Dominant-Negative Attenuation of cAMP-Selective Phosphodiesterase PDE4D Action Affects Learning and Behavior


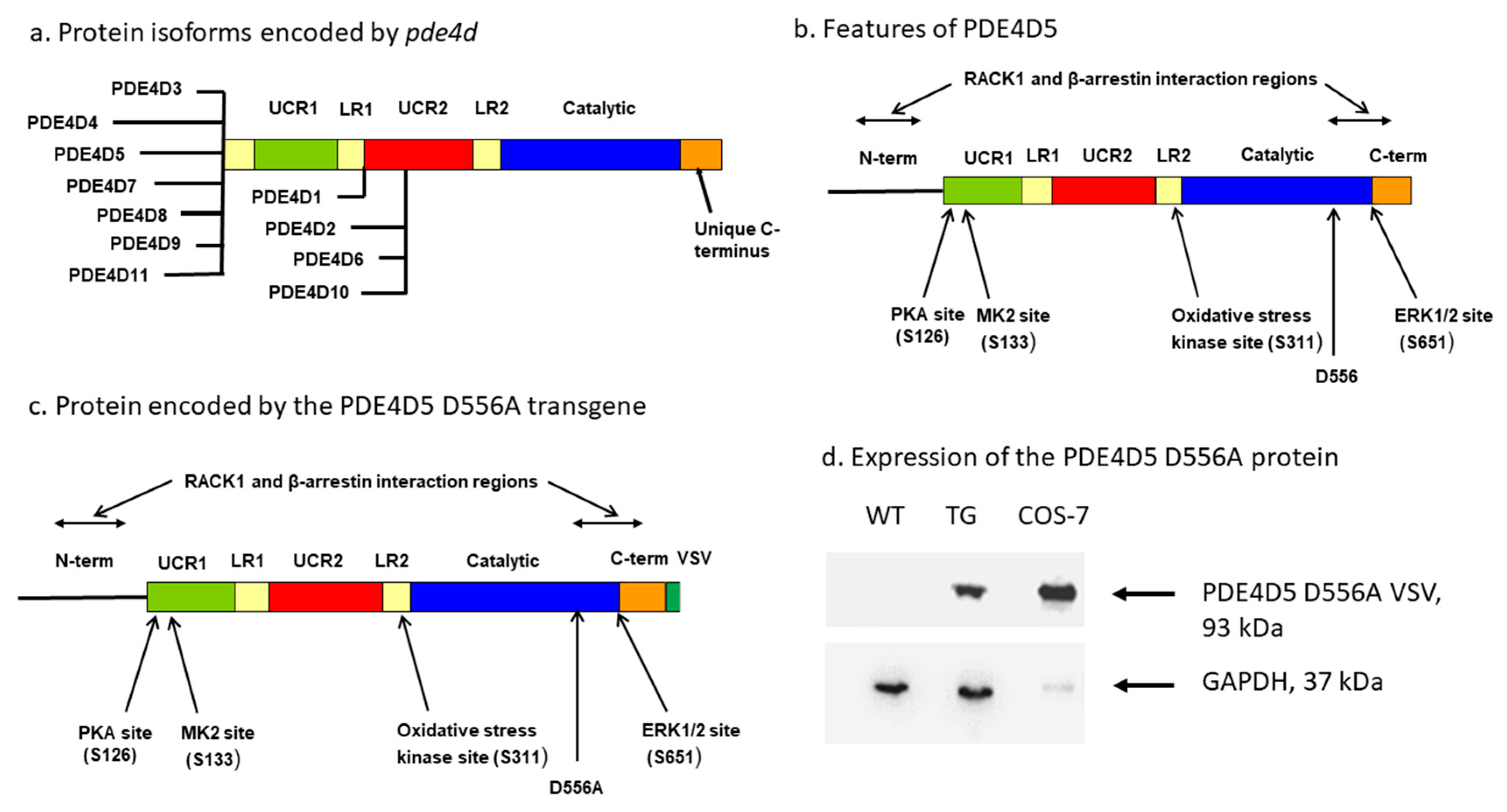{kind=link}
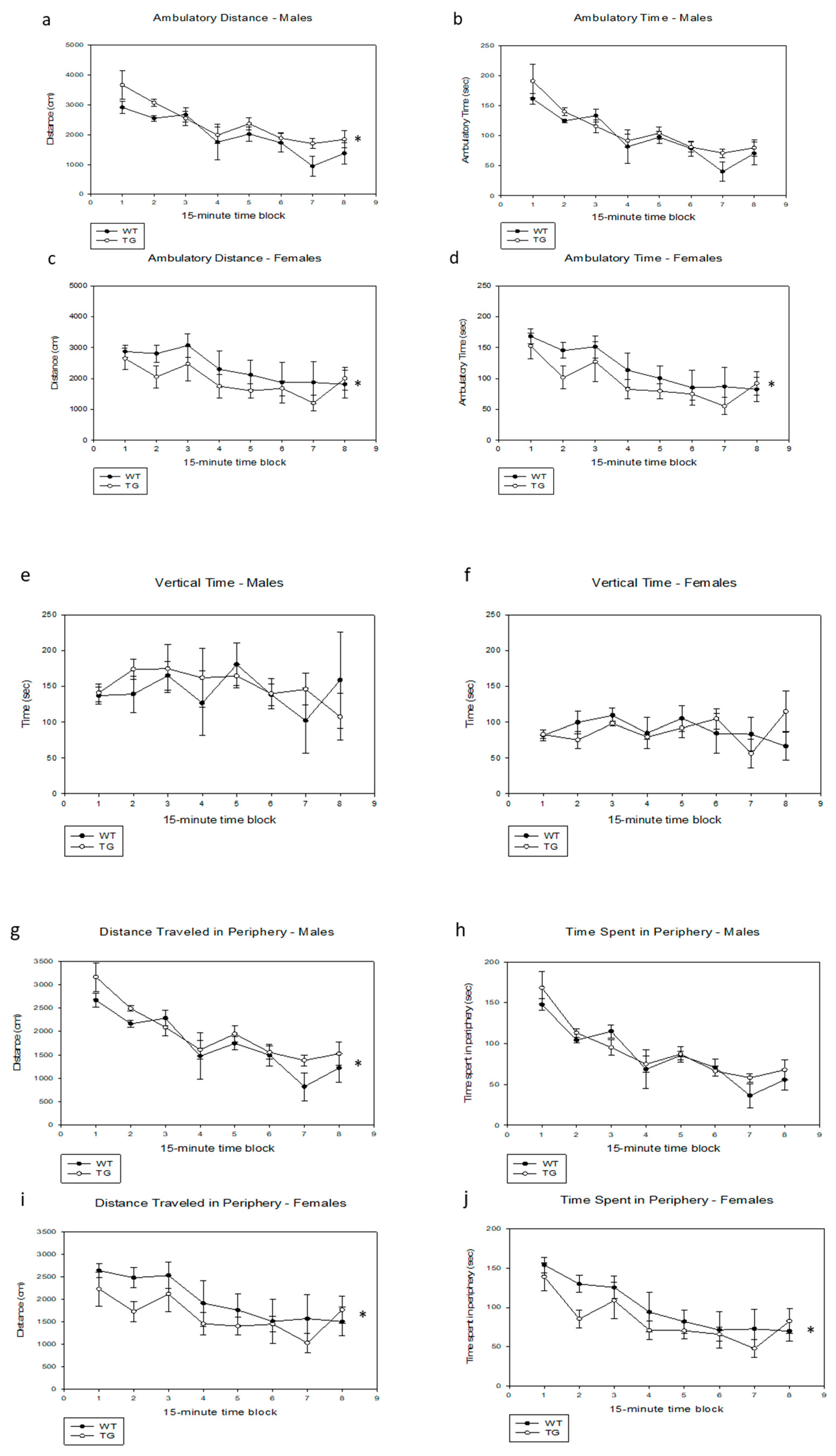{kind=link}
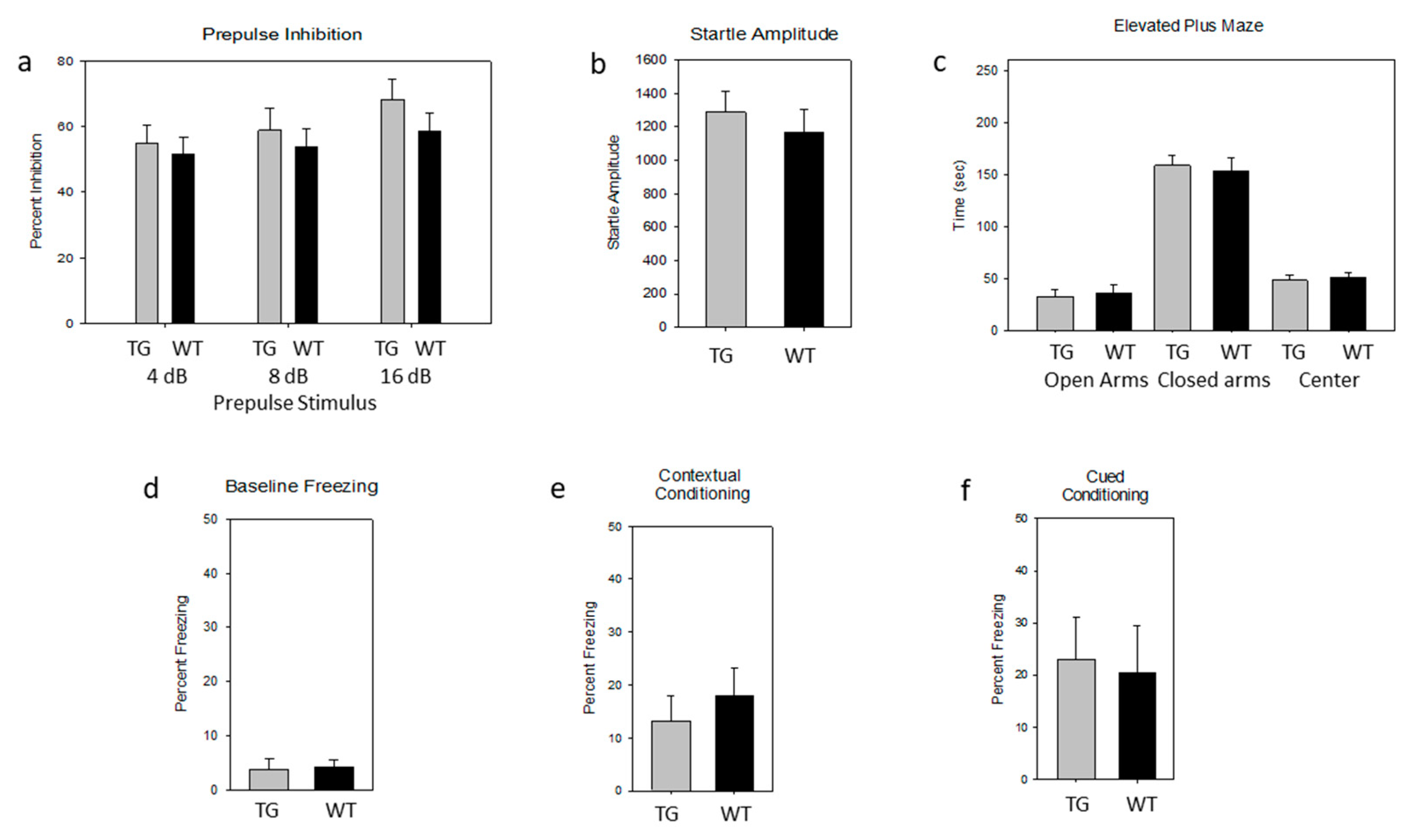{kind=link}
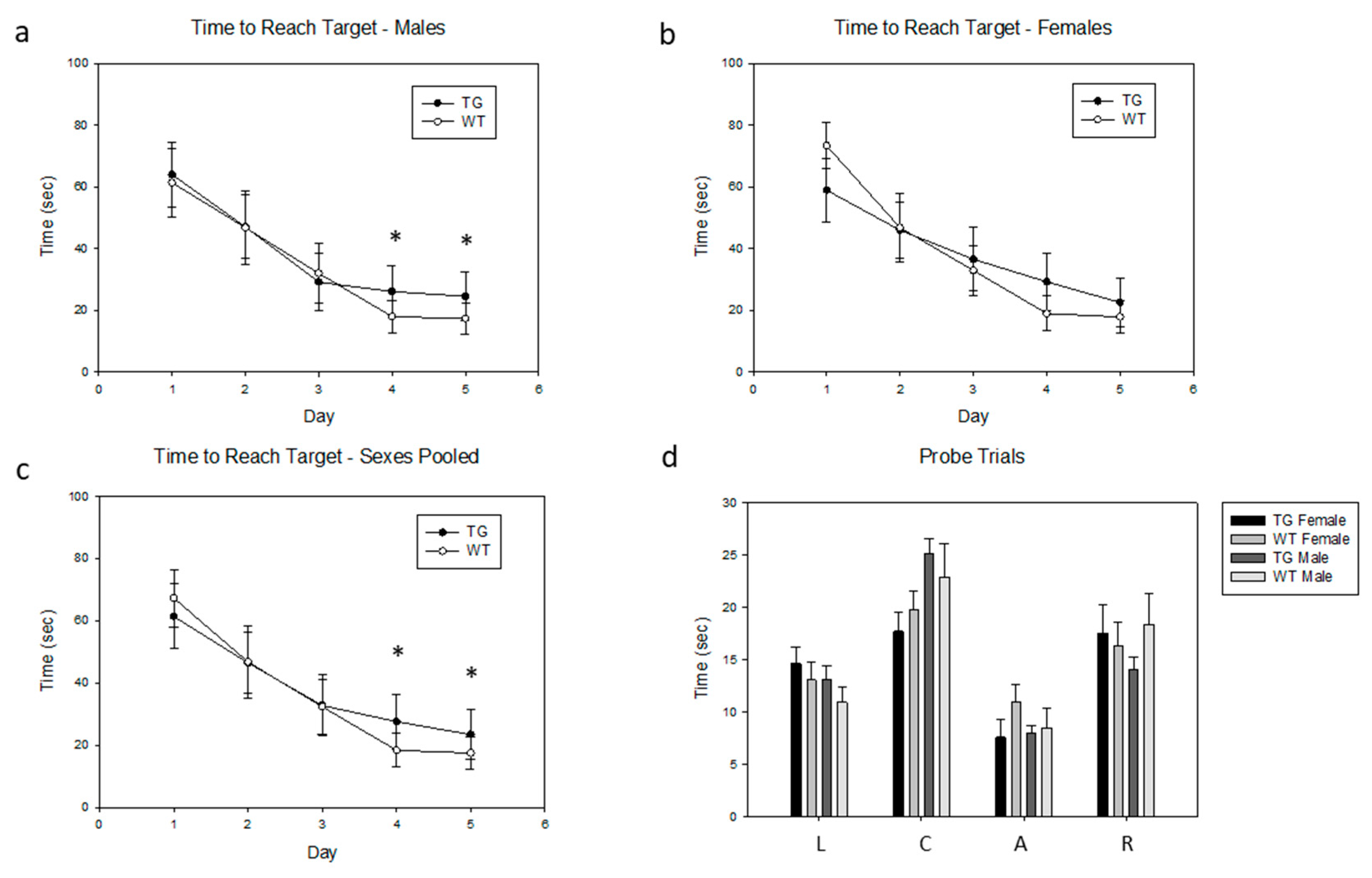{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Mice Expressing a PDE4D5-D556A Transgene in the CNS
2.2. Basic Neurologic Functions in PDE4D5-D556A Transgenic Mice
2.3. Effects of the PDE4D5-D556A Transgene on Activity in a Novel Open Field
2.4. Lack of Effect of the PDE4D5-D556A Transgene on Acoustic Startle and Prepulse Inhibition (PPI)
2.5. Lack of an Effect of the PDE4D5-D556A Transgene on Tests of Anxiety
2.6. Lack of an Effect of the PDE4D5-D556A Transgene on Fear-Associated Conditioning
2.7. Effects of the PDE4D5-D566A Transgene on Spatial Learning
3. Discussion
4. Materials and Methods
4.1. Generation of Transgenic Mice
4.2. Immunoblotting
4.3. Behavioral Assays
4.4. Basic Neurobiological Battery
4.5. Open-Field Testing
4.6. Acoustic Startle and Prepulse Inhibition
4.7. Elevated Plus Maze
4.8. Context-Dependent and Cue-Dependent Fear Conditioning
4.9. Morris Water Maze
4.10. Availability of Data and Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fleischhacker, W.W.; Hinterhuber, H.; Bauer, H.; Pflug, B.; Berner, P.; Simhandl, C.; Wolf, R.; Gerlach, W.; Jaklitsch, H.; Sastre-y-Hernandez, M.; et al. A multicenter double-blind study of three different doses of the new cAMP-phosphodiesterase inhibitor rolipram in patients with major depressive disorder. Neuropsychobiology 1992, 26, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Scott, A.I.; Perini, A.F.; Shering, P.A.; Whalley, L.J. In-patient major depression: Is rolipram as effective as amitriptyline? Eur. J. Clin. Pharmacol. 1991, 40, 127–129. [Google Scholar] [PubMed]
- Krause, W.; Kuhne, G.; Sauerbrey, N. Pharmacokinetics of (+)-rolipram and (-)-rolipram in healthy volunteers. Eur. J. Clin. Pharmacol. 1990, 38, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Hebenstreit, G.F.; Fellerer, K.; Fichte, K.; Fischer, G.; Geyer, N.; Meya, U.; Hernandez, M.; Schony, W.; Schratzer, M.; Soukop, W.; et al. Rolipram in major depressive disorder: Results of a double-blind comparative study with imipramine. Pharmacopsychiatry 1989, 22, 156–160. [Google Scholar] [CrossRef]
- Eckmann, F.; Fichte, K.; Meya, U. Rolipram in major depression: Results of a double-blind comparative study with amitriptyline. Curr. Ther. Res. 1988, 43, 291–295. [Google Scholar]
- Horowski, R.; Sastre-Y-Hernandez, M. Clinical effects of the neurotropic selective cAMP phosphodiesterase inhibitor rolipram in depressed patients: Global evaluation of the preliminary reports. Curr. Ther. Res. 1985, 38, 23–29. [Google Scholar]
- Bobon, D.; Breulet, M.; Gerard-Vandenhove, M.A.; Guiot-Goffioul, F.; Plomteux, G.; Sastre-y-Hernandez, M.; Schratzer, M.; Troisfontaines, B.; Von Frenckell, R.; Wachtel, H. Is phosphodiesterase inhibition a new mechanism of antidepressant action? A double blind double-dummy study between rolipram and desipramine in hospitalized major and/or endogenous depressives. Eur. Arch. Psychiatry Neurol. Sci. 1988, 238, 2–6. [Google Scholar] [CrossRef]
- Zeller, E.; Stief, H.J.; Pflug, B.; Sastre-y-Hernandez, M. Results of a phase II study of the antidepressant effect of rolipram. Pharmacopsychiatry 1984, 17, 188–190. [Google Scholar] [CrossRef]
- Van Duinen, M.A.; Sambeth, A.; Heckman, P.R.; Smit, S.; Tsai, M.; Lahu, G.; Uz, T.; Blokland, A.; Prickaerts, J. Acute administration of roflumilast enhances immediate recall of verbal word memory in healthy young adults. Neuropharmacology 2018, 131, 31–38. [Google Scholar] [CrossRef]
- Heckman, P.R.; Van Duinen, M.A.; Blokland, A.; Uz, T.; Prickaerts, J.; Sambeth, A. Acute administration of roflumilast enhances sensory gating in healthy young humans in a randomized trial. Psychopharmacology (Berl) 2018, 235, 301–308. [Google Scholar] [CrossRef] [Green Version]
- Prickaerts, J.; Heckman, P.R.; Blokland, A. Investigational phosphodiesterase inhibitors in phase I and phase II clinical trials for Alzheimer’s disease. Expert Opin. Investig. Drugs 2017, 26, 1033–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prickaerts, J.; Heckman, P.R.; Blokland, A. Acute treatment with the PDE4 inhibitor roflumilast improves verbal word memory in healthy old individuals: A double-blind placebo-controlled study. Neurobiol. Aging 2019, 77, 37–43. [Google Scholar] [CrossRef]
- Wachtel, H. Potential antidepressant activity of rolipram and other selective cyclic adenosine 3′,5′-monophosphate phosphodiesterase inhibitors. Neuropharmacology 1983, 22, 267–272. [Google Scholar] [CrossRef]
- Wachtel, H.; Schneider, H.H. Rolipram, a novel antidepressant drug, reverses the hypothermia and hypokinesia of monoamine-depleted mice by an action beyond postsynaptic monoamine receptors. Neuropharmacology 1986, 25, 1119–1126. [Google Scholar] [CrossRef]
- Barad, M.; Bourtchouladze, R.; Winder, D.G.; Golan, H.; Kandel, E. Rolipram, a type IV-specific phosphodiesterase inhibitor, facilitates the establishment of long-lasting long-term potentiation and improves memory. Proc. Natl. Acad. Sci. USA 1998, 95, 15020–15025. [Google Scholar] [CrossRef] [Green Version]
- Bach, M.E.; Barad, M.; Son, H.; Zhuo, M.; Lu, Y.F.; Shih, R.; Mansuy, I.; Hawkins, R.D.; Kandel, E.R. Age-related defects in spatial memory are correlated with defects in the late phase of hippocampal long-term potentiation in vitro and are attenuated by drugs that enhance the cAMP signaling pathway. Proc. Natl. Acad. Sci. USA 1999, 96, 5280–5285. [Google Scholar] [CrossRef] [Green Version]
- Titus, D.J.; Sakurai, A.; Kang, Y.; Furones, C.; Jergova, S.; Santos, R.; Sick, T.J.; Atkins, C.M. Phosphodiesterase inhibition rescues chronic cognitive deficits induced by traumatic brain injury. J. Neurosci. 2013, 33, 5216–5226. [Google Scholar] [CrossRef] [Green Version]
- Mueller, E.M.; Hofmann, S.G.; Cherry, J.A. The type IV phosphodiesterase inhibitor rolipram disturbs expression and extinction of conditioned fear in mice. Neuropharmacology 2010, 59, 1–8. [Google Scholar] [CrossRef]
- Nibuya, M.; Nestler, E.J.; Duman, R.S. Chronic antidepressant administration increases the expression of cAMP response element binding protein (CREB) in rat hippocampus. J. Neurosci. 1996, 16, 2365–2372. [Google Scholar] [CrossRef] [Green Version]
- Zhong, P.; Wang, W.; Yu, F.; Nazari, M.; Liu, X.; Liu, Q.S. Phosphodiesterase 4 inhibition impairs cocaine-induced inhibitory synaptic plasticity and conditioned place preference. Neuropsychopharmacology 2012, 37, 2377–2387. [Google Scholar] [CrossRef] [Green Version]
- Villiger, J.W.; Dunn, A.J. Phosphodiesterase inhibitors facilitate memory for passive avoidance conditioning. Behav. Neural Biol. 1981, 31, 354–359. [Google Scholar] [CrossRef]
- Thompson, B.E.; Sachs, B.D.; Kantak, K.M.; Cherry, J.A. The Type IV phosphodiesterase inhibitor rolipram interferes with drug-induced conditioned place preference but not immediate early gene induction in mice. Eur. J. Neurosci. 2004, 19, 2561–2568. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.T.; Zhao, Y.; Huang, Y.; Dorairaj, N.R.; Chandler, L.J.; O’Donnell, J.M. Inhibition of the phosphodiesterase 4 (PDE4) enzyme reverses memory deficits produced by infusion of the MEK inhibitor U0126 into the CA1 subregion of the rat hippocampus. Neuropsychopharmacology 2004, 29, 1432–1439. [Google Scholar] [CrossRef] [PubMed]
- Rutten, K.; Prickaerts, J.H.; Blokland, A. Rolipram reverses scopolamine-induced and time-dependent memory deficits in object recognition by different mechanisms of action. Neurobiol. Learn. Mem. 2006, 85, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.T.; Zhao, Y.; Huang, Y.; Deng, C.; Hopper, A.T.; De Vivo, M.; Rose, G.M.; O’Donnell, J.M. Antidepressant-like effects of PDE4 inhibitors mediated by the high-affinity rolipram binding state (HARBS) of the phosphodiesterase-4 enzyme (PDE4) in rats. Psychopharmacology (Berl) 2006, 186, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Rutten, K.; Basile, J.L.; Prickaerts, J.; Blokland, A.; Vivian, J.A. Selective PDE inhibitors rolipram and sildenafil improve object retrieval performance in adult cynomolgus macaques. Psychopharmacology (Berl) 2008, 196, 643–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutten, K.; Misner, D.L.; Works, M.; Blokland, A.; Novak, T.J.; Santarelli, L.; Wallace, T.L. Enhanced long-term potentiation and impaired learning in phosphodiesterase 4D-knockout (PDE4D) mice. Eur. J. Neurosci. 2008, 28, 625–632. [Google Scholar] [CrossRef]
- Kanes, S.J.; Tokarczyk, J.; Siegel, S.J.; Bilker, W.; Abel, T.; Kelly, M.P. Rolipram: A specific phosphodiesterase 4 inhibitor with potential antipsychotic activity. Neuroscience 2007, 144, 239–246. [Google Scholar] [CrossRef] [Green Version]
- Rutten, K.; Prickaerts, J.; Schaenzle, G.; Rosenbrock, H.; Blokland, A. Sub-chronic rolipram treatment leads to a persistent improvement in long-term object memory in rats. Neurobiol. Learn. Mem. 2008, 90, 569–575. [Google Scholar] [CrossRef]
- Li, Y.F.; Huang, Y.; Amsdell, S.L.; Xiao, L.; O’Donnell, J.M.; Zhang, H.T. Antidepressant- and anxiolytic-like effects of the phosphodiesterase-4 inhibitor rolipram on behavior depend on cyclic AMP response element binding protein-mediated neurogenesis in the hippocampus. Neuropsychopharmacology 2009, 34, 2404–2419. [Google Scholar] [CrossRef] [Green Version]
- Rutten, K.; Van Donkelaar, E.L.; Ferrington, L.; Blokland, A.; Bollen, E.; Steinbusch, H.W.; Kelly, P.A.; Prickaerts, J.H. Phosphodiesterase inhibitors enhance object memory independent of cerebral blood flow and glucose utilization in rats. Neuropsychopharmacology 2009, 34, 1914–1925. [Google Scholar] [CrossRef] [PubMed]
- Vecsey, C.G.; Baillie, G.S.; Jaganath, D.; Havekes, R.; Daniels, A.; Wimmer, M.; Huang, T.; Brown, K.M.; Li, X.Y.; Descalzi, G.; et al. Sleep deprivation impairs cAMP signalling in the hippocampus. Nature 2009, 461, 1122–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.F.; Wang, C.; Lin, H.B.; Li, Y.F.; Huang, Y.; Xu, J.P.; Zhang, H.T. Inhibition of phosphodiesterase-4 reverses memory deficits produced by Abeta25-35 or Abeta1-40 peptide in rats. Psychopharmacology (Berl) 2010, 212, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Yang, X.M.; Zhuo, Y.Y.; Zhou, H.; Lin, H.B.; Cheng, Y.F.; Xu, J.P.; Zhang, H.T. The phosphodiesterase-4 inhibitor rolipram reverses Abeta-induced cognitive impairment and neuroinflammatory and apoptotic responses in rats. Int. J. Neuropsychopharmacol. 2012, 15, 749–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werenicz, A.; Christoff, R.R.; Blank, M.; Jobim, P.F.; Pedroso, T.R.; Reolon, G.K.; Schroder, N.; Roesler, R. Administration of the phosphodiesterase type 4 inhibitor rolipram into the amygdala at a specific time interval after learning increases recognition memory persistence. Learn. Mem. 2012, 19, 495–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiescholleck, V.; Manahan-Vaughan, D. PDE4 inhibition enhances hippocampal synaptic plasticity in vivo and rescues MK801-induced impairment of long-term potentiation and object recognition memory in an animal model of psychosis. Transl. Psychiatry 2012, 2, e89. [Google Scholar] [CrossRef] [Green Version]
- Vanmierlo, T.; Creemers, P.; Akkerman, S.; van Duinen, M.; Sambeth, A.; De Vry, J.; Uz, T.; Blokland, A.; Prickaerts, J. The PDE4 inhibitor roflumilast improves memory in rodents at non-emetic doses. Behav. Brain Res. 2016, 303, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Xu, Y.; Zhang, H.T.; Gurney, M.E.; O’Donnell, J.M. Comparison of the Pharmacological Profiles of Selective PDE4B and PDE4D Inhibitors in the Central Nervous System. Sci. Rep. 2017, 7, 40115. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Zhang, J.; Lu, Y.; Lin, P.; Pan, T.; Zhao, X.; Liu, A.; Wang, Q.; Zhou, W.; Zhang, H.T. Antidepressant-like effects of the phosphodiesterase-4 inhibitor etazolate and phosphodiesterase-5 inhibitor sildenafil via cyclic AMP or cyclic GMP signaling in mice. Metab. Brain Dis. 2014, 29, 673–682. [Google Scholar] [CrossRef]
- Liu, X.; Hao, P.D.; Yang, M.F.; Sun, J.Y.; Mao, L.L.; Fan, C.D.; Zhang, Z.Y.; Li, D.W.; Yang, X.Y.; Sun, B.L.; et al. The phosphodiesterase-4 inhibitor roflumilast decreases ethanol consumption in C57BL/6J mice. Psychopharmacology (Berl) 2017, 234, 2409–2419. [Google Scholar] [CrossRef]
- Zhong, Y.; Zhu, Y.; He, T.; Li, W.; Yan, H.; Miao, Y. Rolipram-induced improvement of cognitive function correlates with changes in hippocampal CREB phosphorylation, BDNF and Arc protein levels. Neurosci. Lett. 2016, 610, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Akar, F.; Mutlu, O.; Celikyurt, I.K.; Ulak, G.; Erden, F.; Bektas, E.; Tanyeri, P. Effects of rolipram and zaprinast on learning and memory in the Morris water maze and radial arm maze tests in naive mice. Drug Res. (Stuttg) 2015, 65, 86–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sierksma, A.S.; van den Hove, D.L.; Pfau, F.; Philippens, M.; Bruno, O.; Fedele, E.; Ricciarelli, R.; Steinbusch, H.W.; Vanmierlo, T.; Prickaerts, J. Improvement of spatial memory function in APPswe/PS1dE9 mice after chronic inhibition of phosphodiesterase type 4D. Neuropharmacology 2014, 77, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Titus, D.J.; Wilson, N.M.; Freund, J.E.; Carballosa, M.M.; Sikah, K.E.; Furones, C.; Dietrich, W.D.; Gurney, M.E.; Atkins, C.M. Chronic Cognitive Dysfunction after Traumatic Brain Injury Is Improved with a Phosphodiesterase 4B Inhibitor. J. Neurosci. 2016, 36, 7095–7108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, N.M.; Gurney, M.E.; Dietrich, W.D.; Atkins, C.M. Therapeutic benefits of phosphodiesterase 4B inhibition after traumatic brain injury. PLoS ONE 2017, 12, e0178013. [Google Scholar] [CrossRef] [PubMed]
- Hedde, J.R.; Hanks, A.N.; Schmidt, C.J.; Hughes, Z.A. The isozyme selective phosphodiesterase-4 inhibitor, ABI-4, attenuates the effects of lipopolysaccharide in human cells and rodent models of peripheral and CNS inflammation. Brain Behav. Immun. 2017, 64, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Ricciarelli, R.; Brullo, C.; Prickaerts, J.; Arancio, O.; Villa, C.; Rebosio, C.; Calcagno, E.; Balbi, M.; van Hagen, B.T.; Argyrousi, E.K.; et al. Memory-enhancing effects of GEBR-32a, a new PDE4D inhibitor holding promise for the treatment of Alzheimer’s disease. Sci. Rep. 2017, 7, 46320. [Google Scholar] [CrossRef]
- Zhang, C.; Xu, Y.; Chowdhary, A.; Fox, D., 3rd; Gurney, M.E.; Zhang, H.T.; Auerbach, B.D.; Salvi, R.J.; Yang, M.; Li, G.; et al. Memory enhancing effects of BPN14770, an allosteric inhibitor of phosphodiesterase-4D, in wild-type and humanized mice. Neuropsychopharmacology 2018, 43, 2299–2309. [Google Scholar] [CrossRef]
- Conti, M.; Beavo, J. Biochemistry and physiology of cyclic nucleotide phosphodiesterases: Essential components in cyclic nucleotide signaling. Annu. Rev. Biochem. 2007, 76, 481–511. [Google Scholar] [CrossRef]
- Maurice, D.H.; Ke, H.; Ahmad, F.; Wang, Y.; Chung, J.; Manganiello, V.C. Advances in targeting cyclic nucleotide phosphodiesterases. Nat. Rev. Drug Discov. 2014, 13, 290–314. [Google Scholar] [CrossRef] [Green Version]
- Baillie, G.S.; Tejeda, G.S.; Kelly, M.P. Therapeutic targeting of 3′,5′-cyclic nucleotide phosphodiesterases: Inhibition and beyond. Nat. Rev. Drug Discov. 2019, 18, 770–796. [Google Scholar] [CrossRef] [PubMed]
- Oba, Y.; Lone, N.A. Efficacy and safety of roflumilast in patients with chronic obstructive pulmonary disease: A systematic review and meta-analysis. Ther. Adv. Respir. Dis. 2013, 7, 13–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papp, K.; Cather, J.C.; Rosoph, L.; Sofen, H.; Langley, R.G.; Matheson, R.T.; Hu, C.; Day, R.M. Efficacy of apremilast in the treatment of moderate to severe psoriasis: A randomised controlled trial. Lancet 2012, 380, 738–746. [Google Scholar] [CrossRef]
- Jarnagin, K.; Chanda, S.; Coronado, D.; Ciaravino, V.; Zane, L.T.; Guttman-Yassky, E.; Lebwohl, M.G. Crisaborole Topical Ointment, 2%: A Nonsteroidal, Topical, Anti-Inflammatory Phosphodiesterase 4 Inhibitor in Clinical Development for the Treatment of Atopic Dermatitis. J. Drugs Dermatol. 2016, 15, 390–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulhall, A.M.; Droege, C.A.; Ernst, N.E.; Panos, R.J.; Zafar, M.A. Phosphodiesterase 4 inhibitors for the treatment of chronic obstructive pulmonary disease: A review of current and developing drugs. Expert Opin. Investig. Drugs 2015, 24, 1597–1611. [Google Scholar] [CrossRef]
- Singh, D.; Martinez, F.J.; Watz, H.; Bengtsson, T.; Maurer, B.T. A dose-ranging study of the inhaled dual phosphodiesterase 3 and 4 inhibitor ensifentrine in COPD. Respir. Res. 2020, 21, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.T.; Huang, Y.; Suvarna, N.U.; Deng, C.; Crissman, A.M.; Hopper, A.T.; De Vivo, M.; Rose, G.M.; O’Donnell, J.M. Effects of the novel PDE4 inhibitors MEM1018 and MEM1091 on memory in the radial-arm maze and inhibitory avoidance tests in rats. Psychopharmacology (Berl) 2005, 179, 613–619. [Google Scholar] [CrossRef]
- Tempesta, D.; Mazza, M.; Serroni, N.; Moschetta, F.S.; Di Giannantonio, M.; Ferrara, M.; De Berardis, D. Neuropsychological functioning in young subjects with generalized anxiety disorder with and without pharmacotherapy. Prog. Neuro-psychopharmacol. Biol. Psychiatry 2013, 45, 236–241. [Google Scholar] [CrossRef]
- Rock, P.L.; Roiser, J.P.; Riedel, W.J.; Blackwell, A.D. Cognitive impairment in depression: A systematic review and meta-analysis. Psychol. Med. 2014, 44, 2029–2040. [Google Scholar] [CrossRef] [Green Version]
- Prosdocimi, T.; Mollica, L.; Donini, S.; Semrau, M.S.; Lucarelli, A.P.; Aiolfi, E.; Cavalli, A.; Storici, P.; Alfei, S.; Brullo, C.; et al. Molecular Bases of PDE4D Inhibition by Memory-Enhancing GEBR Library Compounds. Biochemistry 2018, 57, 2876–2888. [Google Scholar] [CrossRef]
- Cui, S.Y.; Yang, M.X.; Zhang, Y.H.; Zheng, V.; Zhang, H.T.; Gurney, M.E.; Xu, Y.; O’Donnell, J.M. Protection from Amyloid beta Peptide-Induced Memory, Biochemical, and Morphological Deficits by a Phosphodiesterase-4D Allosteric Inhibitor. J. Pharmacol. Exp. Ther 2019, 371, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Gurney, M.E.; Nugent, R.A.; Mo, X.; Sindac, J.A.; Hagen, T.J.; Fox, D., 3rd; O’Donnell, J.M.; Zhang, C.; Xu, Y.; Zhang, H.T.; et al. Design and Synthesis of Selective Phosphodiesterase 4D (PDE4D) Allosteric Inhibitors for the Treatment of Fragile X Syndrome and Other Brain Disorders. J. Med. Chem. 2019, 62, 4884–4901. [Google Scholar] [CrossRef] [PubMed]
- Titus, D.J.; Wilson, N.M.; Alcazar, O.; Calixte, D.A.; Dietrich, W.D.; Gurney, M.E.; Atkins, C.M. A negative allosteric modulator of PDE4D enhances learning after traumatic brain injury. Neurobiol. Learn. Mem. 2018, 148, 38–49. [Google Scholar] [CrossRef] [PubMed]
- Kandel, E.R. The molecular biology of memory storage: A dialogue between genes and synapses. Science 2001, 294, 1030–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, D.A.; Greenberg, M.E. CREB: A mediator of long-term memory from mollusks to mammals. Cell 1994, 79, 5–8. [Google Scholar] [CrossRef]
- Silva, A.J.; Kogan, J.H.; Frankland, P.W.; Kida, S. CREB and memory. Annu. Rev. Neurosci. 1998, 21, 127–148. [Google Scholar] [CrossRef] [Green Version]
- Newton, S.S.; Thome, J.; Wallace, T.L.; Shirayama, Y.; Schlesinger, L.; Sakai, N.; Chen, J.; Neve, R.; Nestler, E.J.; Duman, R.S. Inhibition of cAMP response element-binding protein or dynorphin in the nucleus accumbens produces an antidepressant-like effect. J. Neurosci. 2002, 22, 10883–10890. [Google Scholar] [CrossRef] [Green Version]
- Gloerich, M.; Bos, J.L. Epac: Defining a new mechanism for cAMP action. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 355–375. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.; Dekker, F.J.; Maarsingh, H. Exchange protein directly activated by cAMP (epac): A multidomain cAMP mediator in the regulation of diverse biological functions. Pharmacol. Rev. 2013, 65, 670–709. [Google Scholar] [CrossRef] [Green Version]
- Kaupp, U.B.; Seifert, R. Cyclic nucleotide-gated ion channels. Physiol. Rev. 2002, 82, 769–824. [Google Scholar] [CrossRef] [Green Version]
- Schindler, R.F.; Brand, T. The Popeye domain containing protein family—A novel class of cAMP effectors with important functions in multiple tissues. Prog. Biophys. Mol. Biol. 2016, 120, 28–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, G.; Michaeli, T.; Martins, T.; St John, T.; Steiner, B.; Rodgers, L.; Riggs, M.; Wigler, M.; Ferguson, K. A family of human phosphodiesterases homologous to the dunce learning and memory gene product of Drosophila melanogaster are potential targets for antidepressant drugs. Mol. Cell Biol. 1993, 13, 6558–6571. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.R.; Nicodemus-Johnson, J.; Danziger, R.S. An evolutionary analysis of cAMP-specific Phosphodiesterase 4 alternative splicing. BMC Evol. Biol. 2010, 10, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, G.B.; Rodgers, L.; Riggs, M. Differential CNS expression of alternative mRNA isoforms of the mammalian genes encoding cAMP-specific phosphodiesterases. Gene 1994, 149, 237–244. [Google Scholar] [CrossRef]
- Takahashi, M.; Terwilliger, R.; Lane, C.; Mezes, P.S.; Conti, M.; Duman, R.S. Chronic antidepressant administration increases the expression of cAMP- specific phosphodiesterase 4A and 4B isoforms. J. Neurosci. 1999, 19, 610–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherry, J.A.; Davis, R.L. Cyclic AMP phosphodiesterases are localized in regions of the mouse brain associated with reinforcement, movement, and affect. J. Comp. Neurol. 1999, 407, 287–301. [Google Scholar] [CrossRef]
- Miro, X.; Perez-Torres, S.; Puigdomenech, P.; Palacios, J.M.; Mengod, G. Differential distribution of PDE4D splice variant mRNAs in rat brain suggests association with specific pathways and presynaptical localization. Synapse 2002, 45, 259–269. [Google Scholar] [CrossRef]
- D’Sa, C.; Tolbert, L.M.; Conti, M.; Duman, R.S. Regulation of cAMP-specific phosphodiesterases type 4B and 4D (PDE4) splice variants by cAMP signaling in primary cortical neurons. J. Neurochem. 2002, 81, 745–757. [Google Scholar] [CrossRef]
- D’Sa, C.; Eisch, A.J.; Bolger, G.B.; Duman, R.S. Differential expression and regulation of the cAMP-selective phosphodiesterase type 4A splice variants in rat brain by chronic antidepressant administration. Eur. J. Neurosci. 2005, 22, 1463–1475. [Google Scholar] [CrossRef]
- Reyes-Irisarri, E.; Perez-Torres, S.; Miro, X.; Martinez, E.; Puigdomenech, P.; Palacios, J.M.; Mengod, G. Differential distribution of PDE4B splice variant mRNAs in rat brain and the effects of systemic administration of LPS in their expression. Synapse 2008, 62, 74–79. [Google Scholar] [CrossRef] [Green Version]
- Nishi, A.; Kuroiwa, M.; Miller, D.B.; O’Callaghan, J.P.; Bateup, H.S.; Shuto, T.; Sotogaku, N.; Fukuda, T.; Heintz, N.; Greengard, P.; et al. Distinct roles of PDE4 and PDE10A in the regulation of cAMP/PKA signaling in the striatum. J. Neurosci. 2008, 28, 10460–10471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, F.; Perez-Torres, S.; De Caro, R.; Porzionato, A.; Macchi, V.; Beleta, J.; Gavalda, A.; Palacios, J.M.; Mengod, G. The human area postrema and other nuclei related to the emetic reflex express cAMP phosphodiesterases 4B and 4D. J. Chem. Neuroanat. 2010, 40, 36–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuroiwa, M.; Snyder, G.L.; Shuto, T.; Fukuda, A.; Yanagawa, Y.; Benavides, D.R.; Nairn, A.C.; Bibb, J.A.; Greengard, P.; Nishi, A. Phosphodiesterase 4 inhibition enhances the dopamine D1 receptor/PKA/DARPP-32 signaling cascade in frontal cortex. Psychopharmacology (Berl) 2012, 219, 1065–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, T.; Frey, J.U. Expression of the specific type IV phosphodiesterase gene PDE4B3 during different phases of long-term potentiation in single hippocampal slices of rats in vitro. Neuroscience 2003, 117, 627–638. [Google Scholar] [CrossRef]
- Zhang, K.Y.; Card, G.L.; Suzuki, Y.; Artis, D.R.; Fong, D.; Gillette, S.; Hsieh, D.; Neiman, J.; West, B.L.; Zhang, C.; et al. A glutamine switch mechanism for nucleotide selectivity by phosphodiesterases. Mol. Cell 2004, 15, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Burgin, A.B.; Magnusson, O.T.; Singh, J.; Witte, P.; Staker, B.L.; Bjornsson, J.M.; Thorsteinsdottir, M.; Hrafnsdottir, S.; Hagen, T.; Kiselyov, A.S.; et al. Design of phosphodiesterase 4D (PDE4D) allosteric modulators for enhancing cognition with improved safety. Nat. Biotechnol. 2010, 28, 63–70. [Google Scholar] [CrossRef]
- Francis, S.H.; Blount, M.A.; Corbin, J.D. Mammalian cyclic nucleotide phosphodiesterases: Molecular mechanisms and physiological functions. Physiol. Rev. 2011, 91, 651–690. [Google Scholar] [CrossRef] [Green Version]
- Bolger, G.B.; Erdogan, S.; Jones, R.E.; Loughney, K.; Scotland, G.; Hoffmann, R.; Wilkinson, I.; Farrell, C.; Houslay, M.D. Characterization of five different proteins produced by alternatively spliced mRNAs from the human cAMP-specific phosphodiesterase PDE4D gene. Biochem. J. 1997, 328, 539–548. [Google Scholar] [CrossRef] [Green Version]
- Lamontagne, S.; Meadows, E.; Luk, P.; Normandin, D.; Muise, E.; Boulet, L.; Pon, D.J.; Robichaud, A.; Robertson, G.S.; Metters, K.M.; et al. Localization of phosphodiesterase-4 isoforms in the medulla and nodose ganglion of the squirrel monkey. Brain Res. 2001, 920, 84–96. [Google Scholar] [CrossRef]
- Whitaker, C.M.; Cooper, N.G. The novel distribution of phosphodiesterase-4 subtypes within the rat retina. Neuroscience 2009, 163, 1277–1291. [Google Scholar] [CrossRef] [Green Version]
- Lakics, V.; Karran, E.H.; Boess, F.G. Quantitative comparison of phosphodiesterase mRNA distribution in human brain and peripheral tissues. Neuropharmacology 2010, 59, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Johansson, E.M.; Reyes-Irisarri, E.; Mengod, G. Comparison of cAMP-specific phosphodiesterase mRNAs distribution in mouse and rat brain. Neurosci. Lett. 2012, 525, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Oliva, A.A., Jr.; Kang, Y.; Furones, C.; Alonso, O.F.; Bruno, O.; Dietrich, W.D.; Atkins, C.M. Phosphodiesterase isoform-specific expression induced by traumatic brain injury. J. Neurochem. 2012, 123, 1019–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, M.P.; Adamowicz, W.; Bove, S.; Hartman, A.J.; Mariga, A.; Pathak, G.; Reinhart, V.; Romegialli, A.; Kleiman, R.J. Select 3′, 5′-cyclic nucleotide phosphodiesterases exhibit altered expression in the aged rodent brain. Cell Signal. 2014, 26, 383–397. [Google Scholar] [CrossRef] [PubMed]
- Kelly, M.P. Cyclic nucleotide signaling changes associated with normal aging and age-related diseases of the brain. Cell Signal. 2018, 42, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Gretarsdottir, S.; Thorleifsson, G.; Reynisdottir, S.T.; Manolescu, A.; Jonsdottir, S.; Jonsdottir, T.; Gudmundsdottir, T.; Bjarnadottir, S.M.; Einarsson, O.B.; Gudjonsdottir, H.M.; et al. The gene encoding phosphodiesterase 4D confers risk of ischemic stroke. Nat. Genet. 2003, 35, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Deng, C.; Bugaj-Gaweda, B.; Kwan, M.; Gunwaldsen, C.; Leonard, C.; Xin, X.; Hu, Y.; Unterbeck, A.; De Vivo, M. Cloning and characterization of novel PDE4D isoforms PDE4D6 and PDE4D7. Cell Signal. 2003, 15, 883–891. [Google Scholar] [CrossRef]
- Richter, W.; Jin, S.L.; Conti, M. Splice variants of the cyclic nucleotide phosphodiesterase PDE4D are differentially expressed and regulated in rat tissue. Biochem. J. 2005, 388, 803–811. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekaran, A.; Toh, K.Y.; Low, S.H.; Tay, S.K.; Brenner, S.; Goh, D.L. Identification and characterization of novel mouse PDE4D isoforms: Molecular cloning, subcellular distribution and detection of isoform-specific intracellular localization signals. Cell Signal. 2008, 20, 139–153. [Google Scholar] [CrossRef]
- Lynex, C.N.; Li, Z.; Chen, M.L.; Toh, K.Y.; Low, R.W.; Goh, D.L.; Tay, S.K. Identification and molecular characterization of a novel PDE4D11 cAMP-specific phosphodiesterase isoform. Cell Signal. 2008, 20, 2247–2255. [Google Scholar] [CrossRef]
- Yarwood, S.J.; Steele, M.R.; Scotland, G.; Houslay, M.D.; Bolger, G.B. The RACK1 signaling scaffold protein selectively interacts with the cAMP-specific phosphodiesterase PDE4D5 isoform. J. Biol. Chem. 1999, 274, 14909–14917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, G.B.; McCahill, A.; Yarwood, S.J.; Steele, M.S.; Warwicker, J.; Houslay, M.D. Delineation of RAID1, the RACK1 interaction domain located within the unique N-terminal region of the cAMP-specific phosphodiesterase, PDE4D5. BMC Biochem. 2002, 3, 24. [Google Scholar] [CrossRef] [PubMed]
- Bolger, G.B.; Baillie, G.S.; Li, X.; Lynch, M.J.; Herzyk, P.; Mohamed, A.; Mitchell, L.H.; McCahill, A.; Hundsrucker, C.; Klussmann, E.; et al. Scanning peptide array analyses identify overlapping binding sites for the signalling scaffold proteins, beta-arrestin and RACK1, in cAMP-specific phosphodiesterase PDE4D5. Biochem. J. 2006, 398, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Steele, M.R.; McCahill, A.; Thompson, D.S.; MacKenzie, C.; Isaacs, N.W.; Houslay, M.D.; Bolger, G.B. Identification of a surface on the beta-propeller protein RACK1 that interacts with the cAMP-specific phosphodiesterase PDE4D5. Cell Signal. 2001, 13, 507–513. [Google Scholar] [CrossRef]
- Baillie, G.S.; Adams, D.R.; Bhari, N.; Houslay, T.M.; Vadrevu, S.; Meng, D.; Li, X.; Dunlop, A.; Milligan, G.; Bolger, G.B.; et al. Mapping binding sites for the PDE4D5 cAMP-specific phosphodiesterase to the N- and C-domains of beta-arrestin using spot-immobilized peptide arrays. Biochem. J. 2007, 404, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.J.; Baillie, G.S.; Hyde, E.I.; Li, X.; Houslay, T.M.; McCahill, A.; Dunlop, A.J.; Bolger, G.B.; Klussmann, E.; Adams, D.R.; et al. 1H NMR structural and functional characterisation of a cAMP-specific phosphodiesterase-4D5 (PDE4D5) N-terminal region peptide that disrupts PDE4D5 interaction with the signalling scaffold proteins, beta-arrestin and RACK1. Cell Signal. 2007, 19, 2612–2624. [Google Scholar] [CrossRef]
- Bolger, G.B. RACK1 and beta-arrestin2 attenuate dimerization of PDE4 cAMP phosphodiesterase PDE4D5. Cell Signal. 2016, 28, 706–712. [Google Scholar] [CrossRef] [Green Version]
- Perry, S.J.; Baillie, G.S.; Kohout, T.A.; McPhee, I.; Magiera, M.M.; Ang, K.L.; Miller, W.E.; McLean, A.J.; Conti, M.; Houslay, M.D.; et al. Targeting of cyclic AMP degradation to beta 2-adrenergic receptors by beta-arrestins. Science 2002, 298, 834–836. [Google Scholar] [CrossRef]
- Baillie, G.S.; Sood, A.; McPhee, I.; Gall, I.; Perry, S.J.; Lefkowitz, R.J.; Houslay, M.D. β-Arrestin-mediated PDE4 cAMP phosphodiesterase recruitment regulates beta-adrenoceptor switching from Gs to Gi. Proc. Natl. Acad. Sci. USA 2003, 100, 940–945. [Google Scholar] [CrossRef] [Green Version]
- Bolger, G.B.; McCahill, A.; Huston, E.; Cheung, Y.F.; McSorley, T.; Baillie, G.S.; Houslay, M.D. The unique amino-terminal region of the PDE4D5 cAMP phosphodiesterase isoform confers preferential interaction with beta-arrestins. J. Biol. Chem. 2003, 278, 49230–49238. [Google Scholar] [CrossRef] [Green Version]
- Lynch, M.J.; Baillie, G.S.; Mohamed, A.; Li, X.; Maisonneuve, C.; Klussmann, E.; Van Heeke, G.; Houslay, M.D. RNA Silencing Identifies PDE4D5 as the Functionally Relevant cAMP Phosphodiesterase Interacting with {beta}Arrestin to Control the Protein Kinase A/AKAP79-mediated Switching of the {beta}2-Adrenergic Receptor to Activation of ERK in HEK293B2 Cells. J. Biol. Chem. 2005, 280, 33178–33189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richter, W.; Day, P.; Agrawal, R.; Bruss, M.D.; Granier, S.; Wang, Y.L.; Rasmussen, S.G.; Horner, K.; Wang, P.; Lei, T.; et al. Signaling from beta1- and beta2-adrenergic receptors is defined by differential interactions with PDE4. EMBO J. 2008, 27, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Bradaia, A.; Berton, F.; Ferrari, S.; Luscher, C. beta-Arrestin2, interacting with phosphodiesterase 4, regulates synaptic release probability and presynaptic inhibition by opioids. Proc. Natl. Acad. Sci. USA 2005, 102, 3034–3039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Huston, E.; Lynch, M.J.; Houslay, M.D.; Baillie, G.S. Phosphodiesterase-4 influences the PKA phosphorylation status and membrane translocation of G-protein receptor kinase 2 (GRK2) in HEK-293beta2 cells and cardiac myocytes. Biochem. J. 2006, 394, 427–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, R.; Baillie, G.S.; MacKenzie, S.J.; Yarwood, S.J.; Houslay, M.D. The MAP kinase ERK2 inhibits the cyclic AMP-specific phosphodiesterase HSPDE4D3 by phosphorylating it at Ser579. EMBO J. 1999, 18, 893–903. [Google Scholar] [CrossRef] [Green Version]
- MacKenzie, S.J.; Baillie, G.S.; McPhee, I.; Bolger, G.B.; Houslay, M.D. ERK2 mitogen-activated protein kinase binding, phosphorylation, and regulation of the PDE4D cAMP-specific phosphodiesterases. The involvement of COOH-terminal docking sites and NH2-terminal UCR regions. J. Biol. Chem. 2000, 275, 16609–16617. [Google Scholar] [CrossRef] [Green Version]
- Song, R.S.; Massenburg, B.; Wenderski, W.; Jayaraman, V.; Thompson, L.; Neves, S.R. ERK regulation of phosphodiesterase 4 enhances dopamine-stimulated AMPA receptor membrane insertion. Proc. Natl. Acad. Sci. USA 2013, 110, 15437–15442. [Google Scholar] [CrossRef] [Green Version]
- Mackenzie, K.F.; Wallace, D.A.; Hill, E.V.; Anthony, D.F.; Henderson, D.J.; Houslay, D.M.; Arthur, J.S.; Baillie, G.S.; Houslay, M.D. Phosphorylation of cAMP-specific PDE4A5 (phosphodiesterase-4A5) by MK2 (MAPKAPK2) attenuates its activation through protein kinase A phosphorylation. Biochem. J. 2011, 435, 755–769. [Google Scholar] [CrossRef] [Green Version]
- Houslay, K.F.; Christian, F.; MacLeod, R.; Adams, D.R.; Houslay, M.D.; Baillie, G.S. Identification of a multifunctional docking site on the catalytic unit of phosphodiesterase-4 (PDE4) that is utilised by multiple interaction partners. Biochem. J. 2017, 474, 597–609. [Google Scholar] [CrossRef] [Green Version]
- Sette, C.; Conti, M. Phosphorylation and activation of a cAMP-specific phosphodiesterase by the cAMP-dependent protein kinase. Involvement of serine 54 in the enzyme activation. J. Biol. Chem. 1996, 271, 16526–16534. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, R.; Wilkinson, I.R.; McCallum, J.F.; Engels, P.; Houslay, M.D. cAMP-specific phosphodiesterase HSPDE4D3 mutants which mimic activation and changes in rolipram inhibition triggered by protein kinase A phosphorylation of Ser-54: Generation of a molecular model. Biochem. J. 1998, 333, 139–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oki, N.; Takahashi, S.I.; Hidaka, H.; Conti, M. Short term feedback regulation of cAMP in FRTL-5 thyroid cells. Role of PDE4D3 phosphodiesterase activation. J. Biol. Chem. 2000, 275, 10831–10837. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, S.J.; Baillie, G.S.; McPhee, I.; MacKenzie, C.; Seamons, R.; McSorley, T.; Millen, J.; Beard, M.B.; Van Heeke, G.; Houslay, M.D. Long PDE4 cAMP specific phosphodiesterases are activated by protein kinase A-mediated phosphorylation of a single serine residue in Upstream Conserved Region 1 (UCR1). Br. J. Pharmacol. 2002, 136, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Hill, E.V.; Sheppard, C.L.; Cheung, Y.F.; Gall, I.; Krause, E.; Houslay, M.D. Oxidative stress employs phosphatidyl inositol 3-kinase and ERK signalling pathways to activate cAMP phosphodiesterase-4D3 (PDE4D3) through multi-site phosphorylation at Ser239 and Ser579. Cell Signal. 2006, 18, 2056–2069. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.X.; Hassell, A.M.; Vanderwall, D.; Lambert, M.H.; Holmes, W.D.; Luther, M.A.; Rocque, W.J.; Milburn, M.V.; Zhao, Y.; Ke, H.; et al. Atomic structure of PDE4: Insights into phosphodiesterase mechanism and specificity. Science 2000, 288, 1822–1825. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Peng, M.S.; Chen, Y.; Geng, J.; Robinson, H.; Houslay, M.D.; Cai, J.; Ke, H. Structures of the four subfamilies of phosphodiesterase-4 provide insight into the selectivity of their inhibitors. Biochem. J. 2007, 408, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Cedervall, P.; Aulabaugh, A.; Geoghegan, K.F.; McLellan, T.J.; Pandit, J. Engineered stabilization and structural analysis of the autoinhibited conformation of PDE4. Proc. Natl. Acad. Sci. USA 2015, 112, E1414–E1422. [Google Scholar] [CrossRef] [Green Version]
- Bolger, G.B.; Dunlop, A.J.; Meng, D.; Day, J.P.; Klussmann, E.; Baillie, G.S.; Adams, D.R.; Houslay, M.D. Dimerization of cAMP phosphodiesterase-4 (PDE4) in living cells requires interfaces located in both the UCR1 and catalytic unit domains. Cell Signal. 2015, 27, 756–769. [Google Scholar] [CrossRef] [Green Version]
- Richter, W.; Conti, M. Dimerization of the type 4 cAMP-specific phosphodiesterases is mediated by the upstream conserved regions (UCRs). J. Biol. Chem. 2002, 277, 40212–40221. [Google Scholar] [CrossRef] [Green Version]
- Richter, W.; Conti, M. The oligomerization state determines regulatory properties and inhibitor sensitivity of type 4 cAMP-specific phosphodiesterases. J. Biol. Chem. 2004, 279, 30338–30348. [Google Scholar] [CrossRef] [Green Version]
- Xie, M.; Blackman, B.; Scheitrum, C.; Mika, D.; Blanchard, E.; Lei, T.; Conti, M.; Richter, W. The upstream conserved regions (UCRs) mediate homo- and hetero-oligomerization of type 4 cyclic nucleotide phosphodiesterases (PDE4s). Biochem. J. 2014, 459, 539–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanchard, E.; Zlock, L.; Lao, A.; Mika, D.; Namkung, W.; Xie, M.; Scheitrum, C.; Gruenert, D.C.; Verkman, A.S.; Finkbeiner, W.E.; et al. Anchored PDE4 regulates chloride conductance in wild-type and DeltaF508-CFTR human airway epithelia. FASEB J. 2014, 28, 791–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, S.L.; van Groen, T.; Kadish, I.; Smoot, L.H.M.; Bolger, G.B. Altered phosphorylation, electrophysiology, and behavior on attenuation of PDE4B action in hippocampus. BMC Neurosci. 2017, 18, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGirr, A.; Lipina, T.V.; Mun, H.S.; Georgiou, J.; Al-Amri, A.H.; Ng, E.; Zhai, D.; Elliott, C.; Cameron, R.T.; Mullins, J.G.; et al. Specific Inhibition of Phosphodiesterase-4B Results in Anxiolysis and Facilitates Memory Acquisition. Neuropsychopharmacology 2016, 41, 1080–1092. [Google Scholar] [CrossRef] [PubMed]
- Havekes, R.; Park, A.J.; Tolentino, R.E.; Bruinenberg, V.M.; Tudor, J.C.; Lee, Y.; Hansen, R.T.; Guercio, L.A.; Linton, E.; Neves-Zaph, S.R.; et al. Compartmentalized PDE4A5 Signaling Impairs Hippocampal Synaptic Plasticity and Long-Term Memory. J. Neurosci. 2016, 36, 8936–8946. [Google Scholar] [CrossRef] [Green Version]
- Havekes, R.; Park, A.J.; Tudor, J.C.; Luczak, V.G.; Hansen, R.T.; Ferri, S.L.; Bruinenberg, V.M.; Poplawski, S.G.; Day, J.P.; Aton, S.J.; et al. Sleep deprivation causes memory deficits by negatively impacting neuronal connectivity in hippocampal area CA1. Elife 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.T.; Huang, Y.; Jin, S.L.; Frith, S.A.; Suvarna, N.; Conti, M.; O’Donnell, J.M. Antidepressant-like profile and reduced sensitivity to rolipram in mice deficient in the PDE4D phosphodiesterase enzyme. Neuropsychopharmacology 2002, 27, 587–595. [Google Scholar] [CrossRef] [Green Version]
- Siuciak, J.A.; Chapin, D.S.; McCarthy, S.A.; Martin, A.N. Antipsychotic profile of rolipram: Efficacy in rats and reduced sensitivity in mice deficient in the phosphodiesterase-4B (PDE4B) enzyme. Psychopharmacology (Berl) 2007, 192, 415–424. [Google Scholar] [CrossRef]
- Siuciak, J.A.; McCarthy, S.A.; Chapin, D.S.; Martin, A.N. Behavioral and neurochemical characterization of mice deficient in the phosphodiesterase-4B (PDE4B) enzyme. Psychopharmacology (Berl) 2008, 197, 115–126. [Google Scholar] [CrossRef]
- Zhang, H.T.; Huang, Y.; Masood, A.; Stolinski, L.R.; Li, Y.; Zhang, L.; Dlaboga, D.; Jin, S.L.; Conti, M.; O’Donnell, J.M. Anxiogenic-like behavioral phenotype of mice deficient in phosphodiesterase 4B (PDE4B). Neuropsychopharmacology 2008, 33, 1611–1623. [Google Scholar] [CrossRef] [Green Version]
- Rutten, K.; Wallace, T.L.; Works, M.; Prickaerts, J.; Blokland, A.; Novak, T.J.; Santarelli, L.; Misner, D.L. Enhanced long-term depression and impaired reversal learning in phosphodiesterase 4B-knockout (PDE4B-/-) mice. Neuropharmacology 2011, 61, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.F.; Cheng, Y.F.; Huang, Y.; Conti, M.; Wilson, S.P.; O’Donnell, J.M.; Zhang, H.T. Phosphodiesterase-4D knock-out and RNA interference-mediated knock-down enhance memory and increase hippocampal neurogenesis via increased cAMP signaling. J. Neurosci. 2011, 31, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Hansen, R.T., III; Conti, M.; Zhang, H.T. Mice deficient in phosphodiesterase-4A display anxiogenic-like behavior. Psychopharmacology (Berl) 2014, 231, 2941–2954. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, T.L.; Braun, A.A.; Amos-Kroohs, R.M.; Williams, M.T.; Ostertag, E.; Vorhees, C.V. A new model of Pde4d deficiency: Genetic knock-down of PDE4D enzyme in rats produces an antidepressant phenotype without spatial cognitive effects. Genes Brain Behav. 2012, 11, 614–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.Z.; Zhang, Y.; Liu, Y.Q.; Zhao, N.; Zhang, Y.Z.; Yuan, L.; An, L.; Li, J.; Wang, X.Y.; Qin, J.J.; et al. RNA interference-mediated phosphodiesterase 4D splice variants knock-down in the prefrontal cortex produces antidepressant-like and cognition-enhancing effects. Br. J. Pharmacol. 2013, 168, 1001–1014. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Z.; Yang, W.X.; Zhang, Y.; Zhao, N.; Zhang, Y.Z.; Liu, Y.Q.; Xu, Y.; Wilson, S.P.; O’Donnell, J.M.; Zhang, H.T.; et al. Phosphodiesterase-4D Knock-down in the Prefrontal Cortex Alleviates Chronic Unpredictable Stress-Induced Depressive-Like Behaviors and Memory Deficits in Mice. Sci. Rep. 2015, 5, 11332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Cheng, Y.; Wang, H.; Wang, C.; Wilson, S.P.; Xu, J.; Zhang, H.T. RNA interference-mediated knockdown of long-form phosphodiesterase-4D (PDE4D) enzyme reverses amyloid-beta42-induced memory deficits in mice. J. Alzheimers. Dis. 2014, 38, 269–280. [Google Scholar] [CrossRef]
- Xiang, Y.; Naro, F.; Zoudilova, M.; Jin, S.L.; Conti, M.; Kobilka, B. Phosphodiesterase 4D is required for beta2 adrenoceptor subtype-specific signaling in cardiac myocytes. Proc. Natl. Acad. Sci. USA 2005, 102, 909–914. [Google Scholar] [CrossRef] [Green Version]
- Nino, G.; Hu, A.; Grunstein, J.S.; Grunstein, M.M. Mechanism regulating proasthmatic effects of prolonged homologous beta2-adrenergic receptor desensitization in airway smooth muscle. Am. J. Physiol. Lung Cell Mol. Physiol. 2009, 297, L746–L757. [Google Scholar] [CrossRef] [Green Version]
- Hu, A.; Nino, G.; Grunstein, J.S.; Fatma, S.; Grunstein, M.M. Prolonged heterologous beta2-adrenoceptor desensitization promotes proasthmatic airway smooth muscle function via PKA/ERK1/2-mediated phosphodiesterase-4 induction. Am. J. Physiol. Lung Cell Mol. Physiol. 2008, 294, L1055–L1067. [Google Scholar] [CrossRef] [Green Version]
- Billington, C.K.; Le Jeune, I.R.; Young, K.W.; Hall, I.P. A major functional role for phosphodiesterase 4D5 in human airway smooth muscle cells. Am. J. Respir. Cell Mol. Biol. 2008, 38, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Niimi, K.; Ge, Q.; Moir, L.M.; Ammit, A.J.; Trian, T.; Burgess, J.K.; Black, J.L.; Oliver, B.G. β2-Agonists upregulate PDE4 mRNA but not protein or activity in human airway smooth muscle cells from asthmatic and nonasthmatic volunteers. Am. J. Physiol. Lung Cell Mol. Physiol. 2012, 302, L334–L342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrels, B.; Sandilands, E.; Serrels, A.; Baillie, G.; Houslay, M.D.; Brunton, V.G.; Canel, M.; Machesky, L.M.; Anderson, K.I.; Frame, M.C. A complex between FAK, RACK1, and PDE4D5 controls spreading initiation and cancer cell polarity. Curr. Biol. 2010, 20, 1086–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, S.; Budatha, M.; Dahlman, J.E.; Coon, B.G.; Cameron, R.T.; Langer, R.; Anderson, D.G.; Baillie, G.; Schwartz, M.A. Interaction between integrin alpha5 and PDE4D regulates endothelial inflammatory signalling. Nat. Cell Biol. 2016, 18, 1043–1053. [Google Scholar] [CrossRef] [PubMed]
- Mayford, M.; Baranes, D.; Podsypanina, K.; Kandel, E.R. The 3′-untranslated region of CaMKII alpha is a cis-acting signal for the localization and translation of mRNA in dendrites. Proc. Natl. Acad. Sci. USA 1996, 93, 13250–13255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayford, M.; Bach, M.E.; Huang, Y.Y.; Wang, L.; Hawkins, R.D.; Kandel, E.R. Control of memory formation through regulated expression of a CaMKII transgene. Science 1996, 274, 1678–1683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsien, J.Z.; Chen, D.F.; Gerber, D.; Tom, C.; Mercer, E.H.; Anderson, D.J.; Mayford, M.; Kandel, E.R.; Tonegawa, S. Subregion- and cell type-restricted gene knockout in mouse brain. Cell 1996, 87, 1317–1326. [Google Scholar] [CrossRef] [Green Version]
- Hatcher, J.P.; Jones, D.N.; Rogers, D.C.; Hatcher, P.D.; Reavill, C.; Hagan, J.J.; Hunter, A.J. Development of SHIRPA to characterise the phenotype of gene-targeted mice. Behav. Brain Res. 2001, 125, 43–47. [Google Scholar] [CrossRef]
- Rogers, D.C.; Fisher, E.M.; Brown, S.D.; Peters, J.; Hunter, A.J.; Martin, J.E. Behavioral and functional analysis of mouse phenotype: SHIRPA, a proposed protocol for comprehensive phenotype assessment. Mamm. Genome 1997, 8, 711–713. [Google Scholar] [CrossRef]
- Millar, J.K.; Pickard, B.S.; Mackie, S.; James, R.; Christie, S.; Buchanan, S.R.; Malloy, M.P.; Chubb, J.E.; Huston, E.; Baillie, G.S.; et al. DISC1 and PDE4B are interacting genetic factors in schizophrenia that regulate cAMP signaling. Science 2005, 310, 1187–1191. [Google Scholar] [CrossRef]
- Bradshaw, N.J.; Soares, D.C.; Carlyle, B.C.; Ogawa, F.; Davidson-Smith, H.; Christie, S.; Mackie, S.; Thomson, P.A.; Porteous, D.J.; Millar, J.K. PKA phosphorylation of NDE1 is DISC1/PDE4 dependent and modulates its interaction with LIS1 and NDEL1. J. Neurosci. 2011, 31, 9043–9054. [Google Scholar] [CrossRef] [PubMed]
- Murdoch, H.; Mackie, S.; Collins, D.M.; Hill, E.V.; Bolger, G.B.; Klussmann, E.; Porteous, D.J.; Millar, J.K.; Houslay, M.D. Isoform-selective susceptibility of DISC1/phosphodiesterase-4 complexes to dissociation by elevated intracellular cAMP levels. J. Neurosci. 2007, 27, 9513–9524. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, N.J.; Ogawa, F.; Antolin-Fontes, B.; Chubb, J.E.; Carlyle, B.C.; Christie, S.; Claessens, A.; Porteous, D.J.; Millar, J.K. DISC1, PDE4B, and NDE1 at the centrosome and synapse. Biochem. Biophys. Res. Commun. 2008, 377, 1091–1096. [Google Scholar] [CrossRef] [PubMed]
- Soda, T.; Frank, C.; Ishizuka, K.; Baccarella, A.; Park, Y.U.; Flood, Z.; Park, S.K.; Sawa, A.; Tsai, L.H. DISC1-ATF4 transcriptional repression complex: Dual regulation of the cAMP-PDE4 cascade by DISC1. Mol. Psychiatry 2013, 18, 898–908. [Google Scholar] [CrossRef] [Green Version]
- Clapcote, S.J.; Lipina, T.V.; Millar, J.K.; Mackie, S.; Christie, S.; Ogawa, F.; Lerch, J.P.; Trimble, K.; Uchiyama, M.; Sakuraba, Y.; et al. Behavioral phenotypes of Disc1 missense mutations in mice. Neuron 2007, 54, 387–402. [Google Scholar] [CrossRef] [Green Version]
- Lipina, T.V.; Wang, M.; Liu, F.; Roder, J.C. Synergistic interactions between PDE4B and GSK-3: DISC1 mutant mice. Neuropharmacology 2012, 62, 1252–1262. [Google Scholar] [CrossRef]
- Selcher, J.C.; Nekrasova, T.; Paylor, R.; Landreth, G.E.; Sweatt, J.D. Mice lacking the ERK1 isoform of MAP kinase are unimpaired in emotional learning. Learn. Mem. 2001, 8, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Bourtchuladze, R.; Frenguelli, B.; Blendy, J.; Cioffi, D.; Schutz, G.; Silva, A.J. Deficient long-term memory in mice with a targeted mutation of the cAMP-responsive element-binding protein. Cell 1994, 79, 59–68. [Google Scholar] [CrossRef]
- Kida, S.; Josselyn, S.A.; de Ortiz, S.P.; Kogan, J.H.; Chevere, I.; Masushige, S.; Silva, A.J. CREB required for the stability of new and reactivated fear memories. Nat. Neurosci. 2002, 5, 348–355. [Google Scholar] [CrossRef]
- Barco, A.; Alarcon, J.M.; Kandel, E.R. Expression of constitutively active CREB protein facilitates the late phase of long-term potentiation by enhancing synaptic capture. Cell 2002, 108, 689–703. [Google Scholar] [CrossRef] [Green Version]
- Pittenger, C.; Huang, Y.Y.; Paletzki, R.F.; Bourtchouladze, R.; Scanlin, H.; Vronskaya, S.; Kandel, E.R. Reversible inhibition of CREB/ATF transcription factors in region CA1 of the dorsal hippocampus disrupts hippocampus-dependent spatial memory. Neuron 2002, 34, 447–462. [Google Scholar] [CrossRef] [Green Version]
- Kogan, J.H.; Frankland, P.W.; Blendy, J.A.; Coblentz, J.; Marowitz, Z.; Schutz, G.; Silva, A.J. Spaced training induces normal long-term memory in CREB mutant mice. Curr. Biol. 1997, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.L.; Thomas, S.A.; Villacres, E.C.; Xia, Z.; Simmons, M.L.; Chavkin, C.; Palmiter, R.D.; Storm, D.R. Altered behavior and long-term potentiation in type I adenylyl cyclase mutant mice. Proc. Natl. Acad. Sci. USA 1995, 92, 220–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Shu, X.; Liu, D.; Shang, Y.; Wu, Y.; Pei, L.; Xu, X.; Tian, Q.; Zhang, J.; Qian, K.; et al. EPAC null mutation impairs learning and social interactions via aberrant regulation of miR-124 and Zif268 translation. Neuron 2012, 73, 774–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, D.P.; Jones, K.A.; Woolfrey, K.M.; Burgdorf, J.; Russell, T.A.; Kalmbach, A.; Lee, H.; Yang, C.; Bradberry, M.M.; Wokosin, D.; et al. Social, communication, and cortical structural impairments in Epac2-deficient mice. J. Neurosci. 2012, 32, 11864–11878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, R.G.; Garrud, P.; Rawlins, J.N.; O’Keefe, J. Place navigation impaired in rats with hippocampal lesions. Nature 1982, 297, 681–683. [Google Scholar] [CrossRef]
- Abel, T.; Nguyen, P.V.; Barad, M.; Deuel, T.A.; Kandel, E.R.; Bourtchouladze, R. Genetic demonstration of a role for PKA in the late phase of LTP and in hippocampus-based long-term memory. Cell 1997, 88, 615–626. [Google Scholar] [CrossRef] [Green Version]
- Pittenger, C.; Fasano, S.; Mazzocchi-Jones, D.; Dunnett, S.B.; Kandel, E.R.; Brambilla, R. Impaired bidirectional synaptic plasticity and procedural memory formation in striatum-specific cAMP response element-binding protein-deficient mice. J. Neurosci. 2006, 26, 2808–2813. [Google Scholar] [CrossRef] [Green Version]
- Ahn, S.; Ginty, D.D.; Linden, D.J. A late phase of cerebellar long-term depression requires activation of CaMKIV and CREB. Neuron 1999, 23, 559–568. [Google Scholar] [CrossRef] [Green Version]
- Lonze, B.E.; Riccio, A.; Cohen, S.; Ginty, D.D. Apoptosis, axonal growth defects, and degeneration of peripheral neurons in mice lacking CREB. Neuron 2002, 34, 371–385. [Google Scholar] [CrossRef] [Green Version]
- Burgin, K.E.; Waxham, M.N.; Rickling, S.; Westgate, S.A.; Mobley, W.C.; Kelly, P.T. In situ hybridization histochemistry of Ca2+/calmodulin-dependent protein kinase in developing rat brain. J. Neurosci. 1990, 10, 1788–1798. [Google Scholar] [CrossRef] [PubMed]
- Mayford, M.; Wang, J.; Kandel, E.R.; O’Dell, T.J. CaMKII regulates the frequency-response function of hippocampal synapses for the production of both LTD and LTP. Cell 1995, 81, 891–904. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Deng, K.; Hou, J.; Bryson, J.B.; Barco, A.; Nikulina, E.; Spencer, T.; Mellado, W.; Kandel, E.R.; Filbin, M.T. Activated CREB is sufficient to overcome inhibitors in myelin and promote spinal axon regeneration in vivo. Neuron 2004, 44, 609–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Detke, M.J.; Johnson, J.; Lucki, I. Acute and chronic antidepressant drug treatment in the rat forced swimming test model of depression. Exp. Clin. Psychopharmacol. 1997, 5, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Hikida, T.; Jaaro-Peled, H.; Seshadri, S.; Oishi, K.; Hookway, C.; Kong, S.; Wu, D.; Xue, R.; Andrade, M.; Tankou, S.; et al. Dominant-negative DISC1 transgenic mice display schizophrenia-associated phenotypes detected by measures translatable to humans. Proc. Natl. Acad. Sci. USA 2007, 104, 14501–14506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, S.L.; Richard, F.J.; Kuo, W.P.; D’Ercole, A.J.; Conti, M. Impaired growth and fertility of cAMP-specific phosphodiesterase PDE4D-deficient mice. Proc. Natl. Acad. Sci. USA 1999, 96, 11998–12003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, S.L.; Conti, M. Induction of the cyclic nucleotide phosphodiesterase PDE4B is essential for LPS-activated TNF-alpha responses. Proc. Natl. Acad. Sci. USA 2002, 99, 7628–7633. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Graham, J.M., Jr.; Rimoin, D.L.; Lachman, R.S.; Krejci, P.; Tompson, S.W.; Nelson, S.F.; Krakow, D.; Cohn, D.H. Exome sequencing identifies PDE4D mutations in acrodysostosis. Am. J. Hum. Genet. 2012, 90, 746–751. [Google Scholar] [CrossRef] [Green Version]
- Linglart, A.; Fryssira, H.; Hiort, O.; Holterhus, P.M.; Perez de, N.G.; Argente, J.; Heinrichs, C.; Kuechler, A.; Mantovani, G.; Leheup, B.; et al. PRKAR1A and PDE4D Mutations Cause Acrodysostosis but Two Distinct Syndromes with or without GPCR-Signaling Hormone Resistance. J. Clin. Endocrinol. Metab. 2012, 97, E2328–E2338. [Google Scholar] [CrossRef] [Green Version]
- Michot, C.; Le, G.C.; Goldenberg, A.; Abhyankar, A.; Klein, C.; Kinning, E.; Guerrot, A.M.; Flahaut, P.; Duncombe, A.; Baujat, G.; et al. Exome sequencing identifies PDE4D mutations as another cause of acrodysostosis. Am. J. Hum. Genet. 2012, 90, 740–745. [Google Scholar] [CrossRef] [Green Version]
- Lynch, D.C.; Dyment, D.A.; Huang, L.; Nikkel, S.M.; Lacombe, D.; Campeau, P.M.; Lee, B.; Bacino, C.A.; Michaud, J.L.; Bernier, F.P.; et al. Identification of novel mutations confirms PDE4D as a major gene causing acrodysostosis. Hum. Mutat. 2013, 34, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Lindstrand, A.; Grigelioniene, G.; Nilsson, D.; Pettersson, M.; Hofmeister, W.; Anderlid, B.M.; Kant, S.G.; Ruivenkamp, C.A.; Gustavsson, P.; Valta, H.; et al. Different mutations in PDE4D associated with developmental disorders with mirror phenotypes. J. Med. Genet. 2014, 51, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Brullo, C.; Ricciarelli, R.; Prickaerts, J.; Arancio, O.; Massa, M.; Rotolo, C.; Romussi, A.; Rebosio, C.; Marengo, B.; Pronzato, M.A.; et al. New insights into selective PDE4D inhibitors: 3-(Cyclopentyloxy)-4-methoxybenzaldehyde O-(2-(2,6-dimethylmorpholino)-2-oxoethyl) oxime (GEBR-7b) structural development and promising activities to restore memory impairment. Eur. J. Med. Chem. 2016, 124, 82–102. [Google Scholar] [CrossRef] [PubMed]
- D’Ursi, P.; Guariento, S.; Trombetti, G.; Orro, A.; Cichero, E.; Milanesi, L.; Fossa, P.; Bruno, O. Further Insights in the Binding Mode of Selective Inhibitors to Human PDE4D Enzyme Combining Docking and Molecular Dynamics. Mol. Inform. 2016, 35, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Li, Z.; Huang, Y.Y.; Wu, D.; Luo, H.B. Novel Phosphodiesterase Inhibitors for Cognitive Improvement in Alzheimer’s Disease. J. Med. Chem. 2018. [Google Scholar] [CrossRef]
- Robichaud, A.; Stamatiou, P.B.; Jin, S.L.; Lachance, N.; Macdonald, D.; Laliberte, F.; Liu, S.; Huang, Z.; Conti, M.; Chan, C.C. Deletion of phosphodiesterase 4D in mice shortens alpha(2)-adrenoceptor-mediated anesthesia, a behavioral correlate of emesis. J. Clin. Invest. 2002, 110, 1045–1052. [Google Scholar] [CrossRef]
- Kreis, T.E. Microinjected antibodies against the cytoplasmic domain of vesicular stomatitis virus glycoprotein block its transport to the cell surface. EMBO J. 1986, 5, 931–941. [Google Scholar] [CrossRef]
- Shepherd, M.; McSorley, T.; Olsen, A.E.; Johnston, L.A.; Thomson, N.C.; Baillie, G.S.; Houslay, M.D.; Bolger, G.B. Molecular cloning and subcellular distribution of the novel PDE4B4 cAMP-specific phosphodiesterase isoform. Biochem. J. 2003, 370, 429–438. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Conti, M.; Houslay, M.D.; Farooqui, S.M.; Chen, M.; O’Donnell, J.M. Noradrenergic activity differentially regulates the expression of rolipram-sensitive, high-affinity cyclic AMP phosphodiesterase (PDE4) in rat brain. J. Neurochem. 1997, 69, 2397–2404. [Google Scholar] [CrossRef]
- Shalin, S.C.; Hernandez, C.M.; Dougherty, M.K.; Morrison, D.K.; Sweatt, J.D. Kinase suppressor of Ras1 compartmentalizes hippocampal signal transduction and subserves synaptic plasticity and memory formation. Neuron 2006, 50, 765–779. [Google Scholar] [CrossRef] [Green Version]
- Shalin, S.C.; Zirrgiebel, U.; Honsa, K.J.; Julien, J.P.; Miller, F.D.; Kaplan, D.R.; Sweatt, J.D. Neuronal MEK is important for normal fear conditioning in mice. J. Neurosci. Res. 2004, 75, 760–770. [Google Scholar] [CrossRef] [PubMed]
- Paylor, R.; Crawley, J.N. Inbred strain differences in prepulse inhibition of the mouse startle response. Psychopharmacology (Berl) 1997, 132, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, R.J.; Dalvi, A. Anxiety, defence and the elevated plus-maze. Neurosci. Biobehav. Rev. 1997, 21, 801–810. [Google Scholar] [CrossRef]
- Kim, J.J.; Fanselow, M.S. Modality-specific retrograde amnesia of fear. Science 1992, 256, 675–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levenson, J.M.; O’Riordan, K.J.; Brown, K.D.; Trinh, M.A.; Molfese, D.L.; Sweatt, J.D. Regulation of histone acetylation during memory formation in the hippocampus. J. Biol. Chem. 2004, 279, 40545–40559. [Google Scholar] [CrossRef] [Green Version]
- Moretti, P.; Levenson, J.M.; Battaglia, F.; Atkinson, R.; Teague, R.; Antalffy, B.; Armstrong, D.; Arancio, O.; Sweatt, J.D.; Zoghbi, H.Y. Learning and memory and synaptic plasticity are impaired in a mouse model of Rett syndrome. J. Neurosci. 2006, 26, 319–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, C.A.; Sweatt, J.D. Covalent modification of DNA regulates memory formation. Neuron 2007, 53, 857–869. [Google Scholar] [CrossRef] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bolger, G.B.; Smoot, L.H.M.; van Groen, T. Dominant-Negative Attenuation of cAMP-Selective Phosphodiesterase PDE4D Action Affects Learning and Behavior. Int. J. Mol. Sci. 2020, 21, 5704. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165704
Bolger GB, Smoot LHM, van Groen T. Dominant-Negative Attenuation of cAMP-Selective Phosphodiesterase PDE4D Action Affects Learning and Behavior. International Journal of Molecular Sciences. 2020; 21(16):5704. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165704
Chicago/Turabian StyleBolger, Graeme B., Lisa High Mitchell Smoot, and Thomas van Groen. 2020. "Dominant-Negative Attenuation of cAMP-Selective Phosphodiesterase PDE4D Action Affects Learning and Behavior" International Journal of Molecular Sciences 21, no. 16: 5704. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165704