Induction of Callogenesis, Organogenesis, and Embryogenesis in Non-Meristematic Explants of Bleeding Heart and Evaluation of Chemical Diversity of Key Metabolites from Callus



Abstract
:1. Introduction
2. Results
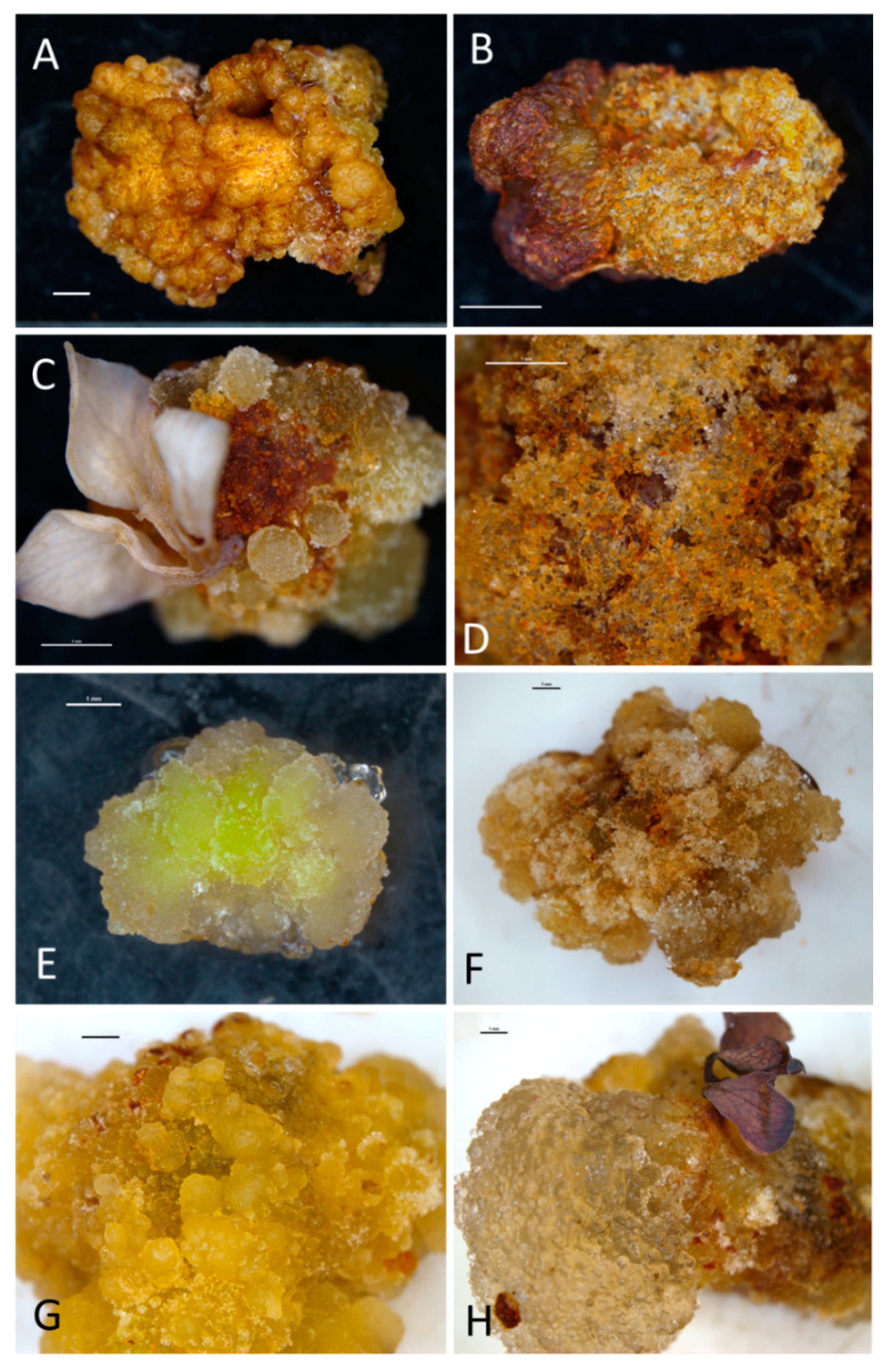
2.1. Morphogenetic Response of Explants—Callogenesis and Organogenesis
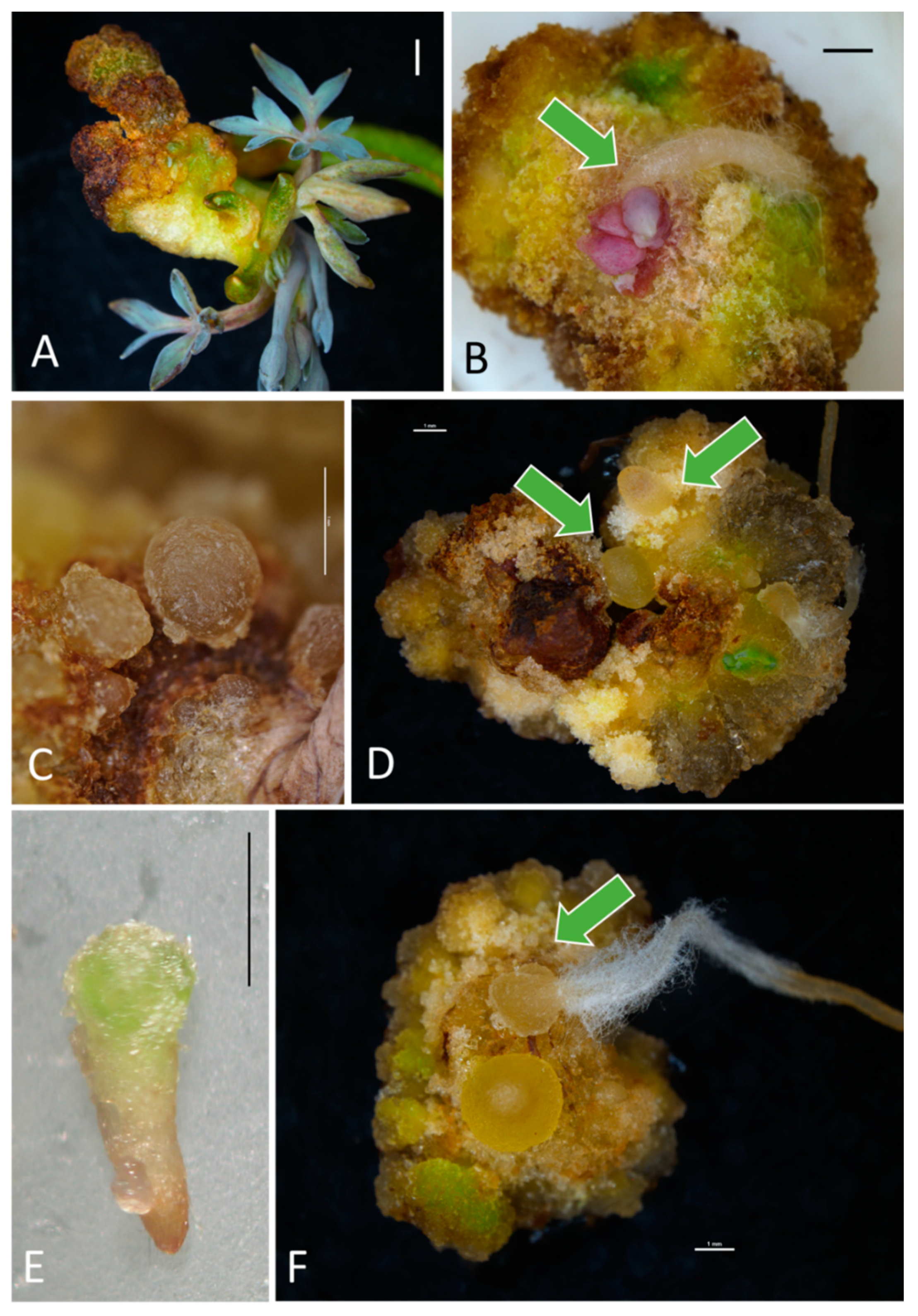
2.2. Morphogenetic Response of Explants—Somatic Embryogenesis
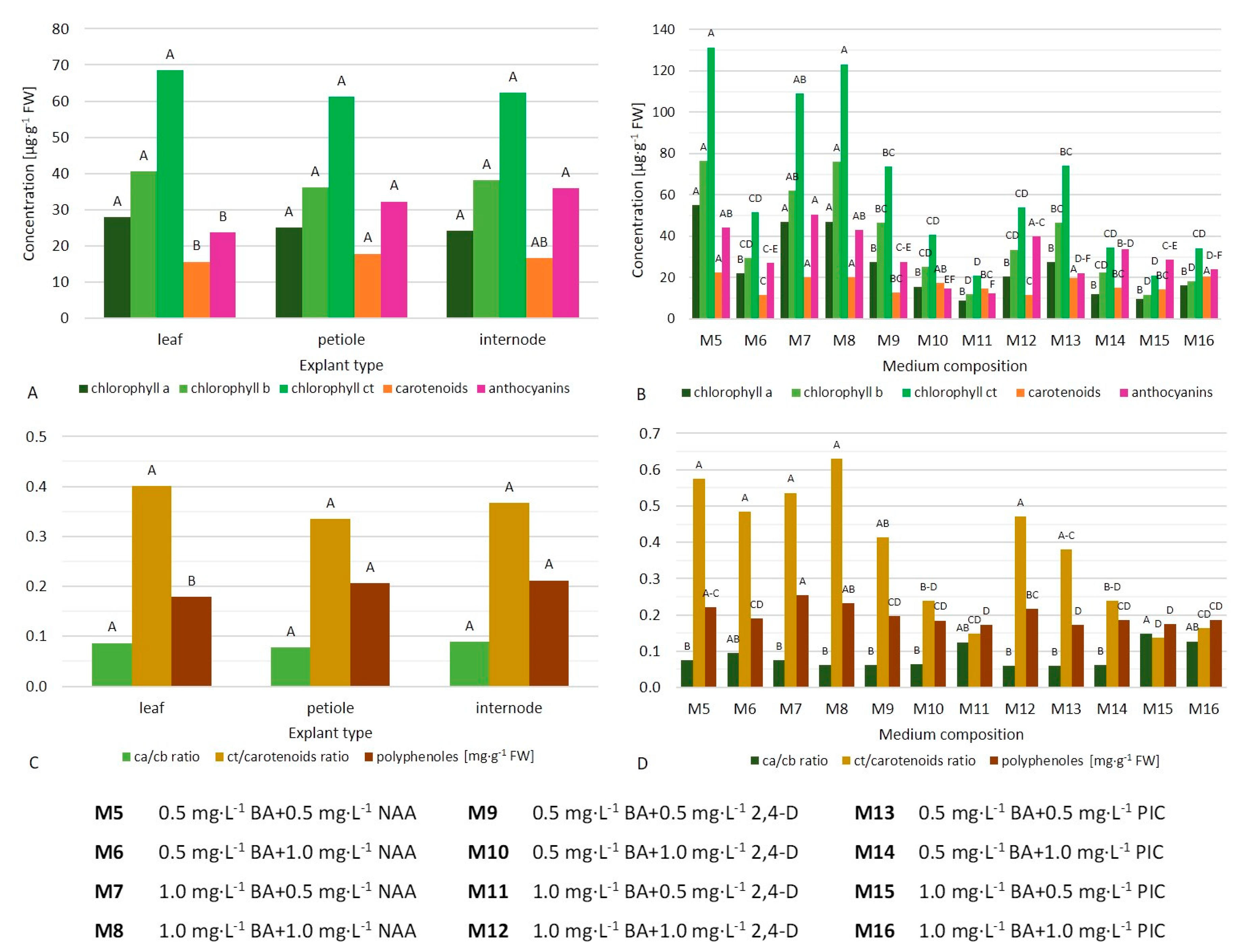
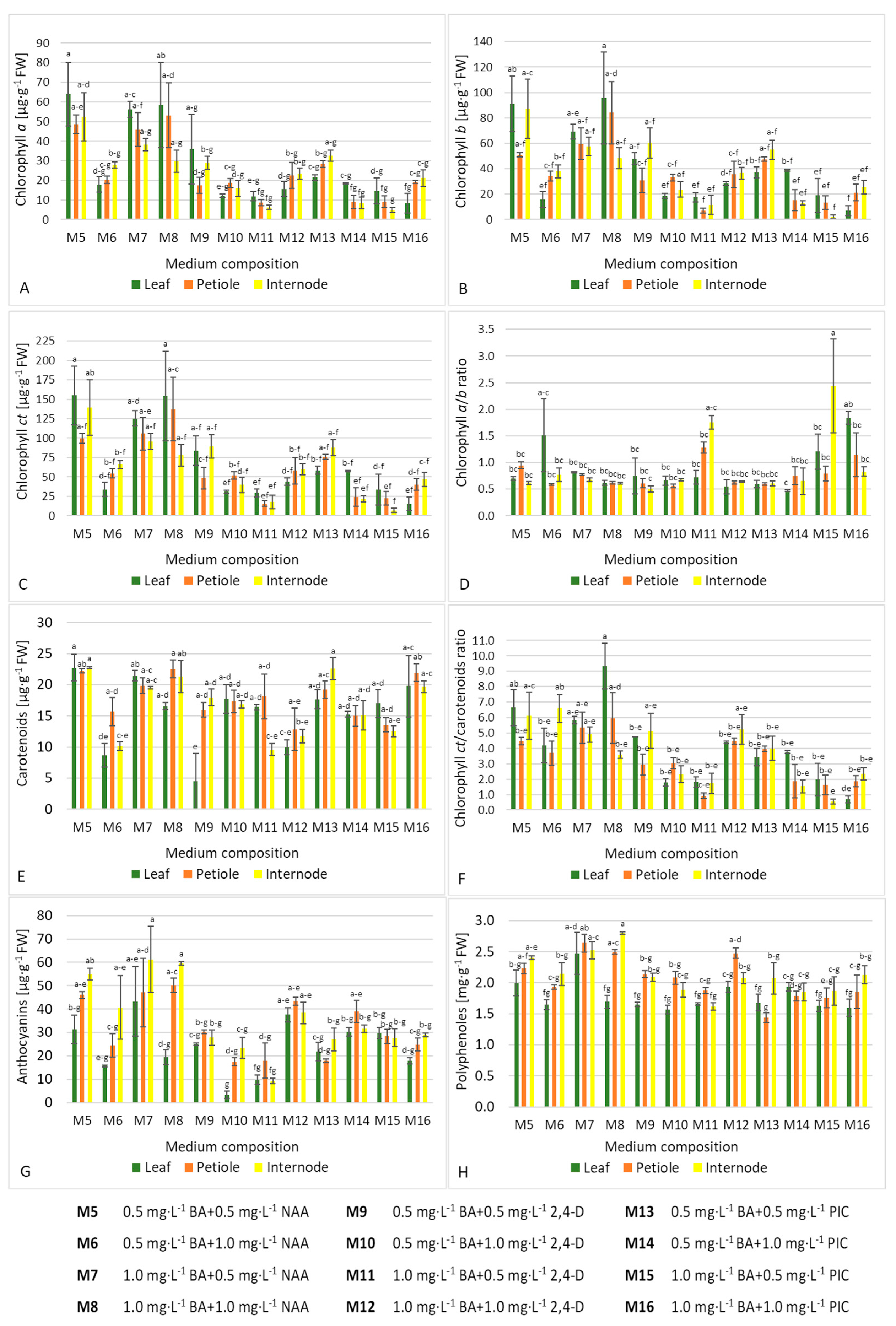
2.3. Spectral Analysis of Primary and Secondary Metabolites in Callus
3. Discussion
3.1. Callogenesis, Adventitious Organogenesis, and Embryogenesis in the Bleeding Heart
3.2. Biosynthesis of Metabolites in Callus
4. Materials and Methods
4.1. Plant Material, Media Preparation, and General Culture Conditions
4.2. Evaluation of a Morphogenetic Response in Explants
4.3. Spectral Assay
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 2,4-D | 2,4-dichlorphenoxyacetic acid |
| ANOVA | Analysis of variance |
| AXs | Auxins |
| BA | 6-benzyladenine |
| BAR | 6-benzyl-adenine riboside |
| ca | Chlorophyll a |
| cb | Chlorophyll b |
| CKs | Cytokinins |
| ct | Total chlorophyll |
| DW | Dry weight |
| FW | Fresh weight |
| IAA | Indole-3-acetic acid |
| IBA | Indole-3-butyric acid |
| KIN | Kinetin |
| M | Medium |
| MS | Murashige and Skoog (1962) medium |
| NAA | 1-naphthaleneacetic acid |
| PGR | Plant growth regulator |
| PIC | Picloram |
| SE | Somatic embryogenesis |
| SH | Schenk and Hildebrandt (1972) medium |
| TDZ | Thidiazuron |
| TOP | Topolin |
| ZEA | Zeatin |
References
- Hodges, L. Bleeding heart: A review for growers. Hort. Technol. 2012, 22, 517–522. [Google Scholar] [CrossRef] [Green Version]
- Iwasa, K.; Kim, C.-W. Biotransformations of protoberberines in cell cultures of Dicentra spectablis. Phytochemistry 1997, 46, 1359–1363. [Google Scholar] [CrossRef]
- Lee, D.-H.; Lee, B.-C.; Yoon, E.-J.; Leem, K.-E.; Park, S.-M.; Pyo, H.-B.; Choe, T.-B. Development of effects of plant extracts on the activity and expression of UVA-induced MMPs (matrix metalloproteases). Int. J. Cosmet. Sci. 2004, 26, 317–319. [Google Scholar] [CrossRef]
- McNulty, J.; Poloczek, J.; Larichev, V.; Werstiuk, N.H.; Griffin, C.; Pandey, S. Discovery of the apoptosis-inducing activity and high accumulation of the butenolides, menisdaurilide and aquilegiolide in Dicentra spectabilis. Planta Med. 2007, 73, 1543–1547. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.H.; Jang, J.H.; Woo, K.W.; Park, J.E.; Lee, K.H.; Jung, H.K.; An, B.; Jung, W.S.; Ham, S.H.; Cho, H.W. Chemical constituents of Dicentra spectabilis and their anti-inflammation effect. J. Appl. Biol. Chem. 2018, 61, 39–46. [Google Scholar] [CrossRef] [Green Version]
- Petruczynik, A.; Plech, T.; Tuzimski, T.; Misiurek, J.; Kaproń, B.; Misiurek, D.; Szultka-Młyńska, M.; Buszewski, B.; Waksmundzka-Hajnos, M. Determination of selected isoquinoline alkaloids from Mahonia aquifolia; Meconopsis cambrica; Corydalis lutea; Dicentra spectabilis; Fumaria officinalis; Macleaya cordata extracts by HPLC-DAD and comparison of their cytotoxic activity. Toxins 2019, 11, 575. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.-G.; Fukuski, Y.; Tahara, S.; Osawa, T. Fungitoxic alkaloids from Hokkaido Papaveraceae. Fitoterapia 2000, 71, 527–534. [Google Scholar] [CrossRef]
- Lee, K.P.; Lee, D.W. Somatic embryogenesis and plant regeneration from seeds of wild Dicentra spectabilis (L.) Lem. Plant Cell Rep. 2003, 22, 105–109. [Google Scholar] [CrossRef]
- Kulus, D. Influence of growth regulators on the development, quality, and physiological state of in vitro-propagated Lamprocapnos spectabilis (L.) Fukuhara. In Vitr. Cell. Dev. Biol. Plant 2020, 56, 447–457. [Google Scholar] [CrossRef]
- Miler, N.; Kulus, D. Microwave treatment can induce chrysanthemum phenotypic and genetic changes. Sci. Hortic. 2018, 227, 223–233. [Google Scholar] [CrossRef]
- Fehér, A. Callus, dedifferentiation, totipotency, somatic embryogenesis: What these terms mean in the era of molecular plant biology? Front. Plant Sci. 2019, 10, 536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Pérez, P.; Lozano-Milo, E.; Landín, M.; Gallego, P.P. Machine learning technology reveals the concealed interactions of phytohormones on medicinal plant in vitro organogenesis. Biomolecules 2020, 10, 746. [Google Scholar] [CrossRef]
- Thorpe, T.A. History of plant tissue culture. Mol. Biotechnol. 2007, 37, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Simon, S.; Petrášek, P. Why plants need more than one type of auxin. Plant Sci. 2011, 180, 454–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobránszki, J.; Mendler-Drienyovszk, N. Cytokinin-induced changes in the chlorophyll content and fluorescence of in vitro apple leaves. J. Plant Physiol. 2014, 171, 1472–1478. [Google Scholar] [CrossRef]
- Lema-Rumińska, J.; Kulus, D. Micropropagation of cacti—A review. Haseltonia 2014, 19, 46–63. [Google Scholar] [CrossRef]
- Murthy, H.N.; Lee, E.; Paek, K. Production of secondary metabolites from cell and organ cultures: Strategies and approaches for biomass improvement and metabolite accumulation. Plant Cell Tiss. Organ Cult. 2014, 118, 1–16. [Google Scholar] [CrossRef]
- Hussain, M.S.; Fareed, S.; Ansari, S.; Rahman, M.A.; Ahmad, I.Z.; Saeed, M. Current approaches toward production of secondary plant metabolites. J. Pharm. Bioallied Sci. 2012, 4, 10–20. [Google Scholar] [CrossRef]
- Filova, A. Production of secondary metabolites in plant tissue cultures. Res. J. Agric. Sci. 2014, 46, 236–245. [Google Scholar]
- Delporte, F.; Pretova, A.; du Jardin, P.; Watillon, B. Morpho-histology and genotype dependence of in vitro morphogenesis in mature embryo cultures of wheat. Protoplasma 2014, 251, 1455–1470. [Google Scholar] [CrossRef] [Green Version]
- Tymoszuk, A.; Zalewska, M.; Lema-Rumińska, J. Regeneration of somatic embryos from in vitro isolated ligulate florets of chrysanthemum. Acta Sci. Pol. Hort. Cult. 2014, 13, 13–22. [Google Scholar]
- Tymoszuk, A.; Miler, N. Silver and gold nanoparticles impact on in vitro adventitious organogenesis in chrysanthemum, gerbera and Cape Primrose. Sci. Hortic. 2019, 257, 108766. [Google Scholar] [CrossRef]
- Ramirez-Malagon, R.; Aguilar-Ramirez, I.; Borodanenko, A.; Perez-Moreno, L.; Barrera-Guerra, J.L.; Nuñez-Palenius, H.G.; Ochoa-Alejo, N. In vitro propagation of ten threatened species of Mammillaria (Cactaceae). In Vitr. Cell. Dev. Biol. Plant 2007, 43, 660–665. [Google Scholar] [CrossRef]
- Sulaiman, I.M. Regeneration of plantlets through organogenesis in the Himalayan yellow poppy, Meconopsis paniculata. Plant Cell Tiss. Organ Cult. 1994, 36, 377–380. [Google Scholar] [CrossRef]
- Park, S.U.; Yu, C.Y.; Chae, Y.A. The first step of biotechnological approaches for alkaloid biosynthesis in Papaveraceae: In vitro plant regenerations. Korean J. Med. Crop Sci. 2004, 12, 428–435. [Google Scholar]
- Economou, A.S. From microcutting rooting to microplant establishment: Key points to consider for maximum success in woody plants. Acta Hortic. 2013, 988, 43–56. [Google Scholar] [CrossRef]
- Tymoszuk, A.; Antkowiak, M. In vitro adventitious organogenesis in Ajania pacifica (Nakai) Bremer et Humphries. BioTechnologia 2018, 4, 335–343. [Google Scholar] [CrossRef]
- Teixeira da Silva, J.A.; Lema-Rumińska, J.; Tymoszuk, A.; Kulpa, D. Regeneration from chrysanthemum flowers: A review. Acta Physiol. Plant. 2015, 37, 36. [Google Scholar] [CrossRef]
- Schenk, R.U.; Hildebrandt, A.C. Medium and techniques for induction and growth of monocotyledonous and dicotyledonous plant cell cultures. Can. J. Bot. 1972, 50, 199–204. [Google Scholar] [CrossRef]
- Elayaraja, D.; Subramanyam, K.; Venkatachalam, V.; Sathish, S.; Kasthurirengan, S.; Ganapathi, A.; Manickavasagam, M. Meta-Topolin (mT) enhances the in vitro regeneration frequency of Sesamum indicum (L.). Biocatal. Agric. Biotechnol. 2019, 21, 101320. [Google Scholar] [CrossRef]
- Doycheva, I.; Yankowa-Tsvetkova, E.; Stanilova, M. Somatic embryogenesis induction in Glaucium flavum Crantz. (Papaveraceae). C. R. l’Académie Bulg. Sci. 2017, 70, 525–530. [Google Scholar]
- Joshi, R.; Kumar, P. Regulation of somatic embryogenesis in crops: A review. Agric. Rev. 2013, 34, 1–20. [Google Scholar]
- Yu, J.; Liu, W.; Liu, J.; Qin, P.; Xu, L. Auxin control of root organogenesis from callus in tissue culture. Front. Plant Sci. 2017, 8, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Chang, C.; Chang, W. Direct somatic embryogenesis on leaf explants of Oncidium Gower Ramsey and subsequent plant regeneration. Plant Cell Rep. 1999, 19, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, M.V.M.; Martins, F.B.; da Cruz, A.C.F.; Pinto de Corvalho, A.C.P.; de Oliveira, E.J.; Otoni, W.C. Somatic embryogenesis in anthurium (Anthurium andraeanum cv. Eidibel) as affected by different explants. Acta Sci. Agron. 2014, 36, 87–98. [Google Scholar] [CrossRef] [Green Version]
- Fierascu, R.C.; Fierascu, I.; Ortan, A.; Georgiev, M.I.; Sieniawska, E. Innovative approaches for recovery of phytoconstituents from medicinal/aromatic plants and biotechnological production. Molecules 2020, 25, 309. [Google Scholar] [CrossRef] [Green Version]
- Mohammad, A.; Alam, P.; Ahmad, M.M.; Ali, A.; Ahmad, J.; Abdin, M.Z. Impact of plant growth regulators (PGRs) on callogenesis and artemisinin content in Artemisia annua L. plants. Indian J. Biotechnol. 2014, 13, 26–33. [Google Scholar]
- Fiedor, J.; Burda, K. Potential role of carotenoids as antioxidants in human health and disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef] [Green Version]
- İnanç, A.L. Chlorophyll: Structural properties, health benefits and its occurrence in virgin olive oils. Akademik Gıda 2011, 9, 26–32. [Google Scholar]
- Li, D.; Wang, P.; Luo, Y.; Zhao, M.; Chen, F. Health benefits of anthocyanins and molecular mechanisms: Update from recent decade. Crit. Rev. Food Sci. Nutr. 2017, 57, 1729–1741. [Google Scholar] [CrossRef]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The role of polyphenols in human health and food systems: A mini-review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef] [Green Version]
- Tarrahi, R.; Rezanejad, F. Callogenesis and production of anthocyanin and chlorophyll in callus cultures of vegetative and floral explants in Rosa gallica and Rosa hybrida (Rosaceae). Turk. J. Bot. 2013, 37, 1145–1154. [Google Scholar] [CrossRef]
- De Matos Nunes, J.; Bertodo, L.O.O.; da Rosa, L.M.G.; Von Poser, G.L.; Rech, S.B. Stress induction of valuable secondary metabolites in Hypericum polyanthemum acclimatized plants. S. Afr. J. Bot. 2014, 94, 182–189. [Google Scholar] [CrossRef] [Green Version]
- Cioć, M.; Kalisz, A.; Żupnik, M.; Pawłowska, B. Different LED Light intensities and 6-benzyladenine concentrations in relation to shoot development, leaf architecture, and photosynthetic pigments of Gerbera jamesonii Bolus in vitro. Agronomy 2019, 9, 358. [Google Scholar] [CrossRef] [Green Version]
- Luczkiewicz, M.; Kokotkiewicz, A.; Glod, D. Plant growth regulators affect biosynthesis and accumulation profile of isoflavone phytoestrogens in high-productive in vitro cultures of Genista tinctoria. Plant Cell Tiss. Organ Cult. 2014, 118, 419–429. [Google Scholar] [CrossRef] [Green Version]
- Erland, L.A.E.; Chattopadhyay, A.; Jones, A.M.P.; Saxena, P.K. Melatonin in plants and plant culture systems: Variability, stability and efficient quantification. Front. Plant Sci. 2016, 7, 1721. [Google Scholar] [CrossRef]
- Ptak, A.; Simlat, M.; Morańska, E.; Skrzypek, E.; Warchoł, M.; Tarakemeh, A.; Laurain-Mattar, D. Exogenous melatonin stimulated Amaryllidaceae alkaloid biosynthesis in in vitro cultures of Leucojum aestivum L. Ind. Crops Prod. 2019, 138, 111458. [Google Scholar] [CrossRef]
- Rakpenthai, A.; Khaksar, G.; Burow, M.; Olsen, C.E.; Sirikantaramas, S. Metabolic changes and increased levels of bioactive compounds in white radish (Raphanus sativus L. cv. 01) sprouts elicited by oligochitosan. Agronomy 2019, 9, 467. [Google Scholar] [CrossRef] [Green Version]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Method. Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Harborne, J.B. Comparative biochemistry of the flavonoids. Phytochemistry 1967, 6, 1569–1573. [Google Scholar] [CrossRef]
- Waterhouse, A.L. Determination of total phenolics. In Current Protocols in Food Analytical Chemistry; Wrolstad, R.E., Ed.; John Wiley & Sons: New York, NY, USA, 2001; pp. I1.1.1–I1.1.8. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Medium Symbol | Plant Growth Regulator (mg·L−1) | Explant Type | Mean | ||||
|---|---|---|---|---|---|---|---|
| BA | Auxin | Leaf | Petiole | Internode | |||
| Callus (%) | |||||||
| M1 | 0.5 | IAA | 1.0 | 66.7 ab | 66.7 ab | 44.4 b | 59.3 C |
| M2 | 0.5 | 2.0 | 88.9 ab | 88.9 ab | 77.8 ab | 85.2 A–C | |
| M3 | 1.0 | 1.0 | 55.5 b | 77.8 ab | 77.7 ab | 70.4 A–C | |
| M4 | 1.0 | 2.0 | 55.5 b | 77.8 ab | 66.7 ab | 66.7 BC | |
| M5 | 0.5 | NAA | 0.5 | 88.9 ab | 55.6 ab | 77.8 ab | 74.1 A–C |
| M6 | 0.5 | 1.0 | 88.9 ab | 77.8 ab | 77.8 ab | 81.5 A–C | |
| M7 | 1.0 | 0.5 | 100 a | 77.8 ab | 88.9 ab | 88.9 AB | |
| M8 | 1.0 | 1.0 | 100 a | 77.8 ab | 100 a | 92.6 AB | |
| M9 | 0.5 | 2,4-D | 0.5 | 100 a | 66.7 ab | 77.8 ab | 81.5 A–C |
| M10 | 0.5 | 1.0 | 100 a | 100 a | 88.9 ab | 96.3 A | |
| M11 | 1.0 | 0.5 | 77.8 ab | 88.9 ab | 88.9 ab | 85.2 A–C | |
| M12 | 1.0 | 1.0 | 100 a | 100 a | 100 a | 100 A | |
| M13 | 0.5 | PIC | 0.5 | 100 a | 100 a | 88.9 ab | 96.3 A |
| M14 | 0.5 | 1.0 | 100 a | 100 a | 100 a | 100 A | |
| M15 | 1.0 | 0.5 | 100 a | 88.9 ab | 100 a | 96.3 A | |
| M16 | 1.0 | 1.0 | 100 a | 100 a | 88.9 ab | 96.3 A | |
| Mean | 65.3 A | 62.4 A | 62.1 A | ||||
| Callus fresh weight [mg] | |||||||
| M1 | 0.5 | IAA | 1.0 | 6.0 ± 2.2 g | 8.3 ± 4.4 g | 12.9 ± 6.3 g | 9.0 E |
| M2 | 0.5 | 2.0 | 12.6 ± 1.0 g | 16.0 ± 5.8 g | 34.0 ± 13.5 fg | 20.9 E | |
| M3 | 1.0 | 1.0 | 20.1 ± 7.2 g | 19.6 ± 14.0 g | 37.5 ± 30 fg | 25.7 E | |
| M4 | 1.0 | 2.0 | 41.8 ± 10.7 fg | 12.9 ± 2.6 g | 20.3 ± 4.7 g | 25.0 E | |
| M5 | 0.5 | NAA | 0.5 | 339.8 ± 70.8 a–g | 147.2 ± 47.3 d–g | 70.0 ± 10.3 e–g | 185.6 CD |
| M6 | 0.5 | 1.0 | 313.6 ± 153.7 a–g | 195.8 ± 50.5 c–g | 231.4 ± 77.1 c–g | 246.9 BC | |
| M7 | 1.0 | 0.5 | 207.4 ± 81.0 c–g | 140.4 ± 35.3 d–g | 55.4 ± 25.1 e–g | 134.4 C–E | |
| M8 | 1.0 | 1.0 | 239.4 ± 74.9 c–g | 99.3 ± 15.4 e–g | 77.8 ± 10.2 e–g | 138.8 C–E | |
| M9 | 0.5 | 2,4-D | 0.5 | 642.8 ± 157.0 a | 466.9 ± 42.4 a–e | 226.1 ± 15.9 c–g | 442.6 A |
| M10 | 0.5 | 1.0 | 548.0 ± 86.4 a–c | 376.1 ± 71.9 a–g | 192.2 ± 55.2 c–g | 372.1 AB | |
| M11 | 1.0 | 0.5 | 617.3 ± 158.8 ab | 521.7 ± 157.2 a–c | 229.4 ± 31.8 c–g | 456.1 A | |
| M12 | 1.0 | 1.0 | 561.6 ± 90.5 a–c | 199.2 ± 57.9 c–g | 228.0 ± 28.7 c–g | 329.6 AB | |
| M13 | 0.5 | PIC | 0.5 | 372.7 ± 20.4 a–g | 479.9 ± 27.2 a–d | 208.7 ± 34.9 c–g | 353.8 AB |
| M14 | 0.5 | 1.0 | 403.7 ± 31.4 a–f | 552.5 ± 31.7 a–c | 250.1 ± 16.9 b–g | 402.1 AB | |
| M15 | 1.0 | 0.5 | 340.8 ± 26.4 a–g | 481.1 ± 48.7 a–d | 280.0 ± 49.5 a–g | 367.3 AB | |
| M16 | 1.0 | 1.0 | 556.8 ± 134.4 a–c | 537.0 ± 8.4 a–c | 258.9 ± 85.1 b–g | 450.9 A | |
| Mean | 326.5 A | 261.6 B | 150.8 C | ||||
| Callus dry weight [%] | |||||||
| M1 | 0.5 | IAA | 1.0 | 30.6 a | 20.5 b–e | 17.5 c–f | 22.8 A |
| M2 | 0.5 | 2.0 | 22.4 b–d | 24.5 a–c | 24.0 a–c | 23.6 A | |
| M3 | 1.0 | 1.0 | 27.1 ab | 25.5 ab | 24.7 a–c | 25.8 A | |
| M4 | 1.0 | 2.0 | 16.1 d–g | 27.3 ab | 30.8 a | 24.7 A | |
| M5 | 0.5 | NAA | 0.5 | 10.0 f–i | 14.4 e–i | 10.3 f–i | 11.6 BC |
| M6 | 0.5 | 1.0 | 9.5 f–i | 10.6 f–i | 14.0 e–i | 11.4 BC | |
| M7 | 1.0 | 0.5 | 11.9 f–i | 14.1 e–i | 9.2 f–i | 11.7 BC | |
| M8 | 1.0 | 1.0 | 11.7 f–i | 13.7 e–i | 13.8 e–i | 13.0 B | |
| M9 | 0.5 | 2,4-D | 0.5 | 9.3 f–i | 10.9 f–i | 12.1 f–i | 10.8 BC |
| M10 | 0.5 | 1.0 | 6.8 g–i | 8.9 f–i | 15.0 e–h | 10.2 BC | |
| M11 | 1.0 | 0.5 | 4.9 i | 8.8 f–i | 9.7 f–i | 7.8 C | |
| M12 | 1.0 | 1.0 | 10.1 f–i | 6.1 hi | 9.1 f–i | 8.4 BC | |
| M13 | 0.5 | PIC | 0.5 | 7.6 g–i | 8.0 f–i | 10.1 f–i | 8.6 BC |
| M14 | 0.5 | 1.0 | 7.3 g–i | 7.6 g–i | 8.5 f–i | 7.8 C | |
| M15 | 1.0 | 0.5 | 7.8 f–i | 9.2 f–i | 11.1 f–i | 9.4 BC | |
| M16 | 1.0 | 1.0 | 6.9 g–i | 7.5 g–i | 8.1 f–i | 7.5 C | |
| Mean | 12.5 B | 13.6 AB | 14.3 A | ||||
| PGRs Concentration | Total Callus (%) | Callus FW (mg) | Callus DW (%) | Non-Embryogenic Callus (%) | Embryogenic Callus (%) | No. of Embryos Per Explant | |
|---|---|---|---|---|---|---|---|
| Cytokinin | Auxin | ||||||
| low | low | 77.8 a | 242.2 ± 37.3 a | 13.4 a | 41.7 ab | 36.1 b | 3.0 ± 0.7 ab |
| low | high | 90.7 a | 260.5 ± 34.1 a | 13.3 a | 31.5 b | 59.3 a | 4.8 ± 0.8 a |
| high | low | 85.2 a | 245.9 ± 38.0 a | 13.7 a | 56.5 a | 28.7 b | 2.2 ± 0.7 b |
| high | high | 88.9 a | 236.1 ± 36.7 a | 13.4 a | 48.8 ab | 40.0 b | 2.5 ± 0.5 ab |
| Medium Symbol | Plant Growth Regulator (mg·L−1) | Explant Type | Mean | ||||
|---|---|---|---|---|---|---|---|
| BA | Auxin | Leaf | Petiole | Internode | |||
| Embryogenic Callus (%) | |||||||
| M1 | 0.5 | IAA | 1.0 | 11.1 bc | 22.2 a–c | 0.0 c | 11.1 DE |
| M2 | 0.5 | 2.0 | 44.4 a–c | 33.3 a–c | 0.0 c | 25.9 CD | |
| M3 | 1.0 | 1.0 | 22.2 a–c | 0.0 c | 0.0 c | 7.4 DE | |
| M4 | 1.0 | 2.0 | 11.1 bc | 33.3 a–c | 0.0 c | 14.8 DE | |
| M5 | 0.5 | NAA | 0.5 | 0.0 c | 0.0 c | 0.0 c | 0.0 E |
| M6 | 0.5 | 1.0 | 55.6 a–c | 44.5 a–c | 22.2 a–c | 40.7 B–D | |
| M7 | 1.0 | 0.5 | 11.1 bc | 0.0 c | 0.0 c | 3.7 DE | |
| M8 | 1.0 | 1.0 | 69.5 a–c | 22.2 a–c | 0.0 c | 30.6 CD | |
| M9 | 0.5 | 2,4-D | 0.5 | 100 a | 55.6 a–c | 33.3 a–c | 63.0 A–C |
| M10 | 0.5 | 1.0 | 88.9 ab | 88.9 ab | 55.6 a–c | 77.8 AB | |
| M11 | 1.0 | 0.5 | 55.6 a–c | 55.6 a–c | 11.1 bc | 40.7 B–D | |
| M12 | 1.0 | 1.0 | 88.9 ab | 22.2 a–c | 11.1 bc | 40.7 B–D | |
| M13 | 0.5 | PIC | 0.5 | 77.8 a–c | 66.7 a–c | 66.7 a–c | 70.4 AB |
| M14 | 0.5 | 1.0 | 100 a | 100 a | 77.8 a–c | 92.6 A | |
| M15 | 1.0 | 0.5 | 55.5 a–c | 88.9 ab | 44.4 a–c | 63.0 A–C | |
| M16 | 1.0 | 1.0 | 77.8 a–c | 88.9 ab | 55.6 a–c | 74.1 AB | |
| Mean | 54.3 A | 45.1 B | 23.6 C | ||||
| No. of embryos per explant | |||||||
| M1 | 0.5 | IAA | 1.0 | 0.3 ± 0.3 cd | 0.8 ± 0.4 b–d | 0.0 d | 0.4 E |
| M2 | 0.5 | 2.0 | 1.6 ± 0.3 b–d | 1.1 ± 0.7 b–d | 0.0 d | 0.9 E | |
| M3 | 1.0 | 1.0 | 1.8 ± 1.0 b–d | 0.0 d | 0.0 d | 0.6 E | |
| M4 | 1.0 | 2.0 | 0.4 ± 0.4 cd | 0.4 ± 0.1 cd | 0.0 d | 0.3 E | |
| M5 | 0.5 | NAA | 0.5 | 0.0 d | 0.0 d | 0.0 d | 0.0 E |
| M6 | 0.5 | 1.0 | 2.0 ± 0.5 b–d | 1.9 ± 1.0 b–d | 0.3 ± 0.2 cd | 1.4 DE | |
| M7 | 1.0 | 0.5 | 0.1 ± 0.1 d | 0.0 d | 0.0 d | 0.0 E | |
| M8 | 1.0 | 1.0 | 2.2 ± 0.2 b–d | 0.3 ± 0.3 cd | 0.0 d | 0.9 E | |
| M9 | 0.5 | 2,4-D | 0.5 | 9.2 ± 2.7 a–d | 7.1 ± 3.6 a–d | 2.3 ± 0.2 b–d | 6.2 A–C |
| M10 | 0.5 | 1.0 | 8.9 ± 1.3 a–d | 9.1 ± 3.1 a–d | 3.9 ± 1.8 a–d | 7.3 AB | |
| M11 | 1.0 | 0.5 | 5.1 ± 4.2 a–d | 4.3 ± 3.4 a–d | 0.1 ± 0.1 d | 3.2 C–E | |
| M12 | 1.0 | 1.0 | 5.6 ± 0.3 a–d | 2.1 ± 2.1 b–d | 1.3 ± 1.3 b–d | 3.0 C–E | |
| M13 | 0.5 | PIC | 0.5 | 8.8 ± 3.5 a–d | 4.7 ± 1.9 a–d | 2.8 ± 0.9 b–d | 5.4 BC |
| M14 | 0.5 | 1.0 | 9.7 ± 1.3 ab | 11.4 ± 0.5 a | 7.2 ± 2.1 a–d | 9.4 A | |
| M15 | 1.0 | 0.5 | 2.9 ± 2.2 b–d | 9.3 ± 3.9 a–c | 2.2 ± 0.6 b–d | 4.8 B–D | |
| M16 | 1.0 | 1.0 | 5.9 ± 1.1 a–d | 6.3 ± 1.7 a–d | 5.9 ± 3.4 a–d | 6.0 A–C | |
| Mean | 4.0 A | 3.7 A | 1.6 B | ||||
| PGRs Concentration | Chlorophyll ct (µg·g−1 FW) | Chlorophyll a/b (µg·g−1 FW) | Carotenoids (µg·g−1 FW) | Chlorophyll ct/Carotenoids | Anthocyanins (µg·g−1 FW) | Polyphenols (mg·g−1 FW) | |
|---|---|---|---|---|---|---|---|
| Cytokinin | Auxin | ||||||
| low | low | 93.0 ± 8.5 a | 0.7 ± 0.04 a | 18.4 ± 1.2 a | 4.6 ± 0.4 a | 31.3 ± 2.4 a | 2.0 ± 0.07 a |
| low | high | 42.2 ± 3.5 b | 0.7 ± 0.09 a | 14.6 ± 0.7 b | 3.2 ± 0.4 ab | 25.1 ± 2.7 a | 1.9 ± 0.05 a |
| high | low | 50.3 ± 9.0 b | 1.2 ± 0.18 a | 16.4 ± 0.8 ab | 2.7 ± 0.4 b | 30.5 ± 4.1 a | 2.0 ± 0.09 a |
| high | high | 70.4 ± 11.0 ab | 0.8 ± 0.14 a | 17.4 ± 1.1 ab | 4.2 ± 0.6 ab | 35.6 ± 2.7 a | 2.1 ± 0.08 a |
| Trait | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
|---|---|---|---|---|---|---|---|---|
| 1. Non-embryogenic callus | 1.00 | |||||||
| 2. Embryogenic callus | −0.87 | 1.00 | ||||||
| 3. No. of embryos | −0.70 | 0.83 | 1.00 | |||||
| 4. Chlorophyll a | 0.36 | −0.39 | −0.34 | 1.00 | ||||
| 5. Chlorophyll b | 0.30 | −0.33 | −0.31 | 0.93 | 1.00 | |||
| 6. Carotenoids | 0.23 | −0.27 | −0.23 | 0.39 | 0.36 | 1.00 | ||
| 7. Anthocyanins | 0.39 | −0.43 | −0.32 | 0.33 | 0.32 | 0.20 | 1.00 | |
| 8. Polyphenols | 0.41 | −0.48 | −0.36 | 0.33 | 0.33 | 0.28 | 0.78 | 1.00 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kulus, D.; Tymoszuk, A. Induction of Callogenesis, Organogenesis, and Embryogenesis in Non-Meristematic Explants of Bleeding Heart and Evaluation of Chemical Diversity of Key Metabolites from Callus. Int. J. Mol. Sci. 2020, 21, 5826. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165826
Kulus D, Tymoszuk A. Induction of Callogenesis, Organogenesis, and Embryogenesis in Non-Meristematic Explants of Bleeding Heart and Evaluation of Chemical Diversity of Key Metabolites from Callus. International Journal of Molecular Sciences. 2020; 21(16):5826. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165826
Chicago/Turabian StyleKulus, Dariusz, and Alicja Tymoszuk. 2020. "Induction of Callogenesis, Organogenesis, and Embryogenesis in Non-Meristematic Explants of Bleeding Heart and Evaluation of Chemical Diversity of Key Metabolites from Callus" International Journal of Molecular Sciences 21, no. 16: 5826. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165826