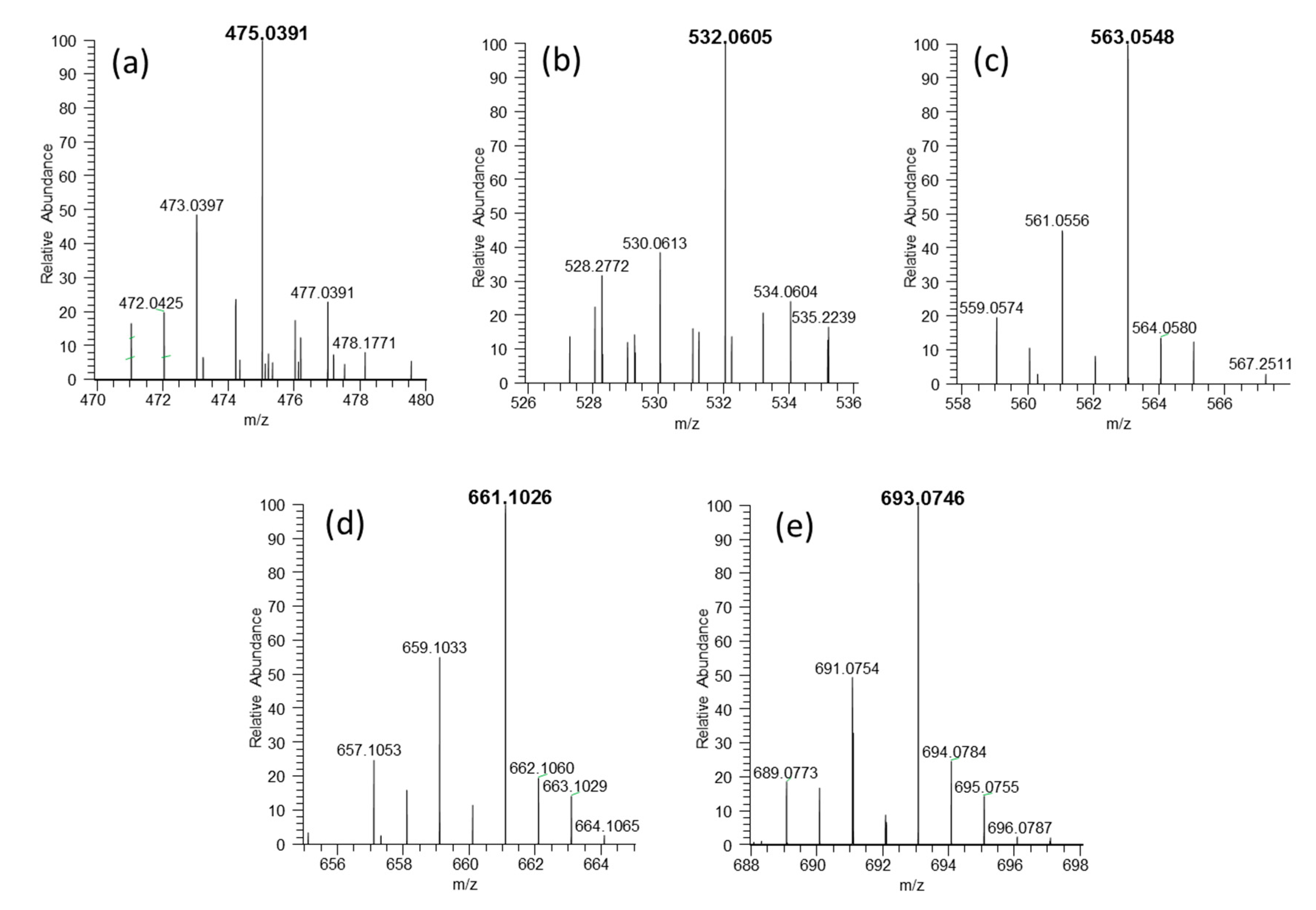
3.11. Identification of Se-Compounds in Water Fraction by ESI MS
3.12. Sample Preparation for SeNP
- Two hundred milligrams of a Se-rich yeast sample were suspended in 5 mL of water, bath sonicated for 1 h in A Branson B2510 ultrasonic bath (Branson, Danbury, CT, USA), and centrifuged at 4500× g for 10 min in a 5804R centrifuge (Eppendorf, Hamburg, Germany).
- The pellet was resuspended with a solution of 5 mL of 4% (m/v) Driselase (Sigma Aldrich, Saint-Quentin Fallavier, France) in 30 mM Tris (Sigma Aldrich, Saint-Quentin Fallavier, France) at pH 7.5, incubated at 25 °C for 17 h, and centrifuged at 4500× g for 10 min.
- The pellet was resuspended with a solution of 5 mL of 4 mg/L protease (Sigma Aldrich, Saint-Quentin Fallavier, France) in 30 mM Tris at pH 7.5, incubated at 37° C for 17 h, and centrifuged at 4500× g for 10 min.
- The pellet was resuspended with a solution of 5 mL of 4% (m/v) sodium dodecyl sulphate (SDS, Sigma Aldrich, Saint-Quentin Fallavier, France), bath sonicated for 1 h, and centrifuged at 4500× g for 10 min.
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hatfield, D.L.; Schweizer, U.; Tsuji, P.A.; Gladyshev, V.N. (Eds.) Selenium: Its Molecular Biology and Role in Human Health, 4th ed.; Springer: New York, NY, USA, 2016; ISBN 978-3-319-41283-2. [Google Scholar]
- Kieliszek, M.; Blazejak, S. Selenium: Significance, and outlook for supplementation. Nutrition 2013, 29, 713–718. [Google Scholar] [CrossRef] [PubMed]
- Arthur, J.R.; McKenzie, R.C.; Beckett, G.J. Selenium in the immune system. J. Nutr. 2003, 133, 1457S–1459S. [Google Scholar] [CrossRef]
- Benstoem, C.; Goetzenich, A.; Kraemer, S.; Benstoem, C.; Goetzenich, A.; Kraemer, S.; Borosch, S.; Manzanares, W.; Hardy, G.; Stoppe, C. Selenium and its supplementation in cardiovascular disease—What do we know? Nutrients 2015, 7, 3094–3118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kieliszek, M.; Lipinski, B. Pathophysiological significance of protein hydrophobic interactions: An emerging hypothesis. Med. Hypotheses 2018, 110, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Nkengfack, G.; Englert, H.; Haddadi, M. Selenium and Immunity. In Nutrition and Immunity; Mahmoudi, M., Rezaei, N., Eds.; Springer: Cham, Switzerland, 2019; pp. 159–165. [Google Scholar] [CrossRef]
- Fairweather-Tait, S.J.; Collings, R.; Hurst, R. Selenium bioavailability: Current knowledge and future research requirements Am. J. Clin. Nutr. 2010, 91, 1484S–1491S. [Google Scholar] [CrossRef] [Green Version]
- Pilon-Smits, E.A. On the ecology of selenium accumulation in plants. Plants 2019, 8, 197. [Google Scholar] [CrossRef] [Green Version]
- Fairweather-Tait, S.J.; Bao, Y.; Broadley, M.R.; Collings, R.; Ford, D.; Hesketh, J.E.; Hurst, R. Selenium in human health and disease. Antioxid. Redox Signal. 2011, 14, 1337–1383. [Google Scholar] [CrossRef]
- Schrauzer, G.N. Selenium yeast: Composition, quality, analysis, and safety. Pure Appl Chem. 2006, 78, 105–109. [Google Scholar] [CrossRef] [Green Version]
- Kieliszek, M. Selenium–fascinating microelement, properties and sources in food. Molecules 2019, 24, 1298. [Google Scholar] [CrossRef] [Green Version]
- Bierla, K.; Szpunar, J.; Yiannikouris, A.; Lobinski, R. Comprehensive speciation of selenium in selenium-rich yeast. TRAC-Trend Anal. Chem. 2012, 41, 122–132. [Google Scholar] [CrossRef]
- Buerth, C.; Tielker, D.; Ernst, J.F. Candida utilis and Cyberlindnera (Pichia) jadinii: Yeast relatives with expanding applications. Appl. Microbiol. Biotechnol. 2016, 100, 6981–6990. [Google Scholar] [CrossRef] [PubMed]
- Bierla, K.; Suzuki, N.; Ogra, Y.; Szpunar, J.; Łobiński, R. Identification and determination of selenohomolanthionine–The major selenium compound in Torula yeast. Food Chem. 2017, 237, 1196–1201. [Google Scholar] [CrossRef] [PubMed]
- Kieliszek, M.; Błażejak, S.; Płaczek, M. Spectrophotometric evaluation of selenium binding by Saccharomyces cerevisiae ATCC MYA-2200 and Candida utilis ATCC 9950 yeast. J. Trace Elem. Med. Biol. 2016, 35, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Kieliszek, M.; Błażejak, S.; Bzducha-Wróbel, A.; Kurcz, A. Effects of selenium on morphological changes in Candida utilis ATCC 9950 yeast cells. Biol. Trace Elem. Res. 2016, 169, 387–393. [Google Scholar] [CrossRef] [Green Version]
- Kieliszek, M.; Błażejak, S.; Bzducha-Wróbel, A.; Kot, A.M. Effect of selenium on lipid and amino acid metabolism in yeast cells. Biol. Trace Elem. Res. 2019, 187, 316–327. [Google Scholar] [CrossRef] [Green Version]
- Moon, J.E.; Heo, W.; Lee, S.H.; Lee, S.H.; Lee, H.G.; Lee, J.H.; Kim, Y.J. Trehalose protects the probiotic yeast Saccharomyces boulardii against oxidative stress-Induced cell death. J. Microbiol. Biotechnol. 2020, 30, 54–61. [Google Scholar] [CrossRef]
- Heinisch, J.J.; Rodicio, R. Stress responses in wine yeast. In Biology of Microorganisms on Grapes, in Must and in Wine; Springer: Cham, Switzerland, 2017; pp. 377–395. [Google Scholar] [CrossRef]
- Türkel, S. Comparative analysis of glycogen and trehalose accumulation in methylotrophic and nonmethylotrophic yeasts. Microbiology 2006, 75, 639–643. [Google Scholar] [CrossRef]
- Arnaudguilhem, C.; Bierla, K.; Ouerdane, L.; Preud’homme, H.; Yiannikouris, A.; Łobiński, R. Selenium metabolomics in yeast using complementary reversed-phase/ hydrophilic ion interaction (HILIC) liquid chromatography-electro-spray hybrid quadrupole trap/Orbitrap mass spectrometry. Anal. Chim. Acta 2012, 757, 26–38. [Google Scholar] [CrossRef]
- Kieliszek, M.; Błażejak, S.; Gientka, I.; Bzducha-Wróbel, A. Accumulation and metabolism of selenium by yeast cells. Appl. Microbiol. Biotechnol. 2015, 99, 5373–5382. [Google Scholar] [CrossRef] [Green Version]
- Fujs, Š.; Gazdag, Z.; Poljšak, B.; Stibilj, V.; Milačič, R.; Pesti, M.; Raspor, M.P.; Batič, M. The oxidative stress response of the yeast Candida intermedia to copper, zinc, and selenium exposure. J. Basic Microbiol. 2005, 45, 125–135. [Google Scholar] [CrossRef]
- Peyroche, G.; Saveanu, C.; Dauplais, M.; Lazard, M.; Beuneu, F.; Decourty, L.; Malabat, C.; Jacquier, A.; Blanquet, S.; Plateau, P. Sodium selenide toxicity is mediated by O2-dependent DNA breaks. PLoS ONE 2012, 7, e36343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kieliszek, M.; Błażejak, S.; Bzducha-Wróbel, A.; Kot, A.M. Effect of selenium on growth and antioxidative system of yeast cells. Mol. Biol. Rep. 2019, 46, 1797–1808. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Yao, X.; Wang, C.; Wang, D.; Wei, G. Transcriptome analysis reveals the mechanism underlying improved glutathione biosynthesis and secretion in Candida utilis during selenium enrichment. J. Biotechnol. 2019, 304, 89–96. [Google Scholar] [CrossRef]
- Asghari-Paskiabi, F.; Imani, M.; Rafii-Tabar, H.; Razzaghi-Abyaneh, M. Physicochemical properties, antifungal activity and cytotoxicity of selenium sulfide nanoparticles green synthesized by Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 2019, 516, 1078–1084. [Google Scholar] [CrossRef]
- Estrada, A.M.; Olivares, L.G.G.; López, E.C.; Serrano, G.R. SelA and SelD genes involved in selenium absorption metabolism in lactic acid bacteria isolated from Mexican cheeses. Int. Dairy J. 2020, 103, 104629. [Google Scholar] [CrossRef]
- Miletić, D.; Turło, J.; Podsadni, P.; Pantić, M.; Nedović, V.; Lević, S.; Nikšić, M. Selenium-enriched Coriolus versicolor mushroom biomass: Potential novel food supplement with improved selenium bioavailability. J. Sci. Food Agric. 2019, 99, 5122–5130. [Google Scholar] [CrossRef]
- Lusa, M.; Help, H.; Honkanen, A.P.; Knuutinen, J.; Parkkonen, J.; Kalasová, D.; Bomberg, M. The reduction of selenium (IV) by boreal Pseudomonas sp. strain T5-6-I–Effects on selenium (IV) uptake in Brassica oleracea. Environ. Res. 2019, 177, 108642. [Google Scholar] [CrossRef] [Green Version]
- Assunção, M.; Martins, L.L.; Mourato, M.P.; Baleiras-Couto, M.M. Effect of selenium on growth and antioxidant enzyme activities of wine related yeasts. World J. Microbiol. Biotechnol. 2015, 31, 1899–1906. [Google Scholar] [CrossRef]
- Wang, Q.; Zuo, Y.; Chen, T.; Yang, Y. Effects of selenium on antioxidant enzymes and photosynthesis in the edible seaweed Gracilaria lemaneiformis. J. Appl. Phycol. 2019, 31, 1303–1310. [Google Scholar] [CrossRef]
- Saharan, R.K.; Sharma, S.C. Effects of glutathione modulation on oxidative stress and enzymatic antioxidant defence in yeast Pachysolen tannophilus. Curr. Microbiol. 2011, 62, 944–949. [Google Scholar] [CrossRef]
- Gao, J.; Yuan, W.; Li, Y.; Bai, F.; Jiang, Y. Synergistic effect of thioredoxin and its reductase from Kluyveromyces marxianus on enhanced tolerance to multiple lignocellulose-derived inhibitors. Microb. Cell Fact. 2017, 16, 181. [Google Scholar] [CrossRef] [Green Version]
- Maqbool, I.; Ponniresan, V.; Govindasamy, K.; Prasad, N.R. Understanding the survival mechanisms of Deinococcus radiodurans against oxidative stress by targeting thioredoxin reductase redox system. Arch. Microbiol. 2019. [Google Scholar] [CrossRef]
- Karamanou, D.A.; Aliferis, K.A. The yeast (Saccharomyces cerevisiae) YCF1 vacuole transporter: Evidence on its implication into the yeast resistance to flusilazole as revealed by GC/EI/MS metabolomics. Pestic. Biochem. Physiol. 2020, 104475. [Google Scholar] [CrossRef]
- De Anchieta Câmara, A., Jr.; Maréchal, P.A.; Tourdot-Maréchal, R.; Husson, F. Dehydration stress responses of yeasts Torulaspora delbrueckii, Metschnikowia pulcherrima and Lachancea thermotolerans: Effects of glutathione and trehalose biosynthesis. Food Microbiol. 2019, 79, 137–146. [Google Scholar] [CrossRef]
- Sharma, S.C.; Anand, M.S. Role of selenium supplementation and heat stress on trehalose and glutathione content in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2006, 133, 1–7. [Google Scholar] [CrossRef]
- Li, H.; Wang, H.L.; Du, J.; Du, G.; Zhan, J.C.; Huang, W.D. Trehalose protects wine yeast against oxidation under thermal stress. World J. Microb. Biot. 2010, 26, 969–976. [Google Scholar] [CrossRef]
- Wilson, W.A.; Roach, P.J.; Montero, M.; Baroja-Fernández, E.; Muñoz, F.J.; Eydallin, G.; Viale, A.M.; Pozueta-Romero, J. Regulation of glycogen metabolism in yeast and bacteria. FEMS Microbiol. Rev. 2010, 34, 952–985. [Google Scholar] [CrossRef] [Green Version]
- Rúa, J.; de Cima, S.; del Valle, P.; Gutiérrez-Larraínzar, M.; Busto, F.; de Arriaga, D. Glycogen and trehalose mobilization by acetic acid in Phycomyces blakesleeanus: Dependence on the anion form. Res. Microbiol. 2008, 159, 200–206. [Google Scholar] [CrossRef]
- Cano, M.; Holland, S.C.; Artier, J.; Burnap, R.L.; Ghirardi, M.; Morgan, J.A.; Yu, J. Glycogen synthesis and metabolite overflow contribute to energy balancing in cyanobacteria. Cell Rep. 2018, 23, 667–672. [Google Scholar] [CrossRef]
- Possik, E.; Pause, A. Glycogen: A must have storage to survive stressful emergencies. Worm 2016, 5, e1005520. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.J.; Chi, Z.; Jiang, H.; Liu, G.L.; Hu, Z.; Chi, Z.M. Cell wall integrity is required for pullulan biosynthesis and glycogen accumulation in Aureobasidium melanogenum P16. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 1516–1526. [Google Scholar] [CrossRef]
- Miletić, D.; Pantić, M.; Sknepnek, A.; Vasiljević, I.; Lazović, M.; Nikšić, M. Influence of selenium yeast on the growth, selenium uptake and mineral composition of Coriolus versicolor mushroom. J. Basic Microbiol. 2020, 60, 331–340. [Google Scholar] [CrossRef]
- Ullah, A.; Sun, B.; Wang, F.; Yin, X.; Xu, B.; Nawazish Ali, N.; Mirani, Z.A.; Mehmood, A.; Naveed, M. Isolation of selenium-resistant under enrichment conditions for selected probiotic Bacillus subtilis (BSN313). J. Food Biochem. 2020, 44, e13227. [Google Scholar] [CrossRef]
- Egressy-Molnár, O.; Ouerdane, L.; Győrfi, J.; Dernovics, M. Analogy in selenium enrichment and selenium speciation between selenized yeast Saccharomyces cerevisiae and Hericium erinaceus (lion’s mane mushroom). LWT-Food Sci. Technol. 2016, 68, 306–312. [Google Scholar] [CrossRef]
- Zhang, G.C.; Wang, D.H.; Wang, D.H.; Wei, G.Y. The mechanism of improved intracellular organic selenium and glutathione contents in selenium-enriched Candida utilis by acid stress. Appl. Microbiol. Biotechnol. 2017, 101, 2131–2141. [Google Scholar] [CrossRef]
- Kieliszek, M.; Kot, A.M.; Bzducha-Wróbel, A.; Błażejak, S.; Gientka, I.; Kurcz, A. Biotechnological use of Candida yeasts in the food industry: A review. Fungal Biol. Rev. 2017, 31, 185–198. [Google Scholar] [CrossRef]
- Rosen, B.P.; Liu, Z. Transport pathways for arsenic and selenium: A minireview. Environ. Int. 2009, 35, 512–515. [Google Scholar] [CrossRef] [Green Version]
- McDermott, J.R.; Rosen, B.P.; Liu, Z. Jen1p: A high affinity selenite transporter in yeast. Mol. Biol. Cell. 2010, 21, 3934–3941. [Google Scholar] [CrossRef] [Green Version]
- Lazard, M.; Blanquet, S.; Fisicaro, P.; Labarraque, G.; Plateau, P. Uptake of selenite by Saccharomyces cerevisiae involves the high and low affinity orthophosphate transporters. J. Biol. Chem. 2010, 42, 32029–32037. [Google Scholar] [CrossRef] [Green Version]
- Byrne, S.L.; Durandeau, K.; Nagy, I.; Barth, S. Identification of ABC transporters from Lolium perenne L. that are regulated by toxic levels of selenium. Planta 2010, 231, 901–911. [Google Scholar] [CrossRef]
- Bierla, K.; Godin, S.; Lobinski, R.; Szpunar, J. Advances in electrospray mass spectrometry for the selenium speciation: Focus on Se-rich yeast. TRAC-Trend Anal. Chem. 2018, 104, 87–94. [Google Scholar] [CrossRef]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (EFSA FEEDAP, Panel); Bampidis, V.; Azimonti, G.; Bastos, M.D.L.; Christensen, H.; Dusemund, B.; Kouba, M.; Durjava, M.K.; López-Alonso, M.; Puente, S.L.; et al. Assessment of the application for renewal of authorisation of selenomethionine produced by Saccharomyces cerevisiae NCYC R397 for all animal species. EFSA J. 2019, 17, e05539. [Google Scholar] [CrossRef]
- Mester, Z.; Willie, S.; Yang, L.; Sturgeon, R.; Caruso, J.A.; Fernandez, M.L.; Fedor, P.; Goldschmidt, R.J.; Goenaga-Infante, H.; Lobinski, R.; et al. Certification of a new selenized yeast reference material (SELM-1) for methionine, selenomethionine and total selenium content and its use in an intercomparison exercise for quantifying these analytes. Anal. Bioanal. Chem. 2006, 385, 168–180. [Google Scholar] [CrossRef]
- Kieliszek, M.; Błażejak, S. Current knowledge on the importance of selenium in food for living organisms: A review. Molecules 2016, 21, 609. [Google Scholar] [CrossRef] [Green Version]
- Schild, F.; Kieffer-Jaquinod, S.; Palencia, A.; Cobessi, D.; Sarret, G.; Zubieta, C.; Jourdain, A.; Dumas, R.; Forge, V.; Testemale, D.; et al. Biochemical and biophysical characterization of the selenium-binding and reducing site in Arabidopsis thaliana homologue to mammals selenium-binding protein 1. J. Biol. Chem. 2014, 289, 31765–31776. [Google Scholar] [CrossRef] [Green Version]
- Ward, P.; Chadha, M.; Connolly, C.; Stalcup, A.; Murphy, R. A comparative assessment of water-soluble selenium metabolites in commercial selenised yeast supplements by liquid chromatography-electrospray ionisation QTOF-MS. Int. J. Mass Spectrom. 2019, 439, 42–52. [Google Scholar] [CrossRef]
- Preud’homme, H.; Far, J.; Gil-Casal, S.; Lobinski, R. Large-scale identification of selenium metabolites by online size-exclusion-reversed phase liquid chromatography with combined inductively coupled plasma (ICP-MS) and electrospray ionization linear trap-Orbitrap mass spectrometry (ESI-MS n). Metallomics 2012, 4, 422–432. [Google Scholar] [CrossRef]
- Kieliszek, M.; Błażejak, S. Speciation Analysis of selenium in Candida utilis yeast cells using HPLC-ICP-MS and UHPLC-ESI-Orbitrap MS Techniques. Appl. Sci. 2018, 8, 2050. [Google Scholar] [CrossRef] [Green Version]
- Lian, S.; Diko, C.S.; Yan, Y.; Li, Z.; Zhang, H.; Ma, Q.; Qu, Y. Characterization of biogenic selenium nanoparticles derived from cell-free extracts of a novel yeast Magnusiomyces ingens. 3 Biotech 2019, 9, 221. [Google Scholar] [CrossRef]
- Faramarzi, S.; Anzabi, Y.; Jafarizadeh-Malmiri, H. Nanobiotechnology approach in intracellular selenium nanoparticle synthesis using Saccharomyces cerevisiae—fabrication and characterization. Arch. Microbiol. 2020. [Google Scholar] [CrossRef]
- Montoro Bustos, A.R.; Purushotham, K.P.; Possolo, A.; Farkas, N.; Vladár, A.E.; Murphy, K.E.; Winchester, M.R. Validation of single particle ICP-MS for routine measurements of nanoparticle size and number size distribution. Anal. Chem. 2018, 90, 14376–14386. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Lamana, J.; Abad-Álvaro, I.; Bierla, K.; Laborda, F.; Szpunar, J.; Lobinski, R. Detection and characterization of biogenic selenium nanoparticles in selenium-rich yeast by single particle ICPMS. J. Anal. At. Spectrom. 2018, 33, 452–460. [Google Scholar] [CrossRef]
- Hamza, F.; Vaidya, A.; Apte, M.; Kumar, A.R.; Zinjarde, S. Selenium nanoparticle-enriched biomass of Yarrowia lipolytica enhances growth and survival of Artemia salina. Enzyme Microb. Technol. 2017, 106, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Samant, S.; Naik, M.; Parulekar, K.; Charya, L.; Vaigankar, D. Selenium reducing Citrobacter fruendii strain KP6 from Mandovi estuary and its potential application in selenium nanoparticle synthesis. Proc. Natl. Acad. Sci. India B 2018, 88, 747–754. [Google Scholar] [CrossRef]
- Kieliszek, M.; Kot, A.M.; Piwowarek, K.; Błażejak, S. Accumulation of selenium in Candida utilis growing in media of increasing concentration of this element. Appl. Sci. 2020, 10, 1439. [Google Scholar] [CrossRef] [Green Version]
- Kieliszek, M.; Błażejak, S.; Bzducha-Wróbel, A. Influence of selenium content in the culture medium on protein profile of yeast cells Candida utilis ATCC 9950. Oxid. Med. Cell. Longev. 2015, 2015, 659750. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Selenium Content in the Medium | Δtlag (h) | Δtlog (h) | ODmin lag | ODmax log | µmax (h−1) | G (h) | ΔOD |
|---|---|---|---|---|---|---|---|
| 0 mg/L | 2 | 22 | 0.54 | 2.2 | 0.063 | 10.09 | 1.934 |
| 30 mg/L | 6 | 20 | 0.49 | 1.9 | 0.067 | 10.29 | 1.578 |
| Sample | Fraction | Selenium (µg/g) | RDS (%) |
|---|---|---|---|
| control | total | 5.0 ± 0.12 c | 2.4 |
| water soluble | 2.0 ± 0.13 d | 6.4 | |
| Se-enriched | total | 1368 ± 69.36 a | 5.1 |
| water soluble | 113 ± 2.67 b | 2.4 |
| Sample | Selenium Compounds | |||||
|---|---|---|---|---|---|---|
| Selenomethionine (SeMet, µg/g) | RSD (%) | Selenocysteine (SeCys, µg/g) | RSD (%) | Selenium Inorganic (Se IV, µg/g) | RSD (%) | |
| SELM-1 | 1162 ± 45.5 | 3.9 | 102 ± 7.6 | 7.4 | 132 ± 8.5 | 6.5 |
| Candida utilis | 138 ± 4.6 | 3.4 | 279 ± 18.1 | 6.5 | 804 ± 45.4 | 5.6 |
| Nr | Formula | Exp. Mass | Error (µg/g) | Name Compounds | Name |
|---|---|---|---|---|---|
| a | C13H23O8N4SSe+ | 475.0387 | −1.9 | selenoglutathione-cysteine | 188609-44-1 or 117135-55-4 * |
| b | C15H26O9N5SSe+ | 532.0599 | −2.3 | glutathione-selenocysteinylglycine | 1357479-87-8 |
| c | C16H27O11N4SSe+ | 563.0544 | −2.2 | glutathione-2,3-DHP-selenocysteine | 1006377-09-8 |
| d | C20H33O12N6SSe+ | 661.1022 | −2.3 | selenoglutathione-glutathione | 161973-63-3 |
| e | C20H33O12N6S2Se+ | 693.0746 | −1.6 | selenodiglutathione | 1052197-78-0 |
| Column | Eluent | Gradient Elution | Temperature (°C) | Flow Rate (mL/min) | Sample Volume (µL) | Detection |
|---|---|---|---|---|---|---|
| C8 Alltima 4.6 × 250 mm | A: 0.1% HFBA * in water B: 0.1% HFBA in MeOH ** | 0–15 min 3% B 15–18 min 3-40% B 18–21 min 40% B 21–23 min 40-3 % B 23–30 min 3% B | 10 | 0.9 | 10 | ICP MS |
| C18 Zorbax Eclipse XBD 4.6 × 150 mm | A: 0.1% FA *** in water B: 0.1% FA in MeOH | 0–2.5 min 3% B 2.5–15 min 3-50% B 15–20 min 50% B 20–20.5 min 40-3 % B 20.5–30 min 3% B | 40 | 1.0 | 10 | ESI MS |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kieliszek, M.; Bierla, K.; Jiménez-Lamana, J.; Kot, A.M.; Alcántara-Durán, J.; Piwowarek, K.; Błażejak, S.; Szpunar, J. Metabolic Response of the Yeast Candida utilis During Enrichment in Selenium. Int. J. Mol. Sci. 2020, 21, 5287. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155287
Kieliszek M, Bierla K, Jiménez-Lamana J, Kot AM, Alcántara-Durán J, Piwowarek K, Błażejak S, Szpunar J. Metabolic Response of the Yeast Candida utilis During Enrichment in Selenium. International Journal of Molecular Sciences. 2020; 21(15):5287. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155287
Chicago/Turabian StyleKieliszek, Marek, Katarzyna Bierla, Javier Jiménez-Lamana, Anna Maria Kot, Jaime Alcántara-Durán, Kamil Piwowarek, Stanisław Błażejak, and Joanna Szpunar. 2020. "Metabolic Response of the Yeast Candida utilis During Enrichment in Selenium" International Journal of Molecular Sciences 21, no. 15: 5287. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155287