Toll-Like Receptor 5 of Golden Pompano Trachinotus ovatus (Linnaeus 1758): Characterization, Promoter Activity and Functional Analysis

 ,
,
Abstract
:1. Introduction
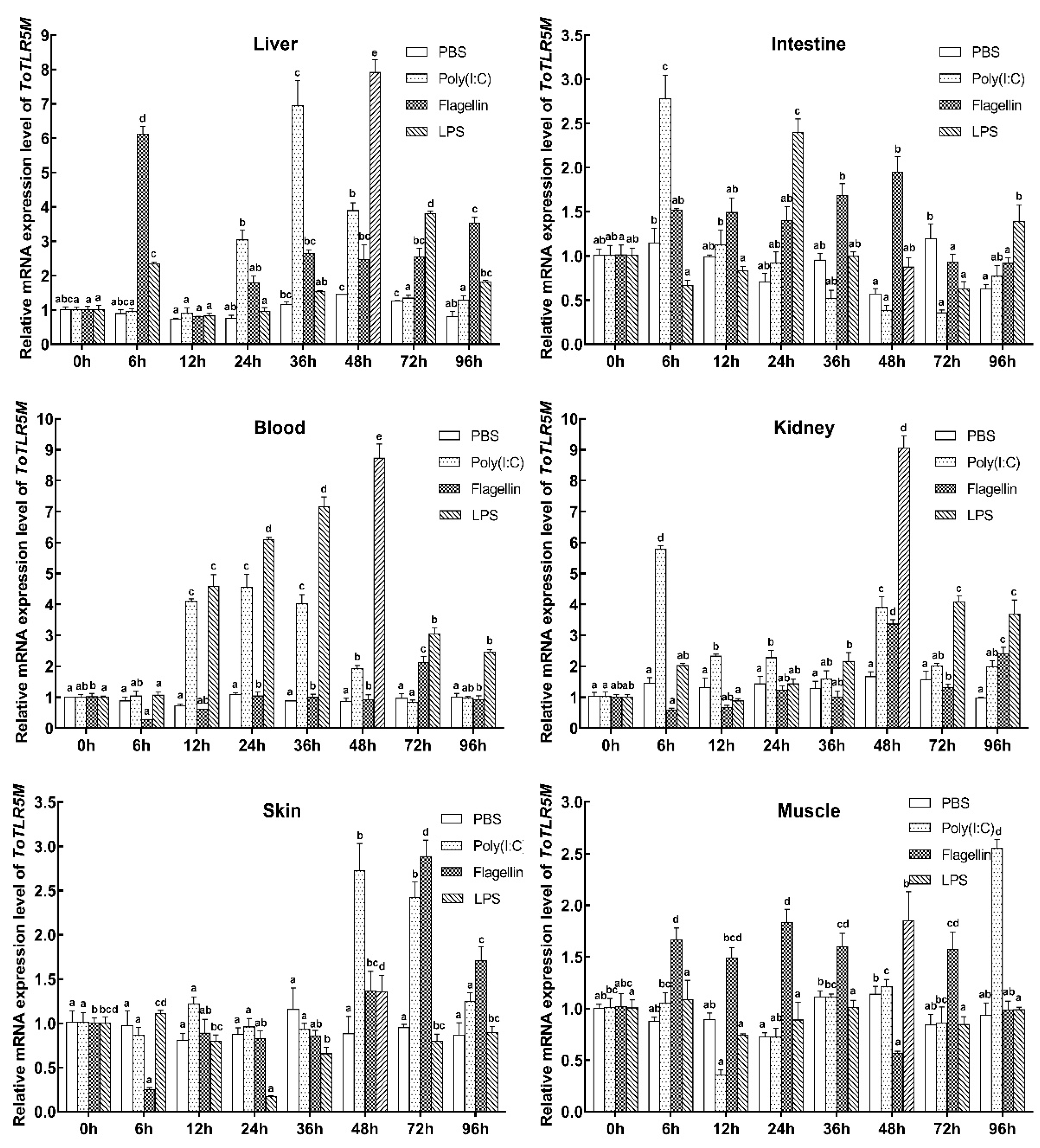
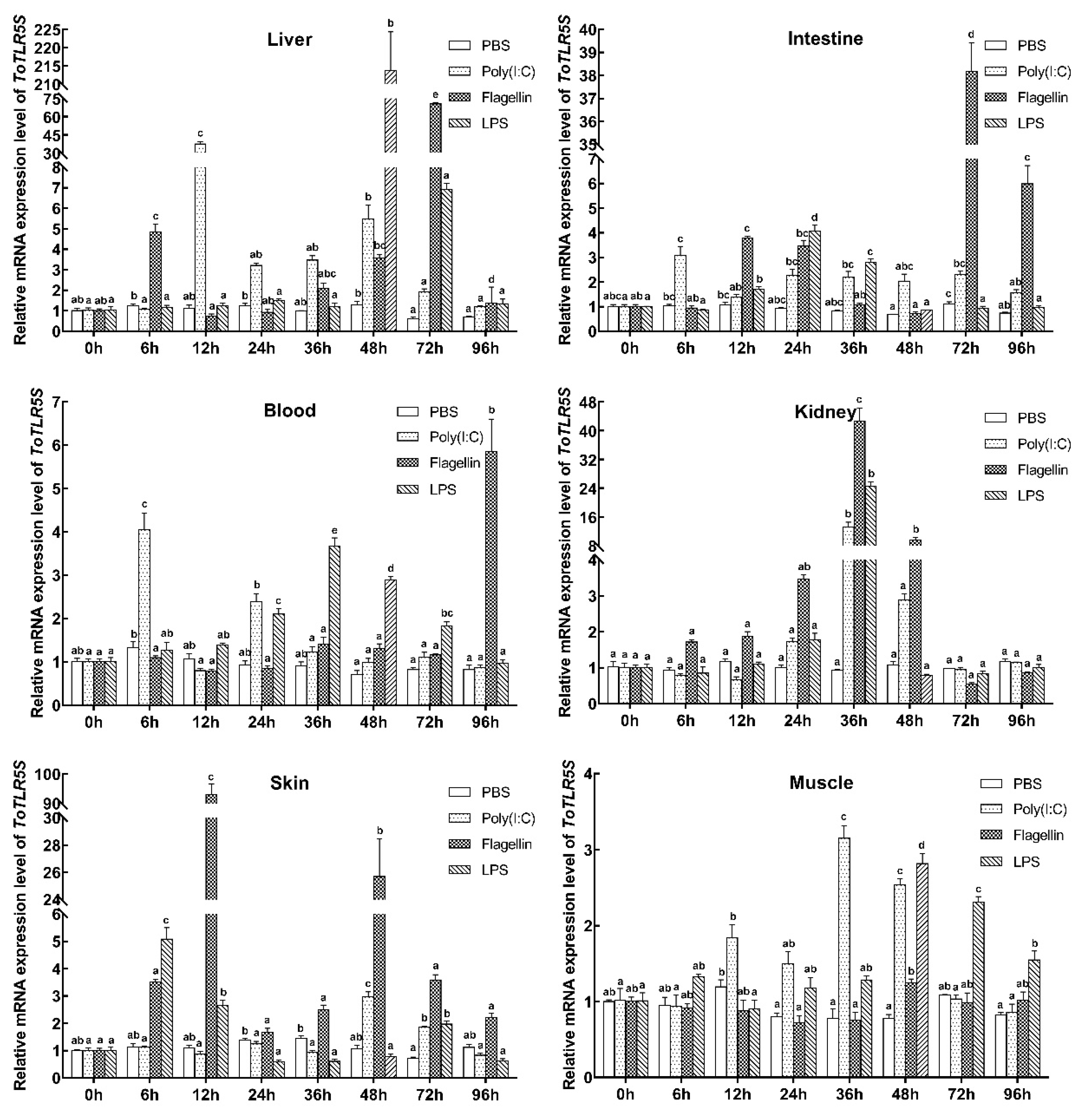
2. Results
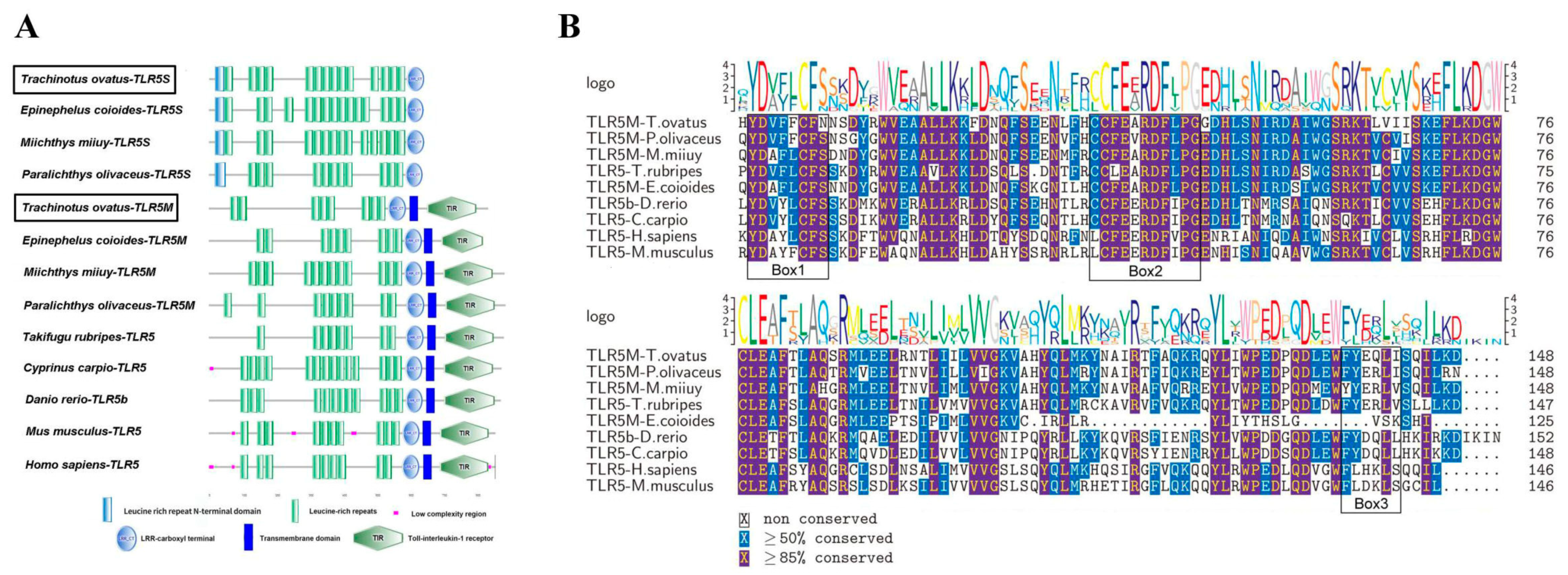
2.1. Sequence Characterization of ToTLR5M and ToTLR5S
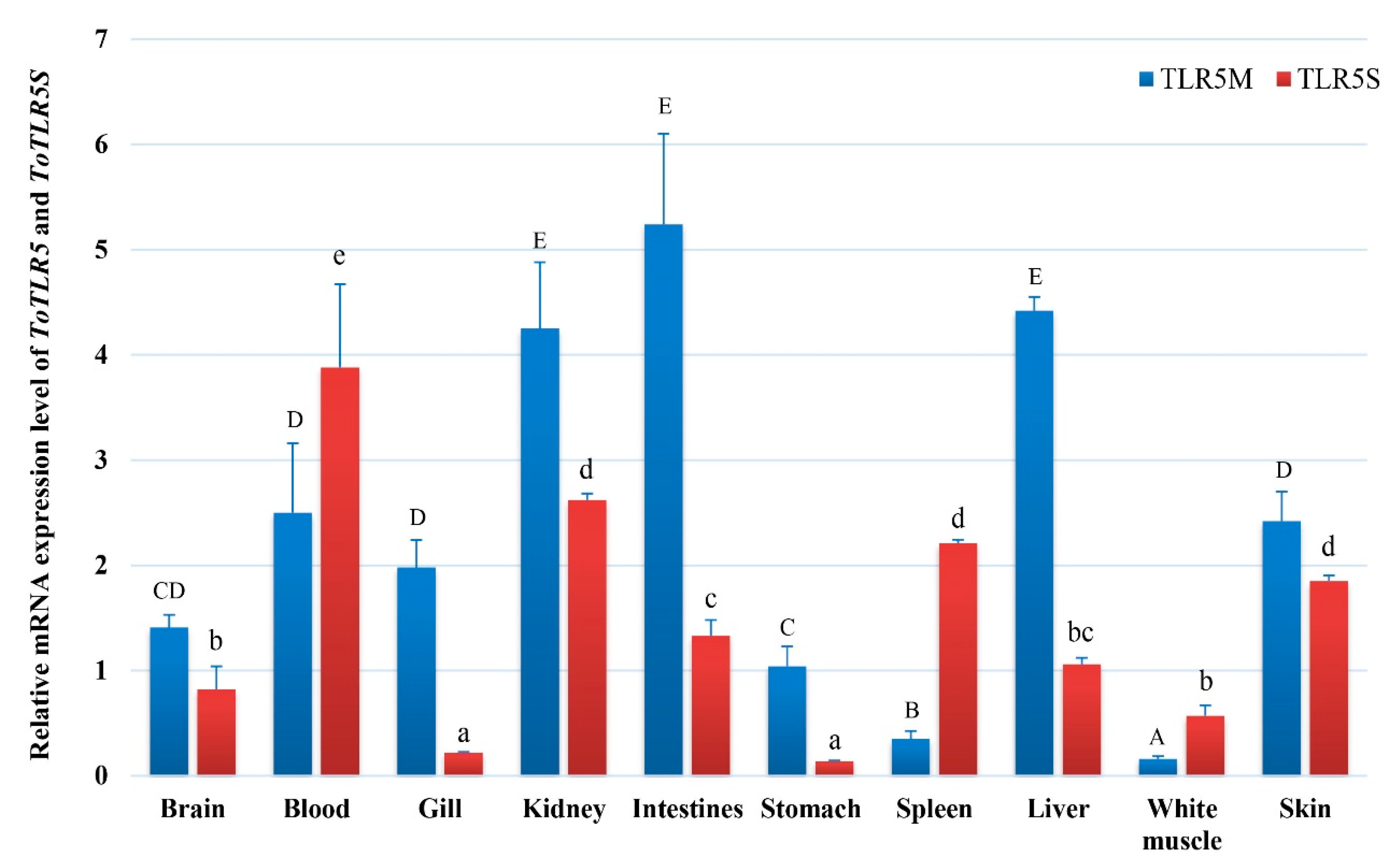
2.2. Tissue Expression of ToTLR5M and ToTLR5S
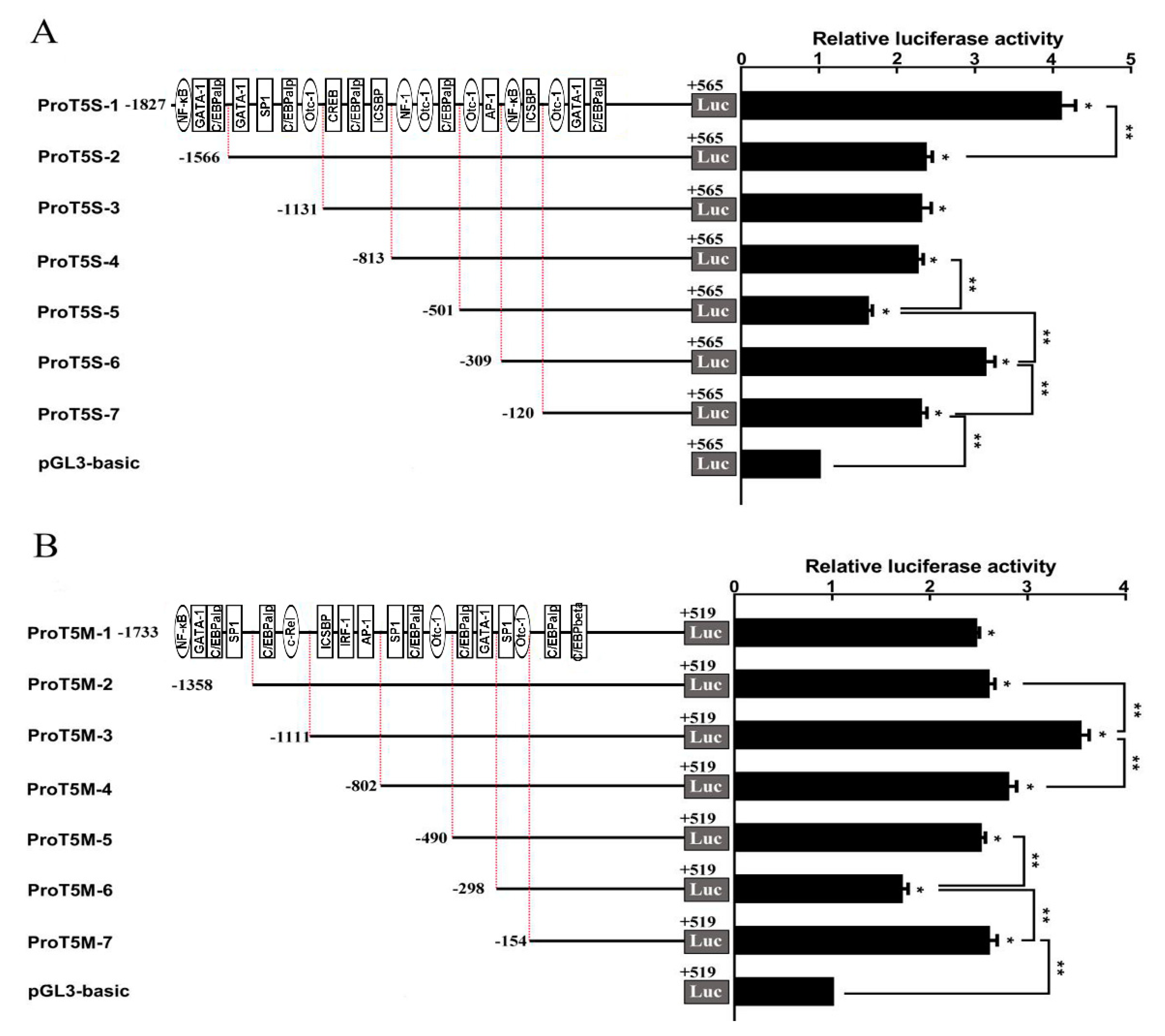
2.3. Promoter Activity of Two ToTLR5s
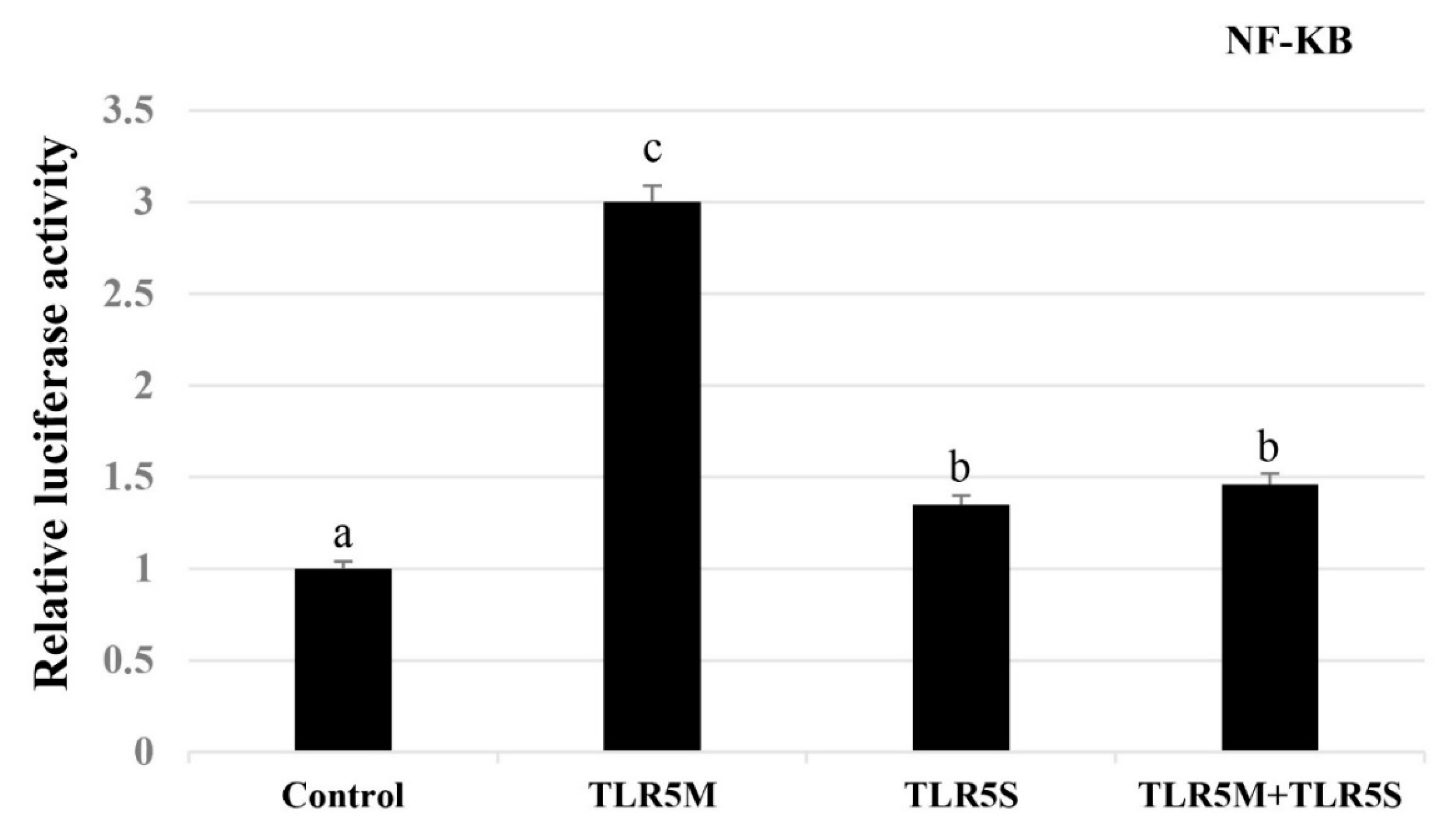
2.4. Effect of Overexpression of Two ToTLR5s on NF-κB Activity
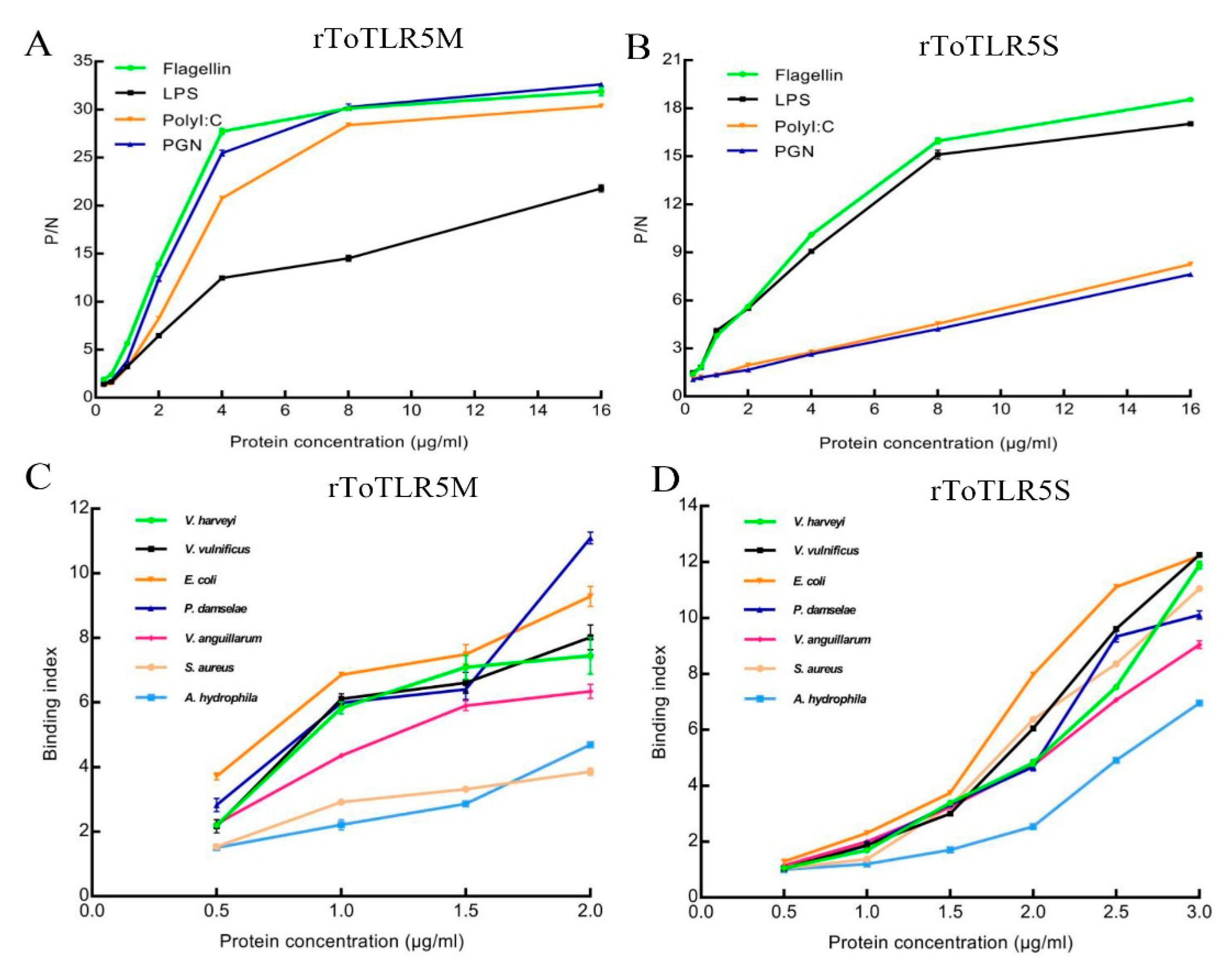
2.5. Binding of Two rToTLR5s to Bacteria and PAMPs
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Fish and Challenge Experiments
4.3. RNA Extraction and Gene Cloning
4.4. Bioinformatics
4.5. Cloning of the 5′-flanking Sequence and Its Promoter Activity
4.6. Quantitative Real-time PCR and Statistical Analysis
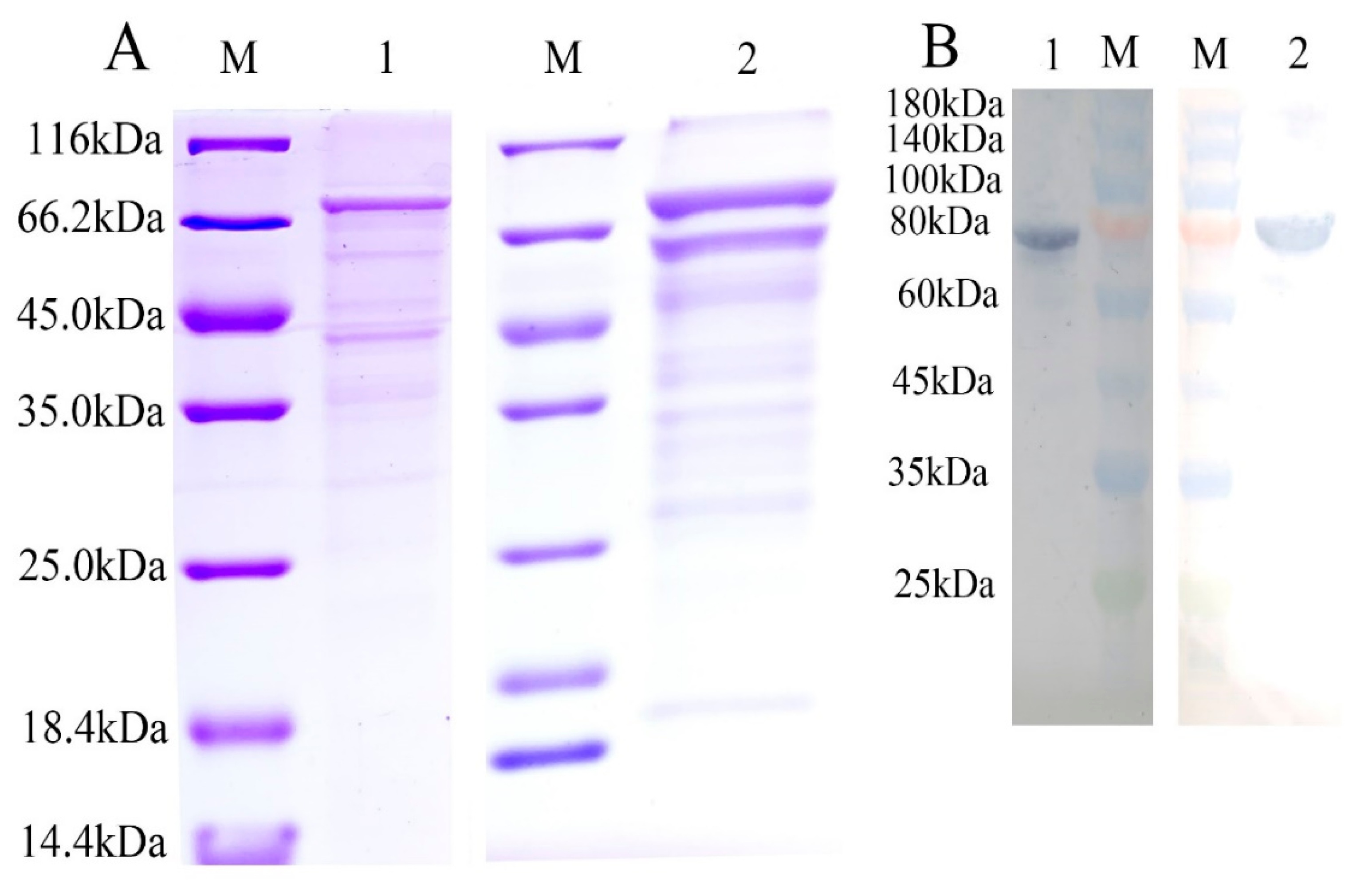
4.7. Expression and Purification of Recombinant Two ToTLR5s
4.8. Assay for the Binding of Bacteria and PAMPs
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| TLRs | Toll-like receptors |
| ORF | open reading frame |
| poly(I:C) | polyinosinic:polycytidylic acid |
| LPS | lipopolysaccharide |
| PAMPs | pathogen-associated molecular patterns |
| PGN | peptidoglycan |
| PRR | pathogen recognition receptor |
| LRRs | leucine-rich repeats |
| GPS | Trachinotus ovatus snout tissue |
| qRT-PCR | quantitative real-time polymerase chain reaction |
| WT | Western blotting |
| SDS-PAGE | sodium dodecyl sulfate polyacrylamide gel electrophoresis |
| ELISA | enzyme-linked immunosorbent assay |
References
- Purcell, M.K.; Smith, K.D.; Hood, L.; Winton, J.R.; Roach, J.C. Conservation of Toll-like receptor signaling pathways in teleost fish. Comp. Biochem. Physiol. Part D Genom. Proteom. 2006, 1, 77–88. [Google Scholar]
- Medzhitov, R. Toll-like receptors and innate immunity. Nat. Rev. Immunol. 2001, 1, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Kaisho, T.; Akira, S. Toll-like receptors. Annu. Rev. Immunol. 2003, 21, 335–376. [Google Scholar] [CrossRef] [PubMed]
- Akira, S.; Takeda, K. Toll-like receptor signalling. Nat. Rev. Immunol. 2004, 4, 499–511. [Google Scholar] [CrossRef]
- Janeway, C.A.; Medzhitov, R. Innate immune recognition. Annu. Rev. Immunol. 2002, 20, 197–216. [Google Scholar] [CrossRef] [Green Version]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [Green Version]
- Kawai, T.; Akira, S. The roles of TLRs, RLRs and NLRs in pathogen recognition. Int. Immunol. 2009, 21, 317–337. [Google Scholar] [CrossRef] [Green Version]
- Palti, Y. Toll-like receptors in bony fish: From genomics to function. Dev. Comp. Immunol. 2011, 35, 1263–1272. [Google Scholar] [CrossRef]
- Means, T.K.; Golenbock, D.T.; Fenton, M.J. Structure and function of Toll-like receptor proteins. Life Sci. 2000, 68, 241–258. [Google Scholar] [CrossRef]
- Gewirtz, A.T.; Navas, T.A.; Lyons, S.; Godowski, P.J.; Madara, J.L. Cutting edge: Bacterial flagellin activates basolaterally expressed TLR5 to induce epithelial proinflammatory gene expression. J. Immunol. 2001, 167, 1882–1885. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, F.; Smith, K.D.; Ozinsky, A.; Hawn, T.R.; Yi, E.C.; Goodlett, D.R.; Eng, J.K.; Akira, S.; Underhill, D.M.; Aderem, A. The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nature 2001, 410, 1099–1103. [Google Scholar] [CrossRef] [PubMed]
- Didierlaurent, A.; Ferrero, I.; Otten, L.A.; Dubois, B.; Reinhardt, M.; Carlsen, H.; Blomhoff, R.; Akira, S.; Kraehenbuhl, J.P.; Sirard, J.C. Flagellin promotes myeloid differentiation factor 88-dependent development of Th2-type response. J. Immunol. 2004, 172, 6922–6930. [Google Scholar] [CrossRef] [PubMed]
- Basu, M.; Swain, B.; Maiti, N.K.; Routray, P.; Samantaa, M. Inductive expression of toll-like receptor 5 (TLR5) and associated downstream signaling molecules following ligand exposure and bacterial infection in the Indian major carp, mrigal (Cirrhinus mrigala). Fish Shellfish Immunol. 2012, 32, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Mizel, S.B.; Honko, A.N.; Moors, M.A.; Smith, P.S.; West, A.P. Induction of macrophage nitric oxide production by Gram-negative flagellin involves signaling via heteromeric Toll-like receptor 5/Toll-like receptor 4 complexes. J. Immunol. 2003, 170, 6217–6223. [Google Scholar] [CrossRef] [PubMed]
- Oshiumi, H.; Tsujita, T.; Shida, K.; Matsumoto, M.; Ikeo, K.; Seya, T. Prediction of the prototype of the human Toll-like receptor gene family from the pufferfish, Fugu rubripes, genome. Immunogenetics 2003, 54, 791–800. [Google Scholar] [CrossRef]
- Tsujita, T.; Tsukada, H.; Nakao, M.; Oshiumi, H.; Matsumoto, M.; Seya, T. Sensing bacterial flagellin by membrane and soluble orthologs of Toll-like receptor 5 in rainbow trout (Onchorhynchus mikiss). J. Biol. Chem. 2004, 279, 48588–48597. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.D.; Asahi, T.; Kondo, H.; Hirono, I.; Aoki, T. Molecular cloning and expression study on Toll-like receptor 5 paralogs in Japanese flounder, Paralichthys olivaceus. Fish Shellfish Immunol. 2010, 29, 630–638. [Google Scholar] [CrossRef]
- Bai, J.; Li, Y.; Deng, Y.; Huang, Y.Q.; He, S.H.; Dai, J.; Zhao, S.Z.; Dan, X.M.; Luo, X.C. Molecular identification and expression analysis of TLR5M and TLR5S from orange-spotted grouper (Epinephelus coioides). Fish Shellfish Immunol. 2017, 63, 97–102. [Google Scholar] [CrossRef]
- Meijer, A.H.; Gabby, K.S.F.; Medina, R.I.A.; He, S.; Bitter, W.; Ewa, S.J.B.; Spaink, H.P. Expression analysis of the Toll-like receptor and TIR domain adaptor families of zebrafish. Mol. Immunol. 2004, 40, 773–783. [Google Scholar] [CrossRef]
- Jiang, Y.; He, L.; Ju, C.; Pei, Y.; Ji, M.; Li, Y.; Liao, L.J.; Jang, S.H.; Zhu, Z.Y.; Wang, Y.P. Isolation and expression of grass carp toll-like receptor 5a (CiTLR5a) and 5b (CiTLR5b) gene involved in the response to flagellin stimulation and grass carp reovirus infection. Fish Shellfish Immunol. 2015, 44, 88–99. [Google Scholar] [CrossRef]
- Umasuthan, N.; Bathige, S.; Thulasitha, W.S.; Jayasooriya, R.G.P.T.; Younhee, S.; Lee, J. Identification of a gene encoding a membrane-anchored toll-like receptor 5 (TLR5M) in Oplegnathus fasciatus that responds to flagellin challenge and activates NF-κB [J]. Fish Shellfish Immunol. 2017, 62, 276–290. [Google Scholar] [CrossRef] [PubMed]
- Jayaramu, P.K.; Tripathi, G.; Kumar, A.P.; Keezhedath, J.; Pathan, M.K.; Kurcheti, P.P. Studies on expression pattern of toll-like receptor 5 (TLR5) in Edwardsiella tarda infected Pangasianodon hypophthalmus. Fish Shellfish Immunol. 2017, 63, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, N.; Kondo, M.; Kono, T.; Sakai, M.; Hikima, J.I. Nonconservation of TLR5 activation site in Edwardsiella tarda flagellin decreases expression of interleukin-1β and NF-κB genes in Japanese flounder, Paralichthys olivaceus. Fish Shellfish Immunol. 2019, 87, 765–771. [Google Scholar] [CrossRef] [PubMed]
- Tsoi, S.; Park, K.C.; Kay, H.H.; O’Brien, T.J.; Podor, E.; Sun, G.; Douglas, S.E.; Brown, L.L.; Johnson, S.C. Identification of a transcript encoding a soluble form of toll-like receptor 5 (TLR5) in Atlantic salmon during Aeromonas salmonicida infection. Vet. Immunol. Immunop. 2006, 109, 183–187. [Google Scholar] [CrossRef]
- Baoprasertkul, P.; Xu, P.; Peatman, E.; Kucuktas, H.; Liu, Z. Divergent Toll-like receptors in catfish, Ictalurus punctatus: TLR5S, TLR20, TLR21. Fish Shellfish Immunol. 2007, 23, 1218–1230. [Google Scholar] [CrossRef]
- Zhang, W.T.; Xiang, J.S.; Li, H.L.; Zhang, N.; Dong, Z.D.; Gao, F.T.; Chen, S.L. Molecular cloning, characterization, and expression of three TLR5S splicing variants in half-smooth tongue sole. J. Fish. Sci. China 2016, 23, 10–20. [Google Scholar]
- Huo, R.; Zhao, X.; Han, J.; Xu, T.J. Genomic organization, evolution and functional characterization of soluble toll-like receptor 5 (TLR5S) in miiuy croaker (Miichthys miiuy). Fish Shellfish Immunol. 2018, 80, 109–114. [Google Scholar] [CrossRef]
- Sun, L.Y.; Guo, H.Y.; Zhu, C.Y.; Ma, Z.H.; Jiang, S.G.; Zhang, D.C. Genetic polymorphism of breeding populations of golden pompano (Trachinotus ovatus). South China Fish Sci. 2014, 10, 67–71. [Google Scholar]
- Zhen, P.L.; Ma, Z.H.; Guo, H.Y.; Jiang, S.G.; Zhang, D.C. Ontogenetic development of caudal skeletons in Trachinotus ovatus larvae, South China. Fish Sci. 2014, 10, 45–50. [Google Scholar]
- Su, Y.L.; Feng, J.; Guo, Z.X.; Xu, L.W.; Wang, J.Y. Histopathological analysis of golden pompano Trachinotus ovatus infected with Photobacterium damselae subsp. piscicida. Mar. Sci. 2012, 36, 75–81. [Google Scholar]
- Bell, J.K.; Mullen, G.E.; Leifer, C.A.; Mazzoni, A.; Davies, D.R.; Segal, D.M. Leucine-rich repeats and pathogen recognition in Toll-like receptors. Trends Immunol. 2003, 24, 528–533. [Google Scholar] [CrossRef]
- Bell, J.K.; Botos, I.; Hall, P.R.; Askins, J.; Shiloach, J.; Segal, D.M.; Davies, D.R. The molecular structure of the Toll-like receptor 3 ligand-binding domain. Proc. Natl. Acad. Sci. USA 2005, 102, 10976–10980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slack, J.L.; Schooley, K.; Bonnert, T.P.; Mitcham, J.L.; Qwarnstrom, E.E.; Sims, J.E.; Dower, S.K. Identification of two major sites in the type I interleukin-1 receptor cytoplasmic region responsible for coupling to pro-inflammatory signaling pathways. J. Biol. Chem. 2000, 275, 4670–4678. [Google Scholar] [CrossRef] [Green Version]
- Bilodeau, A.L.; Waldbieser, G.C. Activation of TLR3 and TLR5 in channel catfish exposed to virulent Edwardsiella ictaluri. Dev. Comp. Immunol. 2005, 29, 713–721. [Google Scholar] [CrossRef]
- Zhang, X.T.; Zhang, G.R.; Shi, Z.C.; Yuan, Y.J.; Zheng, H.; Lin, L.; Wei, K.J.; Ji, W. Expression analysis of nine Toll-like receptors in yellow catfish (Pelteobagrus fulvidraco) responding to Aeromonas hydrophila challenge. Fish Shellfish Immunol. 2017, 63, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Chen, J.; Li, S.; Hu, G.B. Molecular cloning and expression study on Toll-like receptor 5M in turbot, Scophthalmus maximus. Dev. Comp. Immunol. 2018, 85, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Guo, L.; Zhu, K.C.; Guo, H.Y.; Liu, B.; Jiang, S.G.; Zhang, D.C. Genomic structure and molecular characterization of Toll-like receptors 1 and 2 from golden pompano Trachinotus ovatus (Linnaeus, 1758) and their expression response to three types of pathogen associated molecular patterns. Dev. Comp. Immunol. 2018, 86, 34–40. [Google Scholar] [CrossRef]
- Zhu, K.C.; Guo, H.Y.; Zhang, N.; Liu, B.S.; Guo, L.; Jiang, S.G.; Zhang, D.C. Functional characterization of IRF8 regulation of type II IFN in golden pompano (Trachinotus ovatus). Fish Shellfish Immunol. 2019, 94, 1–9. [Google Scholar] [CrossRef]
- Zhu, K.C.; Liu, B.S.; Zhang, N.; Guo, H.Y.; Guo, L.; Jiang, S.G.; Zhang, D.C. Interferon regulatory factor 2 plays a positive role in interferon gamma expression in golden pompano, Trachinotus ovatus (Linnaeus 1758). Fish Shellfish Immunol. 2020, 96, 107–113. [Google Scholar] [CrossRef]
- Zhu, K.C.; Zhang, N.; Liu, B.S.; Guo, L.; Guo, H.Y.; Jiang, S.G.; Zhang, D.C. Functional analysis of IRF1 reveals its role in the activation of the Type I IFN pathway in golden pompano, Trachinotus ovatus (Linnaeus 1758). Int. J. Mol. Sci. 2020, 21, 2652. [Google Scholar] [CrossRef] [Green Version]
- Zhu, K.C.; Guo, H.Y.; Zhang, N.; Guo, L.; Liu, B.S.; Jiang, S.G.; Zhang, D.C. Structural and expression analysis of golden pompano Trachinotus ovatus IRF5 and its role in regulation of type I IFN. Fish Shellfish Immunol. 2020, 97, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J. Cloning, expression and functional analysis of tlr5/tlr22 and associated downstream signaling molecules in qihe crucian carp (Carassius auratus). Ph.D. Thesis, The Henan Normal University, Xinxiang, China, July 2016. [Google Scholar]
- Gao, Q.X.; Yue, Y.F.; Min, M.H.; Peng, S.M.; Shi, Z.H.; Sheng, W.Q.; Zhang, T. Characterization of TLR5 and TLR9 from silver pomfret (Pampus argenteus) and expression profiling in response to bacterial components. Fish Shellfish Immunol. 2018, 80, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Thakur, B.K.; Dasgupta, N.; Ta, A.; Das, S. Physiological TLR5 expression in the intestine is regulated by differential DNA binding of Sp1/Sp3 through simultaneous Sp1 dephosphorylation and Sp3 phosphorylation by two different PKC isoforms. Nucleic Acids Res. 2016, 12, 5658–5672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, J.Y.; Nam, B.H.; Kong, H.J.; Kim, Y.O.; Kim, W.J.; Kim, B.S.; Kim, K.K.; Lee, S.J. Maximal transcriptional activation of piscine soluble Toll-like receptor 5 by the NF-κB subunit p65 and flagellin. Fish Shellfish Immunol. 2011, 6, 881–886. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Shen, Y.; Xu, X.; Hu, M.Y.; Li, J.L. Identification and characterization of the TLR18 gene in grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2015, 2, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, M.; Seya, T. TLR3: Interferon induction by double-stranded RNA including poly (I: C). Adv. Drug Deliver. Rev. 2008, 7, 805–812. [Google Scholar] [CrossRef]
- Zhang, D.C.; Guo, L.; Guo, H.Y.; Zhu, K.C.; Li, S.Q.; Zhang, Y.; Zhang, N.; Liu, B.S.; Jiang, S.G.; Li, J.T. Chromosome-level genome assembly of golden pompano (Trachinotus ovatus) in the family Carangidae. Sci. Data 2019, 6, 216. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.Y.; Zhang, D.C.; Jiang, S.G.; Guo, H.Y.; Zhu, C.Y. Isolation and characterization of 21 polymorphic microstatellites in golden pompano Trachinotus ovatus. Conserv. Genet. Resour. 2013, 5, 1107–1109. [Google Scholar] [CrossRef]
- Yu, Y.; Wei, S.; Wang, Z.; Huang, X.; Huang, Y.; Cai, J.; Li, C.; Qin, Q. Establishment of a new cell line from the snout tissue of golden pompano Trachinotus ovatus, and its application in virus susceptibility. Fish Biol. 2016, 88, 2251–2262. [Google Scholar] [CrossRef]
- Zhu, K.C.; Song, L.; Guo, H.Y.; Guo, L.; Zhang, N.; Liu, B.S.; Jiang, S.G.; Zhang, D.C. Elovl4a participates in LC-PUFA biosynthesis and is regulated by PPARαβ in golden pompano Trachinotus ovatus (Linnaeus 1758). Sci. Rep. 2019, 9, 4684. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using realtime quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Sun, L. CsSAP, a teleost serum amyloid P component, interacts with bacteria, promotes phagocytosis, and enhances host resistance against bacterial and viral infection. Dev. Comp. Immunol. 2016, 55, 12–20. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subject and Primers | Nucleotide Sequence |
|---|---|
| Primers for cDNA sequence cloning | |
| TLR5M-F | GCTAGCATGAGGACGCCGGCCCTTCACT |
| TLR5M-R | GGTACCTCACATAGCAATTGTTGGGATG |
| TLR5S-F | GCTAGCATGTGGCTGCTGGGTCTCCAGG |
| TLR5S-R | GGTACCTTACTGCTGTGTGAGCTGAGCA |
| TLR5M-GSP1 | GACACCAAGGTGAAAAAGATTG |
| TLR5M-GSP2 | GACCAAAAGTAGTGTATTCATA |
| TLR5S-GSP1 | ACCAATGTCACCTTCTTGAGTC |
| TLR5S-GSP2 | ACGTATGTCATGTTGATTTGGG |
| Primers for DNA sequence cloning | |
| TLR5M-F1 | CAACAGCAAAGTTAGATTTACCAAT |
| TLR5M-R1 | CAATTTGAGGTTTAATTGTGTGAAC |
| TLR5M-F2 | AGAGACTCCATACTGACGGAAAGC |
| TLR5M-R2 | AGGAACATGCTATATATCAGCTGT |
| TLR5M-F3 | AAATATGAAGCTACAGCCAGACGC |
| TLR5M-R3 | TGTGAGATTTTGTTTTAATTCTTA |
| TLR5S-F1 | GTTTGTGCTGCACAATCACAGTAATG |
| TLR5S-R1 | CCGTTGTAGGATAGGTCGAGTTTCTG |
| TLR5S-F2 | TCAATTTCCTAAACTTGAACTCTG |
| TLR5S-R2 | GCCCATTTCAGGGGATTTTTTTAT |
| Deletion mutant construction | |
| Pro-TLR5M-F1 | AAGCTTACCTGCGGTATGGAAGAATGCCCTG |
| Pro-TLR5M-F2 | AAGCTTACCCTCGCACATCACTTCCTTAACC |
| Pro-TLR5M-F3 | AAGCTTACCTAGCTACGGAGGACAGGACTG |
| Pro-TLR5M-F4 | AAGCTTACCAGGTCGGCCGCCCAGAAACC |
| Pro-TLR5M-F5 | AAGCTTACCGACTATGGAAAAGTTACAAG |
| Pro-TLR5M-F6 | AAGCTTACCTACAAGTTAAAGAGCAGAGAG |
| Pro-TLR5M-F7 | AAGCTTACCGAAATGCTCCAGGCGGGTCA |
| Pro-TLR5M-R | GGTACCTTAGCTGTCCTCGTCCCACCAAGGCG |
| Pro-TLR5S-F1 | AAGCTTACCGGTCTTTCACTGACTTCCCTAC |
| Pro-TLR5S-F2 | AAGCTTACCTCCAGTGGTGAAAAAGCAGCTG |
| Pro-TLR5S-F3 | AAGCTTACCACCACTCTCTTTAATTATTTACAG |
| Pro-TLR5S-F4 | AAGCTTACCCATTGGGCGCTCAGAAATCACTTG |
| Pro-TLR5S-F5 | AAGCTTACCCTCCTACTCCTAAAAGTACAATA |
| Pro-TLR5S-F6 | AAGCTTACCGTCTTAATATCTGAAAGAGGAA |
| Pro-TLR5S-F7 | AAGCTTACCGATGCTGTTTTCTGTACTACTGAC |
| Pro-TLR5S-R | GGTACCTTACCTCACACTGCTTGGTATAATCC |
| Pro-NF-кB-F | GGTACCGTAAGATCATGTGAACTACC |
| Pro-NF-кB-R | CTCGAGGTATGAAGGTAGTGGTCGTC |
| Primers for qRT-PCR | |
| qTLR5M-F | TTCAGTCACTCATCTTCCTCAG |
| qTLR5M -R | TCTCGTTCAGCCACTTCAG |
| qTLR5S-F | TCAACCTCTCCAACAACTTCA |
| qTLR5S-R | CGGTCATCCAAGCCAGAA |
| EF1α-F | AAGCCAGGTATGGTTGTCAACTTT |
| EF1α-R | CGTGGTGCATCTCCACAGACT |
| Primers for recombinant expression | |
| TLR5M-ED-F | GGATCCGAATTCCGGACTATGCTGGAGCCAAGGGCATTTGC |
| TLR5M-ED-R | GTGGTGCTCGAGTGCGGCCTTATTAATCCTCTTTACATGGTTCGATG |
| TLR5S-ED-F | GGATCCGAATTCCGGACTATGTCATGCCTCATAACGGGCTC |
| TLR5S-ED-R | GTGGTGCTCGAGTGCGGCCTTATTACTGCTGTGTGAGCTGAGCAG |
| Primers for eukaryotic recombinant vector construction | |
| TLR5M-F | TGGACTAGTGGATCCATGAGGACGCCGGCCCTT |
| TLR5M-R | TTTAAACTTAAGCTTCATAGCAATTGTTGGGAT |
| TLR5S-F | TGGACTAGTGGATCCATGTGGCTGCTGGGTCTC |
| TLR5S-R | TTTAAACTTAAGCTTCTGCTGTGTGAGCTGAGC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, K.-C.; Wu, M.; Zhang, D.-C.; Guo, H.-Y.; Zhang, N.; Guo, L.; Liu, B.-S.; Jiang, S.-G. Toll-Like Receptor 5 of Golden Pompano Trachinotus ovatus (Linnaeus 1758): Characterization, Promoter Activity and Functional Analysis. Int. J. Mol. Sci. 2020, 21, 5916. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165916
Zhu K-C, Wu M, Zhang D-C, Guo H-Y, Zhang N, Guo L, Liu B-S, Jiang S-G. Toll-Like Receptor 5 of Golden Pompano Trachinotus ovatus (Linnaeus 1758): Characterization, Promoter Activity and Functional Analysis. International Journal of Molecular Sciences. 2020; 21(16):5916. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165916
Chicago/Turabian StyleZhu, Ke-Cheng, Meng Wu, Dian-Chang Zhang, Hua-Yang Guo, Nan Zhang, Liang Guo, Bao-Suo Liu, and Shi-Gui Jiang. 2020. "Toll-Like Receptor 5 of Golden Pompano Trachinotus ovatus (Linnaeus 1758): Characterization, Promoter Activity and Functional Analysis" International Journal of Molecular Sciences 21, no. 16: 5916. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165916