Halophytic Hordeum brevisubulatum HbHAK1 Facilitates Potassium Retention and Contributes to Salt Tolerance

Abstract
:1. Introduction
2. Results
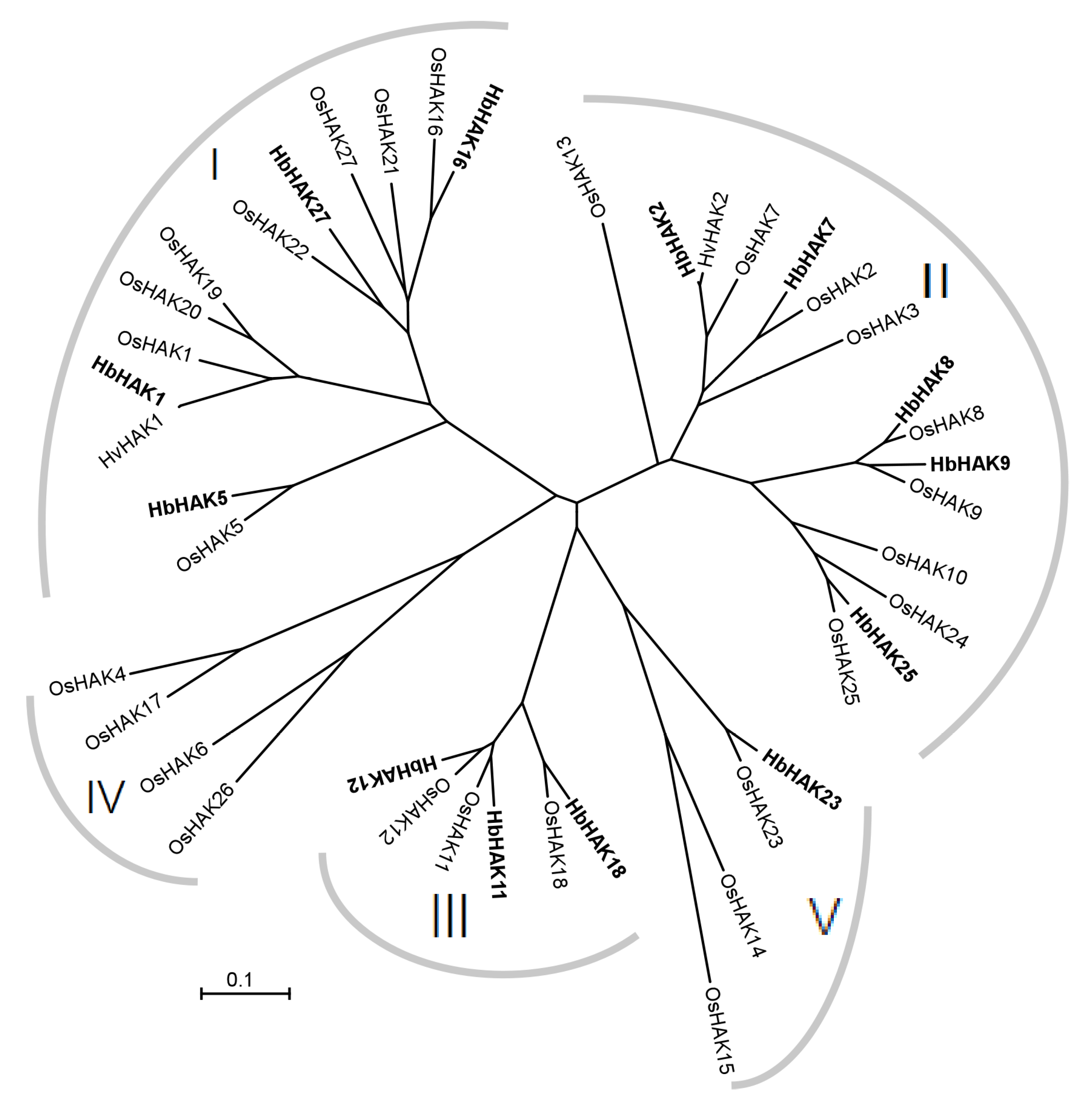
2.1. HAK/KUP/KT Proteins Were Identified According to Transcriptomic Analysis
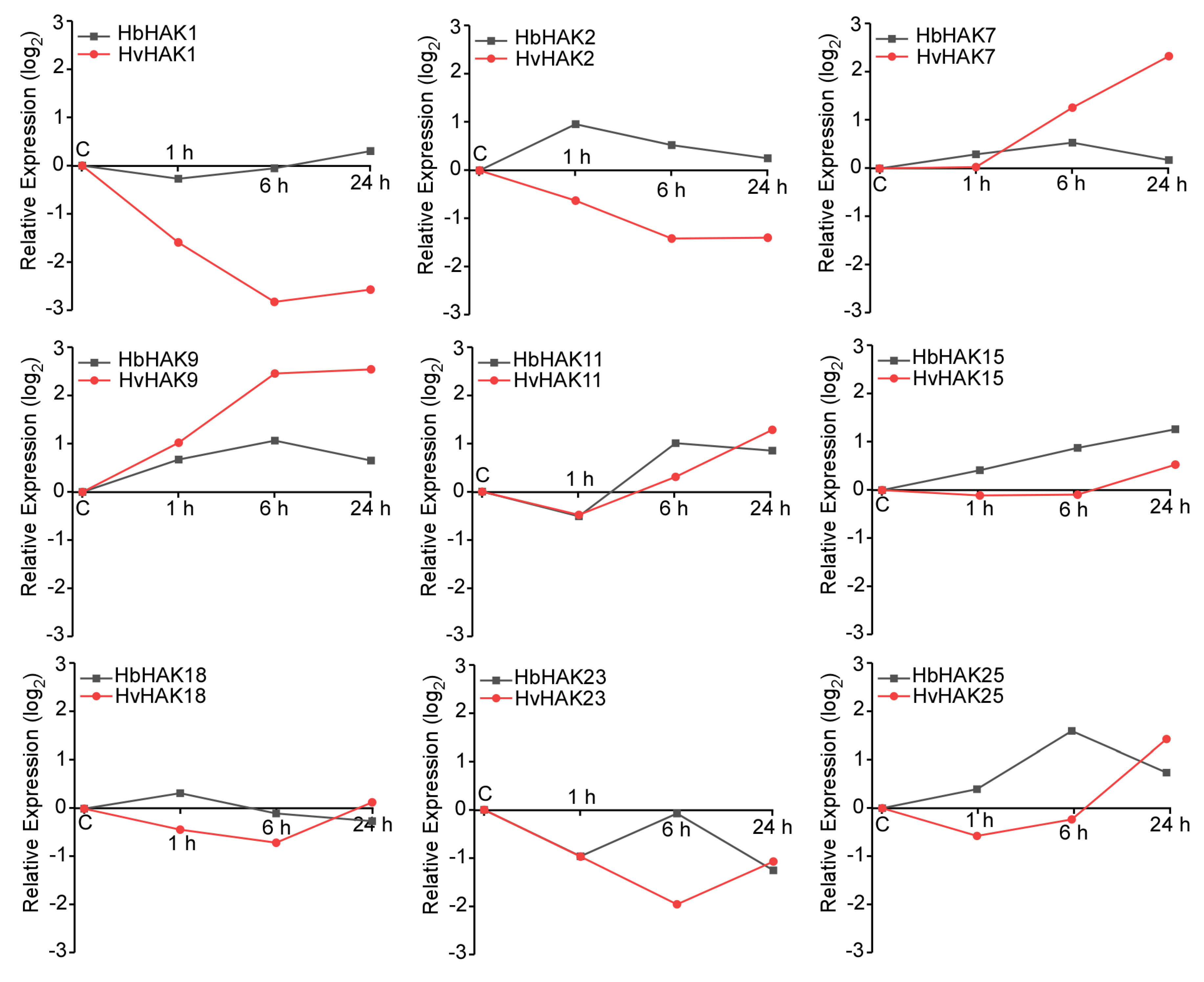
2.2. HbHAK1 Was Upregulated by Salt Stress in Roots with the Differential Expression Patterns
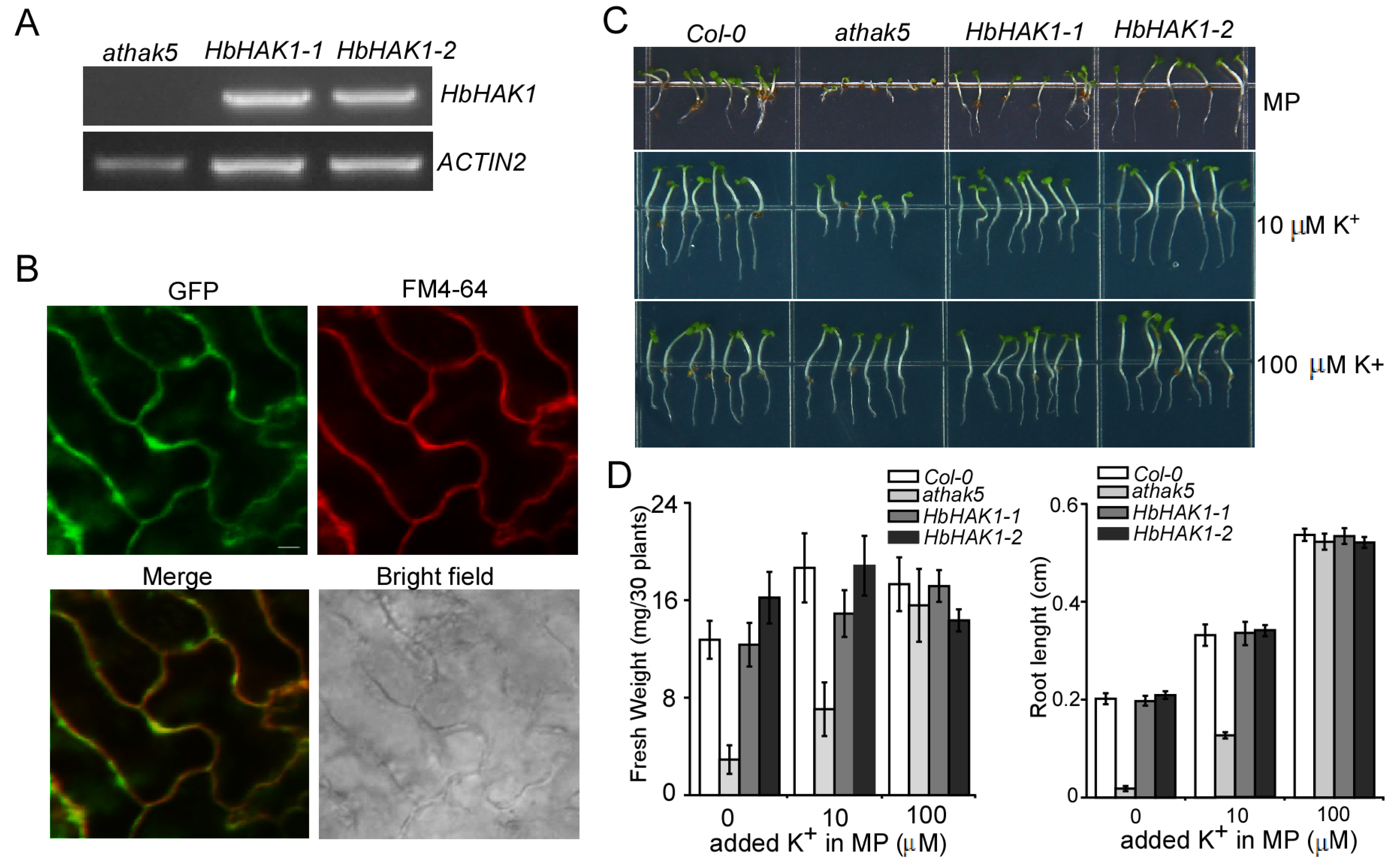
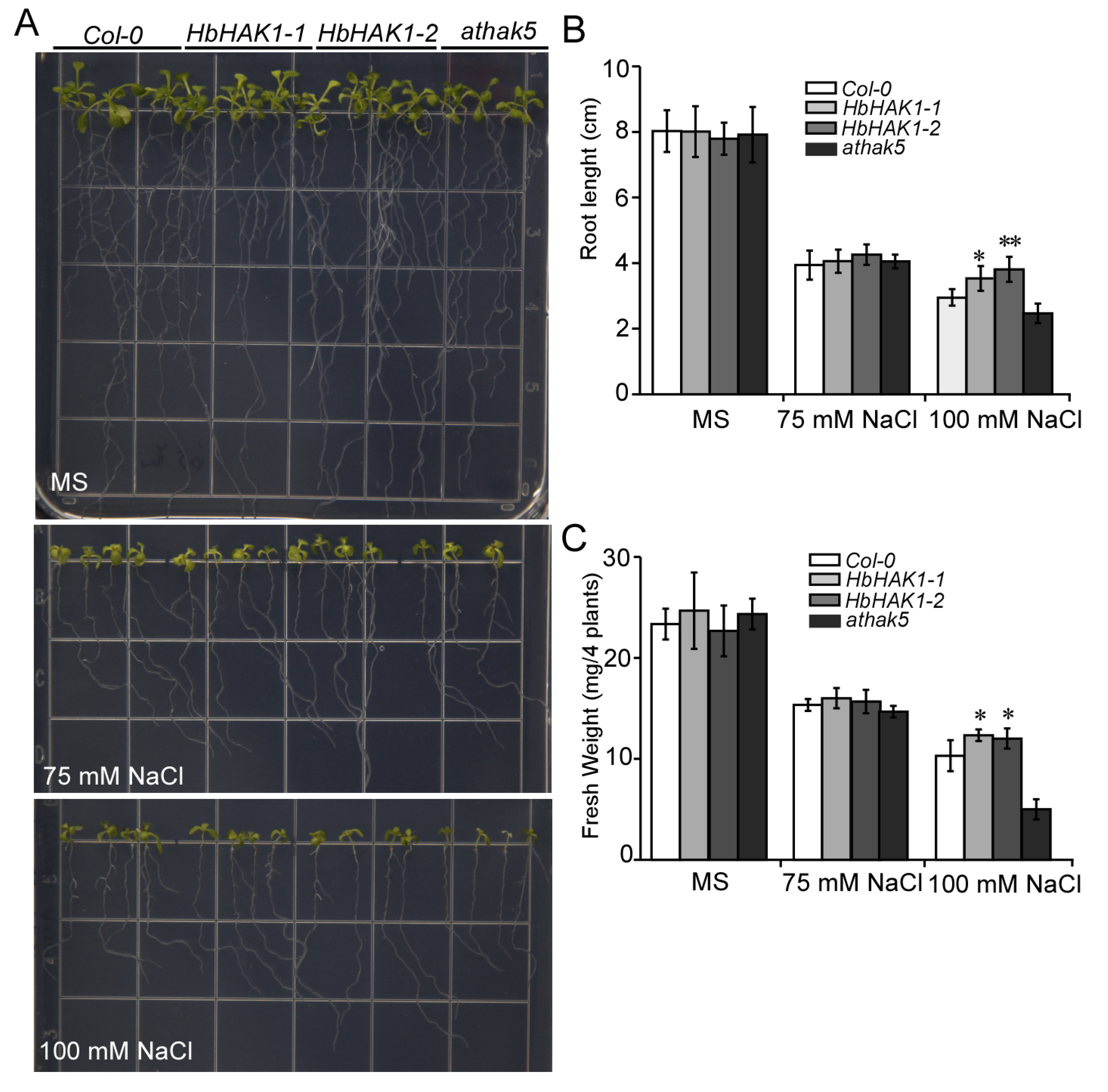
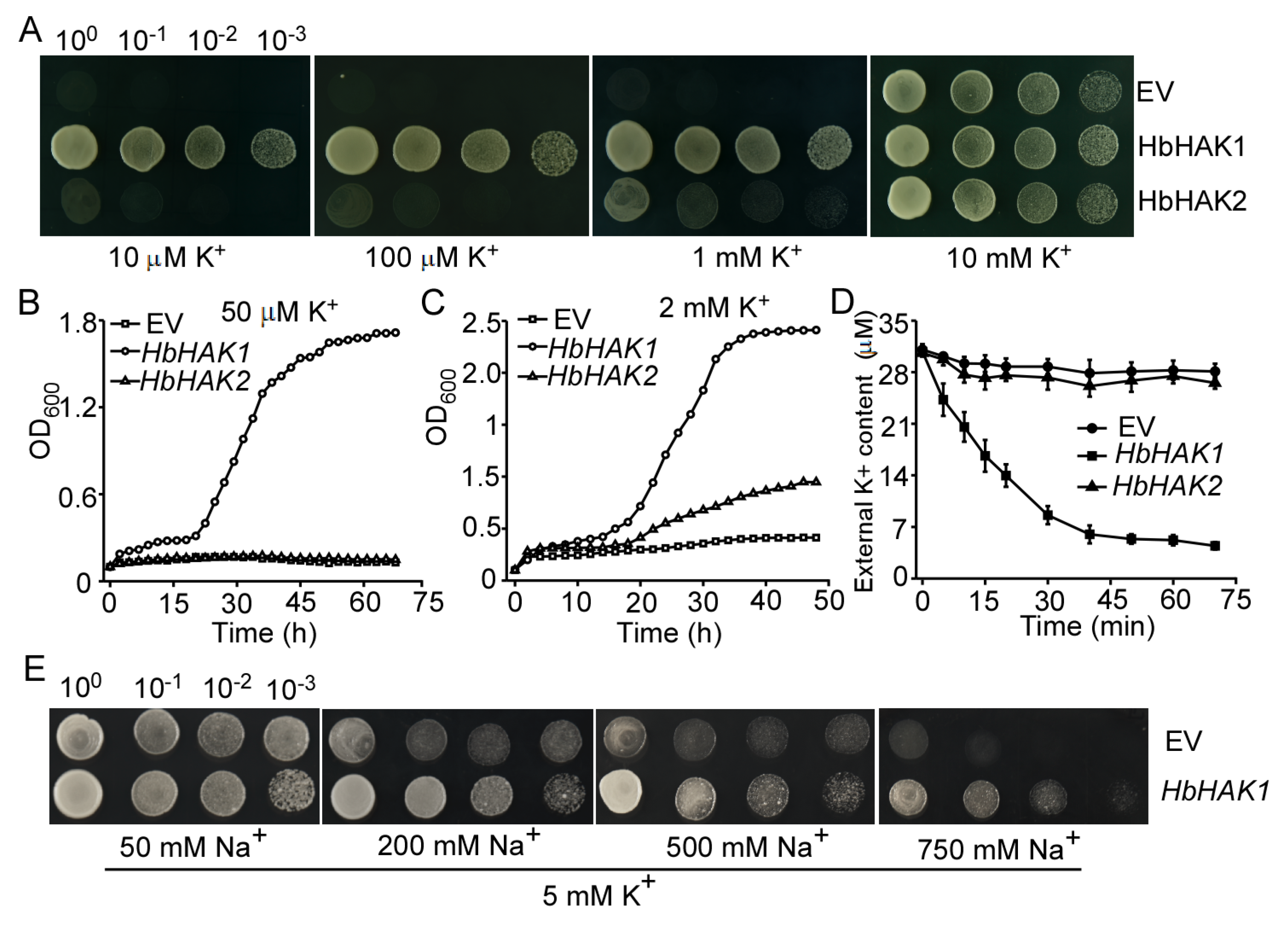
2.3. HbHAK1 Worked as a High-Affinity K+ Transporter on the Plasma Membrane and Involved in Salt Tolerance
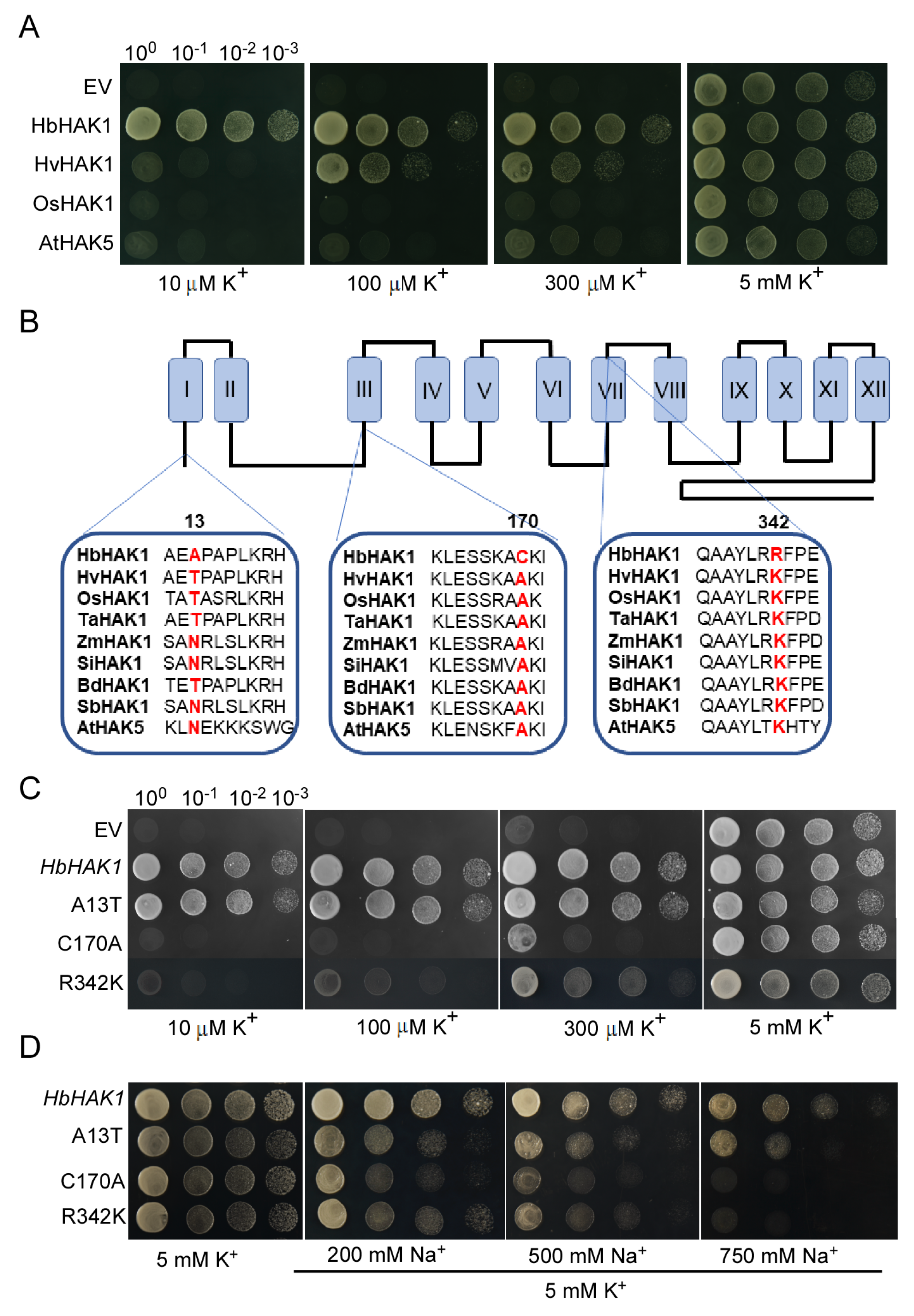
2.4. HbHAK1 Strongly Mediated K+ Uptake Under Low-K+ Conditions or High Na+ Condition
2.5. The Amino Acid Residues C170 and R342 of HbHAK1 Determined the Stronger Transport Activity in Low-K+ Conditions While A13 Under Salt Stress
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Condition
4.2. Isolation of HAK/KUP/KT Family Genes from H. brevisubulatum Transcriptomic Database
4.3. HbHAK1 Expression Transgenic Lines in Arabidopsis athak5
4.4. Subcellular Localization of HbHAK1
4.5. Complementation of HbHAK1 in Cy162 Yeast Strains
4.6. K+ Depletion and Measurement of K+ Contents in Cy162 Yeast Strains
4.7. Site-Directed Mutagenesis of HbHAK1
4.8. Accession Numbers
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Srivastava, A.K.; Shankar, A.; Nalini Chandran, A.K.; Sharma, M.; Jung, K.H.; Suprasanna, P.; Pandey, G.K. Emerging concepts of potassium homeostasis in plants. J. Exp. Bot. 2020, 71, 608–619. [Google Scholar] [CrossRef]
- Shabala, S.; Pottosin, I. Regulation of potassium transport in plants under hostile conditions: Implications for abiotic and biotic stress tolerance. Physiol. Plant 2014, 151, 257–279. [Google Scholar] [CrossRef] [PubMed]
- Shabala, S. Signalling by potassium: Another second messenger to add to the list? J. Exp. Bot. 2017, 68, 4003–4007. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Xu, G.; Alli, A.; Yu, L. Plant HAK/KUP/KT K+ transporters: Function and regulation. Semin. Cell Dev. Biol. 2018, 74, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Nieves-Cordones, M.; Ródenas, R.; Chavanieu, A.; Rivero, R.M.; Martinez, V.; Gaillard, I.; Rubio, F. Uneven HAK/KUP/KT protein diversity among angiosperms: Species distribution and perspectives. Front. Plant Sci. 2016, 7, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gierth, M.; Mäser, P.; Schroeder, J.I. The potassium transporter AtHAK5 functions in K+ deprivation-induced high-affinity K+ uptake and AKT1 K+ channel contribution to K+ uptake kinetics in Arabidopsis roots. Plant Physiol. 2005, 137, 1105–1114. [Google Scholar] [CrossRef] [Green Version]
- Santa-María, G.E.; Rubio, F.; Dubcovsky, J.; Rodríguez-Navarro, A. The HAK1 gene of barley is a member of a large gene family and encodes a high-affinity potassium transporter. Plant Cell 1997, 177, 2281–2289. [Google Scholar]
- Chen, G.; Hu, Q.; Luo, L.; Yang, T.; Zhang, S.; Hu, Y.; Yu, L.; Xu, G. Rice potassium transporter OsHAK1 is essential for maintaining potassium-mediated growth and functions in salt tolerance over low and high potassium concentration ranges. Plant Cell Environ. 2015, 38, 2747–2765. [Google Scholar] [CrossRef]
- Yang, T.; Zhang, S.; Hu, Y.; Wu, F.; Hu, Q.; Chen, G.; Cai, J.; Wu, T.; Moran, N.; Yu, L.; et al. The role of a potassium transporter OsHAK5 in potassium acquisition and transport from roots to shoots in rice at low potassium supply levels. Plant Physiol. 2014, 166, 945–959. [Google Scholar] [CrossRef] [Green Version]
- Alemán, F.; Nieves-Cordones, M.; Martínez, V.; Rubio, F. Differential regulation of the HAK5 genes encoding the high-affinity K+ transporters of Thellungiella halophila and Arabidopsis thaliana. Environ. Exp. Bot. 2009, 65, 263–265. [Google Scholar] [CrossRef]
- Nieves-Cordones, M.; Martínez-Cordero, M.A.; Martínez, V.; Rubio, F. An NH4+ sensitive component dominates high-affinity K+ uptake in tomato plants. Plant Sci. 2007, 172, 273–280. [Google Scholar] [CrossRef]
- Ruiz-Lau, N.; Bojórquez-Quintal, E.; Benito, B.; Echevarría-Machado, I.; Sánchez-Cach, L.A.; Medina-Lara, M.F.; Martínez-Estévez, M. Molecular cloning and functional analysis of a Na+-insensitive K+ transporter of Capsicum chinense Jacq. Front. Plant Sci. 2016, 7, 1980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shabala, S.; Demidchik, V.; Shabala, L.; Cuin, T.; Smith, S.J.; Miller, A.J.; Davies, J.M.; Newman, I.A. Extracellular Ca2+ ameliorates NaCl-induced K+ loss from Arabidopsis root and leaf cells by controlling plasma membrane K+-permeable channels. Plant Physiol. 2006, 1, 1653–1665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carden, D.E.; Walker, D.J.; Flowers, T.J.; Miller, A.J. Single-cell measurements of the contributions of cytosolic Na+ and K+ to Salt Tolerance. Plant Physiol. 2003, 131, 676–683. [Google Scholar] [CrossRef] [Green Version]
- Adams, E.; Abdollahi, P.; Shin, R. Cesium inhibits plant growth through jasmonate signaling in Arabidopsis thaliana. Int. J. Mol. Sci. 2013, 14, 4545–4559. [Google Scholar] [CrossRef] [Green Version]
- Shabala, S.; Cuin, T. Potassium transport and plant salt tolerance. Physiol. Plant 2008, 133, 651–669. [Google Scholar] [CrossRef]
- Rubio, F.; Nieves-Cordones, M.; Horie, T.; Shabala, S. Doing ‘business as usual’ comes with a cost: Evaluating energy cost of maintaining plant intracellular K+ homeostasis under saline conditions. New Phytol. 2020, 225, 1097–1104. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Liu, C.; Gao, Z.; Zhang, Y.; Jiang, H.; Zhu, L.; Ren, D.; Yu, L.; Xu, G.; Qian, Q. OsHAK1, a High-Affinity potassium transporter, positively regulates responses to drought stress in rice. Front. Plant Sci. 2017, 8, 1885. [Google Scholar] [CrossRef]
- Shen, Y.; Shen, L.; Shen, Z.; Jing, W.; Ge, H.; Zhao, J. The potassium transporter OsHAK21 functions in the maintenance of ion homeostasis and tolerance to salt stress in rice. Plant Cell Environ. 2015, 38, 2766–2779. [Google Scholar] [CrossRef]
- Zhang, M.; Liang, X.; Wang, L.; Cao, Y.; Song, W.; Shi, J.; Lai, J.; Jiang, C. A HAK family Na+ transporter confers natural variation of salt tolerance in maize. Nat. Plants 2019, 5, 1297–1308. [Google Scholar] [CrossRef]
- Fita, A.; Rodríguez-Burruezo, A.; Boscaiu, M.; Prohens, J.; Vicente, O. Breeding and domesticating crops adapted to drought and salinity: A new paradigm for increasing food production. Front. Plant Sci. 2015, 6, 978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mano, Y.; Takeda, K. Genetic resources of salt tolerance in wild Hordeum species. Euphytica 1998, 103, 137–141. [Google Scholar] [CrossRef]
- Brassac, J.; Blattner, F.R. Species-Level Phylogeny and Polyploid Relationships in Hordeum (Poaceae) Inferred by Next-Generation Sequencing and In Silico Cloning of Multiple Nuclear Loci. Syst. Biol. 2015, 64, 792–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.F.; Zhang, J.W.; Wu, G.Y.; Wang, H.Z.; Chen, Y.J.; Wei, J.H. HbCIPK2, a novel CBL-interacting protein kinase from halophyte Hordeum brevisubulatum, confers salt and osmotic stress tolerance. Plant Cell Environ. 2012, 35, 1582–1600. [Google Scholar] [CrossRef]
- Wang, C.M.; Xia, Z.R.; Wu, G.Q.; Yuan, H.J.; Wang, X.R.; Li, J.H.; Tian, F.P.; Zhang, Q.; Zhu, X.Q.; He, J.J.; et al. The coordinated regulation of Na+ and K+ in Hordeum brevisubulatum responding to time of salt stress. Plant Sci. 2016, 252, 358–366. [Google Scholar] [CrossRef]
- Zhang, H.W.; Feng, H.; Zhang, J.W.; Ge, R.C.; Zhang, L.L.; Wang, Y.X.; Li, L.G.; Wei, J.H.; Li, R.F. Emerging crosstalk between two signaling pathways coordinates K+ and Na+ homeostasis in the halophyte Hordeum brevisubulatum. J. Exp. Bot. 2020. [Google Scholar] [CrossRef] [PubMed]
- Rubio, F.; Nieves-Cordones, M.; Alemán, F.; Martínez, V. Relative contribution of AtHAK5 and AtAKT1 to K+ uptake in the high-affinity range of concentrations. Physiol. Plant 2008, 134, 598–608. [Google Scholar] [CrossRef]
- Anderson, J.A.; Huprikar, S.S.; Kochian, L.V.; Lucas, W.J.; Gaber, R.F. Functional expression of a probable Arabidopsis thaliana potassium channel in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1992, 89, 3736–3740. [Google Scholar] [CrossRef] [Green Version]
- White, P. Improving potassium acquisition and utilisation by crop plants. J. Plant Nutr. Soil Sci. 2013, 176, 305–316. [Google Scholar] [CrossRef]
- Sentenac, H.; Bonneaud, N.; Minet, M.; Lacroute, F.; Salmon, J.M.; Gaymard, F.; Grignon, C. Cloning and expression in yeast of a plant potassium-ion transport-system. Science 1992, 256, 663–665. [Google Scholar] [CrossRef]
- Nieves-Cordones, M.; Alemán, F.; Martínez, V.; Rubio, F. The Arabidopsis thaliana HAK5 K+ transporter is required for plant growth and K+ acquisition from low K+ solutions under saline conditions. Mol. Plant 2010, 3, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.L.; Wang, Y.X.; Zhang, Q.K.; Jiang, Y.; Zhang, H.W.; Li, R.F. Overexpression of HbMBF1a, encoding multiprotein bridging factor 1 from the halophyte Hordeum brevisubulatum, confers salinity tolerance and ABA insensitivity to transgenic Arabidopsis thaliana. Plant Mol. Biol. 2020, 102, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, T.; Yamane, S.; Yamaguchi, M.; Kato, K.; Shinmyo, A.; Tsunemitsu, Y.; Iwasaki, K.; Ueno, D.; Demura, T. Characterization of rice KT/HAK/KUP potassium transporters and K+ uptake by HAK1 from Oryza sativa. Plant Biotechnol. 2018, 35, 101–111. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Cordero, M.A.; Martínez, V.; Rubio, F. Cloning and functional characterization of the high-affinity K+ transporter HAK1 of pepper. Plant Mol. Biol. 2004, 56, 413–421. [Google Scholar] [CrossRef]
- Mackinnon, R. Potassium channels. FEBS Lett. 2003, 555, 62–65. [Google Scholar] [CrossRef] [Green Version]
- Dreyer, I.; Uozumi, N. Potassium channels in plant cells. FEBS J. 2011, 278, 4293–4303. [Google Scholar] [CrossRef]
- Benito, B.; Haro, R.; Amtmann, A.; Cuin, T.A.; Dreyer, I. The twins K+ and Na+ in plants. J. Plant Physiol. 2014, 171, 723–731. [Google Scholar] [CrossRef]
- Corratgé-Faillie, C.; Jabnoune, M.; Zimmermann, S.; Véry, A.; Fizames, C.; Sentenac, H. Potassium and sodium transport in Non-Animal cells: The Trk/Ktr/HKT Transporter Family. Cell. Mol. Life Sci. 2010, 67, 2511–2532. [Google Scholar] [CrossRef]
- Mangano, S.; Silberstein, S.; Santa-María, G.E. Point mutations in the Barley HvHAK1 potassium transporter lead to improved K+-nutrition and enhanced resistance to salt stress. FEBS Lett. 2008, 582, 3922–3928. [Google Scholar] [CrossRef] [Green Version]
- Rubio, F.; Santa-Maria, G.E.; Rodrigues-Navarro, A. Cloning of Arabidopsis and barley cDNAs encoding HAK potassium transporters in root and shoot cells. Physiol. Plant 2000, 109, 34–43. [Google Scholar] [CrossRef]
- Alemán, F.; Caballero, F.; Ródenas, R.; Rivero, R.M.; Martínez, V.; Rubio, F. The F130S point mutation in the Arabidopsis high-affinity K+ transporter AtHAK5 increases K+ over Na+ and Cs+ selectivity and confers Na+ and Cs+ tolerance to yeast under heterologous expression. Front. Plant Sci. 2014, 5, 430. [Google Scholar] [CrossRef] [PubMed]
- Garciadeblas, B.; Barrero-Gil, J.; Benito, B.; Rodríguez-Navarro, A. Potassium transport systems in the moss Physcomitrella Patens: pphak1 plants reveal the complexity of potassium uptake. Plant J. 2007, 52, 1080–1093. [Google Scholar] [CrossRef] [PubMed]
- Cutler, S.R.; Ehrhardt, D.W.; Griffitts, J.S.; Somerville, C.R. Random GFP::cDNA fusions enable visualization of subcellular structures in cells of Arabidopsis at a high frequency. Proc. Natl. Acad. Sci. USA 2000, 97, 3718–3723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirsch, R.E.; Lewis, B.D.; Spalding, E.P.; Sussman, M.R. A role for the AKT1 potassium channel in plant nutrition. Science 1998, 280, 918–921. [Google Scholar] [CrossRef]
- Brunelli, J.P.; Pall, M.L. A series of yeast/Escherichia coli lambda expression vectors designed for directional cloning of cDNAs and cre/lox-mediated plasmid excision. Yeast 1993, 9, 1309–1318. [Google Scholar] [CrossRef]
- Bañuelos, M.A.; Garciadeblas, B.; Cubero, B.; Rodríguez-Navarro, A. Inventory and functional characterization of the HAK potassium transporters of rice. Plant Physiol. 2002, 130, 784–795. [Google Scholar] [CrossRef] [Green Version]
- Fraile-Escanciano, A.; Kamisugi, Y.; Cuming, A.C.; Rodríguez-Navarro, A.; Benito, B. The SOS1 transporter of Physcomitrella patens mediates sodium efflux in planta. New Phytol. 2010, 188, 750–761. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Protein Length | TMS a | PL b |
|---|---|---|---|
| HbHAK1 | 776 | 12 | plasma membrane |
| HbHAK2 | 777 | 13 | plasma membrane |
| HbHAK5 | 708 | 11 | plasma membrane |
| HbHAK7 | 784 | 13 | plasma membrane |
| HbHAK8 | 789 | 12 | plasma membrane |
| HbHAK9 | 781 | 11 | plasma membrane |
| HbHAK11 | 792 | 14 | plasma membrane |
| HbHAK12 | 763 | 14 | plasma membrane |
| HbHAK16 | 799 | 11 | plasma membrane |
| HbHAK18 | 785 | 14 | plasma membrane |
| HbHAK23 | 945 | 12 | plasma membrane |
| HbHAK25 | 769 | 13 | plasma membrane |
| HbHAK27 | 874 | 12 | plasma membrane |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Xiao, W.; Yu, W.; Jiang, Y.; Li, R. Halophytic Hordeum brevisubulatum HbHAK1 Facilitates Potassium Retention and Contributes to Salt Tolerance. Int. J. Mol. Sci. 2020, 21, 5292. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155292
Zhang H, Xiao W, Yu W, Jiang Y, Li R. Halophytic Hordeum brevisubulatum HbHAK1 Facilitates Potassium Retention and Contributes to Salt Tolerance. International Journal of Molecular Sciences. 2020; 21(15):5292. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155292
Chicago/Turabian StyleZhang, Haiwen, Wen Xiao, Wenwen Yu, Ying Jiang, and Ruifen Li. 2020. "Halophytic Hordeum brevisubulatum HbHAK1 Facilitates Potassium Retention and Contributes to Salt Tolerance" International Journal of Molecular Sciences 21, no. 15: 5292. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155292