Synaptic Organization of the Human Temporal Lobe Neocortex as Revealed by High-Resolution Transmission, Focused Ion Beam Scanning, and Electron Microscopic Tomography

 and
and
Abstract
:1. Introduction
2. Results
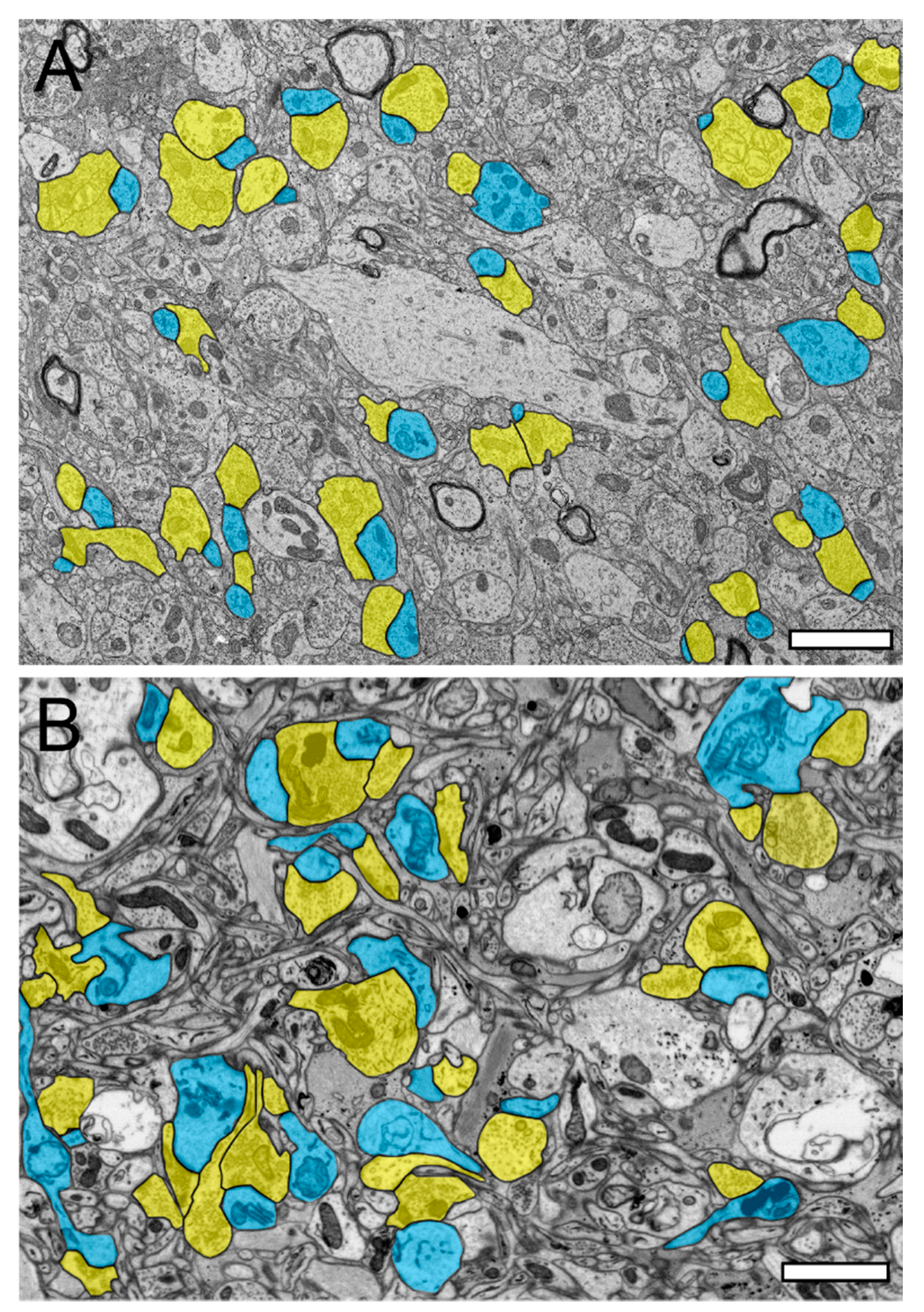
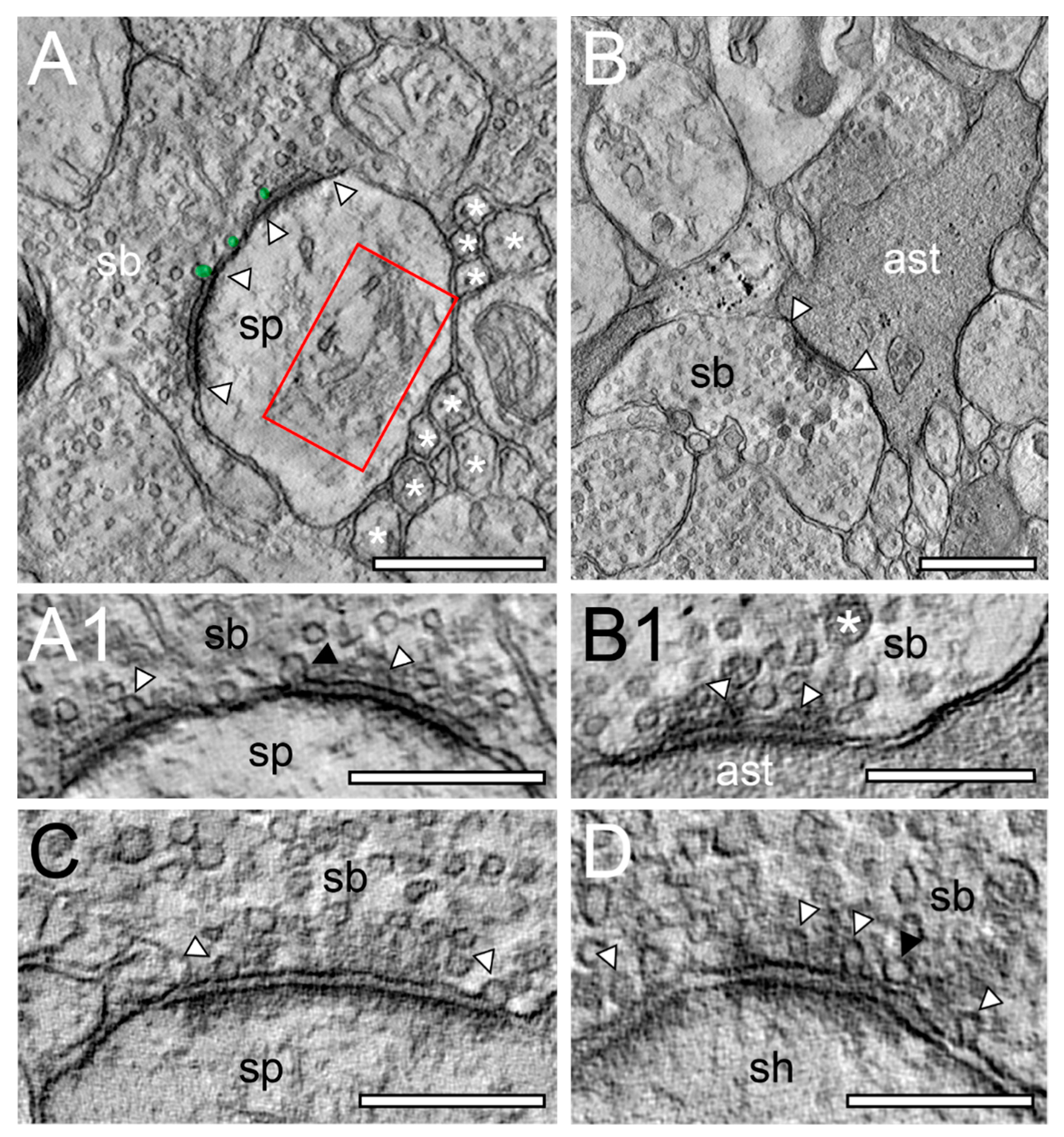
2.1. Comparison of TEM and FIB-SEM to Investigate the Neuropil and Synaptic Organization of the TLN
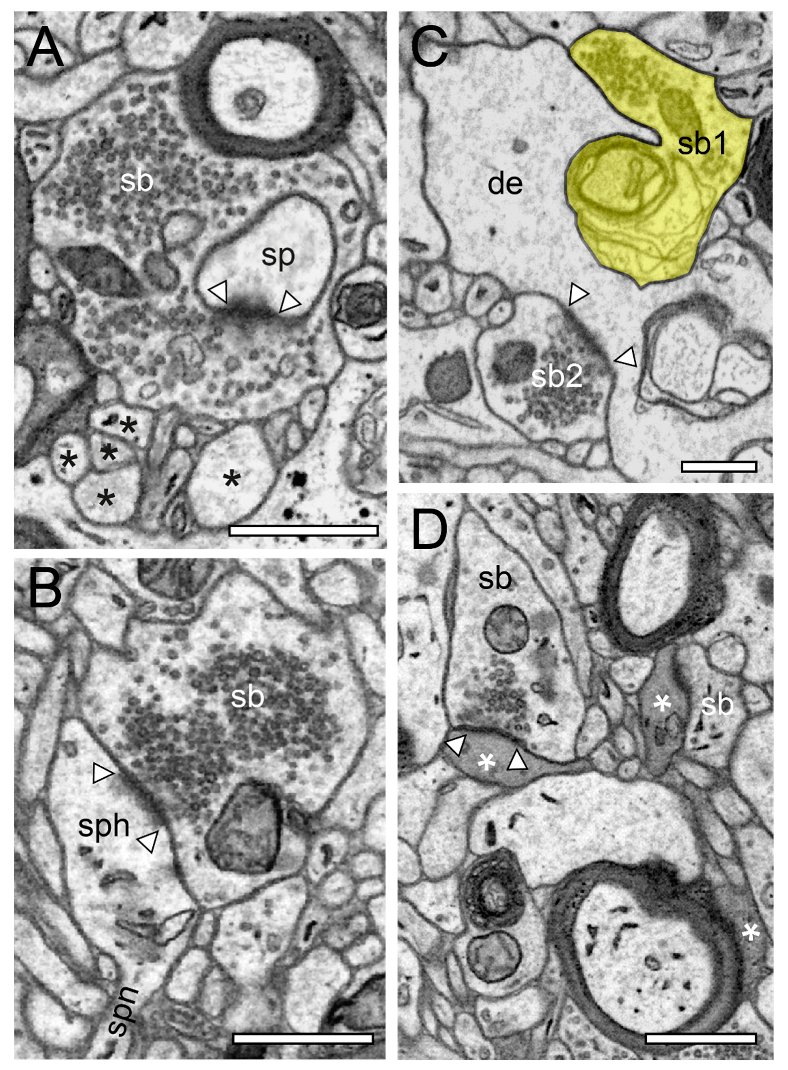
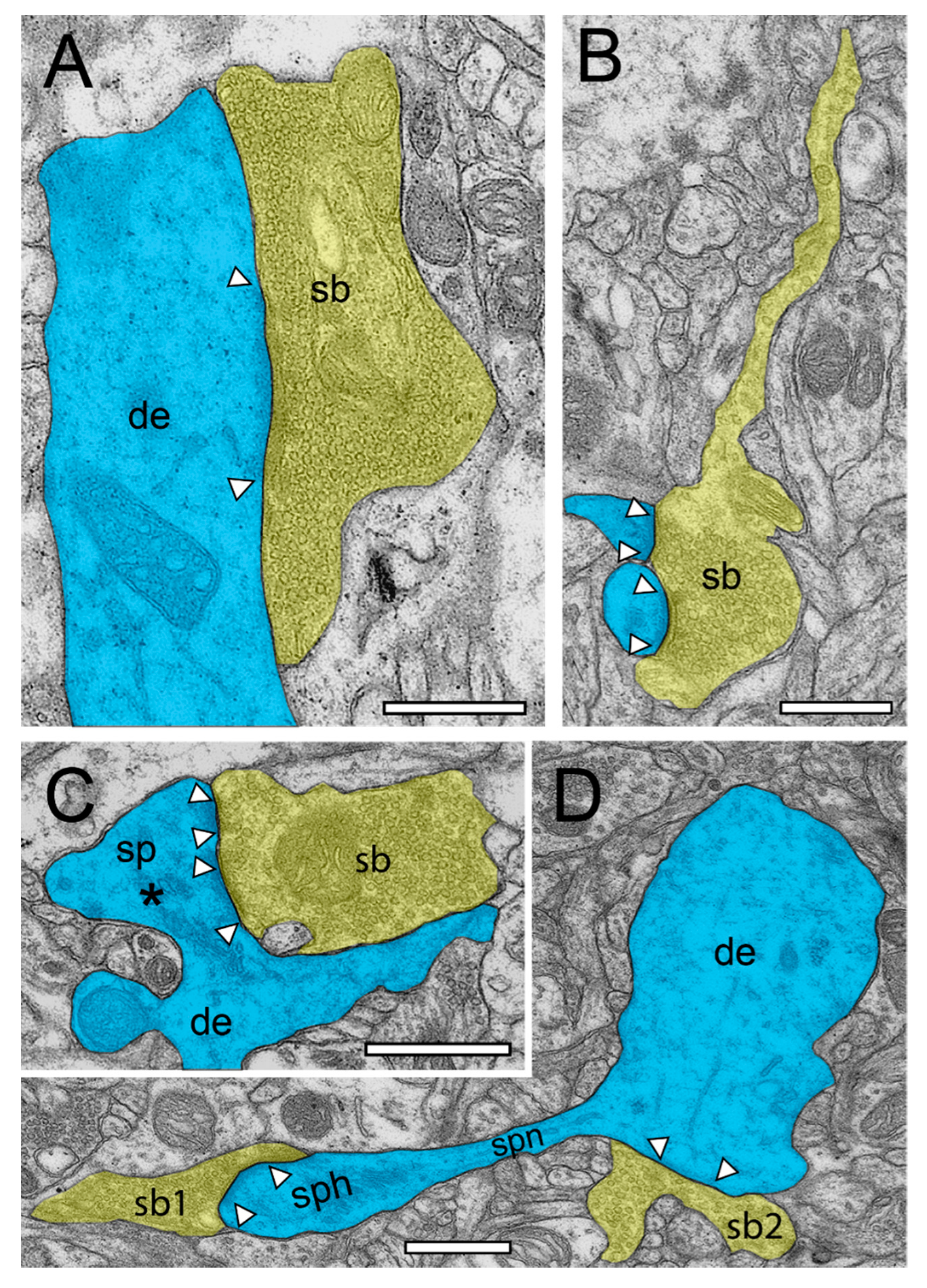
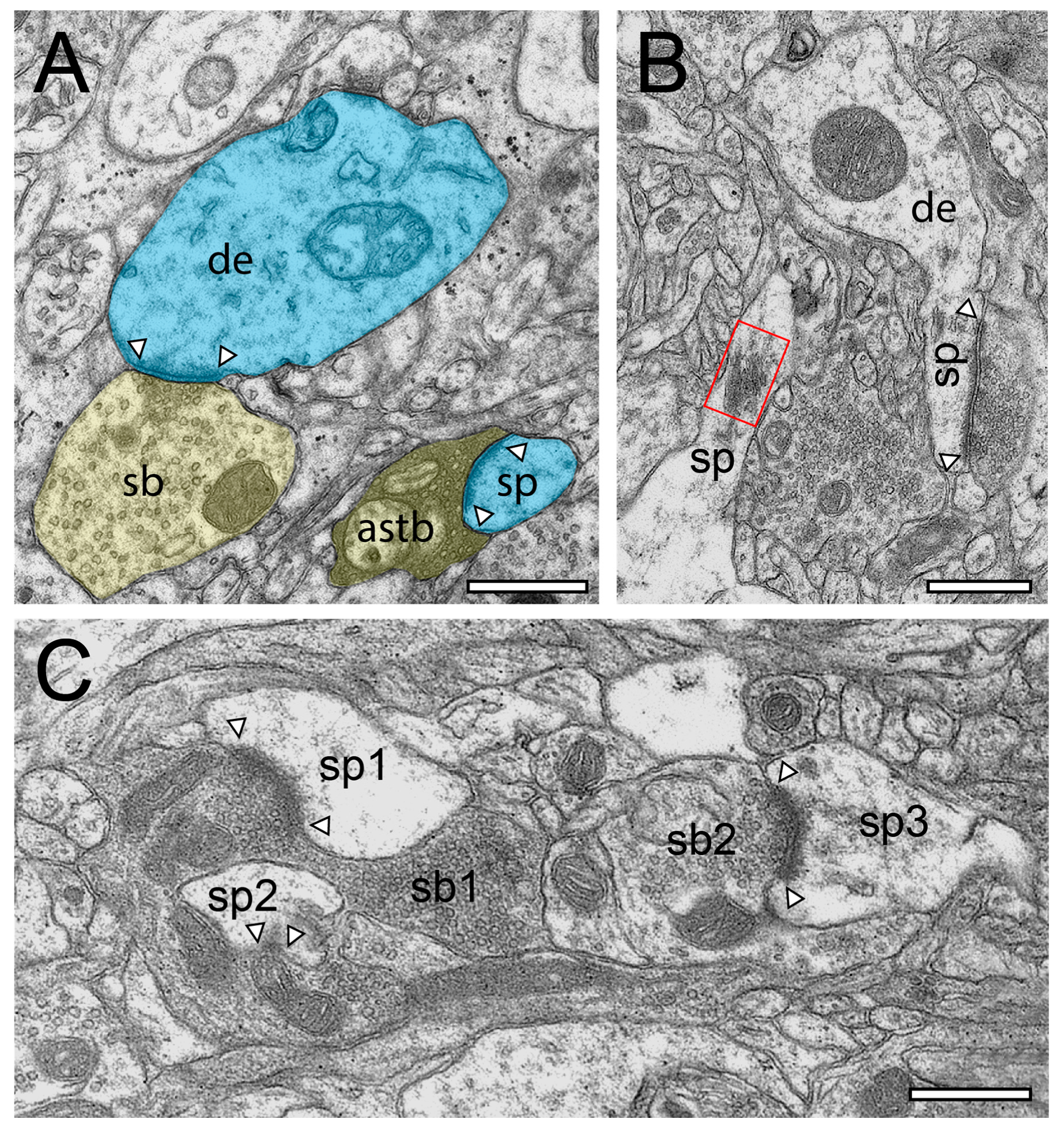
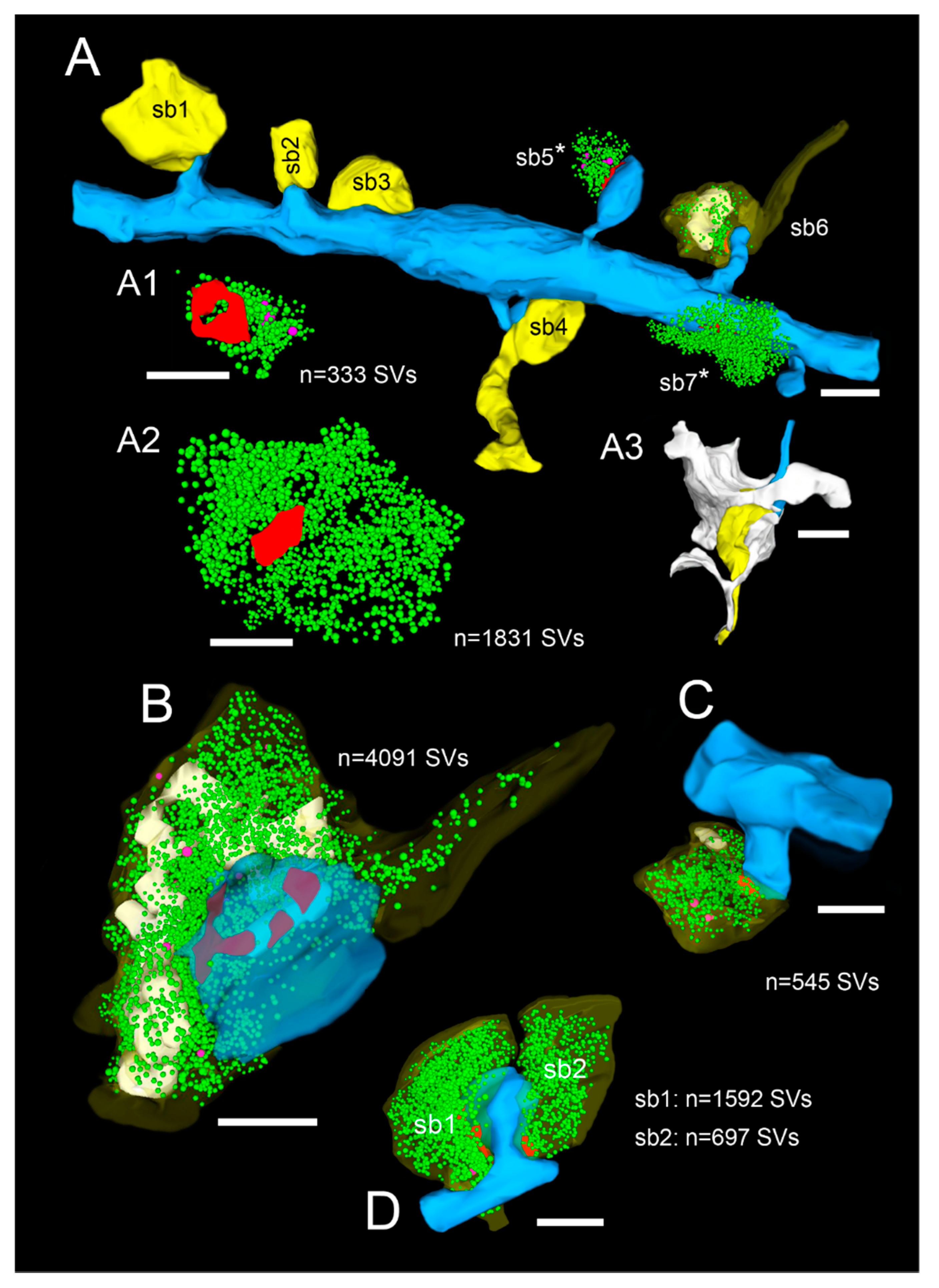
2.2. Very Special Entities: Synaptic Boutons in the Human TLN
2.3. EM Tomography in the Human TLN
3. Discussion
3.1. Methodological Considerations
3.2. Functional Significance Working with Human Tissue Samples
3.3. Perspective to Work with Human Tissue Samples
4. Materials and Methods
4.1. Human Neocortical Tissue Processing for TEM, FIB-SEM, and EM Tomography
4.2. Fixation and Sectioning
4.3. Processing for TEM
4.4. Processing for FIB-SEM
4.5. Serial Ultrathin Sectioning, TEM Data Acquisition, and 3D Volume Reconstructions
4.6. TEM Data Acquisition
4.7. FIB-SEM Data Acquisition
4.8. 3D-Volume Reconstructions
4.9. Analyzing Vesicle Distribution and Pool Sizes
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AZ | Active zone |
| CB | Cacodylate buffer |
| EM | Electron microscopy |
| FIB-SEM | Focused ion beam scanning electron microscopy |
| L | Layer |
| MRI/PET | Magnetic resonance imaging/positron emission tomography |
| PB | Phosphate buffer |
| PO | Propylene oxide |
| RP | Recycling pool |
| RRP | Readily releasable pool |
| RT | Room temperature |
| SB | Synaptic bouton |
| SV | Synaptic vesicle |
| TEM | Transmission electron microscopy |
| TLN | Temporal lobe neocortex |
References
- Von Economo, C.; Koskinas, G.N. Die Cytoarchitektonik der Hirnrinde des Erwachsenen Menschen; Verlag von Julius Springer: Wien, Austria; Berlin, Germany, 1925. [Google Scholar]
- Brodmann, K. Beiträge zur histologischen lokalisation der grosshirnrinde. I. Mitteilung: Die regio rolandica. J. Psychol. Neurol. (Leipzig) 1903, 2, 79–107. [Google Scholar]
- Zilles, K.; Amunts, K. Centenary of brodmann’s map—Conception and fate. Nat. Rev. Neurosci. 2010, 11, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Zilles, K. Brodmann: A pioneer of human brain mapping-his impact on concepts of cortical organization. Brain 2018, 141, 3262–3278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, B.; Schall, M.; Prather, M.; Kapler, E.; Driscoll, L.; Baca, S.; Jacobs, J.; Ford, K.; Wainwright, M.; Tremel, M. Regional dendritic and spine variation in human cerebral cortex: A quantitative golgi study. Cereb. Cortex 2001, 11, 558–571. [Google Scholar] [CrossRef] [PubMed]
- Petanjek, Z.; Judas, M.; Simic, G.; Rasin, M.R.; Uylings, H.B.; Rakic, P.; Kostovic, I. Extraordinary neoteny of synaptic spines in the human prefrontal cortex. Proc. Natl. Acad. Sci. USA 2011, 108, 13281–13286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, A. Golgi, cajal, and the fine structure of the nervous system. Brain Res. Rev. 2007, 55, 256–263. [Google Scholar] [CrossRef] [Green Version]
- Buhl, E.H.; Lübke, J. Intracellular lucifer yellow injection in fixed brain slices combined with retrograde tracing, light and electron microscopy. Neuroscience 1989, 28, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Mikkonen, M.; Pitkanen, A.; Soininen, H.; Alafuzoff, I.; Miettinen, R. Morphology of spiny neurons in the human entorhinal cortex: Intracellular filling with lucifer yellow. Neuroscience 2000, 96, 515–522. [Google Scholar] [CrossRef]
- Benavides-Piccione, R.; Fernaud-Espinosa, I.; Robles, V.; Yuste, R.; DeFelipe, J. Age-Based comparison of human dendritic spine structure using complete three-dimensional reconstructions. Cereb. Cortex 2013, 23, 1798–1810. [Google Scholar] [CrossRef] [Green Version]
- Nieuwenhuys, R.; Broere, C.A.; Cerliani, L. A new myeloarchitectonic map of the human neocortex based on data from the Vogt-Vogt school. Brain Struct. Funct. 2015, 220, 2551–2573. [Google Scholar] [CrossRef]
- Palomero-Gallagher, N.; Zilles, K. Cortical layers: Cyto-, myelo-, receptor- and synaptic architecture in human cortical areas. Neuroimage 2019, 197, 716–741. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.S.; Da Silva, A.B.; Whittaker, R.G.; Woodhall, G.L.; Cunningham, M.O. Human brain slices for epilepsy research: Pitfalls, solutions and future challenges. J. Neurosci. Methods 2016, 260, 221–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molnar, G.; Rozsa, M.; Baka, J.; Holderith, N.; Barzo, P.; Nusser, Z.; Tamas, G. Human pyramidal to interneuron synapses are mediated by multi-vesicular release and multiple docked vesicles. Elife 2016, 5, e18167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szegedi, V.; Paizs, M.; Csakvari, E.; Molnar, G.; Barzo, P.; Tamas, G.; Lamsa, K. Plasticity in single axon glutamatergic connection to GABAergic interneurons regulates complex events in the human neocortex. PLoS Biol. 2016, 14, e2000237. [Google Scholar] [CrossRef] [PubMed]
- Szegedi, V.; Molnar, G.; Paizs, M.; Csakvari, E.; Barzo, P.; Tamas, G.; Lamsa, K. High-Precision fast-spiking basket cell discharges during complex events in the human neocortex. ENeuro 2017, 4. [Google Scholar] [CrossRef] [Green Version]
- Seeman, S.C.; Campagnola, L.; Davoudian, P.A.; Hoggarth, A.; Hage, T.A.; Bosma-Moody, A.; Baker, C.A.; Lee, J.H.; Mihalas, S.; Teeter, C.; et al. Sparse recurrent excitatory connectivity in the microcircuit of the adult mouse and human cortex. Elife 2018, 7, e37349. [Google Scholar] [CrossRef]
- Bocchio, M.; Lukacs, I.P.; Stacey, R.; Plaha, P.; Apostolopoulos, V.; Livermore, L.; Sen, A.; Ansorge, O.; Gillies, M.J.; Somogyi, P.; et al. Group II metabotropic glutamate receptors mediate presynaptic inhibition of excitatory transmission in pyramidal neurons of the human cerebral cortex. Front. Cell Neurosci. 2018, 12, 508. [Google Scholar] [CrossRef]
- Mansvelder, H.D.; Verhoog, M.B.; Goriounova, N.A. Synaptic plasticity in human cortical circuits: Cellular mechanisms of learning and memory in the human brain? Curr. Opin. Neurobiol. 2019, 54, 186–193. [Google Scholar] [CrossRef]
- Blazquez-Llorca, L.; Merchan-Perez, A.; Rodriguez, J.R.; Gascon, J.; DeFelipe, J. FIB-SEM technology and Alzheimer’s disease: Three-Dimensional analysis of human cortical synapses. J. Alzheimer’s Dis. 2013, 34, 995–1013. [Google Scholar] [CrossRef] [Green Version]
- Kay, K.R.; Smith, C.; Wright, A.K.; Serrano-Pozo, A.; Pooler, A.M.; Koffie, R.; Bastin, M.E.; Bak, T.H.; Abrahams, S.; Kopeikina, K.J.; et al. Studying synapses in human brain with array tomography and electron microscopy. Nat. Protoc. 2013, 8, 1366–1380. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.B.; Schumann, C.M. Optimization of electron microscopy for human brains with long-term fixation and fixed-frozen sections. Acta Neuropathol. Commun. 2014, 2, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominguez-Alvaro, M.; Montero-Crespo, M.; Blazquez-Llorca, L.; DeFelipe, J.; Alonso-Nanclares, L. 3D electron microscopy study of synaptic organization of the normal human transentorhinal cortex and its possible alterations in Alzheimer’s disease. ENeuro 2019, 6. [Google Scholar] [CrossRef] [PubMed]
- Yakoubi, R.; Rollenhagen, A.; Von Lehe, M.; Shao, Y.; Sätzler, K.; Lübke, J.H.R. Quantitative three-dimensional reconstructions of excitatory synaptic boutons in layer 5 of the adult human temporal lobe neocortex: A fine-scale electron microscopic analysis. Cereb. Cortex 2019, 29, 2797–2814. [Google Scholar] [CrossRef]
- Yakoubi, R.; Rollenhagen, A.; Von Lehe, M.; Miller, D.; Walkenfort, B.; Hasenberg, M.; Sätzler, K.; Lübke, J.H.R. Ultrastructural heterogeneity of layer 4 excitatory synaptic boutons in the adult human temporal lobe neocortex. Elife 2019, 8, e48373. [Google Scholar] [CrossRef] [PubMed]
- Kiernan, J.A. Anatomy of the temporal lobe. Epilepsy Res. Treat. 2012, 2012, 176157. [Google Scholar] [CrossRef]
- Insausti, R. Comparative neuroanatomical parcellation of the human and nonhuman primate temporal pole. J. Comp. Neurol. 2013, 521, 4163–4176. [Google Scholar] [CrossRef] [PubMed]
- Allone, C.; Lo Buono, V.; Corallo, F.; Pisani, L.R.; Pollicino, P.; Bramanti, P.; Marino, S. Neuroimaging and cognitive functions in temporal lobe epilepsy: A review of the literature. J. Neurol. Sci. 2017, 381, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Tai, X.Y.; Bernhardt, B.; Thom, M.; Thompson, P.; Baxendale, S.; Koepp, M.; Bernasconi, N. Review: Neurodegenerative processes in temporal lobe epilepsy with hippocampal sclerosis: Clinical, pathological and neuroimaging evidence. Neuropathol. Appl. Neurobiol. 2018, 44, 70–90. [Google Scholar] [CrossRef]
- Vaden, J.H.; Banumurthy, G.; Gusarevich, E.S.; Overstreet-Wadiche, L.; Wadiche, J.I. The readily-releasable pool dynamically regulates multivesicular release. Elife 2019, 31, e47434. [Google Scholar] [CrossRef]
- Rollenhagen, A.; Klook, K.; Sätzler, K.; Qi, G.; Anstötz, M.; Feldmeyer, D.; Lübke, J.H.R. Structural determinants underlying the high efficacy of synaptic transmission and plasticity at synaptic boutons in layer 4 of the adult rat ‘barrel cortex’. Brain Struct. Funct. 2015, 220, 3185–3209. [Google Scholar] [CrossRef]
- Rollenhagen, A.; Ohana, O.; Sätzler, K.; Hilgetag, C.C.; Kuhl, D.; Lübke, J.H.R. Structural properties of synaptic transmission and temporal dynamics at excitatory layer 5b synapses in the adult rat somatosensory cortex. Front. Synaptic Neurosci. 2018, 10, 24. [Google Scholar] [CrossRef] [PubMed]
- Knott, G.; Holtmaat, A. Dendritic spine plasticity—Current understanding from in vivo studies. Brain Res. Rev. 2008, 58, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Verstreken, P.; Ly, C.V.; Venken, K.J.; Koh, T.W.; Zhou, Y.; Bellen, H.J. Synaptic mitochondria are critical for mobilization of reserve pool vesicles at Drosophila neuromuscular junctions. Neuron 2005, 47, 365–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verstreken, P.; Ohyama, T.; Bellen, H.J. FM 1-43 labeling of synaptic vesicle pools at the Drosophila neuromuscular junction. Methods Mol. Biol. 2008, 440, 349–369. [Google Scholar] [PubMed] [Green Version]
- Min, R.; Nevian, T. Astrocyte signaling controls spike timing-dependent depression at neocortical synapses. Nat. Neurosci. 2012, 15, 746–753. [Google Scholar] [CrossRef] [PubMed]
- Bopp, R.; Holler-Rickauer, S.; Martin, K.A.C.; Schuhknecht, G.F. An ultrastructural study of the thalamic input to layer 4 of primary motor and primary somatosensory cortex in the mouse. J. Neurosci. 2017, 37, 2435–2448. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Moreno, J.; Rollenhagen, A.; Arlandis, J.; Santuy, A.; Merchán-Peréz, A.; DeFelipe, J.; Lübke, J.H.R.; Clascà, F. Quantitative 3D ultrastructure of thalamocortical synapses from the “Lemniscal” ventral posteromedial nucleus in mouse barrel cortex. Cereb. Cortex 2018, 28, 3159–3175. [Google Scholar] [CrossRef]
- Hsu, A.; Luebke, J.I.; Medalla, M. Comparative ultrastructural features of excitatory synapses in the visual and frontal cortices of the adult mouse and monkey. J. Comp. Neurol. 2017, 525, 2175–2191. [Google Scholar] [CrossRef]
- Medalla, M.; Luebke, J.I. Diversity of glutamatergic synaptic strength in lateral prefrontal versus primary visual cortices in the rhesus monkey. J. Neurosci. 2015, 35, 112–127. [Google Scholar] [CrossRef] [Green Version]
- Sätzler, K.; Söhl, L.F.; Bollmann, J.H.; Borst, J.G.; Frotscher, M.; Sakmann, B.; Lübke, J.H.R. Three-Dimensional reconstruction of a calyx of Held and its postsynaptic principal neuron in the medial nucleus of the trapezoid body. J. Neurosci. 2002, 22, 10567–10579. [Google Scholar] [CrossRef]
- Nicol, M.J.; Walmsley, B. Ultrastructural basis of synaptic transmission between endbulbs of Held and bushy cells in the rat cochlear nucleus. J. Physiol. 2002, 539, 713–723. [Google Scholar] [CrossRef]
- Hoffpauir, B.K.; Grimes, J.L.; Mathers, P.H.; Spirou, G.A. Synaptogenesis of the calyx of Held: Rapid onset of function and one-to-one morphological innervation. J. Neurosci. 2006, 26, 5511–5523. [Google Scholar] [CrossRef] [PubMed]
- Xu-Friedman, M.A.; Regehr, W.G. Ultrastructural contributions to desensitization at cerebellar mossy fiber to granule cell synapses. J. Neurosci. 2003, 23, 2182–2192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamori, J.; Somogyi, J. Differentiation of cerebellar mossy fiber synapses in the rat: A quantitative electron microscope study. J. Comp. Neurol. 1983, 220, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Xu-Friedman, M.A.; Harris, K.M.; Regehr, W.G. Three-dimensional comparison of ultrastructural characteristics at depressing and facilitating synapses onto cerebellar Purkinje cells. J. Neurosci. 2001, 21, 6666–6672. [Google Scholar] [CrossRef] [Green Version]
- Rollenhagen, A.; Sätzler, K.; Rodriguez, E.P.; Jonas, P.; Frotscher, M.; Lübke, J.H.R. Structural determinants of transmission at large hippocampal mossy fiber synapses. J. Neurosci. 2007, 27, 10434–10444. [Google Scholar] [CrossRef] [Green Version]
- Chicurel, M.E.; Harris, K.M. Three-dimensional analysis of the structure and composition of CA3 branched dendritic spines and their synaptic relationships with mossy fiber boutons in the rat hippocampus. J. Comp. Neurol. 1992, 325, 169–182. [Google Scholar] [CrossRef]
- Henze, D.A.; Buzsaki, G. Action potential threshold of hippocampal pyramidal cells in vivo is increased by recent spiking activity. Neuroscience 2001, 105, 121–130. [Google Scholar] [CrossRef]
- Saviane, C.; Silver, R.A. Fast vesicle reloading and a large pool sustain high bandwidth transmission at a central synapse. Nature 2006, 439, 983–987. [Google Scholar] [CrossRef]
- Imig, C.; Min, S.W.; Krinner, S.; Arancillo, M.; Rosenmund, C.; Südhof, T.C.; Rhee, J.S.; Brose, N.; Cooper, B.H. The morphological and molecular nature of synaptic vesicle priming at presynaptic active zones. Neuron 2014, 84, 416–431. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Sugiura, Y.; Südhof, T.C.; Lin, W. Ablation of all synaptobrevin vsnares blocks evoked but not spontaneous neurotransmitter release at neuromuscular synapses. J. Neurosci. 2019, 39, 6049–6066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maus, L.; Lee, C.; Altas, B.; Sertel, S.M.; Weyand, K.; Rizzoli, S.O.; Rhee, J.S.; Brose, N.; Imig, C.; Cooper, B.H. Ultrastructural correlates of presynaptic functional heterogeneity in hippocampal synapses. Cell Rep. 2020, 30, 3632–3643. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Moreno, J.; Porrero, C.; Rollenhagen, A.; Rubio-Teves, M.; Casas-Torremocha, D.; Alonso-Nanclares, L.; Yakoubi, R.; Santuy, A.; Merchan-Pérez, A.; DeFelipe, J.; et al. Area-Specific synapse structure in branched posterior nucleus axons reveals a new level of complexity in thalamocortical networks. J. Neurosci. 2020, 40, 2663–2679. [Google Scholar] [CrossRef] [PubMed]
- Deerinck, T.J.; Bushong, E.; Thor, A.; Ellisman, M.H. NCMIR methods for 3D EM: A new protocol for preparation of biological specimens for serial block face scanning electron microscopy. Available online: https://ncmir.ucsd.edu/sbem-protocol (accessed on 2 August 2020).
- Nguyen, H.B.; Thai, T.Q.; Saitoh, S.; Wu, B.; Saitoh, Y.; Shimo, S.; Fujitani, H.; Otobe, H.; Ohno, N. Conductive resins improve charging and resolution of acquired images in electron microscopic volume imaging. Sci. Rep. 2016, 6, 23721. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.S.; Hayworth, K.J.; Lu, Z.; Grob, P.; Hassan, A.M.; Garcia-Cerdan, J.G.; Niyogi, K.K.; Nogales, E.; Weinberg, R.J.; Hess, H.F. Enhanced FIB-SEM systems for large-volume 3D imaging. Elife 2017, 6, e25916. [Google Scholar] [CrossRef]
- Borst, J.G.; Sakmann, B. Calcium current during a single action potential in a large presynaptic terminal of the rat brainstem. J. Physiol. 1998, 506, 143–157. [Google Scholar] [CrossRef]
- Bischofberger, J.; Geiger, J.R.; Jonas, P. Timing and efficacy of Ca2+ channel activation in hippocampal mossy fiber boutons. J. Neurosci. 2002, 22, 10593–10602. [Google Scholar] [CrossRef] [Green Version]
- Schneggenburger, R.; Neher, E. Intracellular calcium dependence of transmitter release rates at a fast central synapse. Nature 2000, 406, 889–893. [Google Scholar] [CrossRef]
- Helmchen, F.; Borst, J.G.; Sakmann, B. Calcium dynamics associated with a single action potential in a CNS presynaptic terminal. Biophys. J. 1997, 72, 1458–1471. [Google Scholar] [CrossRef] [Green Version]
- Neher, E. Merits and limitations of vesicle pool models in view of heterogeneous populations of synaptic vesicles. Neuron 2015, 87, 1131–1142. [Google Scholar] [CrossRef] [Green Version]
- Chamberland, S.; Tóth, K. Functionally heterogeneous synaptic vesicle pools support diverse synaptic signalling. J. Physiol. 2016, 594, 825–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meinrenken, C.J.; Borst, J.G.; Sakmann, B. Calcium secretion coupling at calyx of Held governed by nonuniform channel-vesicle topography. J. Neurosci. 2002, 22, 1648–1667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freche, D.; Pannasch, U.; Rouach, N.; Holcman, D. Synapse geometry and receptor dynamics modulate synaptic strength. PLoS ONE 2011, 6, e25122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzoli, S.O.; Betz, W.J. Synaptic vesicle pools. Nat. Rev. Neurosci. 2005, 6, 57–69. [Google Scholar] [CrossRef]
- Chiu, W.Z.; Donker Kaat, L.; Boon, A.J.W.; Kamphorst, W.; Schleicher, A.; Zilles, K.; Van Swieten, J.C.; Palomero-Gallagher, N. Multireceptor fingerprints in progressive supranuclear palsy. Alzheimer’s Res. Ther. 2017, 9, 28. [Google Scholar] [CrossRef] [Green Version]
- Palomero-Gallagher, N.; Kedo, O.; Mohlberg, H.; Zilles, K.; Amunts, K. Multimodal mapping and analysis of the cyto- and receptorarchitecture of the human hippocampus. Brain Struct. Funct. 2020, 225, 881–907. [Google Scholar] [CrossRef] [Green Version]
- Van den Heuvel, T.L.; Van der Eerden, A.W.; Manniesing, R.; Ghafoorian, M.; Tan, T.; Andriessen, T.M.; Van de Vyvere, T.; Van den Hauwe, L.; Ter Haar Romeny, B.M.; Goraj, B.M.; et al. Automated detection of cerebral microbleeds in patients with traumatic brain injury. Neuroimage Clin. 2016, 12, 241–251. [Google Scholar] [CrossRef] [Green Version]
- Beliveau, V.; Ganz, M.; Feng, L.; Ozenne, B.; Hojgaard, L.; Fisher, P.M.; Svarer, C.; Greve, D.N.; Knudsen, G.M. A high-resolution in vivo atlas of the human brain’s serotonin system. J. Neurosci. 2017, 37, 120–128. [Google Scholar] [CrossRef]
- Mohan, H.; Verhoog, M.B.; Doreswamy, K.K.; Eyal, G.; Aardse, R.; Lodder, B.N.; Goriounova, N.A.; Asamoah, B.; Groot, C.; Van der Sluis, S.; et al. Dendritic and axonal architecture of individual pyramidal neurons across layers of adult human neocortex. Cereb. Cortex 2015, 25, 4839–4853. [Google Scholar] [CrossRef]
- Reynolds, E.S. The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. J. Cell Biol. 1963, 17, 208–212. [Google Scholar] [CrossRef] [Green Version]
- Mastronarde, D.N. Automated electron microscope tomography using robust prediction of specimen movements. J. Struct. Biol. 2005, 152, 36–51. [Google Scholar] [CrossRef] [PubMed]
- Kremer, J.R.; Mastronarde, D.N.; McIntosh, J.R. Computer visualization of three-dimensional image data using IMOD. J. Struct. Biol. 1996, 116, 71–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species (Strain) | Mouse (C57/BL6)* | Mouse (C57/BL6)** | Mouse (C57/BL6)*** | Rat (Wistar) | Monkey (Macaca mulatta)*** | Human | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Region | L4_M1 | L4_S1 | L4_S1 | L2-3_V1 | L2-3_FC | L4_S1 | L5_S1 | L2-3_V1 | L2-3_LPFC | L4_TLN | L5_TLN |
| Age | — | — | 60–65 days | 2–14 month | 90–120 days | 5–20 years | 20–63 years | ||||
| Synaptic boutons | |||||||||||
| Surface area (µm2) | — | — | 4.67 ± 2.20 | — | — | 3.03 ± 0.71 | 8.19 ± 2.84 | — | — | 2.50 ± 1.78 | 6.09 ± 0.85 |
| Volume (µm3) | 0.17(+)/0.07(−) | 0.31(+)/0.07(−) | 0.46 ± 0.27 | 0.10 ± 0.01 | 0.08 ± 0.03 | 0.20 ± 0.07 | 0.38 ± 0.23 | 0.20 ± 0.04 | 0.30 ± 0.01 | 0.16 ± 0.16 | 0.63 ± 0.17 |
| Active zones | |||||||||||
| Number per bouton | 1.3(+)/1.1(−) | 2.1(+)/1.2(−) | 1.6 | — | — | 1.06 ± 0.06 | 1.12 ± 0.09 | — | — | 1–3 | 1–2 |
| PreAZ surface area (µm2) | — | — | — | — | — | 0.18 ± 0.06 | 0.29 ± 0.19 | — | — | 0.13 ± 0.07 | 0.23 ± 0.05 |
| PSD surface area (µm2) | 0.06(+)/0.56(−) | 0.04(+)/0.04(−) | 0.21 ± 0.11 | 0.08 ± 0.01 | 0.07 ± 0.02 | 0.18 ± 0.06 | 0.31 ± 0.21 | 0.08 ± 0.01 | 0.11 ± 0.01 | 0.13 ± 0.07 | 0.28 ± 0.11 |
| Cleft width (nm) | |||||||||||
| Lateral | — | — | — | — | — | 17.22 ± 1.50 | 15.52 ± 0.39 | — | — | 14.11 ± 0.69 | 17.24 ± 2.21 |
| Central | — | — | — | — | — | 30.22 ± 1.42 | 31.32 ± 1.81 | — | — | 16.47 ± 1.85 | 19.05 ± 2.72 |
| Synaptic vesicles (SVs) | |||||||||||
| Total number | 4846(+)/4861(−) | 5032(+)/5233(−) | 740 ± 285 | — | — | 561.00 ± 108.00 | 811.47 ± 272.25 | 337± 23 | 555 ± 48 | 1820.64 ± 980.34 | 1518.52 ± 303.18 |
| Pool size of SVs in: | |||||||||||
| Putative RRP at p10 nm | — | — | — | — | — | 1.97 ± 2.57 | 3.89 ± 3.35 | — | — | 20.20 ± 18.58 | 5.42 ± 4.09 |
| Putative RRP at p20 nm | — | — | — | — | — | 6.30 ± 6.40 | 11.55 ± 4.16 | — | — | 48.59 ± 39.02 | 15.21 ± 9.09 |
| Putative RP 60–200 nm | — | — | — | — | — | 130.16 ± 20.79 | 162.83 ± 56.37 | — | — | 382.10 ± 248.23 | 181.86 ± 24.20 |
| Putative resting pool >200 nm | — | — | — | — | — | 408.84 ± 100.04 | 599 ± 212.21 | — | — | 1251.82 ± 471.17 | 1264.05 ± 269.91 |
| Surface Area of SBs [µm2] | Number of AZs | Surface Area of AZs [µm2] | |
|---|---|---|---|
| Calyx of Held | 2500 | 554/*155 (85-217) | 0.10 ± 0.08/*0.06 ± 0.12 |
| Cerebellar Mossy Fiber | 69–200/**168–266 | 191–440/**113–176/**~300 | 0.04 ± 0.02 |
| Cerebellar Climbing Fiber | — | 67 | 0.14 ± 0.08 |
| Hippocampal Mossy Fiber | 150–1000 | 2–40 | 0.11 ± 0.07 |
| Experimental Procedure | Time | |
|---|---|---|
| TEM | FIB-SEM | |
| Sample preparation (fixation, vibratome sectioning, osmification, dehydration, embedding, polymerization | 120 h | 125 h |
| Serial sectioning | 3–5 h * | not applicable |
| Sputter coating, sample transfer into SEM + sample stabilization, deposition of protective metal pad, trench milling, and polishing of acquisition plane | not applicable | 5 h |
| Data acquisition (x–y–z dimension; number of x–y images per stack | 1–3 weeks ** | 24–30 h ** |
| Data post-processing (parameter adjustment, bright/contrast, stack alignment, potential cropping) | not applicable | 1 h |
| Total time | approximately 245–485 h ≅ 10–20 days | approximately 160 h ≅ 7 days |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rollenhagen, A.; Walkenfort, B.; Yakoubi, R.; Klauke, S.A.; Schmuhl-Giesen, S.F.; Heinen-Weiler, J.; Voortmann, S.; Marshallsay, B.; Palaz, T.; Holz, U.; et al. Synaptic Organization of the Human Temporal Lobe Neocortex as Revealed by High-Resolution Transmission, Focused Ion Beam Scanning, and Electron Microscopic Tomography. Int. J. Mol. Sci. 2020, 21, 5558. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155558
Rollenhagen A, Walkenfort B, Yakoubi R, Klauke SA, Schmuhl-Giesen SF, Heinen-Weiler J, Voortmann S, Marshallsay B, Palaz T, Holz U, et al. Synaptic Organization of the Human Temporal Lobe Neocortex as Revealed by High-Resolution Transmission, Focused Ion Beam Scanning, and Electron Microscopic Tomography. International Journal of Molecular Sciences. 2020; 21(15):5558. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155558
Chicago/Turabian StyleRollenhagen, Astrid, Bernd Walkenfort, Rachida Yakoubi, Sarah A. Klauke, Sandra F. Schmuhl-Giesen, Jacqueline Heinen-Weiler, Sylvia Voortmann, Brigitte Marshallsay, Tayfun Palaz, Ulrike Holz, and et al. 2020. "Synaptic Organization of the Human Temporal Lobe Neocortex as Revealed by High-Resolution Transmission, Focused Ion Beam Scanning, and Electron Microscopic Tomography" International Journal of Molecular Sciences 21, no. 15: 5558. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155558