Proteomic Research in Peritoneal Dialysis

 ,
,  , ,
, ,
Abstract
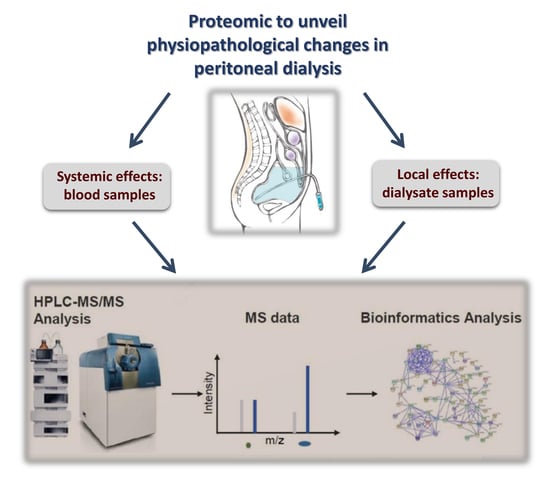
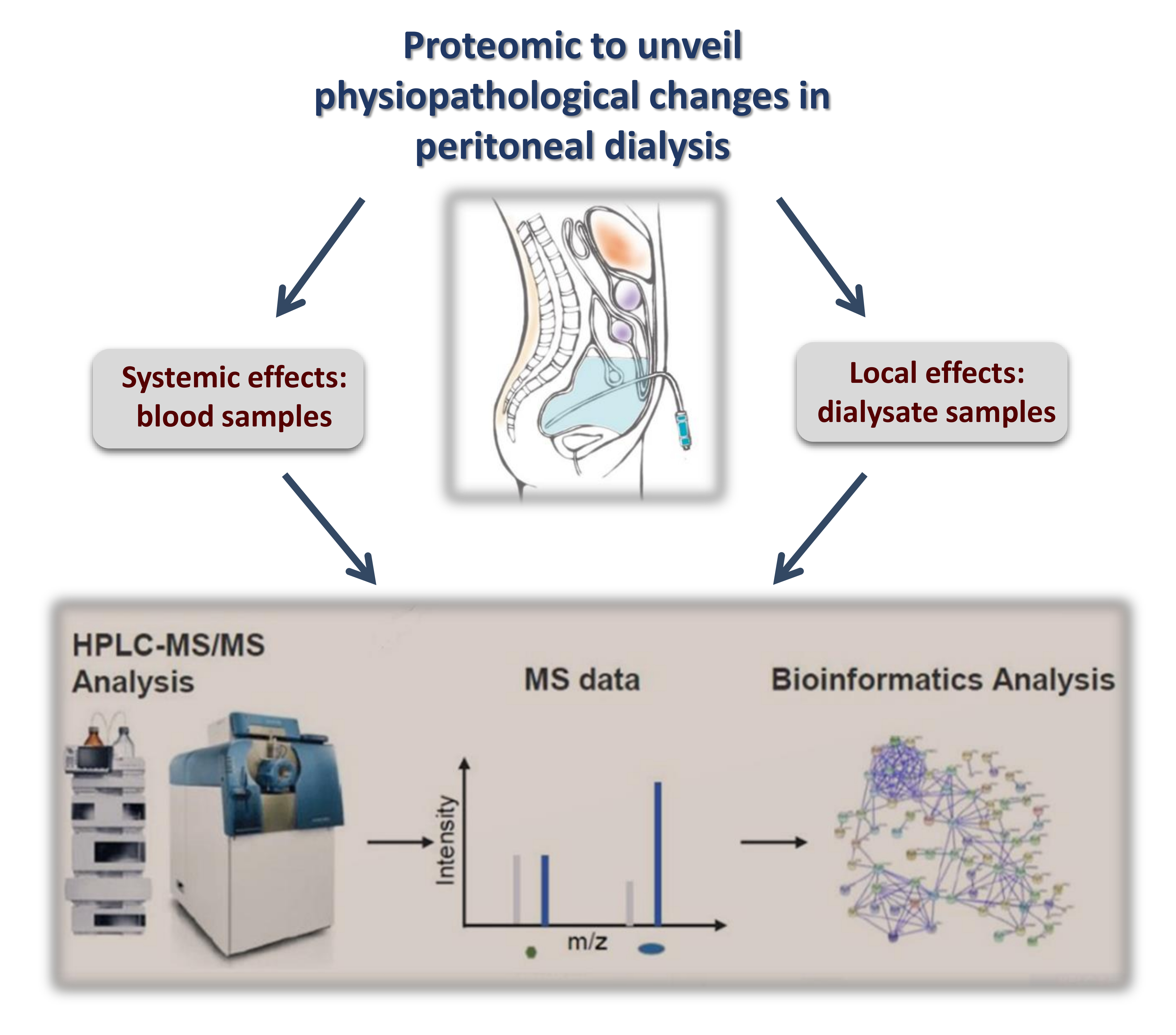
:
1. Introduction
2. Proteomics
3. Proteomic Investigations in Peritoneal Dialysis
3.1. In Vitro Models of PD
3.2. Overview of PDE Proteome
3.3. Effects of Different PD Solutions on PDE Proteome
3.4. Peritoneal Transport Characteristics
3.5. Peritoneal Ultrafiltration Failure
3.6. Peritonitis
3.7. Encapsulating Peritoneal Sclerosis
3.8. Extracellular Vesicles in PDE
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Howell, M.; Walker, R.C.; Howard, K. Cost effectiveness of dialysis modalities: A systemic review of economic evaluations. Appl. Health Econ. Health Policy 2019, 17, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Van Baal, J.O.; Van de Vijver, K.K.; Nieuwland, R.; Van Noorden, C.J.; Van Driel, W.J.; Sturk, A.; Kenter, G.G.; Rokkert, L.G.; Lok, C.A.R. The histophysiology and pathophysiology of the peritoneum. Tissue Cell. 2017, 49, 95–105. [Google Scholar] [CrossRef]
- Gokal, R.; Mallick, R.P. Peritoneal dialysis. Lancet 1999, 353, 823–828. [Google Scholar] [CrossRef]
- Bartosova, M.; Schmitt, C.P. Biocompatible peritoneal dialysis: The target is still way off. Front. Physiol. 2019, 9, 1853. [Google Scholar] [CrossRef]
- Mehrotra, R.; Devuyst, O.; Davies, S.J.; Johnson, D.W. The current state of peritoneal dialysis. J. Am. Soc. Nephrol. 2016, 27, 3238–3252. [Google Scholar] [CrossRef] [Green Version]
- Kramer, A.; Pippias, M.; Noordzij, M.; Stel, V.S.; Afentakis, N.; Ambuhl, P.M.; Andrusev, A.M.; Fuster, E.A.; Arribas, M.F.E.; Åsberg, A.; et al. The European Renal Association–European Dialysis and Transplant Association (ERA-EDTA) Registry Annual Report 2015: A summary. Clin. Kidney J. 2018, 11, 108–122. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.D.; Craig, K.J.; Topley, N.; Von Ruhland, C.; Fallon, M.; Newman, G.R.; Mackenzie, R.K.; Williams, G.T. On behalf of the peritoneal biopsy study group. Morphologic changes in the peritoneal membrane of patients with renal disease. J. Am. Soc. Nephrol. 2002, 13, 470–479. [Google Scholar]
- Bajo, M.A.; Del Peso, G.; Teitelbaum, I. Peritoneal membrane preservation. Semin. Nephrol. 2017, 37, 77–92. [Google Scholar] [CrossRef]
- Davies, S.J.; Phillips, L.; Griffiths, A.; Russell, L.H.; Naish, P.F.; Russell, G.I. What really happens to people on long-term peritoneal dialysis? Kidney Int. 1998, 54, 2207–2217. [Google Scholar] [CrossRef] [Green Version]
- Chaundary, K.; Khanna, R. Biocompatible peritoneal dialysis solutions: Do we have one? Clin. J. Am. Soc. Nephrol. 2010, 5, 723–732. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, C.P.; Aufricht, C. Is there such a thing as biocompatible peritoneal dialysis fluid? Pediatr. Nephrol. 2016, 32, 1835–1843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, C.J. Glucotoxicity in peritoneal dialysis–Solutions for the solution! Adv. Chronic Kidney Dis. 2007, 14, 269–278. [Google Scholar] [CrossRef]
- Wang, I.K.; Lin, C.L.; Chen, H.C.; Lin, S.Y.; Chang, C.T.; Yen, T.H.; Sung, F.C. Risk of new-onset diabetes in end-stage renal disease patients undergoing dialysis: Analysis from registry data of Taiwan. Nephrol. Dial. Transpl. 2018, 33, 670–675. [Google Scholar] [CrossRef]
- Szeto, C.C.; Johnson, D.W. Low GDP solution and glucose-sparing strategies for peritoneal dialysis. Semin. Nephrol. 2017, 37, 30–42. [Google Scholar] [CrossRef]
- Bonomini, M.; Zammit, V.; Divino-Filho, J.C.; Davies, S.J.; Di Liberato, L.; Arduini, A.; Lambie, M. The osmo-metabolic approach: A novel and tantalizing glucose-sparing strategy in peritoneal dialysis. J. Nephrol. 2020, in press. [Google Scholar]
- Aufricht, C.; Beelen, R.; Eberl, M.; Fischbach, M.; Fraser, D.; Jorres, A.; Kratochwill, K.; Lopez Cabrera, M.; Rutherford, P.; Schmitt, C.P.; et al. Biomarker research to improve clinical outcomes of peritoneal dialysis: Consensus of the European Training and Research in Peritoneal Dialysis (EuTRiPD) network. Kidney Int. 2017, 92, 824–835. [Google Scholar] [CrossRef] [Green Version]
- Pauling, L.; Corey, R.B.; Branson, H.R. The structure of proteins; two hydrogen-bonded helical configurations of the polypeptide chain. Proc. Natl. Acad. Sci. USA 1951, 37, 205–211. [Google Scholar] [CrossRef] [Green Version]
- Parker, C.E.; Borchers, C.H. Mass spectrometry based biomarker discovery, verification, and validation—Quality assurance and control of protein biomarker assays. Mol. Oncol. 2014, 8, 840–858. [Google Scholar] [CrossRef]
- Vidova, V.; Spacil, Z. A review on mass spectrometry-based quantitative proteomics: Targeted and data independent acquisition. Anal. Chim. Acta 2017, 964, 7–23. [Google Scholar] [CrossRef] [PubMed]
- Manes, N.P.; Nita-Lazar, A. Application of targeted mass spectrometry in bottom-up proteomics for systems biology research. J. Proteom. 2018, 189, 75–90. [Google Scholar] [CrossRef] [PubMed]
- UniprotKB. Available online: http://www.uniprot.org (accessed on 7 June 2020).
- PeptideAtlas. Available online: http://www.peptideatlas.org (accessed on 7 June 2020).
- PRoteomicsIDEntifications Database (PRIDE). Available online: http://www.ebi.ac.uk/pride (accessed on 7 June 2020).
- Wang, H.Y.; Linc, C.Y.; Chiena, C.C.; Kana, W.C.; Tianf, Y.F.; Liaoh, P.C.; Wui, H.Y.; Su, S.B. Impact of uremic environment on peritoneum: A proteomic view. J. Proteom. 2012, 75, 2053–2063. [Google Scholar] [CrossRef]
- Kratochwill, K.; Lechner, M.; Siehs, C.; Lederhuber, H.C.; Rehulka, P.; Endemann, M.; Kasper, D.C.; Herkner, K.R.; Mayer, B.; Rizzi, A.; et al. Stress responses and conditioning effects in mesothelial cells exposed to peritoneal dialysis fluid. J. Proteome Res. 2009, 8, 1731–1747. [Google Scholar] [CrossRef] [PubMed]
- Lechner, M.; Kratochwill, K.; Lichtenauer, A.; Rehulka, P.; Mayer, B.; Aufricht, C.; Rizzi, A. A proteomic view on the role of glucose in peritoneal dialysis. J. Proteome Res. 2010, 9, 2472–2479. [Google Scholar] [CrossRef] [PubMed]
- Kratochwill, K.; Lechner, M.; Lichtenauer, A.M.; Herzog, R.; Lederhuber, H.C.; Siehs, C.; Endemann, M.; Mayer, B.; Rizzi, A.; Aufricht, C. Interleukin-1 receptor-mediated inflammation impairs the heat shock response of human. Am. J. Pathol. 2011, 178, 1544–1555. [Google Scholar] [CrossRef]
- Herzog, R.; Tarantino, S.; Rudolf, A.; Aufricht, C.; Kratochwill, K.; Witowski, J. Senescence-associated changes in proteome and O-GlcNAcylation pattern in human peritoneal mesothelial cells. BioMed Res. Int. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kratochwill, K.; Bender, T.O.; Lichtenauer, A.M.; Herzog, R.; Tarantino, S.; Bialas, K.; Jörres, A.; Aufricht, C. Cross-omics comparison of stress responses in mesothelial cells exposed to heat- versus filter-sterilized peritoneal dialysis fluids. BioMed Res. Int. 2015. [Google Scholar] [CrossRef] [Green Version]
- Rippe, B. A three-pore model of peritoneal transport. Perit. Dial. Int. 1993, 13, S35–S38. [Google Scholar] [CrossRef] [PubMed]
- Raaijmakers, R.; Pluk, W.; Schroder, C.H.; Gloerich, J.; Cornelissen, E.A.M.; Wessels, H.J.C.T.; Willems, J.L.; Monnens, L.A.H.; Van Den Heuvel, L.P.W.J. Proteomic profiling and identification in peritoneal fluid of children treated by peritoneal dialysis. Nephrol. Dial. Transpl. 2008, 23, 2402–2405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.Y.; Tian, Y.F.; Chien, C.C.; Kan, W.C.; Liao, P.C.; Wu, H.Y.; Su, S.B.; Lin, C.Y. Differential proteomic characterization between normal peritoneal fluid and diabetic peritoneal dialysate. Nephrol. Dial. Transpl. 2010, 25, 1955–1963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.H.; Wang, H.Y.; Lu, C.Y.; Tsai, W.C.; Lin, P.C.; Su, S.B.; Tyan, Y.C. Proteomic profiling for peritoneal dialysate: Differential protein expression in diabetes mellitus. Bio. Med. Res. Int. 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.Y.; Liao, A.C.H.; Huang, C.C.; Liao, P.C.; Chien, C.C.; Kan, W.C.; Wang, H.Y. Comparative proteomic analysis of peritoneal dialysate from chronic glomerulonephritis patients. Bio. Med. Res. Int. 2013. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wen, Q.; Mao, H.P.; Luo, N.; Rong, R.; Fan, J.J.; Yu, X.Q. Developing a reproducible method for the high-resolution separation of peritoneal dialysate proteins on 2-D gels. Protein Expr. Purif. 2013, 89, 96–202. [Google Scholar] [CrossRef]
- Araújo, J.E.; Jorge, S.; Magriço, R.; Teixeira e Costa, F.; Ramos, A.; Reboiro-Jato, M.; Fdez-Riverola, F.; Lodeiro, C.; Capelo, J.L.; Santos, H.M. Classifying patients in peritoneal dialysis by mass spectrometry-based profiling. Talanta 2016, 152, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, E.; Araújo, J.E.; Gómez-Meire, S.; Lodeiro, C.; Perez-Melon, C.; Iglesias-Lamas, E.; Otero-Glez, A.; Capelo, J.L.; Santos, H.M. Proteomics analysis of the peritoneal dialysate effluent reveals the presence of calcium regulation proteins and acute inflammatory response. Clin. Proteom. 2014, 11, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araújo, J.E.; Jorge, S.; Teixeira e Costa, F.; Ramos, A.; Lodeiro, C.; Santos, H.M.; Capelo, J.L. A cost-effective method to get insight into the peritoneal dialysate effluent proteome. J. Proteom. 2016, 145, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Boschetti, E.; Righetti, P.G. The ProteoMiner in the proteomic arena: A non-depleting tool for discovering low-abundance species. J. Proteom. 2008, 71, 255–264. [Google Scholar] [CrossRef]
- Lichtenauer, A.M.; Herzog, R.; Tarantino, S.; Aufricht, C.; Kratochwill, K. Equalizer technology followed by DIGE-based proteomics for detection of cellular proteins in artificial peritoneal dialysis effluents. Electrophoresis 2014, 35, 1387–1394. [Google Scholar] [CrossRef]
- Herzog, R.; Boehm, M.; Unterwurzacher, M.; Wagner, A.; Parapatics, K.; Majek, P.; Mueller, A.C.; Lichtenauer, A.; Bennett, K.L.; Alper, S.L.; et al. Effects of alanyl-glutamine treatment on the peritoneal dialysis effluent proteome reveal pathomechanism-associated molecular signatures. Mol. Cell. Proteom. 2018, 17, 516–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kratochwill, K.; Boehm, M.; Herzog, R.; Gruber, K.; Lichtenauer, A.M.; Kuster, L.; Csaicsich, D.; Gleiss, A.; Alper, S.L.; Aufricht, C.; et al. Addition of alanyl-glutamine to dialysis fluid restores peritoneal cellular stress responses–a first-in-man trial. PLoS ONE 2016, 11, e0165045. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Carpio, V.; Sandoval, P.; Aguilera, A.; Albar-Vizcaino, P.; Perez-Lozano, M.L.; Gonzalez-Mateo, G.T.; Acuna-Ruiz, A.; Garcia-Cantalejo, J.; Botias, P.; Bajo, M.A.; et al. Genomic reprograming analysis of the mesothelial to mesenchymal transition identifies biomarkers in peritoneal dialysis patients. Sci. Rep. 2017, 7, 44941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruschi, M.; Candiano, G.; Santucci, L.; D’Ambrosio, C.; Scaloni, A.; Bonsano, M.; Ghiggeri, G.M.; Verrina, E. Combinatorial peptide ligand library and two dimensional electrophoresis: New frontiers in the study of peritoneal dialysis effluent in pediatric patients. J. Proteom. 2015, 116, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Bucki, R.; Levental, I.; Kulakowska, A.; Janmey, P.A. Plasma gelsolin: Function, prognostic value, and potential therapeutic use. Curr. Protein Pept. Sci. 2008, 9, 541–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuccurullo, M.; Evangelista, C.; Vilasi, A.; Simeoni, M.; Avella, F.; Riccio, E.; Memoli, B.; Malorni, A.; Capasso, G. Proteomic analysis of peritoneal fluid of patients treated by peritoneal dialysis: Effect of glucose concentration. Nephrol. Dial. Transpl. 2011, 26, 1990–1999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mariano, F.; Tetta, C.; Montrucchio, G.; Cavalli, P.L.; Camusssi, G. Role of alpha 1-proteinase inhibitor in restraining peritoneal inflammation in CAPD patients. Kidney Int. 1992, 42, 735–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reixach, N.; Deechongkit, S.; Jiang, X.; Kelly, J.W.; Buxbaum, J. Tissue damage in the amyloidoses: Transthyretin monomers and nonnative oligomers are the major cytotoxic species in tissue culture. Proc. Natl. Acad. Sci. USA 2004, 101, 2817–2822. [Google Scholar] [CrossRef] [Green Version]
- Pesˇic’, I.; Dihazi, G.H.; Muller, G.A.; Jahn, O.; Hoffmann, M.; Eltoweissy, M.; Koziolek, M.; Dihazi, H. Short-time increase of glucose concentration in PDS results in extensive removal and high glycation level of vital proteins during continuous ambulatory peritoneal dialysis. Nephrol. Dial. Transpl. 2011, 26, 2674–2683. [Google Scholar] [CrossRef] [Green Version]
- Dousdampanis, P.; Musso, C.; Trigka, K. Icodextrin and peritoneal dialysis: Advantages and new applications. Int. Urol. Nephrol. 2018, 50, 495–500. [Google Scholar] [CrossRef]
- Goossen, K.; Becker, M.; Marshall, M.R.; Buhn, S.; Breuing, J.; Firanek, C.A.; Hess, S.; Nariai, H.; Sloand, J.A.; Yao, Q.; et al. Icodextrin versus glucose solutions for the once-daily long dwell in peritoneal dialysis: An enriched systematic review and meta-analysis of randomized controlled trials. Am. J. Kidney Dis. 2020, 75, 830–846. [Google Scholar] [CrossRef] [Green Version]
- Bruschi, M.; Candiano, G.; Santucci, L.; Petretto, A.; Mangraviti, S.; Canepa, A.; Perri, K.; Ghiggeri, G.M.; Verrina, E. Proteome profile of peritoneal effluents in children on glucose- or icodextrin-based peritoneal dialysis. Nephrol. Dial. Transpl. 2011, 26, 308–316. [Google Scholar] [CrossRef] [Green Version]
- Vanholder, R.; Pletinck, A.; Schegers, E.; Glorieux, G. Biochemical and clinical impact of organic uremic retention solutes: A comprehensive update. Toxins 2018, 10, 33. [Google Scholar] [CrossRef] [Green Version]
- Del Peso, G.; Jimenez-Heffernan, J.A.; Selgas, R.; Remon, C.; Ossorio, M.; Fernandez-Perpen, A.; Sánchez-Tomero, J.A.; Cirugeda, A.; Se Sousa, E.; Sandoval, P.; et al. Biocompatible dialysis solutions preserve peritoneal mesothelial cell and vessel wall integrity. A case-control study on human biopsies. Perit. Dial. Int. 2015, 36, 129–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaefer, B.; Bartosova, M.; Macher-Goeppinger, S.; Sallay, P.; Voros, P.; Ranchin, B.; Vondrak, K.; Ariceta, G.; Zaloszyc, A.; Bayazit, A.K.; et al. Neutral pH and low-glucose degradation product dialysis fluids induce major early alterations of the peritoneal membrane in children on peritoneal dialysis. Kidney Int. 2018, 94, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Bonomini, M.; Di Liberato, L.; Zammit, V.; Arduini, A. Current opinion on usage of L-carnitine in end-stage renal disease patients on peritoneal dialysis. Molecules 2019, 24, 3449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonomini, M.; Di Silvestre, S.; Di Tomo, P.; Di Pietro, N.; Mandatori, D.; Di Liberato, L.; Sirolli, V.; Chiarelli, F.; Indiveri, C.; Pandolfi, A.; et al. Effect of peritoneal dialysis fluid containing osmometabolic agents on human endothelial cells. Drug Des. Devel. Ther. 2016, 10, 3925–3932. [Google Scholar] [CrossRef] [Green Version]
- Kratochwill, K.; Boehm, M.; Herzog, R.; Lichtenauer, A.M.; Salzer, E.; Lechner, M.; Kuster, L.; Bergmeister, K.; Rizzi, A.; Mayer, B.; et al. Alanyl-glutamine dipeptide restores the cytoprotective stress proteome of mesothelial cells exposed to peritoneal dialysis fluids. Nephrol. Dial. Transpl. 2010, 27, 937–946. [Google Scholar] [CrossRef]
- Herzog, R.; Kuster, L.; Becker, J.; Gluexam, T.; Pils, D.; Spittler, A.; Bhasin, M.K.; Alper, S.L.; Vychytil, A.; Aufricht, C.; et al. Functional and transcriptomic characterization of peritoneal immunemodulation by addition of alanyl-glutamine to dialysis fluid. Sci. Rep. 2017, 7, 6229. [Google Scholar] [CrossRef] [Green Version]
- Vychytil, A.; Herzog, R.; Probst, P.; Ribitsch, W.; Lhotta, K.; Machold-Fabrizii, V.; Wiesholzer, M.; Kaufmann, M.; Salmhofer, H.; Windpessl, M.; et al. A randomized controlled trial of alanyl-glutamine supplementation in peritoneal dialysis fluid to assess impact on biomarkers of peritoneal health. Kidney Int. 2018, 94, 1227–1237. [Google Scholar] [CrossRef] [Green Version]
- Boehm, M.; Herzog, R.; Klinglmüller, F.; Lichtenauer, A.M.; Wagner, A.; Unterwurzacher, M.; Beelen, R.H.J.; Alper, S.L.; Aufricht, C.; Kratochwill, K. The peritoneal surface proteome in a model of chronic peritoneal dialysis reveals mechanisms of membrane damage and preservation. Front. Physiol. 2019, 10, 472. [Google Scholar] [CrossRef]
- Ferrantelli, E.; Liappas, G.; Vila Cuenca, M.; Keuning, E.D.; Foster, T.L.; Vervloet, M.G.; Lopez-Cabrera, M.; Beelen, R.H.J. The dipeptide alanyl-glutamine ameliorates peritoneal fibrosis and attenuates IL-17 dependent pathways during peritoneal dialysis. Kidney Int. 2016, 89, 625–635. [Google Scholar] [CrossRef]
- Twardowski, Z.J. Clinical value of standardized equilibration tests in CAPD patients. Blood Purif. 1989, 7, 95–108. [Google Scholar] [CrossRef]
- Mehrotra, R.; Ravel, V.; Streja, E.; Kuttykrishnan, S.; Adams, S.V.; Katz, R.; Molnar, M.Z.; Kalantar-Zadeh, K. Peritoneal equilibration test and patient outcomes. Clin. J. Am. Soc. Nephrol. 2015, 10, 1990–2001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sritippayawan, S.; Chiangjong, W.; Semangoen, T.; Aiyasanon, N.; Jaetanawanitch, P.; Sinchaikul, S.; Chen, S.T.; Vasuvattakul, S.; Thongboonkerd, V. Proteomic analysis of peritoneal dialysate fluid in patients with different types of peritoneal membranes. J. Prot. Res. 2007, 6, 4356–4362. [Google Scholar] [CrossRef] [Green Version]
- Wen, Q.; Zhang, L.; Mao, H.P.; Tang, X.Q.; Fan, R.R.J.J.; Yu, X.Q. Proteomic analysis in peritoneal dialysis patients with different peritoneal transport characteristics. Biochem. Biophys. Res. Commun. 2013, 438, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Ferrantelli, E.; Farhat, K.; Hipgrave Ederveen, A.L.; Reiding, K.R.; Beelen, R.H.J.; Van Ittersum, F.J.; Wuhrer, M.; Dotz, V. Effluent and serum protein N-glycosylation is associated with inflammation and peritoneal membrane transport characteristic sin peritoneal dialysis patients. Sci. Rep. 2018, 8, 979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kariya, T.; Nishimura, H.; Mizuno, M.; Suzuki, Y.; Matsukawa, Y.; Sakata, F.; Maruyama, S.; Takei, Y.; Ito, Y. TGF-1-VEGF-A pathway induces neoangiogenesis with peritoneal fibrosis in patients undergoing peritoneal dialysis. Am. J. Physiol. Renal Physiol. 2018, 314, F167–F180. [Google Scholar] [CrossRef]
- Wang, A.Y.; Dong, J.; Xu, X.; Davies, S. Volume management as a key dimension of a high-quality PD prescription. Perit Dial Int. 2020, 40, 282–292. [Google Scholar] [CrossRef] [Green Version]
- Teitelbaum, I. Ultrafiltration failure in peritoneal dialysis: A pathophysiologic approach. Blood Purif. 2015, 39, 70–73. [Google Scholar] [CrossRef]
- Krediet, R.T. Ultrafiltration failure is a reflection of peritoneal alterations in patients treated with peritoneal dialysis. Front. Physiol. 2018, 9, 1815. [Google Scholar] [CrossRef] [Green Version]
- Heimburger, O.; Waniewski, J.; Werynski, A.; Lindholm, B. A quantitative description of solute and fluid transport during peritoneal dialysis. Kidney Int. 1992, 41, 1320–1332. [Google Scholar] [CrossRef] [Green Version]
- Bartosova, M.; Schaefer, B.; Bermejo, J.L.; Tarantino, S.; Lasitschka, F.; Macher-Goeppinger, S.; Sinn, P.; Warady, B.A.; Zaloszyc, A.; Parapatics, K.; et al. Complement activation in peritoneal dialysis–induced arteriolopathy. J. Am. Soc. Nephrol. 2018, 29, 268–282. [Google Scholar] [CrossRef]
- Lemmers, R.F.H.; Vilaj, M.; Urda, D.; Agakov, F.; Šimurina, M.; Klaric, L.; Rudan, I.; Campbell, H.; Hayward, C.; Wilson, J.F.; et al. IgG glycan patterns are associated with type 2 diabetes in independent European populations. Biochim. Biophys. Acta 2017, 1861, 2240–2249. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.S.; Heijmans, R.; Meulen, C.K.E.; Lieverse, A.G.; Gornik, O.; Sijbrands, E.J.G.; Lauc, G.; Van Hoek, M. Association of the IgG N-glycome with the course of kidney function in type 2 diabetes. BMJ Open Diabetes Res. Care. 2020, 8, e001026. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.T.; Ravetch, J.V. Functional diversification of IgGs through Fc glycosylation. J. Clin. Invest. 2019, 29, 3492–3498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jegatheesan, D.; Cho, Y.; Johnson, D.W. Clinical studies of interventions to mitigate cardiovascular risk in peritoneal dialysis patients. Semin. Nephrol. 2018, 38, 277–290. [Google Scholar] [CrossRef]
- Stachowska-Pietka, J.; Poleszczuk, J.; Flessner, M.F.; Lindholm, B.; Waniewski, J. Alterations of peritoneal transport characteristics in dialysis patients with ultrafiltration failure: Tissue and capillary components. Nephrol. Dial. Transpl. 2019, 34, 864–870. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.H.; Perl, J.; Teitelbaum, I. Prescribing high-quality peritoneal dialysis: The role of preserving residual kidney function. Perit. Dial. Int. 2020, 40, 274–281. [Google Scholar] [CrossRef] [Green Version]
- Perl, J.; Fuller, D.S.; Bieber, B.A.; Boudville, N.; Kanjanabuch, T.; Ito, Y.; Nessim, S.J.; Piraino, B.M.; Pisoni, R.L.; Robinson, B.M.; et al. Peritoneal dialysis-related infection rats and outcomes: Results from the peritoneal dialysis outcomes and practice patterns study (PDOPPS). Am. J. Kidney Dis. 2020, 76, 42–53. [Google Scholar] [CrossRef] [Green Version]
- Li, P.K.T.; Szeto, C.C.; Piraino, B.; De Arteaga, J.; Fan, S.; Figueiredo, A.E.; Fish, D.N.; Goffin, E.; Kim, Y.L.; Salzer, W.; et al. ISPD peritonitis recommendations: 2016 update on prevention and treatment. Perit. Dial. Int. 2016, 36, 481–508. [Google Scholar] [CrossRef] [Green Version]
- Davies, S.J.; Phillips, L.; Naish, P.F.; Russell, G.I. Peritoneal glucose exposure and changes in membrane solute transport with time on peritoneal dialysis. J. Am. Soc. Nephrol. 2001, 12, 1046–1051. [Google Scholar]
- Schaefer, F.; Borzych-Duzalka, D.; Azocar, M.; Munarriz, R.L.; Sever, L.; Aksu, N.; Sànchez Barbosa, L.; Galan, Y.S.; Xu, H.; Coccia, P.A.; et al. Impact of global economic disparities on practices and outcomes of chronic peritoneal dialysis in children: Insights from the International Pediatric Peritoneal Dialysis Network Registry. Perit. Dial. Int. 2012, 32, 399–409. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.T.; Tsai, C.C.; Chen, C.Y.; Lee, W.J.; Su, C.C.; Wu, Y.J. Proteomic analysis of peritoneal dialysate fluid in patients with dialysis-related peritonitis. Ren. Fail. 2008, 30, 772–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyan, Y.C.; Su, S.B.; Ting, S.S.; Wang, H.Y.; Liao, P.C. A comparative proteomics analysis of peritoneal dialysate before and after the occurrence of peritonitis episode by mass spectrometry. Clin. Chim. Acta 2013, 420, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Guo, N.; Wen, Q.; Li, Z.J.; Xu, R.C.; Peng, F.F.; Yu, X.Q. Optimization and evaluation of magnetic bead separation combined with matrix-assisted laser desorption/ionization time-of-flight mass spectroscopy (MALDI-TOF MS) for proteins profiling of peritoneal dialysis effluent. Int. J. Mol. Sci. 2014, 15, 1162–1175. [Google Scholar] [CrossRef] [PubMed]
- Aldriwesh, M.; Al-Dayan, N.; Barratt, J.; Freestone, P. The iron biology status of peritoneal dialysis patients may be a risk factor for development of infectious peritonitis. Perit. Dial. Int. 2019, 39, 362–374. [Google Scholar] [CrossRef]
- Kawabata, H. Transferrin and transferrin receptors update. Free Rad. Biol. Med. 2019, 133, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Danford, C.J.; Lin, S.C.; Smith, M.P.; Wolf, J.L. Encapsulating peritoneal sclerosis. World J. Gastroenterol. 2018, 24, 3101–3111. [Google Scholar] [CrossRef]
- Brown, E.A.; Bargman, J.; Van Biesen, W.; Chang, M.Y.; Finkelstein, F.O.; Hurst, H.; Johnson, D.W.; Kawanishi, H.; Lambie, M.; De Moraes, T.P.; et al. Length of time on peritoneal dialysis and encapsulating peritoneal sclerosis—Position paper for ISPD: 2017 update. Perit. Dial. Int. 2017, 37, 362–374. [Google Scholar] [CrossRef]
- Alston, H.; Fan, S.; Nakayama, M. Encapsulating peritoneal sclerosis. Semin. Nephrol. 2017, 37, 93–102. [Google Scholar] [CrossRef]
- Maruyama, Y.; Numata, M.; Nakayama, M.; Matsuo, N.; Nordfors, L.; Hosoya, T.; Lindholm, B. Relationship between the -374T/A receptor of advanced glycation end products gene polymorphism and peritoneal solute transport status at the initiation of peritoneal dialysis. Ther. Apher. Dial. 2007, 11, 301–305. [Google Scholar] [CrossRef]
- Betjes, M.G.; Habib, S.M.; Boeschoten, E.W.; Hemke, A.C.; Struijk, D.G.; Westerhuis, R.; Abrahams, A.C.; Korte, M.R. Significant decreasing incidence of encapsulating peritoneal sclerosis in the Dutch population of peritoneal dialysis patients. Perit. Dial. Int. 2017, 37, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Zavvos, V.; Buxton, A.T.; Evans, C.; Lambie, M.; Davies, S.J.; Topley, N.; Wilkie, M.; Summers, A.; Brenchley, P.; Goumenos, D.S.; et al. A prospective, proteomics study identified potential biomarkers of encapsulating peritoneal sclerosis in peritoneal effluent. Kidney Int. 2017, 92, 988–1002. [Google Scholar] [CrossRef] [Green Version]
- Lambie, M.J.; Chess, J.; Summers, A.; Williams, P.F.; Topley, N.; Davies, S.J. GLOBAL Fluid Study Investigators. Peritoneal inflammation precedes encapsulating peritoneal sclerosis: Results from the GLOBAL fluid study. Nephrol. Dial. Transpl. 2016, 31, 480–486. [Google Scholar] [CrossRef] [PubMed]
- Akbari, S.; Abou-Arkoub, R.; Sun, S.; Hiremath, S.; Reunov, A.; McCormick, B.B.; Ruzicka, M.; Burger, D. Microparticle formation in peritoneal dialysis: A proof of concept study. Can. J. Kidney Heal. Dis. 2017, 4, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Carreras-Planella, L.; Soler-Majoral, J.; Rubio-Esteve, C.; Lorano-Ramos, S.I.; Franquesa, M.; Bonet, J.; Troya-Saborido, M.I.; Borras, F.E. Characterization and proteomic profile of extracellular vesicles from peritoneal dialysis efflux. PLoS ONE 2017, 12, e0176987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearson, L.J.; Klaharn, I.Y.; Thongsawang, B.; Manuprasert, W.; Saejew, T.; Somparn, P.; Chuengsaman, P.; Kanjanabuch, T.; Pisitkun, T. Multiple extracellular vesicle types in peritoneal dialysis effluent are prominent and contain known biomarkers. PLoS ONE 2017, 12, e0178601. [Google Scholar] [CrossRef] [Green Version]
- Corciulo, S.; Nicoletti, M.C.; Mastrofrancesco, L.; Milano, S.; Mastrodonato, M.; Carmosino, M.; Gerbino, A.; Corciulo, R.; Russo, R.; Svolto, M.; et al. AQP1-containing exosomes in peritoneal dialysis effluent as biomarker of dialysis efficiency. Cells 2019, 8, 330. [Google Scholar] [CrossRef] [Green Version]
- Monguió-Tortajada, M.; Gálvez-Montón, C.; Bayes-Genis, A.; Roura, S.; Borràs, F.E. Extracellular vesicle isolation methods: Rising impact of size-exclusion chromatography. Cell Mol. Life Sci. 2019, 76, 2369–2382. [Google Scholar] [CrossRef]
- Carreras-Planella, L.; Soler-Majoral, J.; Rubio-Esteve, C.; Morón-Font, M.; Franquesa, M.; Bonal, J.; Troya-Saborido, M.I.; Borràs, F.E. Proteomic profiling of peritoneal dialysis effluent-derived extracellular vesicles: A longitudinal study. J. Nephrol. 2019, 32, 1021–1031. [Google Scholar] [CrossRef]

{kind=link}
| Proteomic Strategy | Condition | Findings | Reference |
|---|---|---|---|
| 2DE followed by LC–MS/MS and WB | Uremia | Higher in uremia: KNG1, apoptosis inhibitor 2, CECR2, and APOA1. | [24] |
| 2DE followed by LC–MS/MS | Diabetes mellitus | Higher in diabetic: DBP, HP, and B2M. Lower in diabetic: C4A and IgK. | [32] |
| RP-nano-UPLC–ESI–MS/MS followed by peptide fragmentation patterning | Diabetic vs. chronic glomerulonephritis | Upregulated in diabetic: APOA-IV, AZGP1, AIF4A1, and HLA-A. Downregulated in diabetic: albumin, AMBP, APOA-I, IgG1-Fc, mutant RBP, and HP alpha2. | [33] |
| 2DE followed by LC–MS/MS | Chronic glomerulonephritis at the beginning of CAPD and after 1 year | Higher at the beginning: IGHM, FGG, and CRP. Higher after 1 year: IGHD, SERPINA1, HRG, APOA-I, and SAP. | [34] |
| ACN- and DTT-based methods before 2-D GE and MS | PD | Loss of DBP, HP, CP may be negative for PD. Removal of adipokine or RBP4 may be positive. | [38] |
| CPLL and 2DE | CPLL treatment in PDE from pediatric patients | Decrease in CPLL-treated samples: albumin, Ig, SERPINA1, TF, and A1M. Decrease along PD of GSN and increase of ITLN1. | [44] |
| 1DE with nano-RP-HPLC–ESI–MS/MS and 2-DE with MALDI-TOF–MS | PD solutions at glucose 1.5%, 2.5%, or 4.25% | Under-expressed in 4.25% glucose: SERPINA1, FGB, APO A-IV, and TTR. | [46] |
| Multiple Affinity Removal LC Column-Human 6, 2DE DIGE, MS and 2D WB | Stay-Safe Balance vs. Physioneal solutions | Increase in higher glucose concentration: AGEs in PDE. | [49] |
| 2DE and MS | 7.5% icodextrin solution vs. 3.86% glucose solution | Higher removal of B2M and CST3 with 7.5% icodextrin solution. | [52] |
| 1D immunoblot, 2D-DIGE, 2D WB, and saturation labeling | Standard PD solution vs. AlaGn-containing PD solution | AlaGln-containing solution reduced PM injury and improved cellular stress. Inhibition of upstream IFG, VEGF, and TGF-β1. | [42] |
| MALDI-Q-TOF–MS and MS/MS | Different transport rates | Increased in high transport: C4A, IGK. | [65] |
| 2D DIGE and MALDI-TOF–MS/MS | Different PM types | Increased in high transport: DBP, C3, APOA1. | [66] |
| MALDI-TOF–MS and glycosylation profile | PM transport rate | Positively associated with triantennary glycans and the α2,6-syalilation of those, and negatively associated with diantennary glycans and the α2,6-syalilation. | [67] |
| LC–MS | CKD and PD on omental arterioles of pediatric patients | CKD: activation of metabolic processes. PD: inflammatory, immunologic, and stress-response cascades. Dialytic glucose correlates with PD vasculopathy and activation of TGF-β pathways. Activated complement system and TGF-β signaling cascade in PD vasculopathy. | [73] |
| Glycosylation profile | Type 2 diabetes | Different IgG N-glycosylation patterns in diabetes. | [75] |
| MALDI-TOF–MS and glycosylation profile | Different PD solutions over time | Increase of an IgG glycosylation pattern over time and in peritonitis. | [67] |
| 2DE and SELDI-TOF–MS | Peritonitis | Increased in peritonitis: B2M. | [84] |
| 2DE and RP-nano-HPLC–ESI–MS/MS | Peritonitis | Higher in peritonitis: HP, SERPINC1. Decreased in peritonitis: HSP70 1A/1B, APOA-1, ITIH4, FGG and FGB, CP, SERPINA1, and AZGP1. | [85] |
| Magnetic bead separation and MALDI-TOF–MS | Peritonitis | Different 1-15 kDa protein and peptide patterns. | [86] |
| MALDI-TOF–MS and radioactive iron-labeled transferrin | Peritonitis | Increased in peritonitis: iron-saturated transferrin. It can also act as bacteria growth source. | [87] |
| 2D SDS-PAGE/MS and iTRAQ | EPS | 3 to 5 years before EPS: changes in COL1A1, g-actin, CFB and CFI, and SERPINA1. 2 years before EPS: GSN, APOA2, APOA4, and HBB. With EPS symptoms: ORM, ITLN1, and AHSG chain B. | [94] |
| Immunogold staining and TEM | Dialysis efficiency | Exosomal AQP1 positively correlates with PD effluent and ultrafiltration, free water transport, and Na sieving. | [99] |
| LC–MS/MS | PM transport rate | Different extracellular vesicles proteome patterns upon PET. | [101] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonomini, M.; Borras, F.E.; Troya-Saborido, M.; Carreras-Planella, L.; Di Liberato, L.; Arduini, A. Proteomic Research in Peritoneal Dialysis. Int. J. Mol. Sci. 2020, 21, 5489. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155489
Bonomini M, Borras FE, Troya-Saborido M, Carreras-Planella L, Di Liberato L, Arduini A. Proteomic Research in Peritoneal Dialysis. International Journal of Molecular Sciences. 2020; 21(15):5489. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155489
Chicago/Turabian StyleBonomini, Mario, Francesc E. Borras, Maribel Troya-Saborido, Laura Carreras-Planella, Lorenzo Di Liberato, and Arduino Arduini. 2020. "Proteomic Research in Peritoneal Dialysis" International Journal of Molecular Sciences 21, no. 15: 5489. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155489