Effect of Triclosan Exposure on Developmental Competence in Parthenogenetic Porcine Embryo during Preimplantation

 , , and
, , and
Abstract
:1. Introduction
2. Results
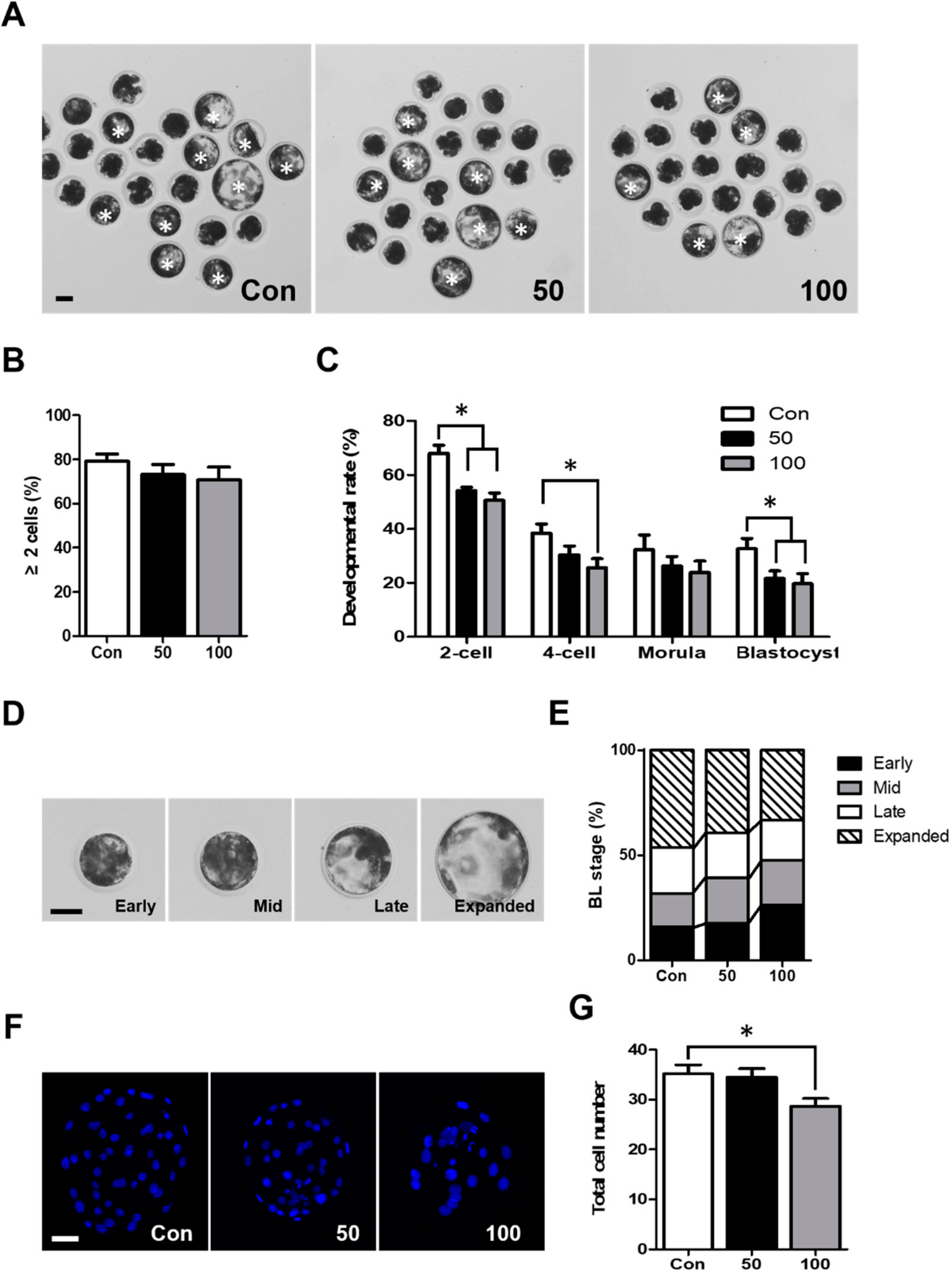
2.1. TCS Exposure Decreased Developmental Competence during Porcine Early Embryonic Development
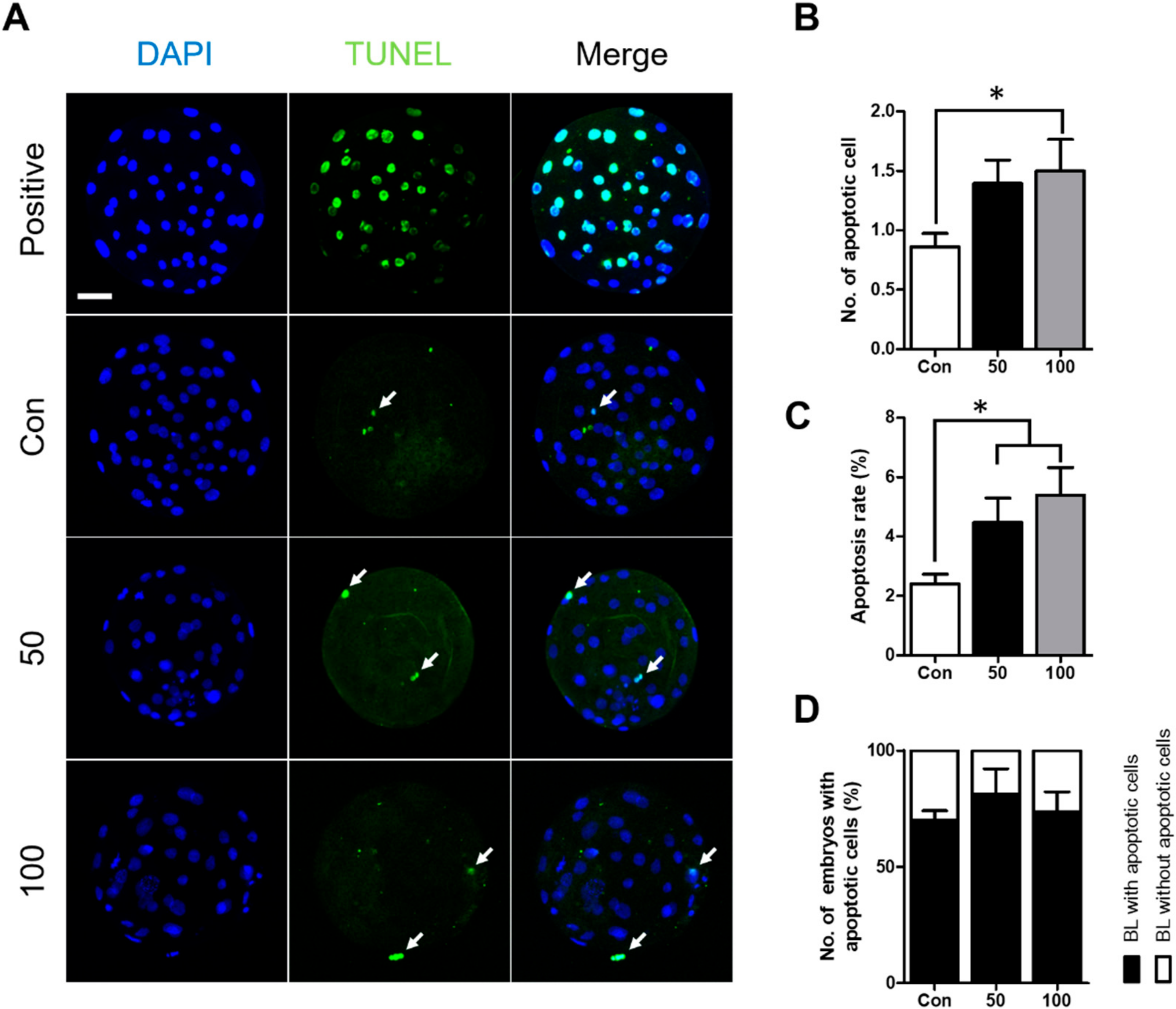
2.2. TCS Can Induce Apoptosis in Porcine Blastocysts
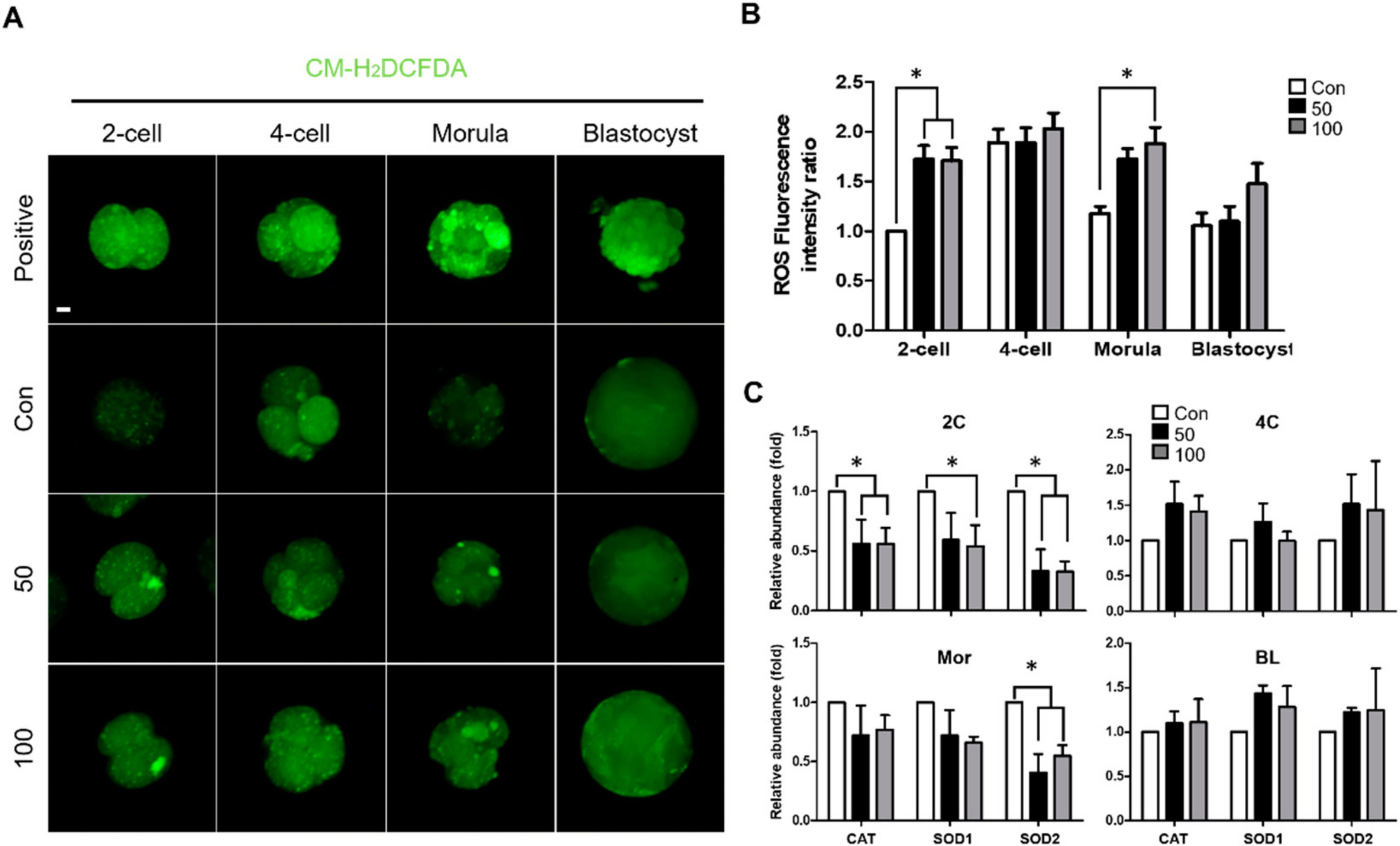
2.3. TCS Increased Oxidative Stress during Early Embryonic Development of Porcine Embryos
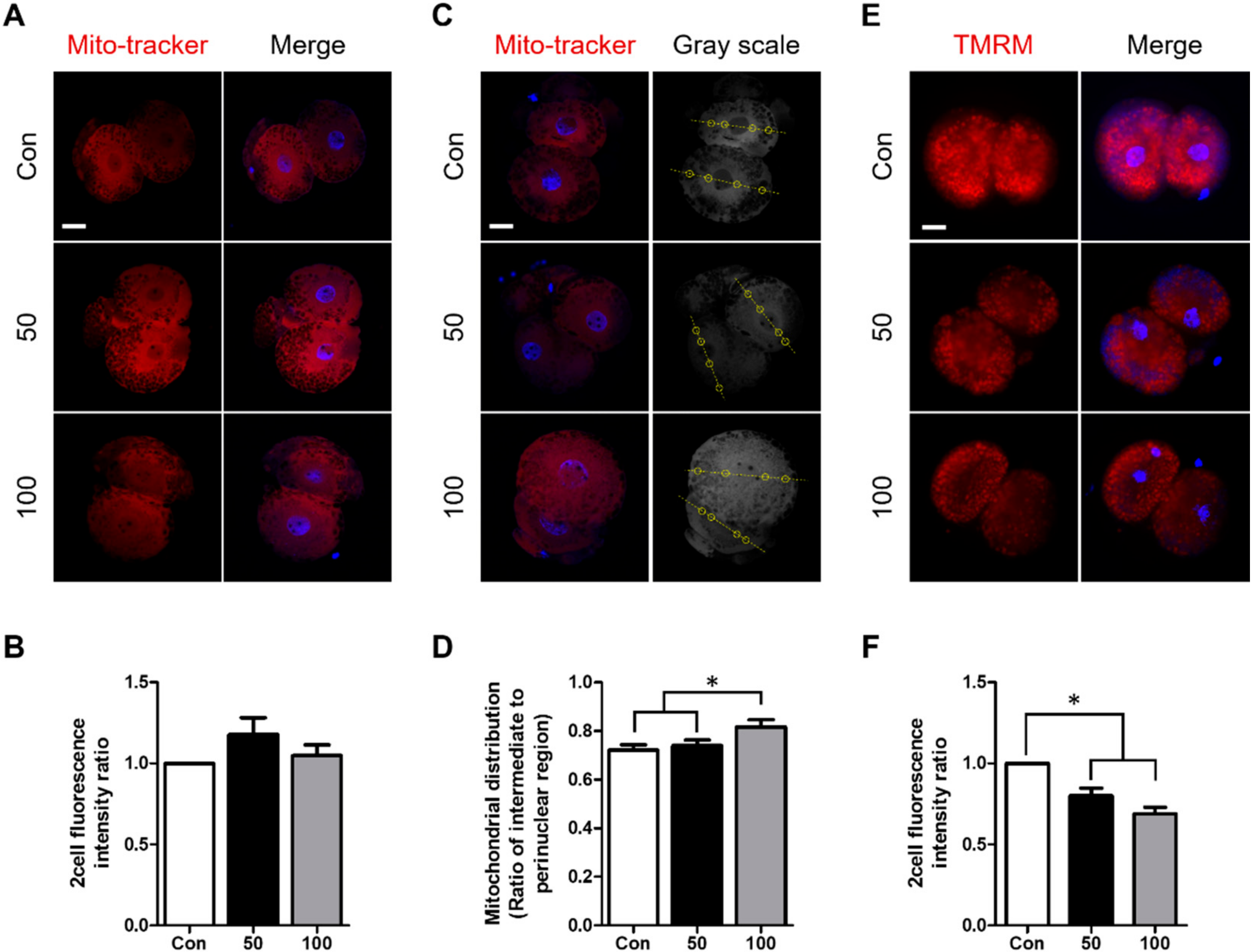
2.4. TCS Treatment Increased Mitochondrial Dysfunction in Porcine Embryos
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Oocyte Collection and In Vitro Maturation (IVM)
4.3. Chemical Treatment
4.4. Parthenogenetic Activation and In Vitro Culture (IVC)
4.5. Terminal Deoxynucleotidyl Transferase-Mediated dUTP-Digoxygenin Nick End-Labeling (TUNEL) Assay
4.6. Measurement of Intercellular ROS Levels
4.7. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.8. MitoTracker Staining
4.9. Quantification of Mitochondrial Distribution
4.10. Measurement of Mitochondrial Membrane Potential
4.11. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| TCS | Triclosan |
| EDC | Endocrine-disrupting chemical |
| BPA | Bisphenol A |
| ROS | Reactive oxygen species |
| MMP PA | Mitochondrial membrane potential Parthenogenetic activation |
| IVC | in vitro culture |
| EBs | Early blastocysts |
| MBs | Mid-blastocysts |
| LBs | Late blastocysts |
| ExBs | Expanded blastocysts |
| TUNEL | Terminal deoxynucleotidyl transferase-mediated dUTP-digoxygenin nick end-labeling |
| IVM | in vitro maturation |
| COCs | Cumulus oocyte complexes |
| BSA | Bovine serum albumin |
| PVA | Polyvinyl alcohol |
| DPBS | Dulbecco’s phosphate-buffered saline |
| qRT-PCR | Quantitative real-time polymerase chain reaction |
| TMRM | Tetramethylrhodamine methyl ester |
References
- Schweizer, H.P. Triclosan: A widely used biocide and its link to antibiotics. FEMS Microbiol. Lett. 2001, 202, 1–7. [Google Scholar] [CrossRef]
- Bhargava, H.N.; Leonard, P.A. Triclosan: Applications and safety. Am. J. Infect. Control 1996, 24, 209–218. [Google Scholar] [CrossRef]
- Weatherly, L.M.; Gosse, J.A. Triclosan exposure, transformation, and human health effects. J. Toxicol. Environ. Health B Crit. Rev. 2017, 20, 447–469. [Google Scholar] [CrossRef] [PubMed]
- Kabir, E.R.; Rahman, M.S.; Rahman, I. A review on endocrine disruptors and their possible impacts on human health. Environ. Toxicol. Pharmacol. 2015, 40, 241–258. [Google Scholar] [CrossRef] [PubMed]
- Valbuena, D.; Martin, J.; de Pablo, J.L.; Remohi, J.; Pellicer, A.; Simon, C. Increasing levels of estradiol are deleterious to embryonic implantation because they directly affect the embryo. Fertil. Steril. 2001, 76, 962–968. [Google Scholar] [CrossRef]
- Sifakis, S.; Androutsopoulos, V.P.; Tsatsakis, A.M.; Spandidos, D.A. Human exposure to endocrine disrupting chemicals: Effects on the male and female reproductive systems. Environ. Toxicol. Pharmacol. 2017, 51, 56–70. [Google Scholar] [CrossRef]
- Guo, J.; Zhao, M.H.; Shin, K.T.; Niu, Y.J.; Ahn, Y.D.; Kim, N.H.; Cui, X.S. The possible molecular mechanisms of bisphenol A action on porcine early embryonic development. Sci. Rep. 2017, 7, 8632. [Google Scholar] [CrossRef]
- Gassman, N.R. Induction of oxidative stress by bisphenol A and its pleiotropic effects. Environ. Mol. Mutagen 2017, 58, 60–71. [Google Scholar] [CrossRef] [Green Version]
- Li, S.J.; Chen, P.; Peres, T.V.; Villahoz, B.F.; Zhang, Z.; Miah, M.R.; Aschner, M. Triclosan induces PC12 cells injury is accompanied by inhibition of AKT/mTOR and activation of p38 pathway. Neurotoxicology 2019, 74, 221–229. [Google Scholar] [CrossRef]
- Ajao, C.; Andersson, M.A.; Teplova, V.V.; Nagy, S.; Gahmberg, C.G.; Andersson, L.C.; Hautaniemi, M.; Kakasi, B.; Roivainen, M.; Salkinoja-Salonen, M. Mitochondrial toxicity of triclosan on mammalian cells. Toxicol. Rep. 2015, 2, 624–637. [Google Scholar] [CrossRef] [Green Version]
- Yuan, M.; Bai, M.Z.; Huang, X.F.; Zhang, Y.; Liu, J.; Hu, M.H.; Zheng, W.Q.; Jin, F. Preimplantation Exposure to Bisphenol A and Triclosan May Lead to Implantation Failure in Humans. Biomed. Res. Int. 2015, 2015, 184–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dennery, P.A. Effects of oxidative stress on embryonic development. Birth Defects Res. C Embryo Today Rev. 2007, 81, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, L.; Xu, J.; Pan, M.H.; Sun, S.C. HT-2 toxin exposure induces mitochondria dysfunction and DNA damage during mouse early embryo development. Reprod. Toxicol. 2019, 85, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Shin, K.T.; Guo, J.; Niu, Y.J.; Cui, X.S. The toxic effect of aflatoxin B1 on early porcine embryonic development. Theriogenology 2018, 118, 157–163. [Google Scholar] [CrossRef]
- Komatsu, K.; Iwase, A.; Mawatari, M.; Wang, J.; Yamashita, M.; Kikkawa, F. Mitochondrial membrane potential in 2-cell stage embryos correlates with the success of preimplantation development. Reproduction 2014, 147, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Honkisz, E.; Zieba-Przybylska, D.; Wojtowicz, A.K. The effect of triclosan on hormone secretion and viability of human choriocarcinoma JEG-3 cells. Reprod. Toxicol. 2012, 34, 385–392. [Google Scholar] [CrossRef]
- Zuckerbraun, H.L.; Babich, H.; May, R.; Sinensky, M.C. Triclosan: Cytotoxicity, mode of action, and induction of apoptosis in human gingival cells in vitro. Eur. J. Oral Sci. 1998, 106, 628–636. [Google Scholar] [CrossRef]
- Santos, R.R.; Schoevers, E.J.; Roelen, B.A. Usefulness of bovine and porcine IVM/IVF models for reproductive toxicology. Reprod. Biol. Endocrinol. 2014, 12, 117. [Google Scholar] [CrossRef] [Green Version]
- Zorova, L.D.; Popkov, V.A.; Plotnikov, E.Y.; Silachev, D.N.; Pevzner, I.B.; Jankauskas, S.S.; Babenko, V.A.; Zorov, S.D.; Balakireva, A.V.; Juhaszova, M.; et al. Mitochondrial membrane potential. Anal. Biochem. 2018, 552, 50–59. [Google Scholar] [CrossRef]
- Petersen, R.C. Triclosan antimicrobial polymers. AIMS Mol. Sci. 2016, 3, 88–103. [Google Scholar] [CrossRef]
- Calafat, A.M.; Ye, X.; Wong, L.Y.; Reidy, J.A.; Needham, L.L. Urinary concentrations of triclosan in the U.S. population: 2003–2004. Environ. Health Perspect. 2008, 116, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Dayan, A.D. Risk assessment of triclosan [Irgasan] in human breast milk. Food Chem. Toxicol. 2007, 45, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Yi, B.R.; Go, R.E.; Hwang, K.A.; Nam, K.H.; Choi, K.C. Methoxychlor and triclosan stimulates ovarian cancer growth by regulating cell cycle- and apoptosis-related genes via an estrogen receptor-dependent pathway. Environ. Toxicol. Pharmacol. 2014, 37, 1264–1274. [Google Scholar] [CrossRef]
- Isom, S.C.; Li, R.F.; Whitworth, K.M.; Prather, R.S. Timing of first embryonic cleavage is a positive indicator of the in vitro developmental potential of porcine embryos derived from in vitro fertilization, somatic cell nuclear transfer and parthenogenesis. Mol. Reprod. Dev. 2012, 79, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Lundin, K.; Bergh, C.; Hardarson, T. Early embryo cleavage is a strong indicator of embryo quality in human IVF. Hum. Reprod. 2001, 16, 2652–2657. [Google Scholar] [CrossRef]
- Vandaele, L.; Mateusen, B.; Maes, D.; de Kruif, A.; Van Soom, A. Is apoptosis in bovine in vitro produced embryos related to early developmental kinetics and in vivo bull fertility? Theriogenology 2006, 65, 1691–1703. [Google Scholar] [CrossRef]
- Booth, P.J.; Watson, T.J.; Leese, H.J. Prediction of porcine blastocyst formation using morphological, kinetic, and amino acid depletion and appearance criteria determined during the early cleavage of in vitro-produced embryos. Biol. Reprod. 2007, 77, 765–779. [Google Scholar] [CrossRef] [Green Version]
- Song, B.S.; Jeong, P.S.; Lee, J.H.; Lee, M.H.; Yang, H.J.; Choi, S.A.; Lee, H.Y.; Yoon, S.B.; Park, Y.H.; Jeong, K.J.; et al. The effects of kinase modulation on in vitro maturation according to different cumulus-oocyte complex morphologies. PLoS ONE 2018, 13, e0205495. [Google Scholar] [CrossRef]
- Rath, D.; Niemann, H.; Torres, C.R. In vitro development to blastocysts of early porcine embryos produced in vivo or in vitro. Theriogenology 1995, 43, 913–926. [Google Scholar] [CrossRef]
- Wirleitner, B.; Schuff, M.; Stecher, A.; Murtinger, M.; Vanderzwalmen, P. Pregnancy and birth outcomes following fresh or vitrified embryo transfer according to blastocyst morphology and expansion stage, and culturing strategy for delayed development. Hum. Reprod. 2016, 31, 1685–1695. [Google Scholar] [CrossRef]
- Ribeiro, E.S.; Gerger, R.P.; Ohlweiler, L.U.; Ortigari, I., Jr.; Mezzalira, J.C.; Forell, F.; Bertolini, L.R.; Rodrigues, J.L.; Ambrosio, C.E.; Miglino, M.A.; et al. Developmental potential of bovine hand-made clone embryos reconstructed by aggregation or fusion with distinct cytoplasmic volumes. Cloning Stem Cells 2009, 11, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Lai, L.; Mao, J.; Im, G.S.; Bonk, A.; Prather, R.S. Apoptosis and in vitro development of preimplantation porcine embryos derived in vitro or by nuclear transfer. Biol. Reprod. 2003, 69, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Yueh, M.F.; Taniguchi, K.; Chen, S.; Evans, R.M.; Hammock, B.D.; Karin, M.; Tukey, R.H. The commonly used antimicrobial additive triclosan is a liver tumor promoter. Proc. Natl. Acad. Sci. USA 2014, 111, 17200–17205. [Google Scholar] [CrossRef] [Green Version]
- Liang, S.; Jin, Y.X.; Yuan, B.; Zhang, J.B.; Kim, N.H. Melatonin enhances the developmental competence of porcine somatic cell nuclear transfer embryos by preventing DNA damage induced by oxidative stress. Sci. Rep. 2017, 7, 111–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Che, M.J.; Liu, C.; Liu, H.G.; Fu, X.W.; Hou, Y.P. Toxicity and related mechanisms of dihydroartemisinin on porcine oocyte maturation in vitro. Toxicol. Appl. Pharmacol. 2018, 341, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.J.; Zhou, W.; Guo, J.; Nie, Z.W.; Shin, K.T.; Kim, N.H.; Lv, W.F.; Cui, X.S. C-Phycocyanin protects against mitochondrial dysfunction and oxidative stress in parthenogenetic porcine embryos. Sci. Rep. 2017, 7, 169–192. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Jin, M.; Nguyen, S.H.; Mao, L.; Li, J.; Coin, L.J.M.; Yuan, Z.; Guo, J. Non-antibiotic antimicrobial triclosan induces multiple antibiotic resistance through genetic mutation. Environ. Int. 2018, 118, 257–265. [Google Scholar] [CrossRef]
- Wang, F.; Xu, R.; Zheng, F.; Liu, H. Effects of triclosan on acute toxicity, genetic toxicity and oxidative stress in goldfish (Carassius auratus). Exp. Anim. 2018, 67, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Dubey, D.; Srivastav, A.K.; Singh, J.; Chopra, D.; Qureshi, S.; Kushwaha, H.N.; Singh, N.; Ray, R.S. Photoexcited triclosan induced DNA damage and oxidative stress via p38 MAP kinase signaling involving type I radicals under sunlight/UVB exposure. Ecotoxicol. Environ. Saf. 2019, 174, 270–282. [Google Scholar] [CrossRef]
- Ishihara, Y.; Takemoto, T.; Ishida, A.; Yamazaki, T. Protective actions of 17beta-estradiol and progesterone on oxidative neuronal injury induced by organometallic compounds. Oxid. Med. Cell. Longev. 2015, 2015, 343–706. [Google Scholar] [CrossRef] [Green Version]
- Ahn, K.C.; Zhao, B.; Chen, J.; Cherednichenko, G.; Sanmarti, E.; Denison, M.S.; Lasley, B.; Pessah, I.N.; Kultz, D.; Chang, D.P.; et al. In vitro biologic activities of the antimicrobials triclocarban, its analogs, and triclosan in bioassay screens: Receptor-based bioassay screens. Environ. Health Perspect. 2008, 116, 1203–2310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, C.; Hsu, W.L.; Hong, W.F.; Cheng, W.T.; Yang, Y. Estrogen receptor is expressed in pig embryos during preimplantation development. Mol. Reprod. Dev. 2000, 55, 83–88. [Google Scholar] [CrossRef]
- Yang, S.G.; Park, H.J.; Lee, S.M.; Kim, J.W.; Kim, M.J.; Kim, I.S.; Jegal, H.-G.; Koo, D.-B. Reduction of Mitochondrial Derived Superoxide by Mito-TEMPO Improves Porcine Oocyte Maturation In Vitro. J. Anim. Reprod. Biotechnol. 2019, 34, 10–19. [Google Scholar]
- Li, G.; Li, J.; Hao, R.; Guo, Y. Activation of catalase activity by a peroxisome-localized small heat shock protein Hsp17.6CII. J. Genet. Genom. 2017, 44, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Patrushev, N.; Seidel-Rogol, B.; Salazar, G. Angiotensin II requires zinc and downregulation of the zinc transporters ZnT3 and ZnT10 to induce senescence of vascular smooth muscle cells. PLoS ONE 2012, 7, e33211. [Google Scholar] [CrossRef] [Green Version]
- Zou, L.; Mi, C.; Yu, H.; Gu, W.; Teng, Y. Characterization of the interaction between triclosan and catalase. RSC Adv. 2017, 7, 9031–9036. [Google Scholar] [CrossRef] [Green Version]
- Mi, C.; Teng, Y.; Wang, X.; Yu, H.; Huang, Z.; Zong, W.; Zou, L. Molecular interaction of triclosan with superoxide dismutase (SOD) reveals a potentially toxic mechanism of the antimicrobial agent. Ecotoxicol. Environ. Saf. 2018, 153, 78–83. [Google Scholar] [CrossRef]
- Van der Bliek, A.M.; Sedensky, M.M.; Morgan, P.G. Cell Biology of the Mitochondrion. Genetics 2017, 207, 843–871. [Google Scholar] [CrossRef] [Green Version]
- West, A.P. Mitochondrial dysfunction as a trigger of innate immune responses and inflammation. Toxicology 2017, 391, 54–63. [Google Scholar] [CrossRef]
- Nagai, S.; Mabuchi, T.; Hirata, S.; Shoda, T.; Kasai, T.; Yokota, S.; Shitara, H.; Yonekawa, H.; Hoshi, K. Correlation of abnormal mitochondrial distribution in mouse oocytes with reduced developmental competence. Tohoku J. Exp. Med. 2006, 210, 137–144. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Frolova, A.I.; Purcell, S.; Adastra, K.; Schoeller, E.; Chi, M.M.; Schedl, T.; Moley, K.H. Mitochondrial dysfunction and apoptosis in cumulus cells of type I diabetic mice. PLoS ONE 2010, 5, e15901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, H.; Satoh, M.; Kabashima, K. Distributions of Mitochondria and the Cytoskeleton in Hamster Embryos Developed In Vivo and In Vitro. J. Mamm. Ova Res. 2006, 23, 128–134. [Google Scholar] [CrossRef]
- Ludwig, T.E.; Squirrell, J.M.; Palmenberg, A.C.; Bavister, B.D. Relationship between development, metabolism, and mitochondrial organization in 2-cell hamster embryos in the presence of low levels of phosphate. Biol. Reprod. 2001, 65, 1648–1654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Gao, Y.Y.; Chen, L.; Nie, Z.W.; Cheng, W.; Liu, X.; Schatten, H.; Zhang, X.; Miao, Y.L. Melatonin prevents postovulatory oocyte aging and promotes subsequent embryonic development in the pig. Aging (Albany N. Y.) 2017, 9, 1552–1564. [Google Scholar] [CrossRef] [Green Version]
- Teplova, V.V.; Belosludtsev, K.N.; Kruglov, A.G. Mechanism of triclosan toxicity: Mitochondrial dysfunction including complex II inhibition, superoxide release and uncoupling of oxidative phosphorylation. Toxicol. Lett. 2017, 275, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Byun, H.O.; Kim, H.Y.; Lim, J.J.; Seo, Y.H.; Yoon, G. Mitochondrial dysfunction by complex II inhibition delays overall cell cycle progression via reactive oxygen species production. J. Cell. Biochem. 2008, 104, 1747–1759. [Google Scholar] [CrossRef] [PubMed]
- Yoon, Y.S.; Byun, H.O.; Cho, H.; Kim, B.K.; Yoon, G. Complex II defect via down-regulation of iron-sulfur subunit induces mitochondrial dysfunction and cell cycle delay in iron chelation-induced senescence-associated growth arrest. J. Biol. Chem. 2003, 278, 51577–51586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Squirrell, J.M.; Lane, M.; Bavister, B.D. Altering intracellular pH disrupts development and cellular organization in preimplantation hamster embryos. Biol. Reprod. 2001, 64, 1845–1854. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences | GenBank Accession No. | Product Size (bp) |
|---|---|---|---|
| Catalase | F: 5′-TGT ACC CGC TAT TCT GGG GA-3′ | NM_214301.2 | 119 |
| R: 5′-ACA TGG GCG ATA AGA CCC CT-3′ | |||
| SOD1 | F: 5′-GGT GGG CCA AAG GAT CAA GA-3′ | NM_001190422.1 | 220 |
| R: 5′-CCA CCC GGT TTC CTA GTT CT-3′ | |||
| SOD2 | F: 5′-GGT GGA GGC CAC ATC AAT CA-3′ | NM_214127.2 | 80 |
| R: 5′-CCA CCT CCG GTG TAG TTA GT-3′ | |||
| GAPDH | F: 5′-CCC TGA GAC ACG ATG GTG AA-3′ | NM_001206359.1 | 127 |
| R: 5′-GGG ACT CTG TGC TAC CAC TT-3′ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.J.; Park, H.-J.; Lee, S.; Kang, H.-G.; Jeong, P.-S.; Park, S.H.; Park, Y.-H.; Lee, J.-H.; Lim, K.S.; Lee, S.H.; et al. Effect of Triclosan Exposure on Developmental Competence in Parthenogenetic Porcine Embryo during Preimplantation. Int. J. Mol. Sci. 2020, 21, 5790. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165790
Kim MJ, Park H-J, Lee S, Kang H-G, Jeong P-S, Park SH, Park Y-H, Lee J-H, Lim KS, Lee SH, et al. Effect of Triclosan Exposure on Developmental Competence in Parthenogenetic Porcine Embryo during Preimplantation. International Journal of Molecular Sciences. 2020; 21(16):5790. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165790
Chicago/Turabian StyleKim, Min Ju, Hyo-Jin Park, Sanghoon Lee, Hyo-Gu Kang, Pil-Soo Jeong, Soo Hyun Park, Young-Ho Park, Jong-Hee Lee, Kyung Seob Lim, Seung Hwan Lee, and et al. 2020. "Effect of Triclosan Exposure on Developmental Competence in Parthenogenetic Porcine Embryo during Preimplantation" International Journal of Molecular Sciences 21, no. 16: 5790. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165790