Consequences of Vitamin A Deficiency: Immunoglobulin Dysregulation, Squamous Cell Metaplasia, Infectious Disease, and Death

 , ,
, ,
Abstract
:1. Introduction
2. Results
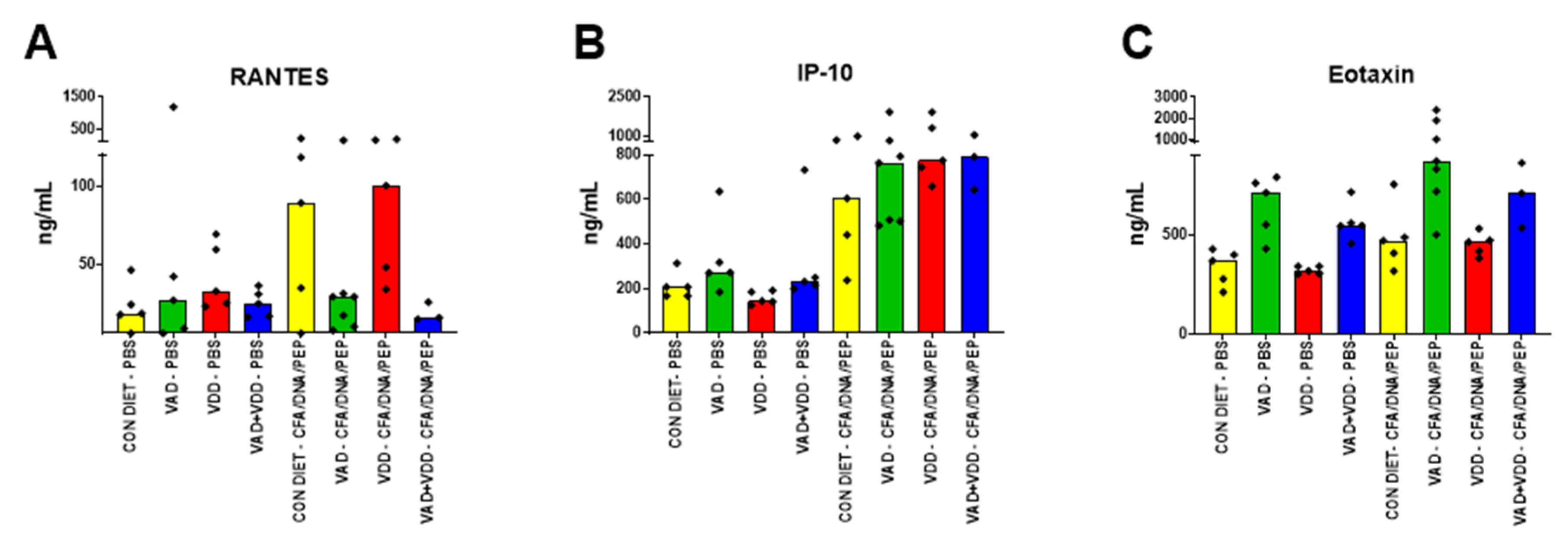
2.1. Abnormal Immunoglobulin and Cytokine/Chemokine Profiles in VAD and VAD+VDD Mice
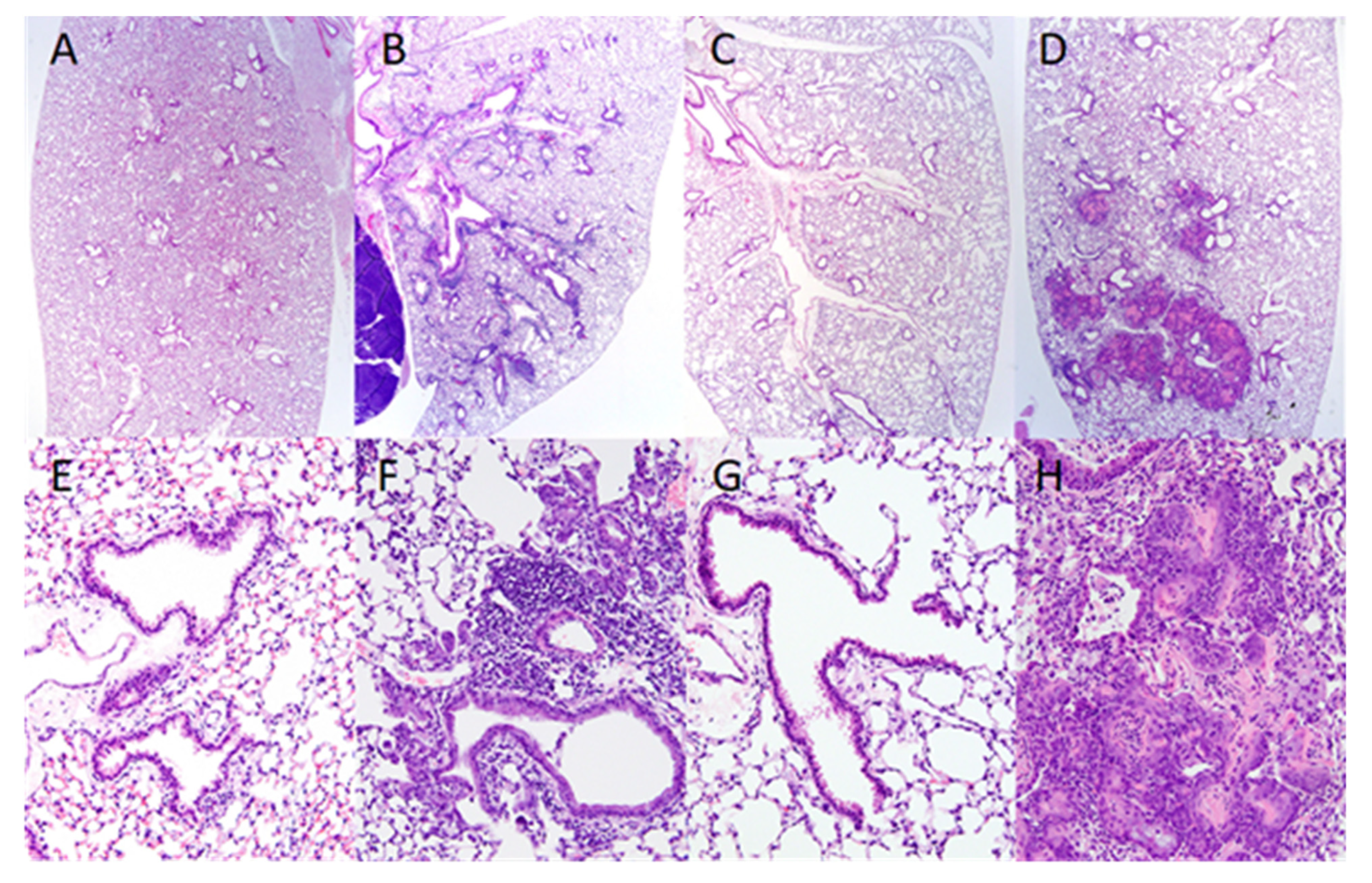
2.2. Squamous Cell Metaplasia of the Renal Pelvis and Ascending Bacterial Urinary Tract Infections (UTI) in VAD and VAD+VDD Mice
2.3. Abnormal Epithelial Cell Barriers in the Airways of VAD Mice Infected with a Respiratory Virus
3. Discussion
3.1. VAD, Keratinizing Metaplasia, UTIs and Respiratory Tract Disease
3.2. Dysregulation of Adaptive and Innate Immune Parameters in VAD Mice
3.3. Nuclear Receptors and Cross-Regulation
3.4. Defining Cause-Effect Relationships In Vivo
3.5. A focus on the Cross Regulatory Signals of Nuclear Receptors in Clinical Studies
4. Materials and Methods
4.1. Animal Models
4.2. CFA or CFA/DNA/PEP Injections
4.3. SeV infections
4.4. Total Serum Immunoglobulin Analyses
4.5. Histology
4.6. Cytokines
4.7. Statistical Analyses
Author Contributions
Funding
Conflicts of Interest
References
- Hussey, G.D.; Klein, M. A randomized, controlled trial of vitamin A in children with severe measles. N. Engl. J. Med. 1990, 323, 160–164. [Google Scholar] [CrossRef]
- Andersen, A.; Fisker, A.B.; Rodrigues, A.; Martins, C.; Ravn, H.; Lund, N.; Biering-Sorensen, S.; Benn, C.S.; Aaby, P. National Immunization Campaigns with Oral Polio Vaccine Reduce All-Cause Mortality: A Natural Experiment within Seven Randomized Trials. Front. Public Health 2018, 6, 13. [Google Scholar] [CrossRef]
- Vitamin A supplementation: Who, when and how. Community Eye Health 2013, 26, 71.
- Gilbert, C. What is vitamin A and why do we need it? Community Eye Health 2013, 26, 65. [Google Scholar] [PubMed]
- Sommer, A. Vitamin A, infectious disease, and childhood mortality: A 2 solution? J. Infect. Dis. 1993, 167, 1003–1007. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.G.; Oshansky, C.M.; Bajracharya, R.; Tang, L.; Sun, Y.; Wong, S.S.; Webby, R.; Thomas, P.G.; Hurwitz, J.L. Retinol binding protein and vitamin D associations with serum antibody isotypes, serum influenza virus-specific neutralizing activities and airway cytokine profiles. Clin. Exp. Immunol. 2016, 183, 239–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurwitz, J.L.; Jones, B.G.; Penkert, R.R.; Gansebom, S.; Sun, Y.; Tang, L.; Bramley, A.M.; Jain, S.; McCullers, J.A.; Arnold, S.R. Low Retinol-Binding Protein and Vitamin D Levels Are Associated with Severe Outcomes in Children Hospitalized with Lower Respiratory Tract Infection and Respiratory Syncytial Virus or Human Metapneumovirus Detection. J. Pediatr. 2017, 187, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Mora, J.R.; Iwata, M.; von Andrian, U.H. Vitamin effects on the immune system: Vitamins A and D take centre stage. Nat. Rev. Immunol. 2008, 8, 685–698. [Google Scholar] [CrossRef] [Green Version]
- Mora, J.R.; von Andrian, U.H. Retinoic acid: An educational “vitamin elixir” for gut-seeking T cells. Immunity 2004, 21, 458–460. [Google Scholar] [CrossRef] [Green Version]
- Penkert, R.R.; Jones, B.G.; Hacker, H.; Partridge, J.F.; Hurwitz, J.L. Vitamin A differentially regulates cytokine expression in respiratory epithelial and macrophage cell lines. Cytokine 2017, 91, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Evans, R.M.; Mangelsdorf, D.J. Nuclear Receptors, RXR, and the Big Bang. Cell 2014, 157, 255–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Luca, L.M. Retinoids and their receptors in differentiation, embryogenesis, and neoplasia. FASEB J. 1991, 5, 2924–2933. [Google Scholar] [CrossRef] [PubMed]
- Shaw, N.; Elholm, M.; Noy, N. Retinoic acid is a high affinity selective ligand for the peroxisome proliferator-activated receptor beta/delta. J. Biol. Chem. 2003, 278, 41589–41592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Napoli, J.L. Functions of Intracellular Retinoid Binding-Proteins. Subcell Biochem. 2016, 81, 21–76. [Google Scholar] [CrossRef] [Green Version]
- Hurwitz, J.L.; Penkert, R.R.; Xu, B.; Fan, Y.; Partridge, J.F.; Maul, R.W.; Gearhart, P.J. Hotspots for Vitamin-Steroid-Thyroid Hormone Response Elements Within Switch Regions of Immunoglobulin Heavy Chain Loci Predict a Direct Influence of Vitamins and Hormones on B Cell Class Switch Recombination. Viral Immunol. 2016, 29, 132–136. [Google Scholar] [CrossRef] [Green Version]
- Jones, B.G.; Sealy, R.E.; Penkert, R.R.; Surman, S.L.; Maul, R.W.; Neale, G.; Xu, B.; Gearhart, P.J.; Hurwitz, J.L. Complex sex-biased antibody responses: Estrogen receptors bind estrogen response elements centered within immunoglobulin heavy chain gene enhancers. Int. Immunol. 2018. [Google Scholar] [CrossRef]
- Jones, B.G.; Penkert, R.R.; Xu, B.; Fan, Y.; Neale, G.; Gearhart, P.J.; Hurwitz, J.L. Binding of estrogen receptors to switch sites and regulatory elements in the immunoglobulin heavy chain locus of activated B cells suggests a direct influence of estrogen on antibody expression. Mol. Immunol. 2016, 77, 97–102. [Google Scholar] [CrossRef] [Green Version]
- Sealy, R.E.; Jones, B.G.; Surman, S.L.; Penkert, R.R.; Pelletier, S.; Neale, G.; Hurwitz, J.L. Will Attention by Vaccine Developers to the Host’s Nuclear Hormone Levels and Immunocompetence Improve Vaccine Success? Vaccines (Basel) 2019, 7, 26. [Google Scholar] [CrossRef] [Green Version]
- Stavnezer, J.; Schrader, C.E. IgH chain class switch recombination: Mechanism and regulation. J. Immunol. 2014, 193, 5370–5378. [Google Scholar] [CrossRef] [Green Version]
- Birshtein, B.K. Epigenetic Regulation of Individual Modules of the immunoglobulin heavy chain locus 3′ Regulatory Region. Front. Immunol. 2014, 5, 163. [Google Scholar] [CrossRef] [Green Version]
- Billiau, A.; Matthys, P. Modes of action of Freund’s adjuvants in experimental models of autoimmune diseases. J. Leukoc. Biol. 2001, 70, 849–860. [Google Scholar] [PubMed]
- Desai, D.D.; Krishnan, M.R.; Swindle, J.T.; Marion, T.N. Antigen-specific induction of antibodies against native mammalian DNA in nonautoimmune mice. J. Immunol. 1993, 151, 1614–1626. [Google Scholar] [PubMed]
- Desai, D.D.; Marion, T.N. Induction of anti-DNA antibody with DNA-peptide complexes. Int. Immunol. 2000, 12, 1569–1578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnan, M.R.; Wang, C.; Marion, T.N. Anti-DNA autoantibodies initiate experimental lupus nephritis by binding directly to the glomerular basement membrane in mice. Kidney Int. 2012, 82, 184–192. [Google Scholar] [CrossRef] [Green Version]
- Minano, F.J.; Fernandez-Alonso, A.; Benamar, K.; Myers, R.K.; Sancibrian, M.; Ruiz, R.M.; Armengol, J.A. Macrophage inflammatory protein-1beta (MIP-1beta) produced endogenously in brain during E. coli fever in rats. Eur. J. Neurosci. 1996, 8, 424–428. [Google Scholar] [CrossRef]
- Heslop, R.; Bojang, A.L.; Jarju, S.; Mendy, J.; Mulwa, S.; Secka, O.; Mendy, F.S.; Owolabi, O.; Kampmann, B.; Sutherland, J.S. Changes in Host Cytokine Patterns of TB Patients with Different Bacterial Loads Detected Using 16S rRNA Analysis. PLoS ONE 2016, 11, e0168272. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, Y.; Klein, T.W.; Friedman, H. Induction of cytokine granulocyte-macrophage colony-stimulating factor and chemokine macrophage inflammatory protein 2 mRNAs in macrophages by Legionella pneumophila or Salmonella typhimurium attachment requires different ligand-receptor systems. Infect. Immun. 1996, 64, 3062–3068. [Google Scholar] [CrossRef] [Green Version]
- Allahverdian, S.; Harada, N.; Singhera, G.K.; Knight, D.A.; Dorscheid, D.R. Secretion of IL-13 by airway epithelial cells enhances epithelial repair via HB-EGF. Am. J. Respir. Cell Mol. Biol. 2008, 38, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Bucco, R.A.; Zheng, W.L.; Davis, J.T.; Sierra-Rivera, E.; Osteen, K.G.; Chaudhary, A.K.; Ong, D.E. Cellular retinoic acid-binding protein(II) presence in rat uterine epithelial cells correlates with their synthesis of retinoic acid. Biochemistry 1997, 36, 4009–4014. [Google Scholar] [CrossRef]
- Rudraraju, R.; Jones, B.G.; Surman, S.L.; Sealy, R.E.; Thomas, P.G.; Hurwitz, J.L. Respiratory tract epithelial cells express retinaldehyde dehydrogenase ALDH1A and enhance IgA production by stimulated B cells in the presence of vitamin A. PLoS ONE 2014, 9, e86554. [Google Scholar] [CrossRef] [Green Version]
- Nabeyrat, E.; Besnard, V.; Corroyer, S.; Cazals, V.; Clement, A. Retinoic acid-induced proliferation of lung alveolar epithelial cells: Relation with the IGF system. Am. J. Physiol. 1998, 275, L71–L79. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.H.; Gaffar, A.; Alamgir, S.M. Xerophthalmia, protein-calorie malnutrition, and infections in children. J. Pediatr. 1979, 95, 651–656. [Google Scholar] [CrossRef]
- Chandra, R.K. Increased bacterial binding to respiratory epithelial cells in vitamin A deficiency. Bmj 1988, 297, 834–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackfan, K.D.; Wohlbach, S.B. Vitamin A deficiency in infants: A clinical and pathological study. J. Pediatr. 1933, 3, 679–706. [Google Scholar] [CrossRef]
- Pinnock, C. Vitamin A. Nurs. J. India 1991, 82, 307–308. [Google Scholar]
- Sommer, A. Mortality associated with mild, untreated xerophthalmia. Trans. Am. Ophthalmol. Soc. 1983, 81, 825–853. [Google Scholar]
- Sommer, A.; Katz, J.; Tarwotjo, I. Increased risk of respiratory disease and diarrhea in children with preexisting mild vitamin A deficiency. Am. J. Clin. Nutr. 1984, 40, 1090–1095. [Google Scholar] [CrossRef]
- Bloch, C.E. Further clinical investigations into the diseases arising in consequence of a deficiency in the fat-soluble A factor. Am. J. Dis. Child. 1924, 659–667. [Google Scholar] [CrossRef]
- Kavukcu, S.; Soylu, A.; Turkmen, M.; Sarioglu, S.; Buyukgebiz, B.; Gure, A. The role of vitamin A in preventing renal scarring secondary to pyelonephritis. BJU Int. 1999, 83, 1055–1059. [Google Scholar] [CrossRef]
- Shaikh, N.; Ewing, A.L.; Bhatnagar, S.; Hoberman, A. Risk of renal scarring in children with a first urinary tract infection: A systematic review. Pediatrics 2010, 126, 1084–1091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neveus, T. Can postpyelonephritic renal scarring be prevented? Pediatr. Nephrol. 2013, 28, 187–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faust, W.C.; Diaz, M.; Pohl, H.G. Incidence of post-pyelonephritic renal scarring: A meta-analysis of the dimercapto-succinic acid literature. J. Urol. 2009, 181, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Benador, D.; Benador, N.; Slosman, D.; Mermillod, B.; Girardin, E. Are younger children at highest risk of renal sequelae after pyelonephritis? Lancet 1997, 349, 17–19. [Google Scholar] [CrossRef]
- Jakobsson, B.; Berg, U.; Svensson, L. Renal scarring after acute pyelonephritis. Arch. Dis. Child. 1994, 70, 111–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kavukcu, S.; Turkmen, M.; Sevinc, N.; Soylu, A.; Derebek, E.; Buyukgebiz, B. Serum vitamin A and beta-carotene concentrations and renal scarring in urinary tract infections. Arch. Dis. Child. 1998, 78, 271–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kavukcu, S.; Turkmen, M.A.; Soylu, A. Could the effective mechanisms of retinoids on nephrogenesis be also operative on the amelioration of injury in acquired renal lesions? Pediatr. Nephrol. 2001, 16, 689–690. [Google Scholar] [CrossRef]
- Yilmaz, A.; Bahat, E.; Yilmaz, G.G.; Hasanoglu, A.; Akman, S.; Guven, A.G. Adjuvant effect of vitamin A on recurrent lower urinary tract infections. Pediatr. Int. 2007, 49, 310–313. [Google Scholar] [CrossRef]
- Wagner, J. Potential role of retinoids in the therapy of renal disease. Nephrol. Dial. Transplant. 2001, 16, 441–444. [Google Scholar] [CrossRef] [Green Version]
- Sobouti, B.; Hooman, N.; Movahed, M. The effect of vitamin E or vitamin A on the prevention of renal scarring in children with acute pyelonephritis. Pediatr. Nephrol. 2013, 28, 277–283. [Google Scholar] [CrossRef]
- Bennett, R.T.; Mazzaccaro, R.J.; Chopra, N.; Melman, A.; Franco, I. Suppression of renal inflammation with vitamins A and E in ascending pyelonephritis in rats. J. Urol. 1999, 161, 1681–1684. [Google Scholar] [CrossRef]
- Ayazi, P.; Moshiri, S.A.; Mahyar, A.; Moradi, M. The effect of vitamin A on renal damage following acute pyelonephritis in children. Eur. J. Pediatr. 2011, 170, 347–350. [Google Scholar] [CrossRef] [PubMed]
- Flores-Mireles, A.L.; Walker, J.N.; Caparon, M.; Hultgren, S.J. Urinary tract infections: Epidemiology, mechanisms of infection and treatment options. Nat. Rev. Microbiol. 2015, 13, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.M.; Wittenberg, J.F.; Bryan, G.T. Effect of avitaminosis A and hypervitaminosis A on urinary bladder carcinogenicity of N-(4-(5-Nitro-2-furyl)-2-thiazolyl)formamide. Cancer Res. 1976, 36, 2334–2339. [Google Scholar] [PubMed]
- Stephensen, C.B.; Blount, S.R.; Schoeb, T.R.; Park, J.Y. Vitamin A deficiency impairs some aspects of the host response to influenza A virus infection in BALB/c mice. J. Nutr. 1993, 123, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Shields, P.A.; Jeffery, P.K. The combined effects of vitamin A-deficiency and cigarette smoke on rat tracheal epithelium. Br. J. Exp. Pathol. 1987, 68, 705–717. [Google Scholar] [PubMed]
- Penkert, R.R.; Surman, S.L.; Jones, B.G.; Sealy, R.E.; Vogel, P.; Neale, G.; Hurwitz, J.L. Vitamin A deficient mice exhibit increased viral antigens and enhanced cytokine/chemokine production in nasal tissues following respiratory virus infection despite the presence of FoxP3+ T cells. Int. Immunol. 2016, 28, 139–152. [Google Scholar] [CrossRef]
- Barsky, S.H.; Roth, M.D.; Kleerup, E.C.; Simmons, M.; Tashkin, D.P. Histopathologic and molecular alterations in bronchial epithelium in habitual smokers of marijuana, cocaine, and/or tobacco. J. Natl. Cancer Inst. 1998, 90, 1198–1205. [Google Scholar] [CrossRef] [Green Version]
- Mathe, G.; Gouveia, J.; Hercend, T.; Gros, F.; Dorval, T.; Hazon, J.; Misset, J.L.; Schwarzenberg, L.; Ribaud, P.; Lemaigre, G.; et al. Correlation between precancerous bronchial metaplasia and cigarette consumption, and preliminary results of retinoid treatment. Cancer Detect. Prev. 1982, 5, 461–466. [Google Scholar]
- Peters, E.J.; Morice, R.; Benner, S.E.; Lippman, S.; Lukeman, J.; Lee, J.S.; Ro, J.Y.; Hong, W.K. Squamous metaplasia of the bronchial mucosa and its relationship to smoking. Chest 1993, 103, 1429–1432. [Google Scholar] [CrossRef]
- Wistuba, I.I.; Behrens, C.; Milchgrub, S.; Bryant, D.; Hung, J.; Minna, J.D.; Gazdar, A.F. Sequential molecular abnormalities are involved in the multistage development of squamous cell lung carcinoma. Oncogene 1999, 18, 643–650. [Google Scholar] [CrossRef] [Green Version]
- Giroux, V.; Rustgi, A.K. Metaplasia: Tissue injury adaptation and a precursor to the dysplasia-cancer sequence. Nat. Rev. Cancer 2017, 17, 594–604. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.Y.; Lotan, R. Retinoids and their receptors in cancer development and chemoprevention. Crit. Rev. Oncol. Hemat. 2002, 41, 41–55. [Google Scholar] [CrossRef]
- Wald, N.; Idle, M.; Boreham, J.; Bailey, A. Low serum-vitamin-A and subsequent risk of cancer. Preliminary results of a prospective study. Lancet 1980, 2, 813–815. [Google Scholar] [CrossRef]
- Willett, W.C. Vitamin A and lung cancer. Nutr. Rev. 1990, 48, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Shareck, M.; Rousseau, M.C.; Koushik, A.; Siemiatycki, J.; Parent, M.E. Inverse Association between Dietary Intake of Selected Carotenoids and Vitamin C and Risk of Lung Cancer. Front. Oncol. 2017, 7, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, N.; Su, X.; Wang, Z.; Dai, B.; Kang, J. Association of Dietary Vitamin A and beta-Carotene Intake with the Risk of Lung Cancer: A Meta-Analysis of 19 Publications. Nutrients 2015, 7, 9309–9324. [Google Scholar] [CrossRef]
- Bukhari, M.H.; Qureshi, S.S.; Niazi, S.; Asef, M.; Naheed, M.; Khan, S.A.; Chaudhry, N.A.; Tayyab, M.; Hasan, M. Chemotherapeutic/chemopreventive role of retinoids in chemically induced skin carcinogenesis in albino mice. Int. J. Dermatol. 2007, 46, 1160–1165. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Y.; Hill, J.; Kim, H.T.; Shen, Q.; Bissonnette, R.P.; Lamph, W.W.; Brown, P.H. The rexinoid, bexarotene, prevents the development of premalignant lesions in MMTV-erbB2 mice. Br. J. Cancer 2008, 98, 1380–1388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, R.C.; Kelloff, G.J.; Detrisac, C.J.; Steele, V.E.; Thomas, C.F.; Sigman, C.C. Chemoprevention of OH-BBN-induced bladder cancer in mice by oltipraz, alone and in combination with 4-HPR and DFMO. Anticancer Res. 1994, 14, 5–11. [Google Scholar] [PubMed]
- Pisano, C.; Vesci, L.; Fodera, R.; Ferrara, F.F.; Rossi, C.; De Cesare, M.; Zuco, V.; Pratesi, G.; Supino, R.; Zunino, F. Antitumor activity of the combination of synthetic retinoid ST1926 and cisplatin in ovarian carcinoma models. Ann. Oncol. 2007, 18, 1500–1505. [Google Scholar] [CrossRef]
- Shah, R.K.; Valdez, T.A.; Wang, Z.; Shapshay, S.M. Pulsed-dye laser and retinoic acid delay progression of oral squamous cell carcinoma: A murine model. Laryngoscope 2001, 111, 1203–1208. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wen, W.; Yi, Y.; Zhang, Z.; Lubet, R.A.; You, M. Preventive effects of bexarotene and budesonide in a genetically engineered mouse model of small cell lung cancer. Cancer Prev. Res. (Phila.) 2009, 2, 1059–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edelman, M.J.; Smith, R.; Hausner, P.; Doyle, L.A.; Kalra, K.; Kendall, J.; Bedor, M.; Bisaccia, S. Phase II trial of the novel retinoid, bexarotene, and gemcitabine plus carboplatin in advanced non-small-cell lung cancer. J. Clin. Oncol. 2005, 23, 5774–5778. [Google Scholar] [CrossRef] [PubMed]
- Peterlin, P.; Garnier, A.; Tissot, A.; Garandeau, C.; Houreau-Langlard, D.; Hourmant, M.; Vantyghem, S.; Bonnet, A.; Guillaume, T.; Bene, M.C.; et al. Successful treatment of acute promyelocytic leukemia with arsenic trioxide and all-trans retinoic acid in a double lung and kidney transplanted patient. Ann. Hematol. 2016, 95, 1737–1738. [Google Scholar] [CrossRef] [PubMed]
- Recchia, F.; Sica, G.; Candeloro, G.; Necozione, S.; Bisegna, R.; Bratta, M.; Rea, S. Beta-interferon, retinoids and ta/moxifen in metastatic breast cancer: Long-term follow-up of a phase II study. Oncol. Rep. 2009, 21, 1011–1016. [Google Scholar] [CrossRef] [Green Version]
- Colombo, N.; Formelli, F.; Cantu, M.G.; Parma, G.; Gasco, M.; Argusti, A.; Santinelli, A.; Montironi, R.; Cavadini, E.; Baglietto, L.; et al. A phase I-II preoperative biomarker trial of fenretinide in ascitic ovarian cancer. Cancer Epidemiol. Biomark. 2006, 15, 1914–1919. [Google Scholar] [CrossRef] [Green Version]
- Alpha-Tocopherol, Beta Carotene Cancer Prevention Study Group. The effect of vitamin E and beta carotene on the incidence of lung cancer and other cancers in male smokers. N. Engl. J. Med. 1994, 330, 1029–1035. [Google Scholar] [CrossRef]
- De Vries, N.; Van Zandwijk, N.; Pastorino, U. The EUROSCAN study: A progress report. Am. J. Otolaryngol. 1993, 14, 62–66. [Google Scholar] [CrossRef]
- Hennekens, C.H.; Buring, J.E.; Manson, J.E.; Stampfer, M.; Rosner, B.; Cook, N.R.; Belanger, C.; LaMotte, F.; Gaziano, J.M.; Ridker, P.M.; et al. Lack of effect of long-term supplementation with beta carotene on the incidence of malignant neoplasms and cardiovascular disease. N. Engl. J. Med. 1996, 334, 1145–1149. [Google Scholar] [CrossRef] [Green Version]
- Omenn, G.S. Chemoprevention of lung cancers: Lessons from CARET, the beta-carotene and retinol efficacy trial, and prospects for the future. Eur. J. Cancer Prev. 2007, 16, 184–191. [Google Scholar] [CrossRef]
- Satia, J.A.; Littman, A.; Slatore, C.G.; Galanko, J.A.; White, E. Long-term use of beta-carotene, retinol, lycopene, and lutein supplements and lung cancer risk: Results from the VITamins And Lifestyle (VITAL) study. Am. J. Epidemiol. 2009, 169, 815–828. [Google Scholar] [CrossRef] [PubMed]
- Omenn, G.S.; Goodman, G.E.; Thornquist, M.D.; Balmes, J.; Cullen, M.R.; Glass, A.; Keogh, J.P.; Meyskens, F.L.; Valanis, B.; Williams, J.H.; et al. Effects of a combination of beta carotene and vitamin A on lung cancer and cardiovascular disease. N. Engl. J. Med. 1996, 334, 1150–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, M.E.; Jacobs, E.T.; Baron, J.A.; Marshall, J.R.; Byers, T. Dietary supplements and cancer prevention: Balancing potential benefits against proven harms. J. Natl. Cancer Inst. 2012, 104, 732–739. [Google Scholar] [CrossRef] [PubMed]
- Druesne-Pecollo, N.; Latino-Martel, P.; Norat, T.; Barrandon, E.; Bertrais, S.; Galan, P.; Hercberg, S. Beta-carotene supplementation and cancer risk: A systematic review and metaanalysis of randomized controlled trials. Int. J. Cancer 2010, 127, 172–184. [Google Scholar] [CrossRef]
- Neuhouser, M.L.; Patterson, R.E.; Thornquist, M.D.; Omenn, G.S.; King, I.B.; Goodman, G.E. Fruits and vegetables are associated with lower lung cancer risk only in the placebo arm of the beta-carotene and retinol efficacy trial (CARET). Cancer Epidemiol. Biomark. Prev. 2003, 12, 350–358. [Google Scholar]
- Russell, R.M. The enigma of beta-carotene in carcinogenesis: What can be learned from animal studies. J. Nutr. 2004, 134, 262S–268S. [Google Scholar] [CrossRef]
- Cerdan, C.; Martin, Y.; Brailly, H.; Courcoul, M.; Flavetta, S.; Costello, R.; Mawas, C.; Birg, F.; Olive, D. IL-1 alpha is produced by T lymphocytes activated via the CD2 plus CD28 pathways. J. Immunol. 1991, 146, 560–564. [Google Scholar]
- Culley, F.J.; Pennycook, A.M.; Tregoning, J.S.; Dodd, J.S.; Walzl, G.; Wells, T.N.; Hussell, T.; Openshaw, P.J. Role of CCL5 (RANTES) in viral lung disease. J. Virol. 2006, 80, 8151–8157. [Google Scholar] [CrossRef] [Green Version]
- Rammal, A.; Tewfik, M.; Rousseau, S. Differences in RANTES and IL-6 levels among chronic rhinosinusitis patients with predominant gram-negative and gram-positive infection. J. Otolaryngol. Head Neck Surg. 2017, 46, 7. [Google Scholar] [CrossRef] [Green Version]
- Rudraraju, R.; Surman, S.L.; Jones, B.G.; Sealy, R.; Woodland, D.L.; Hurwitz, J.L. Reduced frequencies and heightened CD103 expression among virus-induced CD8(+) T cells in the respiratory tract airways of vitamin A-deficient mice. Clin. Vaccine Immunol. 2012, 19, 757–765. [Google Scholar] [CrossRef]
- Crawford, A.; Angelosanto, J.M.; Nadwodny, K.L.; Blackburn, S.D.; Wherry, E.J. A role for the chemokine RANTES in regulating CD8 T cell responses during chronic viral infection. PLoS Pathog. 2011, 7, e1002098. [Google Scholar] [CrossRef] [PubMed]
- Surman, S.L.; Jones, B.G.; Rudraraju, R.; Sealy, R.E.; Hurwitz, J.L. Intranasal administration of retinyl palmitate with a respiratory virus vaccine corrects impaired mucosal IgA response in the vitamin A-deficient host. Clin. Vaccine Immunol. 2014, 21, 598–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surman, S.L.; Jones, B.G.; Sealy, R.E.; Rudraraju, R.; Hurwitz, J.L. Oral retinyl palmitate or retinoic acid corrects mucosal IgA responses toward an intranasal influenza virus vaccine in vitamin A deficient mice. Vaccine 2014, 32, 2521–2524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surman, S.L.; Penkert, R.R.; Jones, B.G.; Sealy, R.E.; Hurwitz, J.L. Vitamin supplementation at the time of immunization with cold-adapted influenza virus vaccine corrects poor antibody responses in mice deficient for vitamins A and D. Clin. Vaccine Immunol. 2016, 23, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Surman, S.L.; Rudraraju, R.; Sealy, R.; Jones, B.; Hurwitz, J.L. Vitamin A deficiency disrupts vaccine-induced antibody-forming cells and the balance of IgA/IgG isotypes in the upper and lower respiratory tract. Viral Immunol. 2012, 25, 341–344. [Google Scholar] [CrossRef]
- Jones, B.G.; Penkert, R.R.; Surman, S.L.; Sealy, R.E.; Pelletier, S.; Berns, H.; Hurwitz, J.L. Background check: Profound differences in serum antibody isotypes among C57BL/6 mouse substrains discourage substrain interchanges in immunology experiments. Immunol. Lett. 2019, 216, 9–11. [Google Scholar] [CrossRef]
- Jones, B.G.; Penkert, R.R.; Surman, S.L.; Sealy, R.E.; Pelletier, S.; Xu, B.; Neale, G.; Maul, R.W.; Gearhart, P.J.; Hurwitz, J.L. Matters of life and death: How estrogen and estrogen receptor binding to the immunoglobulin heavy chain locus may influence outcomes of infection, allergy, and autoimmune disease. Cell. Immunol. 2019, 346, 103996. [Google Scholar] [CrossRef]
- Jones, B.G.; Sealy, R.E.; Penkert, R.R.; Surman, S.L.; Birshtein, B.K.; Xu, B.; Neale, G.; Maul, R.W.; Gearhart, P.J.; Hurwitz, J.L. From Influenza Virus Infections to Lupus: Synchronous Estrogen Receptor alpha and RNA Polymerase II Binding Within the Immunoglobulin Heavy Chain Locus. Viral Immunol. 2020, 33, 307–315. [Google Scholar] [CrossRef]
- Patel, N.; Penkert, R.R.; Jones, B.G.; Sealy, R.E.; Surman, S.L.; Sun, Y.; Tang, L.; DeBeauchamp, J.; Webb, A.; Richardson, J.; et al. Baseline Serum Vitamin A and D Levels Determine Benefit of Oral Vitamin A&D Supplements to Humoral Immune Responses Following Pediatric Influenza Vaccination. Viruses 2019, 11, 907. [Google Scholar] [CrossRef] [Green Version]
- Bresee, J.S.; Fischer, M.; Dowell, S.F.; Johnston, B.D.; Biggs, V.M.; Levine, R.S.; Lingappa, J.R.; Keyserling, H.L.; Petersen, K.M.; Bak, J.R.; et al. Vitamin A therapy for children with respiratory syncytial virus infection: A multicenter trial in the United States. Pediatr. Infect. Dis. J. 1996, 15, 777–782. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diets | Treatments | ||||||
|---|---|---|---|---|---|---|---|
| PBS | CFA | CFA/DNA/PEP | TOTAL | ||||
| Female | Male | Female | Male | Female | Male | All | |
| CONTROL | |||||||
| Fraction | 0/8 | 0/11 | 1/10 | 0/13 | 0/11 | 0/9 | 1/62 |
| Percent | 0% | 0% | 10% | 0% | 0% | 0% | 2% |
| VAD | |||||||
| Fraction | 3/5 | 7/11 | 6/8 | 2/7 | 4/7 | 7/10 | 29/48 |
| Percent | 60% | 64% | 75% | 29% | 57% | 70% | 60% |
| VDD | |||||||
| Fraction | 1/12 | 0/12 | 0/10 | 0/14 | 0/7 | 0/14 | 1/69 |
| Percent | 8% | 0% | 0% | 0% | 0% | 0% | 1% |
| VAD+VDD | |||||||
| Fraction | 0/5 | 6/9 | 7/8 | 4/6 | 3/3 | 6/8 | 26/39 |
| Percent | 0% | 67% | 88% | 67% | 100% | 75% | 67% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Surman, S.L.; Penkert, R.R.; Sealy, R.E.; Jones, B.G.; Marion, T.N.; Vogel, P.; Hurwitz, J.L. Consequences of Vitamin A Deficiency: Immunoglobulin Dysregulation, Squamous Cell Metaplasia, Infectious Disease, and Death. Int. J. Mol. Sci. 2020, 21, 5570. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155570
Surman SL, Penkert RR, Sealy RE, Jones BG, Marion TN, Vogel P, Hurwitz JL. Consequences of Vitamin A Deficiency: Immunoglobulin Dysregulation, Squamous Cell Metaplasia, Infectious Disease, and Death. International Journal of Molecular Sciences. 2020; 21(15):5570. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155570
Chicago/Turabian StyleSurman, Sherri L., Rhiannon R. Penkert, Robert E. Sealy, Bart G. Jones, Tony N. Marion, Peter Vogel, and Julia L. Hurwitz. 2020. "Consequences of Vitamin A Deficiency: Immunoglobulin Dysregulation, Squamous Cell Metaplasia, Infectious Disease, and Death" International Journal of Molecular Sciences 21, no. 15: 5570. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155570