Extracellular ATP as an Inter-Kingdom Signaling Molecule: Release Mechanisms by Bacteria and Its Implication on the Host


{kind=link}
Abstract
:1. Principles of Purinergic Signaling
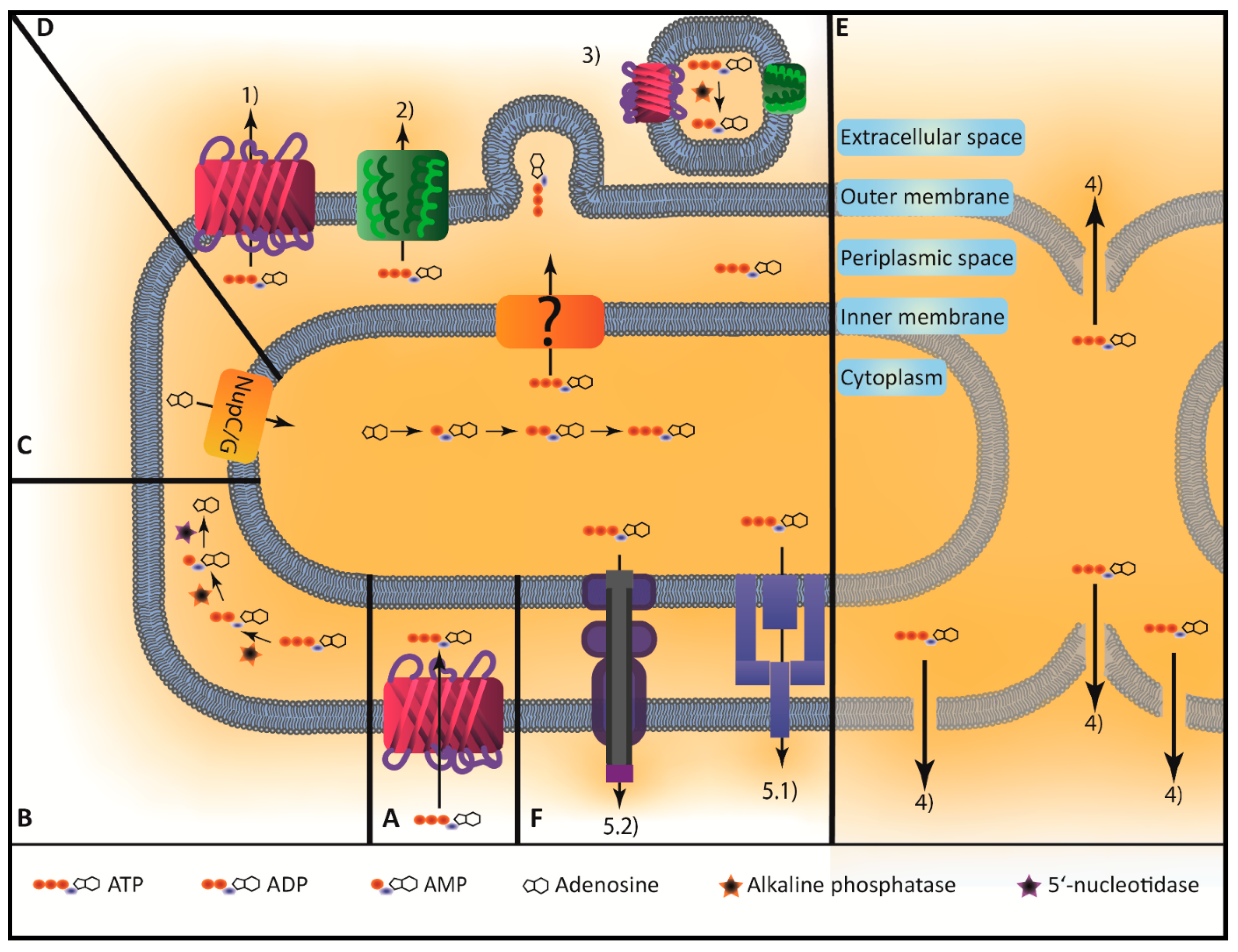
2. Secretion and Uptake of ATP by Microorganisms
2.1. Bacterial eATP under Homeostatic Conditions
2.2. Bacterial eATP During Growth
2.3. Uptake of eATP by Bacteria and Intracellular Fate
3. Comparison of ATP Secretion Mechanisms between Prokaryotic and Eukaryotic Cells
3.1. Release Mechanisms of Eukaryotic Cells
3.2. Release Mechanisms of Prokaryotic Cells
4. Extracellular ATP as an Inter-Kingdom Signaling Molecule
5. Bacterial ATP as Potential Virulence Factor in Inflammation and Sepsis
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ADP | Adenosine 5′-diphosphate |
| AMP | Adenosine 5′-monophosphate |
| ATP | Adenosine 5′-triphosphate |
| BHI | Brain heart infusion |
| cAMP | 3′:5′-cyclic adenosine monophosphate |
| DAMP | Danger associated molecular pattern |
| eATP | Extracellular adenosine 5′-triphosphate |
| ICAM-1 | Intercellular adhesion molecule 1 |
| LB | Luria–Bertani broth |
| MS | Mechanosensitive |
| OMV | Outer membrane vesicles |
| PAMP | Pathogen associated molecular pattern |
| PLCβ | Phospholipase C β |
| Tfh | Follicular T helper cells |
| TH17 | IL-17 producing helper T cells |
| TSA | Tryptic soy agar |
| UDP | Uridine 5′-biphosphate |
| UTP | Uridine 5′-triphosphate |
| VCAM-1 | Vascular cell adhesion molecule 1 |
| VNUT | Vesicular nucleotide transporter |
References
- Burnstock, G. Purinergic signalling. Br. J. Pharmacol. 2006, 147, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Venereau, E.; Ceriotti, C.; Bianchi, M.E. DAMPs from cell death to new life. Front. Immunol. 2015, 6, 422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagerberg, S.K.; Patel, P.; Andersen, L.W.; Lui, X.; Donnino, M.W.; Praetorius, H.A. Erythrocyte P2X1 receptor expression is correlated with change in haematocrit in patients admitted to the ICU with blood pathogen-positive sepsis. Crit. Care 2018, 22, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Ouyang, Y.; Liu, B.; Ma, X.; Ding, R. Platelet activation and antiplatelet therapy in sepsis: A narrative review. Thromb. Res. 2018, 166, 28–36. [Google Scholar] [CrossRef]
- Idzko, M.; Ferrari, D.; Eltzschig, H.K. Nucleotide signalling during inflammation. Nature 2014, 509, 310–317. [Google Scholar] [CrossRef] [Green Version]
- Eltzschig, H.K.; Sitkovsky, M.V.; Robson, S.C. Purinergic signaling during inflammation. New Engl. J. Med. 2012, 367, 2322–2333. [Google Scholar] [CrossRef] [Green Version]
- Ledderose, C.; Bao, Y.; Kondo, Y.; Fakhari, M.; Slubowski, C.; Zhang, J.; Junger, W.G. Purinergic signaling and the immune response in sepsis: A review. Clin. Ther. 2016, 38, 1054–1065. [Google Scholar] [CrossRef] [Green Version]
- Burnstock, G. Historical review: ATP as a neurotransmitter. Trends Pharmacol. Sci. 2006, 27, 166–176. [Google Scholar] [CrossRef]
- Antonio, L.S.; Stewart, A.P.; Xu, X.J.; Varanda, W.A.; Lagnado, R.D.M.; Edwardson, J.M. P2X4 receptors interact with both P2X2 and P2X7 receptors in the form of homotrimers. Br. J. Pharmacol. 2011, 163, 1069–1077. [Google Scholar] [CrossRef] [Green Version]
- Burnstock, G. Introduction to the special issue on purinergic receptors. Adv. Exp. Med. Biol. 2017, 1051, 1–6. [Google Scholar] [CrossRef]
- Burnstock, G. P2X ion channel receptors and inflammation. Purinergic Signal. 2016, 12, 59–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junger, W.G. Immune cell regulation by autocrine purinergic signalling. Nat. Rev. Immunol. 2011, 11, 201–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonioli, L.; Pacher, P.; Vizi, E.S.; Hasko, G. CD39 and CD73 in immunity and inflammation. Trends Mol. Med. 2013, 19, 355–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasko, G.; Linden, J.; Cronstein, B.; Pacher, P. Adenosine receptors: Therapeutic aspects for inflammatory and immune diseases. Nat. Rev. Drug Discov. 2008, 7, 759–770. [Google Scholar] [CrossRef] [PubMed]
- Fredholm, B.B.; Adriaan, I.J.; Jacobson, K.A.; Linden, J.; Muller, C.E. International union of basic and clinical pharmacology. LXXXI. Nomenclature and classification of adenosine receptors--an update. Pharmacol. Rev. 2011, 63, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Cronstein, B.N.; Daguma, L.; Nichols, D.; Hutchison, A.J.; Williams, M. The adenosine/neutrophil paradox resolved: Human neutrophils possess both A1 and A2 receptors that promote chemotaxis and inhibit O2 generation, respectively. J. Clin. Investig. 1990, 85, 1150–1157. [Google Scholar] [CrossRef]
- Ohta, A.; Sitkovsky, M. Role of G-protein-coupled adenosine receptors in downregulation of inflammation and protection from tissue damage. Nature 2001, 414, 916–920. [Google Scholar] [CrossRef]
- Wallace, K.L.; Linden, J. Adenosine A2A receptors induced on iNKT and NK cells reduce pulmonary inflammation and injury in mice with sickle cell disease. Blood 2010, 116, 5010–5020. [Google Scholar] [CrossRef] [Green Version]
- Cauwels, A.; Rogge, E.; Vandendriessche, B.; Shiva, S.; Brouckaert, P. Extracellular ATP drives systemic inflammation, tissue damage and mortality. Cell Death Dis. 2014, 5, e1102. [Google Scholar] [CrossRef] [Green Version]
- Cekic, C.; Linden, J. Purinergic regulation of the immune system. Nat. Rev. Immunol. 2016, 16, 177–192. [Google Scholar] [CrossRef]
- Csoka, B.; Nemeth, Z.H.; Toro, G.; Koscso, B.; Kokai, E.; Robson, S.C.; Enjyoji, K.; Rolandelli, R.H.; Erdelyi, K.; Pacher, P.; et al. CD39 improves survival in microbial sepsis by attenuating systemic inflammation. FASEB J. Publ. Fed. Am. Soc. Exp. Biol. 2015, 29, 25–36. [Google Scholar] [CrossRef] [Green Version]
- Fredholm, B.B.; Abbracchio, M.P.; Burnstock, G.; Daly, J.W.; Harden, T.K.; Jacobson, K.A.; Leff, P.; Williams, M. Nomenclature and classification of purinoceptors. Pharmacol. Rev. 1994, 46, 143–156. [Google Scholar]
- Fountain, S.J. Primitive ATP-activated P2X receptors: Discovery, function and pharmacology. Front. Cell. Neurosci. 2013, 7, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koshlukova, S.E.; Lloyd, T.L.; Araujo, M.W.; Edgerton, M. Salivary histatin 5 induces non-lytic release of ATP from candida albicans leading to cell death. J. Biol. Chem. 1999, 274, 18872–18879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, X.; Malhotra, R.; Guidotti, G. ATP uptake in the Golgi and extracellular release require Mcd4 protein and the vacuolar H+-ATPase. J. Biol. Chem. 2003, 278, 33436–33444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyum, R.; Guidotti, G. Glucose-dependent, cAMP-mediated ATP efflux from saccharomyces cerevisiae. microbiology 1997, 143, 1901–1908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakubowski, H.; Goldman, E. Evidence for cooperation between cells during sporulation of the yeast saccharomyces cerevisiae. Mol. Cell. Biol. 1988, 8, 5166–5178. [Google Scholar] [CrossRef] [Green Version]
- Inami, A.; Kiyono, H.; Kurashima, Y. ATP as a Pathophysiologic mediator of bacteria-host crosstalk in the gastrointestinal tract. Int. J. Mol. Sci. 2018, 19, 2371. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, E.P.; Alexeeva, Y.V.; Pham, D.K.; Wright, J.P.; Nicolau, D.V. ATP level variations in heterotrophic bacteria during attachment on hydrophilic and hydrophobic surfaces. Int. Microbiol. J. Span. Soc. Microbiol. 2006, 9, 37–46. [Google Scholar]
- Dosch, M.; Zindel, J.; Jebbawi, F.; Melin, N.; Taltavull, D.S.; Stroka, D.; Candinas, D.; Beldi, G. Connexin-43-dependent ATP release mediates macrophage activation during sepsis. eLife 2019, 8. [Google Scholar] [CrossRef]
- Eltzschig, H.K.; Eckle, T.; Mager, A.; Kuper, N.; Karcher, C.; Weissmuller, T.; Boengler, K.; Schulz, R.; Robson, S.C.; Colgan, S.P. ATP release from activated neutrophils occurs via connexin 43 and modulates adenosine-dependent endothelial cell function. Circ. Res. 2006, 99, 1100–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lasko, D.R.; Wang, D.I. On-line monitoring of intracellular ATP concentration in escherichia coli fermentations. Biotechnol. Bioeng. 1996, 52, 364–372. [Google Scholar] [CrossRef]
- Takaine, M.; Ueno, M.; Kitamura, K.; Imamura, H.; Yoshida, S. Reliable imaging of ATP in living budding and fission yeast. J. Cell Sci. 2019, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaginuma, H.; Kawai, S.; Tabata, K.V.; Tomiyama, K.; Kakizuka, A.; Komatsuzaki, T.; Noji, H.; Imamura, H. Diversity in ATP concentrations in a single bacterial cell population revealed by quantitative single-cell imaging. Sci. Rep. 2014, 4, 6522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwase, T.; Shinji, H.; Tajima, A.; Sato, F.; Tamura, T.; Iwamoto, T.; Yoneda, M.; Mizunoe, Y. Isolation and identification of ATP-secreting bacteria from mice and humans. J. Clin. Microbiol. 2010, 48, 1949–1951. [Google Scholar] [CrossRef] [Green Version]
- Hironaka, I.; Iwase, T.; Sugimoto, S.; Okuda, K.; Tajima, A.; Yanaga, K.; Mizunoe, Y. Glucose triggers ATP secretion from bacteria in a growth-phase-dependent manner. Appl. Environ. Microbiol. 2013, 79, 2328–2335. [Google Scholar] [CrossRef] [Green Version]
- Mempin, R.; Tran, H.; Chen, C.; Gong, H.; Kim Ho, K.; Lu, S. Release of extracellular ATP by bacteria during growth. BMC Microbiol. 2013, 13, 301. [Google Scholar] [CrossRef] [Green Version]
- Bekker, M.; Vries, S.D.; Beek, A.T.; Hellingwerf, K.J.; Mattos, M.J.D. Respiration of escherichia coli can be fully uncoupled via the nonelectrogenic terminal cytochrome bd-II oxidase. J. Bacteriol. 2009, 191, 5510–5517. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Hellingwerf, K.J.; Mattos, M.J.D.; Bekker, M. Uncoupling of substrate-level phosphorylation in escherichia coli during glucose-limited growth. Appl. Environ. Microbiol. 2012, 78, 6908–6913. [Google Scholar] [CrossRef] [Green Version]
- Unden, G.; Steinmetz, P.A.; Dunnwald, P.D. The aerobic and anaerobic respiratory chain of escherichia coli and salmonella enterica: Enzymes and energetics. EcoSal Plus 2014, 6. [Google Scholar] [CrossRef]
- Watanabe, K.; Tomioka, S.; Tanimura, K.; Oku, H.; Isoi, K. Uptake of AMP, ADP, and ATP in escherichia coli W. Biosci. Biotechnol. Biochem. 2011, 75, 7–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez, C.L.; Corradi, G.; Lauri, N.; Freixa, I.M.; Denis, M.F.L.; Enrique, N.; Mate, S.M.; Milesi, V.; Ostuni, M.A.; Herlax, V.; et al. Dynamic regulation of extracellular ATP in escherichia coli. Biochem. J. 2017, 474, 1395–1416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakehi, M.; Usuda, Y.; Tabira, Y.; Sugimoto, S. Complete deficiency of 5′-nucleotidase activity in escherichia coli leads to loss of growth on purine nucleotides but not of their excretion. J. Mol. Microbiol. Biotechnol. 2007, 13, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Passariello, C.; Forleo, C.; Micheli, V.; Schippa, S.; Leone, R.; Mangani, S.; Thaller, M.C.; Rossolini, G.M. Biochemical characterization of the class B acid phosphatase (AphA) of escherichia coli MG1655. Biochim. Biophys. Acta 2006, 1764, 13–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thaller, M.C.; Schippa, S.; Bonci, A.; Cresti, S.; Rossolini, G.M. Identification of the gene (aphA) encoding the class B acid phosphatase/phosphotransferase of escherichia coli MG1655 and characterization of its product. FEMS Microbiol. Lett. 1997, 146, 191–198. [Google Scholar] [CrossRef]
- Proudfoot, M.; Kuznetsova, E.; Brown, G.; Rao, N.N.; Kitagawa, M.; Mori, H.; Savchenko, A.; Yakunin, A.F. General enzymatic screens identify three new nucleotidases in escherichia coli. biochemical characterization of SurE, YfbR, and YjjG. J. Biol. Chem. 2004, 279, 54687–54694. [Google Scholar] [CrossRef] [Green Version]
- Patching, S.G.; Baldwin, S.A.; Baldwin, A.D.; Young, J.D.; Gallagher, M.P.; Henderson, P.J.; Herbert, R.B. The nucleoside transport proteins, NupC and NupG, from escherichia coli: Specific structural motifs necessary for the binding of ligands. Org. Biomol. Chem. 2005, 3, 462–470. [Google Scholar] [CrossRef]
- Meyrat, A.; Ballmoos, C.V. ATP synthesis at physiological nucleotide concentrations. Sci. Rep. 2019, 9, 3070. [Google Scholar] [CrossRef] [PubMed]
- Dosch, M.; Gerber, J.; Jebbawi, F.; Beldi, G. Mechanisms of ATP release by inflammatory cells. Int. J. Mol. Sci. 2018, 19, 1222. [Google Scholar] [CrossRef] [Green Version]
- Chekeni, F.B.; Elliott, M.R.; Sandilos, J.K.; Walk, S.F.; Kinchen, J.M.; Lazarowski, E.R.; Armstrong, A.J.; Penuela, S.; Laird, D.W.; Salvesen, G.S.; et al. Pannexin 1 channels mediate ‘find-me’ signal release and membrane permeability during apoptosis. Nature 2010, 467, 863–867. [Google Scholar] [CrossRef] [Green Version]
- Saez, P.J.; Vargas, P.; Shoji, K.F.; Harcha, P.A.; Dumenil, A.M.L.; Saez, J.C. ATP promotes the fast migration of dendritic cells through the activity of pannexin 1 channels and P2X7 receptors. Sci. Signal. 2017, 10. [Google Scholar] [CrossRef] [Green Version]
- Woehrle, T.; Yip, L.; Elkhal, A.; Sumi, Y.; Chen, Y.; Yao, Y.; Insel, P.A.; Junger, W.G. Pannexin-1 hemichannel-mediated ATP release together with P2X1 and P2X4 receptors regulate T-cell activation at the immune synapse. Blood 2010, 116, 3475–3484. [Google Scholar] [CrossRef] [Green Version]
- Riteau, N.; Baron, L.; Villeret, B.; Guillou, N.; Savigny, F.; Ryffel, B.; Rassendren, F.; Bert, M.L.; Gombault, A.; Couillin, I. ATP release and purinergic signaling: A common pathway for particle-mediated inflammasome activation. Cell Death Dis. 2012, 3, e403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Qin, W.; Xu, X.; Xiong, Y.; Zhang, Y.; Zhang, H.; Sun, B. Endotoxin-induced autocrine ATP signaling inhibits neutrophil chemotaxis through enhancing myosin light chain phosphorylation. Proc. Natl. Acad. Sci. USA 2017, 114, 4483–4488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imura, Y.; Morizawa, Y.; Komatsu, R.; Shibata, K.; Shinozaki, Y.; Kasai, H.; Moriishi, K.; Moriyama, Y.; Koizumi, S. Microglia release ATP by exocytosis. Glia 2013, 61, 1320–1330. [Google Scholar] [CrossRef] [PubMed]
- Lazarowski, E.R. Vesicular and conductive mechanisms of nucleotide release. Purinergic Signal. 2012, 8, 359–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudhof, T.C.; Rothman, J.E. Membrane fusion: Grappling with SNARE and SM proteins. Science 2009, 323, 474–477. [Google Scholar] [CrossRef] [Green Version]
- Kato, Y.; Hiasa, M.; Ichikawa, R.; Hasuzawa, N.; Kadowaki, A.; Iwatsuki, K.; Shima, K.; Endo, Y.; Kitahara, Y.; Inoue, T.; et al. Identification of a vesicular ATP release inhibitor for the treatment of neuropathic and inflammatory pain. Proc. Natl. Acad. Sci. USA 2017. [Google Scholar] [CrossRef] [Green Version]
- Ren, H.; Teng, Y.; Tan, B.; Zhang, X.; Jiang, W.; Liu, M.; Jiang, W.; Du, B.; Qian, M. Toll-like receptor-triggered calcium mobilization protects mice against bacterial infection through extracellular ATP release. Infect. Immun. 2014, 82, 5076–5085. [Google Scholar] [CrossRef] [Green Version]
- Sakaki, H.; Tsukimoto, M.; Harada, H.; Moriyama, Y.; Kojima, S. Autocrine regulation of macrophage activation via exocytosis of ATP and activation of P2Y11 receptor. PLoS ONE 2013, 8, e59778. [Google Scholar] [CrossRef] [Green Version]
- Virgilio, F.D.; Vuerich, M. Purinergic signaling in the immune system. Auton. Neurosci. 2015, 191, 117–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.Z.; Nikaido, H. Efflux-mediated drug resistance in bacteria: An update. Drugs 2009, 69, 1555–1623. [Google Scholar] [CrossRef] [PubMed]
- Poole, K. Efflux-mediated antimicrobial resistance. J. Antimicrob. Chemother. 2005, 56, 20–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masi, M.; Refregiers, M.; Pos, K.M.; Pages, J.M. Mechanisms of envelope permeability and antibiotic influx and efflux in gram-negative bacteria. Nat. Microbiol. 2017, 2, 17001. [Google Scholar] [CrossRef]
- Piddock, L.J. Multidrug-resistance efflux pumps—Not just for resistance. Nat. Rev. Microbiol. 2006, 4, 629–636. [Google Scholar] [CrossRef]
- Costa, T.R.; Rodrigues, C.F.; Meir, A.; Prevost, M.S.; Redzej, A.; Trokter, M.; Waksman, G. Secretion systems in Gram-negative bacteria: Structural and mechanistic insights. Nat. Rev. Microbiol. 2015, 13, 343–359. [Google Scholar] [CrossRef]
- Green, E.R.; Mecsas, J. Bacterial secretion systems: An overview. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Benz, R.; Schmid, A.; Maier, C.; Bremer, E. Characterization of the nucleoside-binding site inside the Tsx channel of escherichia coli outer membrane. Reconstitution experiments with lipid bilayer membranes. Eur. J. Biochem. 1988, 176, 699–705. [Google Scholar] [CrossRef]
- Maier, C.; Bremer, E.; Schmid, A.; Benz, R. Pore-forming activity of the Tsx protein from the outer membrane of escherichia coli. demonstration of a nucleoside-specific binding site. J. Biol. Chem. 1988, 263, 2493–2499. [Google Scholar]
- Benz, R.; Schmid, A.; Nakae, T.; Scheperkeuter, G.H.V. Pore formation by LamB of escherichia coli in lipid bilayer membranes. J. Bacteriol. 1986, 165, 978–986. [Google Scholar] [CrossRef] [Green Version]
- Samartzidou, H.; Delcour, A.H. Distinct sensitivities of OmpF and PhoE porins to charged modulators. FEBS Lett. 1999, 444, 65–70. [Google Scholar] [CrossRef] [Green Version]
- Heller, K.B.; Wilson, T.H. Selectivity of the escherichia coli outer membrane porins ompC and ompF. FEBS Lett. 1981, 129, 253–255. [Google Scholar] [CrossRef] [Green Version]
- Gehring, K.B.; Nikaido, H. Existence and purification of porin heterotrimers of escherichia coli K12 OmpC, OmpF, and PhoE proteins. J. Biol. Chem. 1989, 264, 2810–2815. [Google Scholar] [PubMed]
- Booth, I.R.; Edwards, M.D.; Black, S.; Schumann, U.; Miller, S. Mechanosensitive channels in bacteria: Signs of closure? Nat. Rev. Microbiol. 2007, 5, 431–440. [Google Scholar] [CrossRef]
- Berrier, C.; Coulombe, A.; Szabo, I.; Zoratti, M.; Ghazi, A. Gadolinium ion inhibits loss of metabolites induced by osmotic shock and large stretch-activated channels in bacteria. Eur. J. Biochem. 1992, 206, 559–565. [Google Scholar] [CrossRef]
- Schwechheimer, C.; Kuehn, M.J. Outer-membrane vesicles from gram-negative bacteria: Biogenesis and functions. Nat. Rev. Microbiol. 2015, 13, 605–619. [Google Scholar] [CrossRef] [Green Version]
- Proietti, M.; Perruzza, L.; Scribano, D.; Pellegrini, G.; D’Antuono, R.; Strati, F.; Raffaelli, M.; Gonzalez, S.F.; Thelen, M.; Hardt, W.D.; et al. ATP released by intestinal bacteria limits the generation of protective IgA against enteropathogens. Nat. Commun. 2019, 10, 250. [Google Scholar] [CrossRef]
- Hartmann, R.; Hennig, S.B.B.; Schwarz, U. Murein hydrolases in the envelope of escherichia coli. properties in situ and solubilization from the envelope. Eur. J. Biochem. 1974, 41, 203–208. [Google Scholar] [CrossRef]
- Schwarz, U.; Asmus, A.; Frank, H. Autolytic enzymes and cell division of escherichia coli. J. Mol. Biol. 1969, 41, 419–429. [Google Scholar] [CrossRef]
- Leive, L. Studies on the permeability change produced in coliform bacteria by ethylenediaminetetraacetate. J. Biol. Chem. 1968, 243, 2373–2380. [Google Scholar]
- Leduc, M.; Heijenoort, J.V. Autolysis of escherichia coli. J. Bacteriol. 1980, 142, 52–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, A.L. Death of bacteria in growing culture. J. Bacteriol. 1959, 77, 623–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proietti, M.; Cornacchione, V.; Jost, T.R.; Romagnani, A.; Faliti, C.E.; Perruzza, L.; Rigoni, R.; Radaelli, E.; Caprioli, F.; Preziuso, S.; et al. ATP-gated ionotropic P2X7 receptor controls follicular T helper cell numbers in Peyer’s patches to promote host-microbiota mutualism. Immunity 2014, 41, 789–801. [Google Scholar] [CrossRef] [Green Version]
- Scribano, D.; Petrucca, A.; Pompili, M.; Ambrosi, C.; Bruni, E.; Zagaglia, C.; Prosseda, G.; Nencioni, L.; Casalino, M.; Polticelli, F.; et al. Polar localization of PhoN2, a periplasmic virulence-associated factor of Shigella flexneri, is required for proper IcsA exposition at the old bacterial pole. PLoS ONE 2014, 9, e90230. [Google Scholar] [CrossRef] [PubMed]
- Perruzza, L.; Gargari, G.; Proietti, M.; Fosso, B.; D’Erchia, A.M.; Faliti, C.E.; Jost, T.R.; Scribano, D.; Mauri, L.; Colombo, D.; et al. T follicular helper cells promote a beneficial gut ecosystem for host metabolic homeostasis by sensing microbiota-derived extracellular ATP. Cell Rep. 2017, 18, 2566–2575. [Google Scholar] [CrossRef] [Green Version]
- Atarashi, K.; Nishimura, J.; Shima, T.; Umesaki, Y.; Yamamoto, M.; Onoue, M.; Yagita, H.; Ishii, N.; Evans, R.; Honda, K.; et al. ATP drives lamina propria T(H)17 cell differentiation. Nature 2008, 455, 808–812. [Google Scholar] [CrossRef]
- Abbasian, B.; Shair, A.; O’Gorman, D.B.; Diaz, A.M.P.; Brennan, L.; Engelbrecht, K.; Koenig, D.W.; Reid, G.; Burton, J.P. Potential role of extracellular ATP released by bacteria in bladder infection and contractility. mSphere 2019, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmermann, H.; Zebisch, M.; Strater, N. Cellular function and molecular structure of ecto-nucleotidases. Purinergic Signal. 2012, 8, 437–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skals, M.; Bjaelde, R.G.; Reinholdt, J.; Poulsen, K.; Vad, B.S.; Otzen, D.E.; Leipziger, J.; Praetorius, H.A. Bacterial RTX toxins allow acute ATP release from human erythrocytes directly through the toxin pore. J. Biol. Chem. 2014, 289, 19098–19109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skals, M.; Jorgensen, N.R.; Leipziger, J.; Praetorius, H.A. Alpha-hemolysin from escherichia coli uses endogenous amplification through P2X receptor activation to induce hemolysis. Proc. Natl. Acad. Sci. USA 2009, 106, 4030–4035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xi, C.; Wu, J. dATP/ATP, a multifunctional nucleotide, stimulates bacterial cell lysis, extracellular DNA release and biofilm development. PLoS ONE 2010, 5, e13355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Q.; Tan, K.S. The danger signal extracellular ATP is an inducer of fusobacterium nucleatum biofilm dispersal. Front. Cell. Infect. Microbiol. 2016, 6, 155. [Google Scholar] [CrossRef] [Green Version]
- Binderman, I.; Gadban, N.; Yaffe, A. Extracellular ATP is a key modulator of alveolar bone loss in periodontitis. Arch. Oral Biol. 2017, 81, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Quah, S.Y.; Tan, K.S. Secreted adenosine triphosphate from aggregatibacter actinomycetemcomitans triggers chemokine response. Mol. Oral Microbiol. 2016, 31, 423–434. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.; Atanasova, K.R.; Bui, P.Q.; Lee, J.; Hung, S.C.; Yilmaz, O.; Ojcius, D.M. Porphyromonas gingivalis attenuates ATP-mediated inflammasome activation and HMGB1 release through expression of a nucleoside-diphosphate kinase. Microbes Infect. 2015, 17, 369–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sansom, F.M.; Newton, H.J.; Crikis, S.; Cianciotto, N.P.; Cowan, P.J.; d’Apice, A.J.; Hartland, E.L. A bacterial ecto-triphosphate diphosphohydrolase similar to human CD39 is essential for intracellular multiplication of legionella pneumophila. Cell. Microbiol. 2007, 9, 1922–1935. [Google Scholar] [CrossRef]
- Thammavongsa, V.; Kern, J.W.; Missiakas, D.M.; Schneewind, O. Staphylococcus aureus synthesizes adenosine to escape host immune responses. J. Exp. Med. 2009, 206, 2417–2427. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.; Zhang, Y.; Smith, O.N.C.; Frank, K.L.; Guenther, B.D.; Kern, M.; Schlievert, P.M.; Herzberg, M.C. Ecto-5′-nucleotidase: A candidate virulence factor in streptococcus sanguinis experimental endocarditis. PLoS ONE 2012, 7, e38059. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Yao, Y.; Sumi, Y.; Li, A.; To, U.K.; Elkhal, A.; Inoue, Y.; Woehrle, T.; Zhang, Q.; Hauser, C.; et al. Purinergic signaling: A fundamental mechanism in neutrophil activation. Sci. Signal. 2010, 3. [Google Scholar] [CrossRef] [Green Version]
- Hechler, B.; Gachet, C. Purinergic receptors in thrombosis and inflammation. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 2307–2315. [Google Scholar] [CrossRef] [Green Version]
- Robertson, J.; Lang, S.; Lambert, P.A.; Martin, P.E. Peptidoglycan derived from staphylococcus epidermidis induces Connexin43 hemichannel activity with consequences on the innate immune response in endothelial cells. Biochem. J. 2010, 432, 133–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maitre, B.; Magnenat, S.; Heim, V.; Ravanat, C.; Evans, R.J.; Salle, H.D.L.; Gachet, C.; Hechler, B. The P2X1 receptor is required for neutrophil extravasation during lipopolysaccharide-induced lethal endotoxemia in mice. J. Immunol. 2015, 194, 739–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minasyan, H. Sepsis: Mechanisms of bacterial injury to the patient. Scand. J. Trauma Resusc. Emerg. Med. 2019, 27, 19. [Google Scholar] [CrossRef] [Green Version]
- Krezalek, M.A.; Hyoju, S.; Zaborin, A.; Okafor, E.; Chandrasekar, L.; Bindokas, V.; Guyton, K.; Montgomery, C.P.; Daum, R.S.; Zaborina, O.; et al. Can methicillin-resistant staphylococcus aureus silently travel from the gut to the wound and cause postoperative infection? Modeling the “trojan horse hypothesis”. Ann. Surg. 2018, 267, 749–758. [Google Scholar] [CrossRef] [PubMed]
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Hari, M.S.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.D.; Coopersmith, C.M.; et al. The third international consensus definitions for sepsis and septic shock (sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Vincent, J.L.; Marshall, J.C.; Silva, S.A.N.; Francois, B.; Loeches, I.M.; Lipman, J.; Reinhart, K.; Antonelli, M.; Pickkers, P.; Njimi, H.; et al. Assessment of the worldwide burden of critical illness: The intensive care over nations (ICON) audit. Lancet. Respir. Med. 2014, 2, 380–386. [Google Scholar] [CrossRef]
- Lecut, C.; Faccinetto, C.; Delierneux, C.; Oerle, R.V.; Spronk, H.M.; Evans, R.J.; Benna, J.E.; Bours, V.; Oury, C. ATP-gated P2X1 ion channels protect against endotoxemia by dampening neutrophil activation. J. Thromb. Haemost. 2012, 10, 453–465. [Google Scholar] [CrossRef]
- Santana, P.T.; Benjamim, C.F.; Martinez, C.G.; Kurtenbach, E.; Takiya, C.M.; Silva, R.C. The P2X7 receptor contributes to the development of the exacerbated inflammatory response associated with sepsis. J. Innate Immun. 2015, 7, 417–427. [Google Scholar] [CrossRef]
- Csoka, B.; Nemeth, Z.H.; Szabo, I.; Davies, D.L.; Varga, Z.V.; Paloczi, J.; Falzoni, S.; Virgilio, F.D.; Muramatsu, R.; Yamashita, T.; et al. Macrophage P2X4 receptors augment bacterial killing and protect against sepsis. JCI Insight 2018, 3. [Google Scholar] [CrossRef] [Green Version]
- Csoka, B.; Nemeth, Z.H.; Toro, G.; Idzko, M.; Zech, A.; Koscso, B.; Spolarics, Z.; Antonioli, L.; Cseri, K.; Erdelyi, K.; et al. Extracellular ATP protects against sepsis through macrophage P2X7 purinergic receptors by enhancing intracellular bacterial killing. Faseb J. Publ. Fed. Am. Soc. Exp. Biol. 2015, 29, 3626–3637. [Google Scholar] [CrossRef] [Green Version]
- Greve, A.S.; Skals, M.; Fagerberg, S.K.; Tonnus, W.; Eriksen, S.E.; Evans, R.J.; Linkermann, A.; Praetorius, H.A. P2X1, P2X4, and P2X7 receptor knock out mice expose differential outcome of sepsis induced by alpha-haemolysin producing escherichia coli. Front. Cell. Infect. Microbiol. 2017, 7, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spari, D.; Beldi, G. Extracellular ATP as an Inter-Kingdom Signaling Molecule: Release Mechanisms by Bacteria and Its Implication on the Host. Int. J. Mol. Sci. 2020, 21, 5590. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155590
Spari D, Beldi G. Extracellular ATP as an Inter-Kingdom Signaling Molecule: Release Mechanisms by Bacteria and Its Implication on the Host. International Journal of Molecular Sciences. 2020; 21(15):5590. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155590
Chicago/Turabian StyleSpari, Daniel, and Guido Beldi. 2020. "Extracellular ATP as an Inter-Kingdom Signaling Molecule: Release Mechanisms by Bacteria and Its Implication on the Host" International Journal of Molecular Sciences 21, no. 15: 5590. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155590