Induction of Glucoraphasatin Biosynthesis Genes by MYB29 in Radish (Raphanus sativus L.) Roots

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
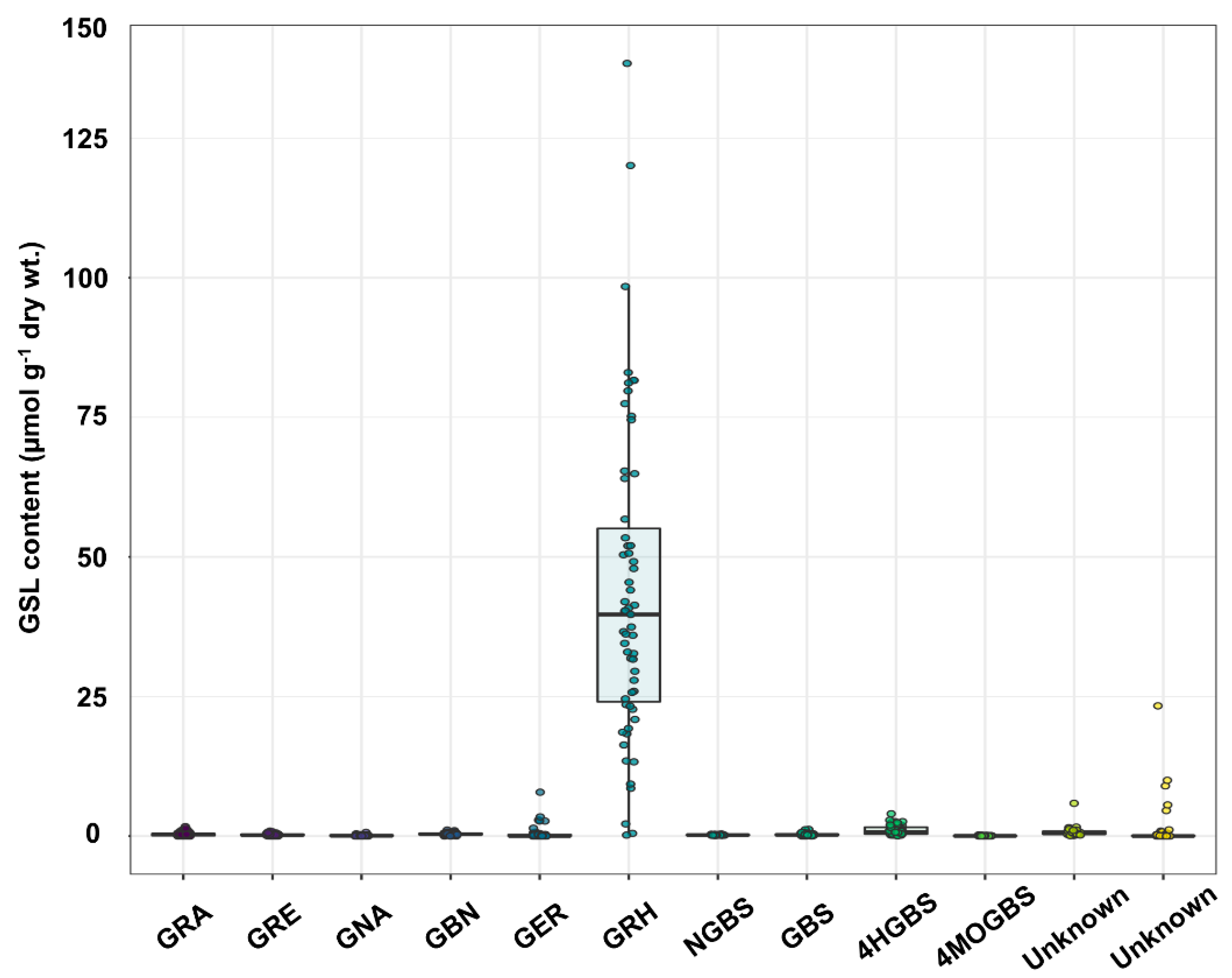
2.1. GSL Profiles in Radish Roots
2.2. RNA Sequencing and Mapping of Radish Reference Genomes
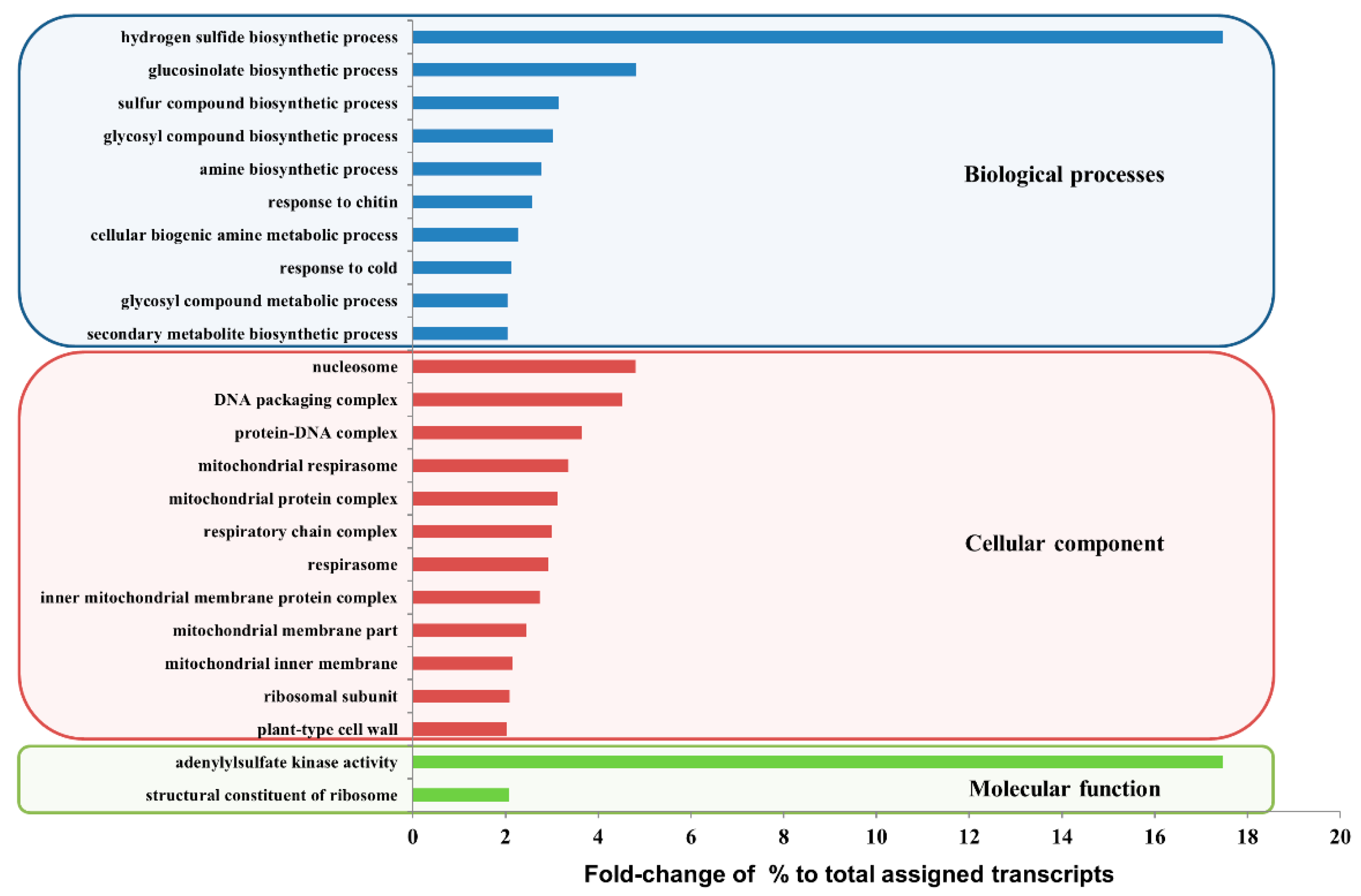
2.3. Analysis of Differentially Expressed Genes between Roots of the Radish Accessions
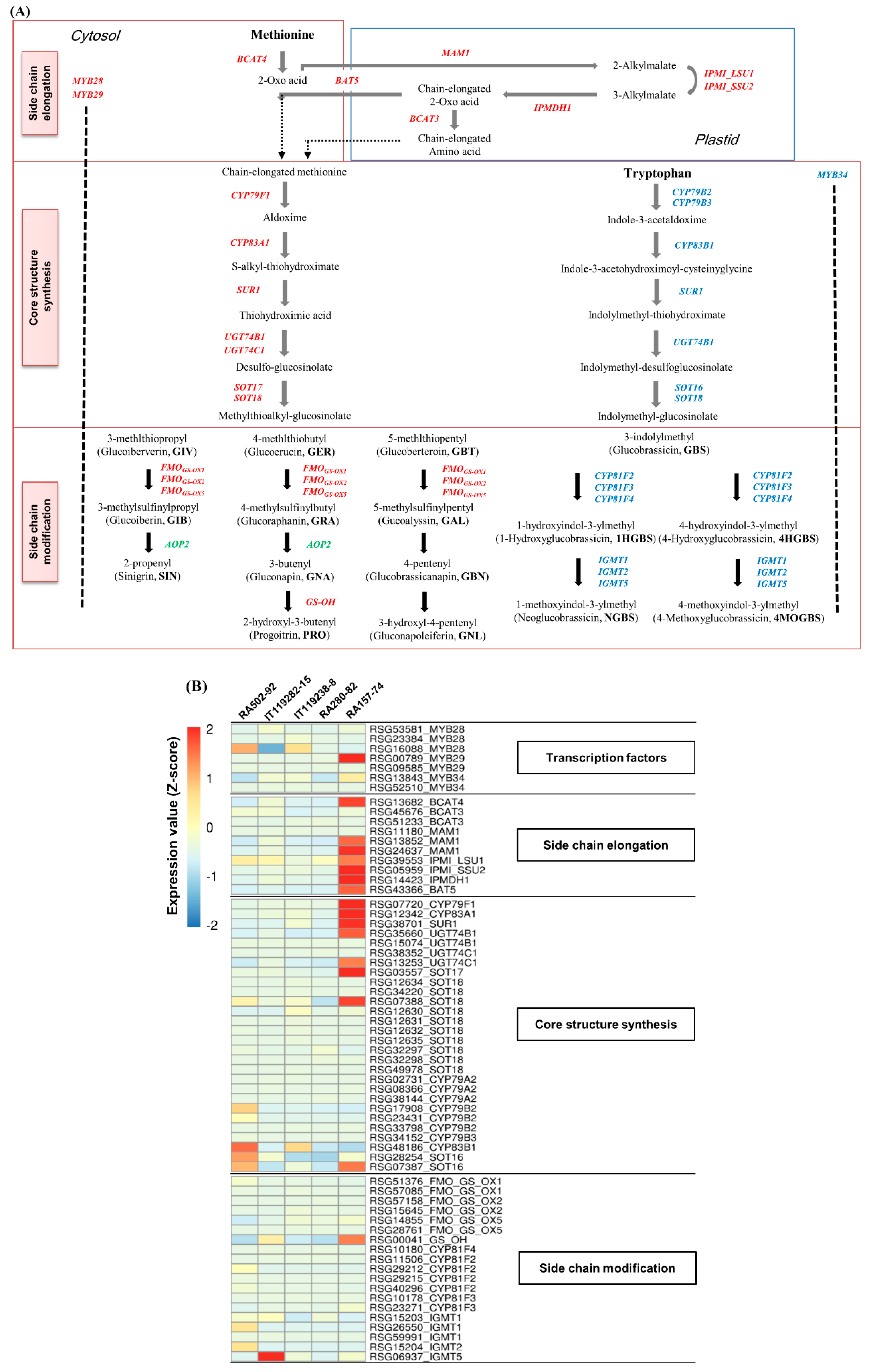
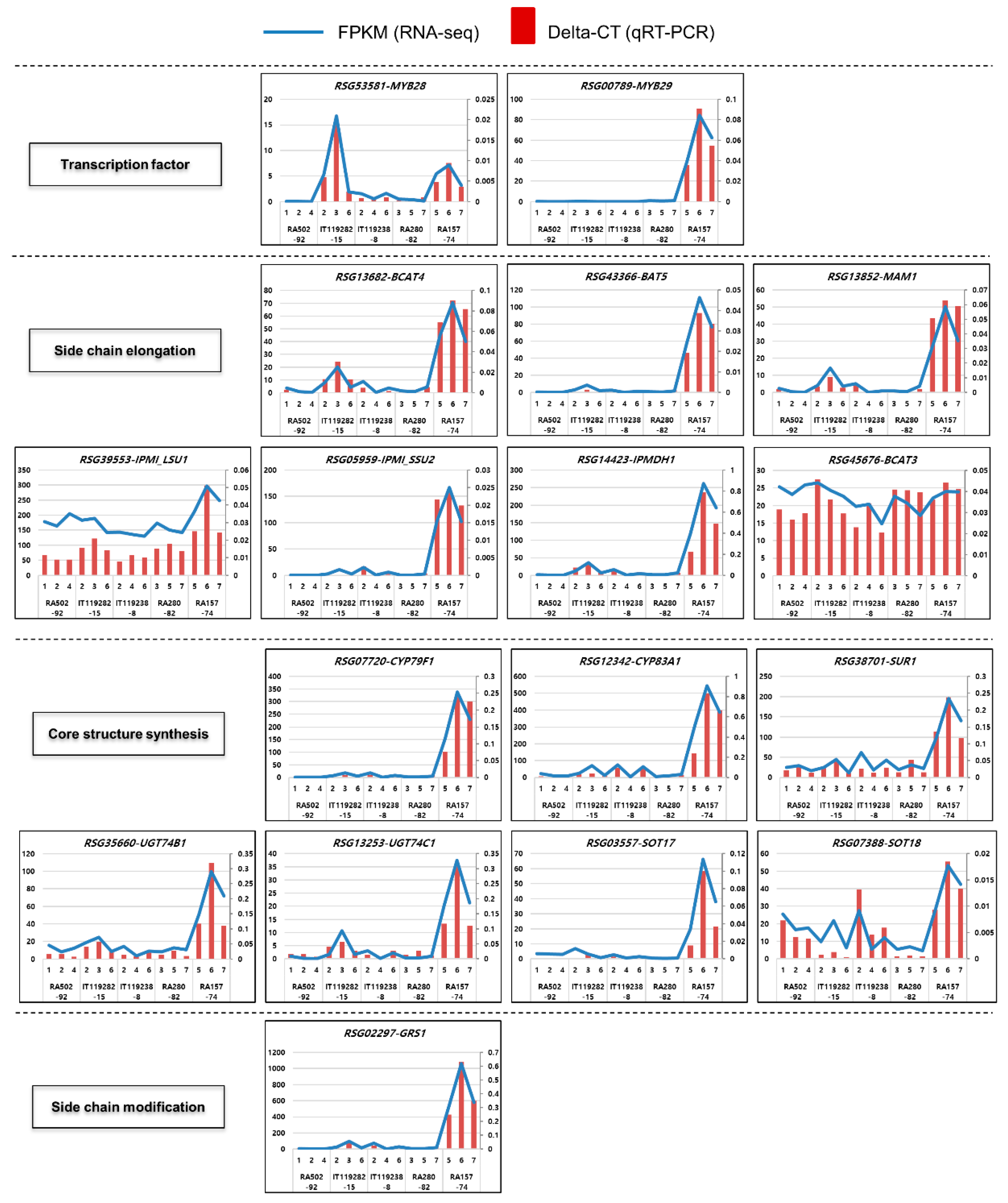
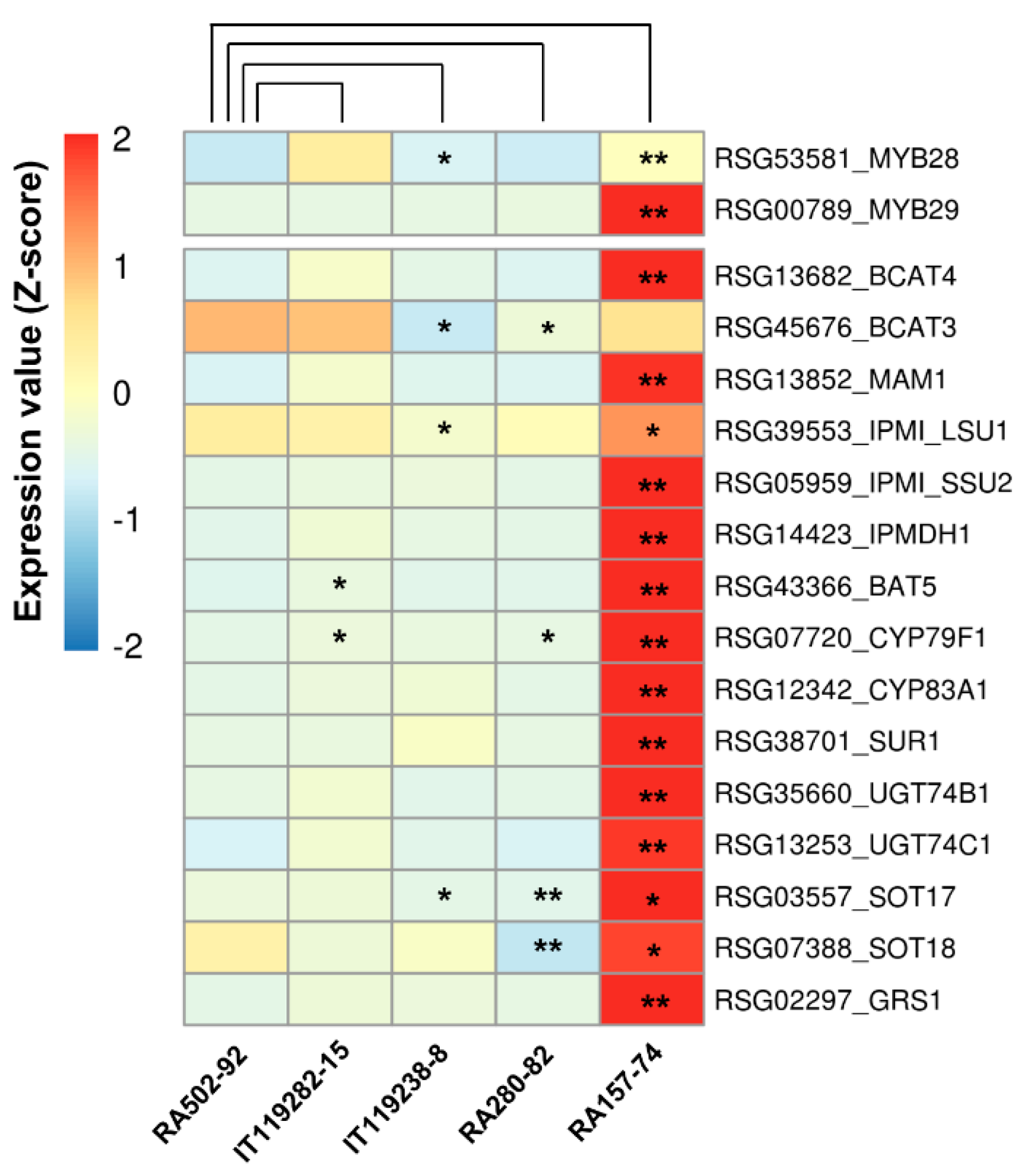
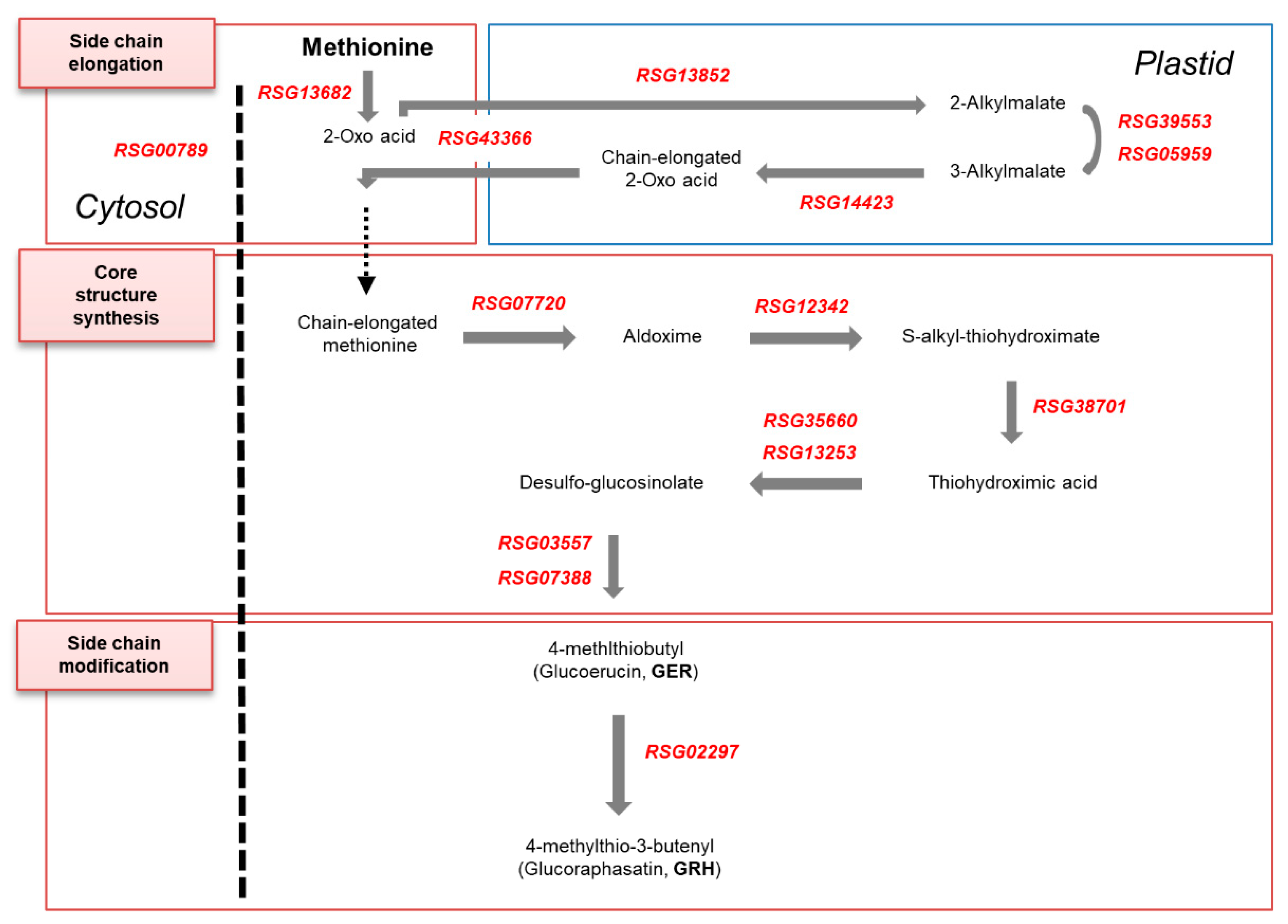
2.4. Identification and Expression Analysis of GSL Biosynthesis Genes in Radish Roots
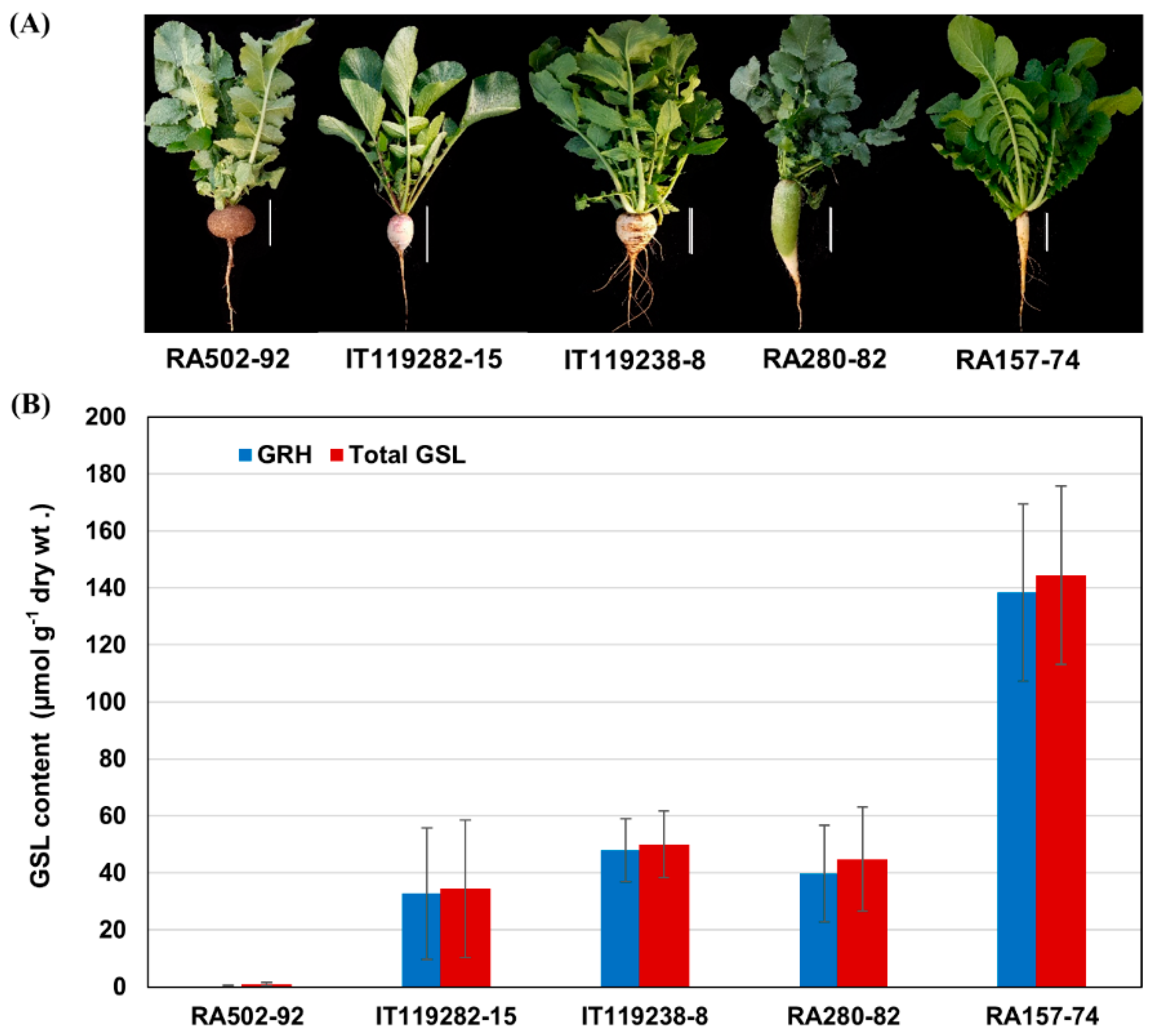
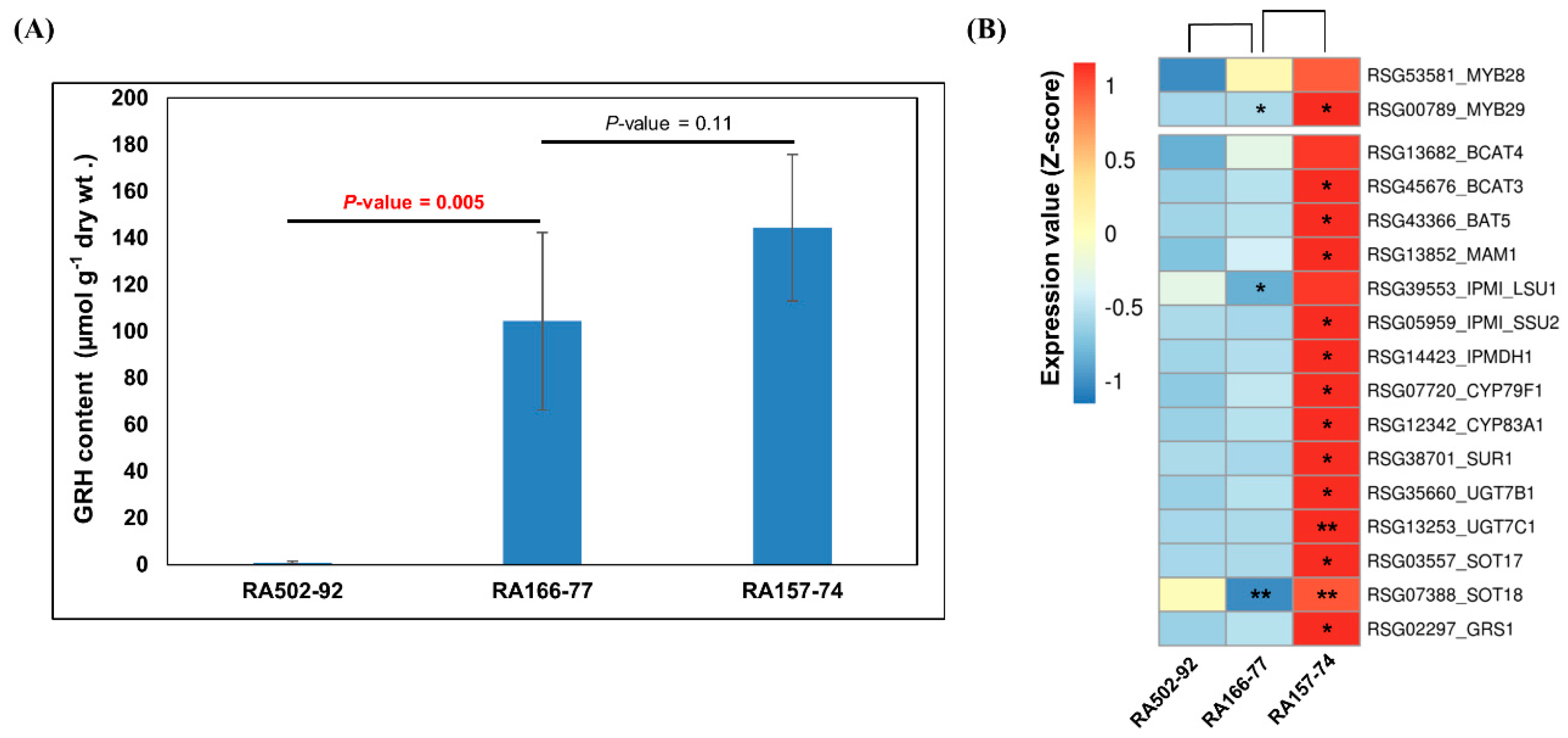
2.5. GRH Content and Expression of GRH Biosynthesis Genes in Radish Roots
3. Discussion
3.1. Abundant GRH Content and Functional Possibility for Radish Breeding
3.2. Induction of GRH Biosynthesis by MYB29 Transcription Factor in Radish Roots
3.3. Expression Profiling of GRH Biosynthesis Genes in Radish Roots
4. Materials and Methods
4.1. Plant Materials
4.2. Determination of GSL Content
4.3. Total RNA Isolation and RNA-Seq Analysis
4.4. Analysis of Differentially Expressed Genes Involved in GSL Biosynthesis
4.5. Quantitative Real-Time PCR
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| GSL | Glucosinolate |
| GSLs | Glucosinolates |
| GRH | Glucoraphasatin |
| GRA | Glucoraphanin |
| GRE | Glucoraphenin |
| GNA | Gluconapin |
| GBN | Glucobrassicanapin |
| GER | Glucoerucin |
| 4HGBS | 4-hydroxyglucobrassicin |
| 4MOGBS | 4-methoxyglucobrassicin |
| NGBS | Neoglucobrassicin |
| GBS | Glucobrassicin |
| LGRHA | Low GRH accession |
| HGRHA | High GRH accession |
| GO | Gene ontology |
| DEGs | Differentially expressed genes |
| 7MTH | 7-Methylthioheptyl |
| 8MTO | 8-Methylthiooctyl |
| 4MTB | 4-Methlthiobutyl |
| 3MTP | 3-Methylthiopropyl |
| BCATs | Branched-chain aminotransferases |
| MAMs | Methylthioalkylmalate synthases |
| IPMIs | Isopropylmalate isomerases |
| IPMDH | Isopropylmalate dehydrogenase |
| BAT5 | Bile acid transporter 5 |
| CYP | Cytochrome P450 monooxygenase |
| SUR1 | S-alkyl-thiohydroximate lyase |
| UGT74 | UDP-glucosyl transferase 74 |
| SOTs | Sulfotransferases |
| FMOGS-OXs | Flavin-monooxygenase glucosinolate S-oxygenases |
| MYB | MYB transcription factor |
| GRS1 | Glucoraphasatin synthase 1 |
| GS-OH | 2-oxoglutarate and Fe (II)-dependent oxygenase |
| IGMTs | O-methyltransferases |
| AOP2 | 2-oxoglutarate-dependent dioxygenase |
| qRT-PCR | Quantitative real-time Polymerase Chain Reaction |
| RsActin | Raphanus sativus actin |
| MeJA | Methyl jasmonate |
| DS-GSL | Desulfo-glucosinolate |
| DH | Doubled haploid |
| FDR | False discovery rate |
| ITCs | Isothiocyanates |
| Wt. | Weights |
References
- Jeon, J.; Bong, S.J.; Park, J.S.; Park, Y.K.; Arasu, M.V.; Al-Dhabi, N.A.; Park, S.U. De novo transcriptome analysis and glucosinolate profiling in watercress (Nasturtium officinale R. Br.). BMC Genom. 2017, 18, 401. [Google Scholar] [CrossRef]
- Wittstock, U.; Halkier, B.A. Glucosinolate research in the Arabidopsis era. Trends Plant Sci. 2002, 7, 254–270. [Google Scholar] [CrossRef]
- Halkier, B.A.; Gershenzon, J. Biology and Biochemistry of Glucosinolates. Annu. Rev. Plant Biol. 2006, 57. [Google Scholar] [CrossRef] [Green Version]
- Wiesner, M.; Zrenner, R.; Krumbein, A.; Glatt, H.; Schreiner, M. Genotypic variation of the glucosinolate profile in pak choi (Brassica rapa ssp. chinensis). J. Agric. Food Chem. 2013, 61, 1943–1953. [Google Scholar] [CrossRef]
- Lee, J.; Noh, Y.-H.; Park, K.-H.; Kim, D.-S.; Jeong, H.T.; Lee, H.-S.; Min, S.R.; Kim, H. Environmentally friendly fertilizers can enhance yield and bioactive compounds in Chinese cabbage (Brassica rapa ssp. pekinensis). Turk. J. Agric. For. 2019, 43, 138–150. [Google Scholar] [CrossRef]
- Guo, R.; Huang, Z.; Deng, Y.; Chen, X.; XuHan, X.; Lai, Z. Comparative Transcriptome Analyses Reveal a Special Glucosinolate Metabolism Mechanism in Brassica alboglabra Sprouts. Front. Plant Sci. 2016, 7, 1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robin, A.H.; Yi, G.E.; Laila, R.; Yang, K.; Park, J.I.; Kim, H.R.; Nou, I.S. Expression Profiling of Glucosinolate Biosynthetic Genes in Brassica oleracea L. var. capitata Inbred Lines Reveals Their Association with Glucosinolate Content. Molecules 2016, 21, 787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padilla, G.; Cartea, M.E.; Velasco, P.; de Haro, A.; Ordas, A. Variation of glucosinolates in vegetable crops of Brassica rapa. Phytochemistry 2007, 68, 536–545. [Google Scholar] [CrossRef]
- Liu, S.; Liu, Y.; Yang, X.; Tong, C.; Edwards, D.; Parkin, I.A.; Zhao, M.; Ma, J.; Yu, J.; Huang, S.; et al. The Brassica oleracea genome reveals the asymmetrical evolution of polyploid genomes. Nat Commun. 2014, 5, 3930. [Google Scholar] [CrossRef]
- Lee, Y.S.; Ku, K.M.; Becker, T.M.; Juvik, J.A. Chemopreventive glucosinolate accumulation in various broccoli and collard tissues: Microfluidic-based targeted transcriptomics for by-product valorization. PLoS ONE 2017, 12, e0185112. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Pan, Y.; Liu, Z.; Zhu, X.; Zhai, L.; Xu, L.; Yu, R.; Gong, Y.; Liu, L. De novo transcriptome sequencing of radish (Raphanus sativus L.) and analysis of major genes involved in glucosinolate metabolism. BMC Genom. 2013, 14, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitsui, Y.; Shimomura, M.; Komatsu, K.; Namiki, N.; Shibata-Hatta, M.; Imai, M.; Katayose, Y.; Mukai, Y.; Kanamori, H.; Kurita, K.; et al. The radish genome and comprehensive gene expression profile of tuberous root formation and development. Sci. Rep. 2015, 5, 10835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, M.-S.; Chung, J.-H.; Park, B.-S.; Kim, J.S. Analysis of Sugars Content by Genotypes in 82 Radish (Raphanus sativus L.). Korean J. Plant Res. 2018, 31, 53–465. [Google Scholar] [CrossRef]
- Hashida, T.; Nakatsuji, R.; Budahn, H.; Schrader, O.; Peterka, H.; Fujimura, T.; Kubo, N.; Hirai, M. Construction of a chromosome-assigned, sequence-tagged linkage map for the radish, Raphanus sativus L. and QTL analysis of morphological traits. Breed Sci. 2013, 63, 218–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Wang, J.; Qiu, Y.; Liu, T.; Song, J.; Li, X. Identification of ’Xinlimei’ radish candidate genes associated with anthocyanin biosynthesis based on a transcriptome analysis. Gene 2018, 657, 81–91. [Google Scholar] [CrossRef]
- Hanlon, P.R.; Barnes, D.M. Phytochemical composition and biological activity of 8 varieties of radish (Raphanus sativus L.) sprouts and mature taproots. J. Food Sci. 2011, 76, C185–C192. [Google Scholar] [CrossRef]
- Ko, H.-C.; Sung, J.-S.; Hur, O.-S.; Baek, H.-J.; Jeon, Y.-A.; Luitel, B.P.; Ryu, K.-Y.; Kim, J.-B.; Rhee, J.-H. Comparison of Glucosinolate Contents in Leaves and Roots of Radish (Raphanus spp.). Korean J. Plant Res. 2017, 30, 579–589. [Google Scholar] [CrossRef]
- Yi, G.; Lim, S.; Chae, W.B.; Park, J.E.; Park, H.R.; Lee, E.J.; Huh, J.H. Root Glucosinolate Profiles for Screening of Radish (Raphanus sativus L.) Genetic Resources. J. Agric. Food Chem 2016, 64, 61–70. [Google Scholar] [CrossRef]
- Malik, M.S.; Riley, M.B.; Norsworthy, J.K.; Bridges, W., Jr. Glucosinolate profile variation of growth stages of wild radish (Raphanus raphanistrum). J. Agric. Food Chem. 2010, 58, 3309–3315. [Google Scholar] [CrossRef]
- Ishida, M.; Nagata, M.; Ohara, T.; Kakizaki, T.; Hatakeyama, K.; Nishio, T. Small variation of glucosinolate composition in Japanese cultivars of radish (Raphanus sativus L.) requires simple quantitative analysis for breeding of glucosinolate component. Breed Sci. 2012, 62, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Lee, H.W.; Li, Z.; Lu, Y.; Zou, L.; Ong, C.N. Simultaneous Quantification of 22 Glucosinolates in 12 Brassicaceae Vegetables by Hydrophilic Interaction Chromatography-Tandem Mass Spectrometry. ACS Omega 2018, 3, 15546–15553. [Google Scholar] [CrossRef] [PubMed]
- Papi, A.; Orlandi, M.; Bartolini, G.; Barillari, J.; Iori, R.; Paolini, M.; Ferroni, F.; Grazia Fumo, M.; Pedulli, G.F.; Valgimigli, L. Cytotoxic and antioxidant activity of 4-methylthio-3-butenyl isothiocyanate from Raphanus sativus L. (Kaiware Daikon) sprouts. J. Agric. Food Chem. 2008, 56, 875–883. [Google Scholar] [CrossRef] [PubMed]
- Sarwar, M.; Kirkegaard, J.A. Biofumigation potential of brassicas: II. Eff. Environ. Ontog. Glucosinolate Prod. Implic. Screen. Plant Soil 1998, 201, 91–101. [Google Scholar]
- Seo, M.S.; Jin, M.; Sohn, S.H.; Kim, J.S. Expression profiles of BrMYB transcription factors related to glucosinolate biosynthesis and stress response in eight subspecies of Brassica rapa. FEBS Open Biol. 2017, 7, 1646–1659. [Google Scholar] [CrossRef]
- Sonderby, I.E.; Burow, M.; Rowe, H.C.; Kliebenstein, D.J.; Halkier, B.A. A complex interplay of three R2R3 MYB transcription factors determines the profile of aliphatic glucosinolates in Arabidopsis. Plant Physiol. 2010, 153, 348–363. [Google Scholar] [CrossRef] [Green Version]
- Knill, T.; Schuster, J.; Reichelt, M.; Gershenzon, J.; Binder, S. Arabidopsis branched-chain aminotransferase 3 functions in both amino acid and glucosinolate biosynthesis. Plant Physiol. 2008, 146, 1028–1039. [Google Scholar] [CrossRef] [Green Version]
- Hasan, M.; Friedt, W.; Pons-Kuhnemann, J.; Freitag, N.M.; Link, K.; Snowdon, R.J. Association of gene-linked SSR markers to seed glucosinolate content in oilseed rape (Brassica napus ssp. napus). Theor. Appl. Genet. 2008, 116, 1035–1049. [Google Scholar] [CrossRef]
- Sonderby, I.E.; Geu-Flores, F.; Halkier, B.A. Biosynthesis of glucosinolates—gene discovery and beyond. Trends Plant Sci. 2010, 15, 283–290. [Google Scholar] [CrossRef]
- Nugroho, A.B.D.; Han, N.; Pervitasari, A.N.; Kim, D.H.; Kim, J. Differential expression of major genes involved in the biosynthesis of aliphatic glucosinolates in intergeneric Baemoochae (Brassicaceae) and its parents during development. Plant Mol. Biol. 2020, 102, 171–184. [Google Scholar] [CrossRef]
- Kakizaki, T.; Kitashiba, H.; Zou, Z.; Li, F.; Fukino, N.; Ohara, T.; Nishio, T.; Ishida, M. A 2-Oxoglutarate-Dependent Dioxygenase Mediates the Biosynthesis of Glucoraphasatin in Radish. Plant Physiol. 2017, 173, 1583–1593. [Google Scholar] [CrossRef] [Green Version]
- Baenas, N.; Garcia-Viguera, C.; Moreno, D.A. Biotic elicitors effectively increase the glucosinolates content in Brassicaceae sprouts. J. Agric. Food Chem. 2014, 62, 1881–1889. [Google Scholar] [CrossRef] [PubMed]
- De Nicola, G.R.; Bagatta, M.; Pagnotta, E.; Angelino, D.; Gennari, L.; Ninfali, P.; Rollin, P.; Iori, R. Comparison of bioactive phytochemical content and release of isothiocyanates in selected brassica sprouts. Food Chem. 2013, 141, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Dinkova-Kostova, A.T.; Kostov, R.V. Glucosinolates and isothiocyanates in health and disease. Trends Mol. Med. 2012, 18, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Soundararajan, P.; Kim, J.S. Anti-Carcinogenic Glucosinolates in Cruciferous Vegetables and Their Antagonistic Effects on Prevention of Cancers. Molecules 2018, 23, 2983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishida, M.; Hara, M.; Fukino, N.; Kakizaki, T.; Morimitsu, Y. Glucosinolate metabolism, functionality and breeding for the improvement of Brassicaceae vegetables. Breed Sci. 2014, 64, 48–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aarabi, F.; Kusajima, M.; Tohge, T.; Konishi, T.; Gigolashvili, T.; Takamune, M.; Sasazaki, Y.; Watanabe, M.; Nakashita, H.; Fernie, A.R.; et al. Sulfur deficiency-induced repressor proteins optimize glucosinolate biosynthesis in plants. Sci. Adv. 2016, 2, e1601087. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.J.; Chiu, Y.C.; Kim, N.K.; Park, H.M.; Lee, C.H.; Juvik, J.A.; Ku, K.M. Cultivar-Specific Changes in Primary and Secondary Metabolites in Pak Choi (Brassica Rapa, Chinensis Group) by Methyl Jasmonate. Int. J. Mol. Sci. 2017, 18, 1004. [Google Scholar] [CrossRef] [Green Version]
- Pangesti, N.; Reichelt, M.; van de Mortel, J.E.; Kapsomenou, E.; Gershenzon, J.; van Loon, J.J.; Dicke, M.; Pineda, A. Jasmonic Acid and Ethylene Signaling Pathways Regulate Glucosinolate Levels in Plants during Rhizobacteria-Induced Systemic Resistance Against a Leaf-Chewing Herbivore. J. Chem. Ecol. 2016, 42, 1212–1225. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Yang, H.; Ren, L.; Chen, W.; Liu, L.; Liu, F.; Zeng, L.; Yan, R.; Chen, K.; Fang, X. Jasmonic Acid-Mediated Aliphatic Glucosinolate Metabolism Is Involved in Clubroot Disease Development in Brassica napus L. Front. Plant Sci. 2018, 9, 750. [Google Scholar] [CrossRef]
- Hirai, M.Y.; Sugiyama, K.; Sawada, Y.; Tohge, T.; Obayashi, T.; Suzuki, A.; Araki, R.; Sakurai, N.; Suzuki, H.; Aoki, K.; et al. Omics-based identification of Arabidopsis Myb transcription factors regulating aliphatic glucosinolate biosynthesis. Proc. Natl. Acad. Sci. USA 2007, 104, 6478–6483. [Google Scholar] [CrossRef] [Green Version]
- Gigolashvili, T.; Yatusevich, R.; Rollwitz, I.; Humphry, M.; Gershenzon, J.; Flugge, U.I. The plastidic bile acid transporter 5 is required for the biosynthesis of methionine-derived glucosinolates in Arabidopsis thaliana. Plant Cell 2009, 21, 1813–1829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czerniawski, P.; Bednarek, P. Glutathione S-Transferases in the Biosynthesis of Sulfur-Containing Secondary Metabolites in Brassicaceae Plants. Front. Plant Sci. 2018, 9, 1639. [Google Scholar] [CrossRef] [PubMed]
- Klein, M.; Papenbrock, J. Kinetics and substrate specificities of desulfo-glucosinolate sulfotransferases in Arabidopsis thaliana. Physiol. Plant. 2009, 135, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq--a Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, 106. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classification | Common Name | Abbreviation | Chemical Name | Molecular Formula |
|---|---|---|---|---|
| Aliphatic | Glucoraphanin | GRA | 4-(Methylsulfinyl)butyl | C12H23NO10S3 |
| Glucoraphenin | GRE | 4-Methylsulfinyl-3-butenyl | C12H21NO10S3 | |
| Gluconapin | GNA | 3-Butenyl | C11H19NO9S2 | |
| Glucobrassicanapin | GBN | 4-Pentenyl | C12H21NO9S2 | |
| Glucoerucin | GER | 4-(Methylthio)butyl | C12H23NO9S3 | |
| Glucoraphasatin | GRH | 4-Methylthio-3-butenyl | C12H21NO9S3 | |
| Indolic | 4-Hydroxyglucobrassicin | 4HGBS | 4-Hydroxyindol-3-ylmethyl | C16H20N2O10S2 |
| 4-Methoxyglucobrassicin | 4MOGBS | 4-Methoxyindol-3-ylmethyl | C17H22N2O10S2 | |
| Neoglucobrassicin | NGBS | 1-Methoxyindol-3-ylmethyl | C17H22N2O10S2 | |
| Glucobrassicin | GBS | 3-Indolylmethyl | C16H20N2O9S2 |
| Accessions | Origin | Species | Selfing Generation | GRA | GRE | GNA | GBN | GER | GRH | NGBS | GBS | 4HGBS | 4MOGBS | Unknown | Unknown | Total GSLs |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RA502-92 | Hungary | Raphanus sativus convar. sativus | S2 | ND | ND | 0.27 ± 0.05 | 0.04 ± 0.04 | ND | 0.17 ± 0.30 | 0.17 ± 0.04 | 0.01 ± 0.01 | 0.05 ± 0.06 | 0.08 ± 0.07 | 0.09 ± 0.15 | ND | 0.35 ± 0.05 |
| IT119282-15 | Lebanon | Raphanus sativus var. sativus | S3 | 0.08 ± 0.13 | 0.12 ± 0.12 | ND | 0.66 ± 0.39 | ND | 32.70 ± 23.09 | 0.19 ± 0.05 | 0.23 ± 0.21 | 0.16 ± 0.14 | 0.01 ± 0.02 | 0.27 ± 0.22 | ND | 62.35 ± 9.14 |
| IT119238-8 | Iran | Raphanus sativus var. sativus | S2 | 0.17 ± 0.29 | 0.39 ± 0.31 | ND | 0.46 ± 0.18 | ND | 47.92 ± 11.01 | 0.18 ± 0.02 | 0.20 ± 0.09 | 0.36 ± 0.23 | 0.07 ± 0.01 | 0.35 ± 0.10 | ND | 38.55 ± 5.43 |
| RA280-82 | Kazakhstan | Raphanus sativus convar. sativus | S3 | 0.74 ± 0.24 | 0.32 ± 0.14 | 0.23 ± 0.05 | 0.56 ± 0.26 | ND | 39.69 ± 16.99 | 0.17 ± 0.04 | 0.38 ± 0.19 | 2.06 ± 0.58 | 0.02 ± 0.04 | 0.68 ± 0.34 | ND | 60.12 ± 6.30 |
| RA157-74 | Portugal | Raphanus sativus convar. sativus | S3 | 0.85 ± 0.47 | 0.49 ± 0.37 | 0.22 ± 0.02 | 0.71 ± 0.22 | ND | 138.39 ± 31.11 | 0.19 ± 0.08 | 0.25 ± 0.11 | 1.68 ± 0.50 | 0.19 ± 0.04 | 1.51 ± 0.35 | 0.08 ± 0.13 | 122.46 ± 14.39 |
| Gene ID | BLAST | |
|---|---|---|
| RSG02297 | nr BLAST | PREDICTED: 1-aminocyclopropane-1-carboxylate oxidase homolog 5-like, BAW81934.1| GLUCORAPHASATIN SYNTHASE 1 (Raphanus sativus) |
| InterProScan | IPR026992 (Pfam); Non-haem dioxygenase N-terminal domain, | |
| IPR005123 (Pfam); Oxoglutarate/iron-dependent dioxygenase | ||
| Araport11 | AT1G03410.2 | 2-oxoglutarate (2OG) and Fe (II)-dependent oxygenase superfamily protein | Chr1:844782-846463 REVERSE LENGTH = 361 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, J.-N.; Won, S.Y.; Seo, M.-S.; Lee, J.; Lee, S.M.; Kwon, S.-J.; Kim, J.S. Induction of Glucoraphasatin Biosynthesis Genes by MYB29 in Radish (Raphanus sativus L.) Roots. Int. J. Mol. Sci. 2020, 21, 5721. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165721
Kang J-N, Won SY, Seo M-S, Lee J, Lee SM, Kwon S-J, Kim JS. Induction of Glucoraphasatin Biosynthesis Genes by MYB29 in Radish (Raphanus sativus L.) Roots. International Journal of Molecular Sciences. 2020; 21(16):5721. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165721
Chicago/Turabian StyleKang, Ji-Nam, So Youn Won, Mi-Suk Seo, Jeongyeo Lee, Si Myung Lee, Soo-Jin Kwon, and Jung Sun Kim. 2020. "Induction of Glucoraphasatin Biosynthesis Genes by MYB29 in Radish (Raphanus sativus L.) Roots" International Journal of Molecular Sciences 21, no. 16: 5721. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165721