PD-L1 in Systemic Immunity: Unraveling Its Contribution to PD-1/PD-L1 Blockade Immunotherapy

 , , ,
, , ,
Abstract
:1. Introduction
2. PD-L1 as a Tissue Biomarker. Problems and Limitations
3. Systemic Biomarkers and PD-L1/PD-1 as Potential Systemic Biomarkers
3.1. Soluble Serum Biomarkers
3.2. PD-L1 Expression on Circulating Tumor Cells
3.3. PD-L1 Expression on Systemic Myeloid Populations
3.3.1. Monocytes
3.3.2. Macrophages
3.3.3. Dendritic Cells
3.3.4. Myeloid-Derived Suppressor Cells (MDSCs)
3.3.5. Granulocytes/Neutrophils
3.4. PD-L1 Expression on Systemic Lymphoid Populations
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PD-1 | Programmed death 1 |
| PD-L1 | Programmed death ligand 1 |
| ICI | Immune checkpoint inhibition |
| FDA | Food and drug administration |
| PFS | progression-free survival |
| OS | overall survival |
| NSCLC | non-small cell lung cancer |
| ORR | overall response rates |
| irAEs | immune related adverse events |
| IHC | immunohistochemistry |
| APC | antigen presenting cells |
| TME | tumor microenvironment |
| CTC | circulating tumor cells |
| CTLA4 | cytotoxic T-lymphocyte-associated protein 4 |
| DC | Dendritic cells |
| IFNγ | interferon gamma |
| IL6 | interleukin 6 |
| sPD-L1 | soluble PD-L1 |
| NCM | nonclassical monocytes |
| IM | intermediate monocytes |
| CM | classical monocytes |
| CCL2 | C-C Motif Chemokine Ligand 2 |
| NK | natural killer cells |
| IL10 | interleukin 10 |
| CX3CR1 | CX3C chemokine receptor 1 |
| GM-CSF | granulocyte monocyte colony stimulating factor |
| M-CSF | macrophage colony stimulating factor |
| IL-4 | interleukin 4 |
| IL-13 | interleukin 13 |
| TAM | tumor-associated macrophages |
| MAM | metastasis associated macrophages |
| ROS | reactive oxygen species |
| PD-L2 | Programmed cell death ligand 2 |
| Treg | regulatory T cells |
| CIKs | cytokine-induced killer cells |
| MHC | Major histocompatibility complex |
| MDSCs | myeloid derived suppressor cells |
| iNOS | inducible nitric oxide synthase |
| mMDSCs | monocytic MDSCs |
| gMDSCs | granulocytic MDSCs |
| PMN | polymorphonuclear cells |
| HLA-DR | human leucocyte antigen DR |
| CRC | colorectal cancer |
| NLR | neutrophil-to-lymphocyte ratio |
| TIL | tumor infiltrating lymphocytes |
| sPD-L1 | soluble PD-L1 |
References
- Brahmer, J.; Reckamp, K.L.; Baas, P.; Crino, L.; Eberhardt, W.E.; Poddubskaya, E.; Antonia, S.; Pluzanski, A.; Vokes, E.E.; Holgado, E.; et al. Nivolumab versus Docetaxel in Advanced Squamous-Cell Non–Small-Cell Lung Cancer. N. Engl. J. Med. 2015, 373, 123–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borghaei, H.; Paz-Ares, L.; Horn, L.; Spigel, D.; Steins, M.; Ready, N.; Chow, L.; Vokes, E.; Felip, E.; Holgado, E.; et al. Nivolumab versus Docetaxel in Advanced Nonsquamous Non–Small-Cell Lung Cancer. N. Engl. J. Med. 2015, 373, 1627–1639. [Google Scholar] [CrossRef] [PubMed]
- Herbst, R.S.; Baas, P.; Kim, D.-W.; Felip, E.; Perez-Gracia, J.L.; Han, J.; Molina, J.; Kim, J.; Arvis, C.; Ahn, M.; et al. Pembrolizumab versus docetaxel for previously treated, PD-L1-positive, advanced non-small-cell lung cancer (KEYNOTE-010): A randomised controlled trial. Lancet 2016, 387, 1540–1550. [Google Scholar] [CrossRef]
- Rittmeyer, A.; Barlesi, F.; Waterkamp, D.; Park, K.; Ciardiello, F.; von Pawel, J.; Gadgeel, S.; Hida, T.; Kowalski, D.; Cobo Dols, M.; et al. Atezolizumab versus docetaxel in patients with previously treated non-small-cell lung cancer (OAK): A phase 3, open-label, multicentre randomised controlled trial. Lancet 2017, 389, 255–265. [Google Scholar] [CrossRef]
- Reck, M.; Rodríguez-Abreu, D.; Robinson, A.G.; Hui, R.; Csoszi, T.; Fulop, A.; Gottfried, M.; Peled, N.; Tafreshi, A.; Cuffe, S.; et al. Pembrolizumab versus Chemotherapy for PD-L1–Positive Non–Small-Cell Lung Cancer. N. Engl. J. Med. 2016, 375, 1823–1833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langer, C.; Gadgeel, S.; Borghaei, H.; Papadimitrakopoulou, V.; Patnaik, A.; Powell, S.; Gentzler, R.; Martins, R.; Stevenson, J.; Jalal, S.; et al. Carboplatin and pemetrexed with or without pembrolizumab for advanced, non-squamous non-small-cell lung cancer: A randomised, phase 2 cohort of the open-label KEYNOTE-021 study. Lancet Oncol. 2016, 17, 1497–1508. [Google Scholar] [CrossRef]
- Socinski, M.A.; Jotte, R.M.; Cappuzzo, F.; Orlandi, F.; Stroyakovskiy, D.; Nogami, N.; Rodriguez-Abreu, D.; Moro-Sibilot, D.; Thomas, C.; Barlesi, F.; et al. Atezolizumab for First-Line Treatment of Metastatic Nonsquamous NSCLC. N. Engl. J. Med. 2018, 378, 2288–2301. [Google Scholar] [CrossRef]
- Gandhi, L.; Rodríguez-Abreu, D.; Gadgeel, S.; Esteban, E.; Felip, E.; De Angelis, F.; Domine, M.; Clingan, P.; Hochmair, M.; Powell, S.; et al. Pembrolizumab plus Chemotherapy in Metastatic Non–Small-Cell Lung Cancer. N. Engl. J. Med. 2018, 378, 2078–2092. [Google Scholar] [CrossRef]
- Paz-Ares, L.; Luft, A.; Vicente, D.; Tafreshi, A.; Gumus, M.; Mazieres, J.; Hermes, B.; Senler, F.; Csoszi, T.; Fulop, A.; et al. Pembrolizumab plus Chemotherapy for Squamous Non–Small-Cell Lung Cancer. N. Engl. J. Med. 2018, 379, 2040–2051. [Google Scholar] [CrossRef]
- Jotte, R.M.; Cappuzzo, F.; Vynnychenko, I.; Stroyakovskiy, D.; Rodriguez-Abreu, D.; Hussein, M.; Soo, R.; Conter, H.; Kozuki, T.; Silva, C.; et al. IMpower131: Primary PFS and safety analysis of a randomized phase III study of atezolizumab + carboplatin + paclitaxel or nab-paclitaxel vs. carboplatin + nab-paclitaxel as 1L therapy in advanced squamous NSCLC. J. Clin. Oncol. 2018, 36 (Suppl. 18), LBA9000. [Google Scholar] [CrossRef]
- Hellmann, M.D.; Rizvi, N.A.; Goldman, J.W.; Gettinger, S.; Borghaei, S.; Brahmer, J.; Ready, N.; Gerber, D.; Chow, L.; Juergens, R.; et al. Nivolumab plus ipilimumab as first-line treatment for advanced non-small-cell lung cancer (CheckMate 012): Results of an open-label, phase 1, multicohort study. Lancet Oncol. 2017, 18, 31–41. [Google Scholar] [CrossRef] [Green Version]
- Remon, J.; Mezquita, L.; Corral, J.; Vilariño, N.; Reguart, N. Immune-related adverse events with immune checkpoint inhibitors in thoracic malignancies: Focusing on non-small cell lung cancer patients. J. Thorac. Dis. 2018, 10 (Suppl. 13), S1516–S1533. [Google Scholar] [CrossRef] [PubMed]
- Passat, T.; Touchefeu, Y.; Gervois, N.; Jarry, A.; Bossard, C.; Bennouna, J. Physiopathological mechanisms of immune-related adverse events induced by anti-CTLA-4, anti-PD-1 and anti-PD-L1 antibodies in cancer treatment. Bull. Cancer 2018, 105, 1033–1041. [Google Scholar] [CrossRef] [PubMed]
- Lindeman, N.I.; Cagle, P.T.; Aisner, D.L.; Arcila, M.; Beasley, M.; Bernicker, E.; Colasacco, C.; Dacic, S.; Hirsch, F.; Kerr, K.; et al. Updated molecular testing guideline for the selection of lung cancer patients for treatment with targeted tyrosine kinase inhibitors. Arch. Pathol. Lab. Med. 2018, 142, 321–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaderbhaï, C.; Tharin, Z.; Ghiringhelli, F. The role of molecular profiling to predict the response to immune checkpoint inhibitors in lung cancer. Cancers 2019, 11, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofman, P.; Heeke, S.; Alix-Panabières, C.; Pantel, K. Liquid biopsy in the era of immuno-oncology: Is it ready for prime-time use for cancer patients? Ann. Oncol. 2019, 30, 1448–1459. [Google Scholar] [CrossRef] [Green Version]
- Evans, M.; O’Sullivan, B.; Smith, M.; Taniere, P. Predictive markers for anti-PD-1/PD-L1 therapy in non-small cell lung cancer-where are we? Transl. Lung Cancer Res. 2018, 7, 682–690. [Google Scholar] [CrossRef]
- Mathai, R.A.; Vidya, R.V.S.; Reddy, B.S.; Thomas, L.; Udupa, K.; Kolesar, J.; Rao, M. Potential Utility of Liquid Biopsy as a Diagnostic and Prognostic Tool for the Assessment of Solid Tumors: Implications in the Precision Oncology. J. Clin. Med. 2019, 8, 373. [Google Scholar] [CrossRef] [Green Version]
- Pasini, L.; Ulivi, P. Liquid Biopsy for the Detection of Resistance Mechanisms in NSCLC: Comparison of Different Blood Biomarkers. J. Clin. Med. 2019, 8, 998. [Google Scholar] [CrossRef] [Green Version]
- Lantuejoul, S.; Damotte, D.; Hofman, V.; Adam, J. Programmed death ligand 1 immunohistochemistry in non-small cell lung carcinoma. J. Thorac. Dis. 2019, 11 (Suppl. 1), S89–S101. [Google Scholar] [CrossRef]
- Bocanegra, A.; Fernandez, G.; Zuazo, M.; Arasanz, H.; Garcia-Granda, M.J.; Hernandez, C.; Ibañez, M.; Hernandez-Marin, B.; Martinez-Aguillo, M.; Lecumberri, M.J.; et al. PD-L1 expression in systemic immune cell populations as a potential predictive biomarker of responses to PD-L1/PD-1 blockade therapy in lung cancer. Int. J. Mol. Sci. 2019, 20, 1631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Chung, J.H. PD-L1 Testing in Non-small Cell Lung Cancer: Past, present, and future. J. Pathol. Transl. Med. 2019, 53, 199–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mcallister, S.S.; Weinberg, R.A. The tumour-induced systemic environment as a critical regulator of cancer progression and metastasis. Nat. Cell Biol. 2014, 16, 717–727. [Google Scholar] [CrossRef] [PubMed]
- Spitzer, M.H.; Carmi, Y.; Reticker-Flynn, N.E.; Kwek, S.; Madhireddy, D.; Martins, M.; Gherardini, F.; Prestwood, T.; Chabon, J.; Bendall, S.; et al. Systemic Immunity Is Required for Effective Cancer Immunotherapy. Cell 2017, 168, 487–502. [Google Scholar] [CrossRef] [PubMed]
- Laplane, L.; Duluc, D.; Bikfalvi, A.; Larmonier, N.; Pradeu, T. Beyond the tumour microenvironment. Int. J. Cancer 2019, 145, 2611–2618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okuma, Y.; Wakui, H.; Utsumi, H.; Sagawa, Y.; Hosomi, Y.; Kuwano, K.; Homma, S. Soluble Programmed Cell Death Ligand 1 as a Novel Biomarker for Nivolumab Therapy for Non–Small-cell Lung Cancer. Clin. Lung Cancer 2018, 19, 410–417. [Google Scholar] [CrossRef]
- Meyo, M.T.; Jouinot, A.; Giroux-Leprieur, E.; Fabre, E.; Wislez, M.; Alifano, M.; Leroy, K.; Boudou-Rouquette, P.; Tlemsani, C.; Khoudour, N.; et al. Predictive value of soluble PD-1, PD-L1, VEGFA, CD40 ligand and CD44 for nivolumab therapy in advanced non-small cell lung cancer: A case-control study. Cancers 2020, 12, 473. [Google Scholar] [CrossRef] [Green Version]
- Costantini, A.; Julié, C.; Dumenil, C.; Helias-Rodzewicz, Z.; Tisserand, J.; Dumoulin, J.; Giraud, V.; Labrune, S.; Chinet, T.; Emile, J.; et al. Predictive role of plasmatic biomarkers in advanced non-small cell lung cancer treated by nivolumab. Oncoimmunology 2018, 7, e1452581. [Google Scholar] [CrossRef] [Green Version]
- Ando, K.; Hamada, K.; Watanabe, M.; Ohkuma, R.; Shida, M.; Onoue, R.; Kubota, Y.; Matsui, H.; Ishiguro, T.; Hirasawa, Y.; et al. Plasma levels of soluble PD-L1 correlate with tumor regression in patients with lung and gastric cancer treated with immune checkpoint inhibitors. Anticancer Res. 2019, 39, 5195–5201. [Google Scholar] [CrossRef]
- Chiarucci, C.; Cannito, S.; Daffinà, M.G.; Amato, G.; Giacobini, G.; Cutaia, O.; Lofiego, M.; Fazio, C.; Giannarelli, D.; Danielli, R.; et al. Circulating levels of PD-L1 in mesothelioma patients from the NIBIT-MESO-1 study: Correlation with survival. Cancers 2020, 12, 361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Mahoney, K.M.; Giobbie-Hurder, A.; Zhao, F.; Lee, S.; Liao, X.; Rodig, S.; Li, J.; Wu, X.; Butterfield, L.; et al. Soluble PD-L1 as a biomarker in malignant melanoma treated with checkpoint blockade. Cancer Immunol. Res. 2017, 5, 480–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guibert, N.; Delaunay, M.; Lusque, A.; Boubekeur, N.; Rouquette, I.; Clermont, E.; Mourlanette, J.; Gouin, S.; Dormoy, I.; Favre, G.; et al. PD-L1 expression in circulating tumor cells of advanced non-small cell lung cancer patients treated with nivolumab. Lung Cancer. 2018, 120, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Nicolazzo, C.; Raimondi, C.; Mancini, M.; Caponnetto, S.; Gradilone, A.; Gandini, O.; Mastromartino, M.; Del Bene, G.; Prete, A.; Longo, F.; et al. Monitoring PD-L1 positive circulating tumor cells in non-small cell lung cancer patients treated with the PD-1 inhibitor Nivolumab. Sci. Rep. 2016, 6, 31726. [Google Scholar] [CrossRef] [PubMed]
- Jacquelot, N.; Roberti, M.P.; Enot, D.P.; Rusakiewicz, S.; Ternes, N.; Jegou, S.; Woods, D.; Sodre, A.; Hansen, M.; Meirow, Y.; et al. Predictors of responses to immune checkpoint blockade in advanced melanoma. Nat. Commun. 2017, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Lang, J. Soluble PD-1 and PD-L1: Predictive and prognostic significance in cancer. Oncotarget 2017, 8, 97671–97682. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, H.; Chen, H.; Wang, W.; Chen, X.; Geng, Q.; Xia, Z.; Lu, Y. Serum levels of soluble programmed death ligand 1 predict treatment response and progression free survival in multiple myeloma. Oncotarget 2015, 6, 41228–41236. [Google Scholar] [CrossRef] [Green Version]
- Rossille, D.; Gressier, M.; Damotte, D.; Maucort-Bouch, D.; Pangault, C.; Semana, G.; Gouill, S.; Haioun, C.; Tarte, K.; Lamy, T.; et al. High level of soluble programmed cell death ligand 1 in blood impacts overall survival in aggressive diffuse large B-cell lymphoma: Results from a French multicenter clinical trial. Leukemia 2014, 28, 2367–2375. [Google Scholar] [CrossRef]
- Liu, S.; Zhu, Y.; Zhang, C.; Meng, X.; Sun, B.; Zhang, G.; Fan, Y.; Kang, X. The Clinical Significance of Soluble Programmed Cell Death-Ligand 1 (sPD-L1) in Patients With Gliomas. Front. Oncol. 2020, 10, 9. [Google Scholar] [CrossRef]
- Lu, T.; Chen, Y.; Li, J.; Guo, Q.; Lin, W.; Zheng, Y.; Su, Y.; Zong, J.; Lin, S.; Ye, Y.; et al. High soluble programmed death-ligand 1 predicts poor prognosis in patients with nasopharyngeal carcinoma. OncoTargets Ther. 2020, 13, 1757–1765. [Google Scholar] [CrossRef] [Green Version]
- Frigola, X.; Inman, B.A.; Lohse, C.M.; Krco, K.; Cheville, J.; Thompson, R.; Leibovich, B.; Blute, M.; Dong, H.; Kwon, E. Identification of a soluble form of B7-H1 that retains immunosuppressive activity and is associated with aggressive renal cell carcinoma. Clin. Cancer Res. 2011, 17, 1915–1923. [Google Scholar] [CrossRef] [Green Version]
- Jovanovic, D.; Milenkovic, M.R.; Stevuljevic, J.K.; Markovic, J.; Ceriman, V.; Kontic, M.; Trifunovic, V. Membrane PD-L1 expression and soluble PD-L1 plasma levels in idiopathic pulmonary fibrosis-a pilot study. J. Thorac. Dis. 2018, 10, 6660–6669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalmarzi, R.N.; Fattahi, N.; Kaviani, Z.; Ataee, P.; Mansouri, M.; Moradi, G.; Yousefzade, A.; Abbassi, J. Inverse correlation of soluble programmed cell death-1 ligand-1 (sPD-L1) with eosinophil count and clinical severity in allergic rhinitis patients. Allergol. Int. 2017, 66, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Aarslev, K.; Dige, A.; Greisen, S.R.; Kreutzfeldt, M.; Jessen, N.; Vilstrup, H.; Deleuran, B.; Gronbaek, H. Soluble programmed death-1 levels are associated with disease activity and treatment response in patients with autoimmune hepatitis. Scand. J. Gastroenterol. 2017, 52, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Yamagiwa, S.; Ishikawa, T.; Waguri, N.; Sugitani, S.; Kamimura, K.; Tsuchiya, A.; Takamura, M.; Kawai, H.; Terai, S. Increase of soluble programmed cell death ligand 1 in patients with chronic hepatitis C. Int. J. Med. Sci. 2017, 14, 403–411. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Guo, H.; Li, S.; Liu, C.; Ding, S.; Huang, Y.; Fang, C.; Hu, J. Soluble programmed death-1 ligand 1(sPD-L1) is significantly reduced in the serum of type 1 diabetes patients. Acta Diabetol. 2018, 55, 515–517. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, Q.; Shi, B.; Xu, P.; Hu, Z.; Bai, L.; Zhang, X. Development of a sandwich ELISA for evaluating soluble PD-L1 (CD274) in human sera of different ages as well as supernatants of PD-L1 + cell lines. Cytokine 2011, 56, 231–238. [Google Scholar] [CrossRef]
- Okuyama, M.; Mezawa, H.; Kawai, T.; Urashima, M. Elevated Soluble PD-L1 in Pregnant Women’s Serum Suppresses the Immune Reaction. Front. Immunol. 2019, 10, 86. [Google Scholar] [CrossRef] [Green Version]
- Giuliano, M.; Shaikh, A.; Lo, H.C.; Arpino, G.; De Placido, S.; Zhang, X.; Cristofanilli, M.; Schiff, R.; Trivedi, M. Perspective on circulating tumor cell clusters: Why it takes a village to metastasize. Cancer Res. 2018, 78, 845–852. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Balic, M.; El-Ashry, D.; Cote, R.J. Circulating Tumor Cells: Strategies for Capture, Analyses, and Propagation. Cancer J. 2018, 24, 70–77. [Google Scholar] [CrossRef]
- Hong, Y.; Fang, F.; Zhang, Q. Circulating tumor cell clusters: What we know and what we expect. Int. J. Oncol. 2016, 49, 2206–2216. [Google Scholar] [CrossRef] [Green Version]
- Bankó, P.; Lee, S.-Y.; Nagygyörgy, V.; Zrinyi, M.; Chae, C.; Cho, D.; Telekes, A. Technologies for circulating tumor cell separation from whole blood. J. Hematol Oncol. 2019, 12, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aceto, N.; Bardia, A.; Miyamoto, D.T.; Donaldson, M.; Wittner, B.; Spencer, J.; Yu, M.; Pely, A.; Engstrom, A.; Zhu, H.; et al. Circulating Tumor Cell Clusters are Oligoclonal Precursors of Breast Cancer Metastasis. Cell 2014, 158, 1110–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, Y.; Yagi, S.; Akamatsu, H.; Kanai, K.; Hayata, A.; Tokudome, N.; Akamatsu, K.; Higuchi, M.; Kanbara, H.; Nakanishi, M.; et al. Heterogeneous Expression of Programmed Death Receptor-ligand 1 on Circulating Tumor Cells in Patients With Lung Cancer. Clin. Lung Cancer. 2019, 20, 270–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilie, M.; Szafer-Glusman, E.; Hofman, V.; Chamorey, E.; Lalvee, S.; Selva, E.; Leroy, S.; Marquette, C.; Kowanetz, M.; Hedge, P.; et al. Detection of PD-L1 in circulating tumor cells and white blood cells from patients with advanced non-small-cell lung cancer. Ann. Oncol. 2018, 29, 193–199. [Google Scholar] [CrossRef]
- Tang, H.; Liang, Y.; Anders, R.A.; Taube, J.; Qiu, X.; Mulgaonkar, A.; Liu, X.; Harrington, S.; Guo, J.; Xin, Y.; et al. PD-L1 on host cells is essential for PD-L1 blockade–mediated tumor regression. J. Clin. Investig. 2018, 128, 580–588. [Google Scholar] [CrossRef] [Green Version]
- Haderk, F.; Schulz, R.; Iskar, M.; Cid, L.; Worst, T.; Willmund, K.; Schulz, A.; Warnken, U.; Seiler, J.; Benner, A.; et al. Tumor-derived exosomes modulate PD-L1 expression in monocytes. Sci. Immunol. 2017, 2, eaah5509. [Google Scholar] [CrossRef] [Green Version]
- Bianchini, M.; Duchene, J.; Santovito, D.; Schloss, M.; Evrard, M.; Winkels, H.; Aslani, M.; Mohanta, S.; Horckmans, M.; Blanchet, X.; et al. PD-L1 expression on nonclassical monocytes reveals their origin and immunoregulatory function. Sci. Immunol. 2019, 4, eaar3054. [Google Scholar] [CrossRef]
- Vari, F.; Arpon, D.; Keane, C.; Hertzberg, M.; Talaulikar, D.; Jain, S.; Cui, Q.; Han, E.; Tobin, J.; Bird, R.; et al. Immune evasion via PD-1/PD-L1 on NK cells and monocyte/macrophages is more prominent in Hodgkin lymphoma than DLBCL. Blood 2018, 131, 1809–1819. [Google Scholar] [CrossRef] [Green Version]
- Kitamura, T.; Doughty-Shenton, D.; Cassetta, L.; Fragkogianni, S.; Brownlie, D.; Kato, Y.; Carragher, N.; Pollard, J. Monocytes differentiate to immune suppressive precursors of metastasis-associated macrophages in mouse models of metastatic breast cancer. Front. Immunol. 2018, 8, 2004. [Google Scholar] [CrossRef] [Green Version]
- Sugiura, D.; Maruhashi, T.; Okazaki, I.-M.; Shimizu, K.; Maeda, T.; Takemoto, T.; Okazaki, T. Restriction of PD-1 function by cis-PD-L1/CD80 interactions is required for optimal T cell responses. Science 2019, 364, 558–566. [Google Scholar] [CrossRef]
- Mayoux, M.; Roller, A.; Pulko, V.; Sammicheli, S.; Chen, S.; Sum, E.; Jost, C.; Fransen, M.; Buser, R.; Kowanetz, M.; et al. Dendritic cells dictate responses to PD-L1 blockade cancer immunotherapy. Sci. Transl. Med. 2020, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.; Song, Y.; Richardson, P.; Munshi, N.; Chauhan, D.; Anderson, K.C. Targeting PD-1-PD-L1 immune checkpoint in plasmacytoid dendritic cell interactions with T cells, natural killer cells and multiple myeloma cells. Leukemia 2015, 29, 1441–1444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Bakthavatsalam, R.; Meng, Z.; Li, Z.; Li, W.; Perkins, J.; Reyes, J. PD-L1 signal on liver dendritic cells is critical for Foxp3 +CD4+CD25+ treg and liver tolerance induction in mice. Transpl. Proc. 2013, 45, 1853–1855. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Song, Z.; Xiao, J.; Liu, X.; Luo, Y.; Yang, Z.; Lou, R.; Li, A. Blocking the PD-1/PD-L1 axis in dendritic cell-stimulated Cytokine-Induced killer cells with pembrolizumab enhances their therapeutic effects against hepatocellular carcinoma. J. Cancer 2019, 10, 2578–2587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, J.S.; Gibney, G.; Kudchadkar, R.R.; Yu, B.; Cheng, P.; Martinez, A.; Kroeger, J.; Richards, A.; McCormick, L.; Moberg, V.; et al. Phase I/II study of metastatic melanoma patients treated with nivolumab who had progressed after ipilimumab. Cancer Immunol. Res. 2016, 4, 345–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, C.; Redd, P.S.; Lee, J.R.; Savage, N.; Liu, K. The expression profiles and regulation of PD-L1 in tumor-induced myeloid-derived suppressor cells. Oncoimmunology 2016, 5, e1247135. [Google Scholar] [CrossRef] [Green Version]
- Califano, J.A.; Khan, Z.; Noonan, K.A.; Rudraraju, L.; Zhang, Z.; Wang, H.; Goodman, S.; Gourin, C.; Ha, P.; Fakhry, C.; et al. Tadalafil augments tumor specific immunity in patients with head and neck squamous cell carcinoma. Clin. Cancer Res. 2015, 21, 30–38. [Google Scholar] [CrossRef] [Green Version]
- De Kleijn, S.; Langereis, J.D.; Leentjens, J.; Kox, M.; Netea, M.; Koenderman, L.; Ferwerda, G.; Pickkers, P.; Hermans, P. IFN-γ-Stimulated Neutrophils Suppress Lymphocyte Proliferation through Expression of PD-L1. PLoS ONE 2013, 8, e72249. [Google Scholar] [CrossRef] [Green Version]
- Bharat, A.; McQuattie-Pimentel, A.C.; Budinger, G.S. Non-Classical Monocytes in Tissue Injury and Cancer. Oncotarget 2017, 8, 1061671–1061672. [Google Scholar] [CrossRef]
- Stankovic, B.; Bjørhovde, H.A.K.; Skarshaug, R.; Aamodt, H.; Frafjord, A.; Muller, E.; Hammarstrom, C.; Beraki, K.; Baekkevold, E.; Woldbaek, P.; et al. Immune Cell Composition in Human Non-small Cell Lung Cancer. Front. Immunol. 2019, 9, 3101. [Google Scholar] [CrossRef] [Green Version]
- Hanna, R.N.; Cekic, C.; Sag, D.; Tacke, R.; Thomas, G.; Nowyhed, H.; Herrley, E.; Rasquinha, N.; McArdle, S.; Wu, R.; et al. Patrolling monocytes control tumor metastasis to the lung. Science 2015, 350, 985–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyakudomi, M.; Matsubara, T.; Hyakudomi, R.; Yamamoto, T.; Kinugasa, S.; Yamanoi, A.; Maruyama, R.; Tanaka, T. Increased expression of Fractalkine is correlated with a better prognosis and an increased number of both CD8+ T cells and natural killer cells in gastric adenocarcinoma. Ann. Surg Oncol. 2008, 15, 1775–1782. [Google Scholar] [CrossRef] [PubMed]
- Park, M.H.; Lee, J.S.; Yoon, J.H. High expression of CX3CL1 by tumor cells correlates with a good prognosis and increased tumor-infiltrating CD8+ T cells, natural killer cells, and dendritic cells in breast carcinoma. J. Surg. Oncol. 2012, 106, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.; Heishi, T.; Khan, O.F.; Kowalski, P.; Incio, J.; Rahbari, N.; Chung, E.; Clark, J.; Willett, C.; Luster, A.; et al. Ly6Clo monocytes drive immunosuppression and confer resistance to anti-VEGFR2 cancer therapy. J. Clin. Investig. 2017, 127, 3039–3051. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Zhou, Q.; Zhu, H.; Jing, Y.; Zhang, X.; Yang, Y.; Hu, Q.; Huang, X.; Ni, Y. Frequency of circulating CD14 ++ CD16 + intermediate monocytes as potential biomarker for the diagnosis of oral squamous cell carcinoma. J. Oral Pathol. Med. 2018, 47, 923–929. [Google Scholar] [CrossRef]
- Subimerb, C.; Pinlaor, S.; Lulitanond, V.; Khuntikeo, N.; Okada, S.; McGrath, M.; Wongkham, S. Circulating CD14+CD16+monocyte levels predict tissue invasive character of cholangiocarcinoma. Clin. Exp. Immunol. 2010, 161, 471–479. [Google Scholar] [CrossRef]
- Romano, E.; Kusio-Kobialka, M.; Foukas, P.G.; Baumgaertner, P.; Meyer, C.; Ballabeni, P.; Michielin, O.; Weide, B.; Romero, P.; Speiser, D. Ipilimumab-dependent cell-mediated cytotoxicity of regulatory T cells ex vivo by nonclassical monocytes in melanoma patients. Proc. Natl. Acad. Sci. USA 2015, 112, 6140–6145. [Google Scholar] [CrossRef] [Green Version]
- Krieg, C.; Nowicka, M.; Guglietta, S.; Schindler, S.; Hartmann, F.; Weber, L.; Dummer, R.; Robinson, M.; Levesque, M.; Becher, B. High-dimensional single-cell analysis predicts response to anti-PD-1 immunotherapy. Nat. Med. 2018, 24, 144–153. [Google Scholar] [CrossRef]
- Zhou, J.; Tang, Z.; Gao, S.; Li, C.; Feng, Y.; Zhou, X. Tumor-Associated Macrophages: Recent Insights and Therapies. Front. Oncol. 2020, 10, 188. [Google Scholar] [CrossRef]
- Gordon, S.R.; Maute, R.L.; Dulken, B.W.; Hutter, G.; George, B.; McCracken, M.; Gupta, R.; Tsai, J.; Sinha, R.; Corey, D.; et al. PD-1 expression by tumour-associated macrophages inhibits phagocytosis and tumour immunity. Nature 2017, 545, 495–499. [Google Scholar] [CrossRef]
- Dhupkar, P.; Gordon, N.; Stewart, J.; Kleinerman, E.S. Anti-PD-1 therapy redirects macrophages from an M2 to an M1 phenotype inducing regression of OS lung metastases. Cancer Med. 2018, 7, 2654–2664. [Google Scholar] [CrossRef] [PubMed]
- Qian, B.; Li, J.; Zhang, H.; Kitamura, T.; Zhang, J.; Campion, L.; Kaiser, E.; Snyder, L.; Pollard, J. CCL2 recruits inflammatory monocytes to facilitate breast tumor metastasis. Nature 2011, 475, 222–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, W.; Li, X.; Porter, J.L.; Ostrodi, D.; Yang, B.; Li, J.; Wang, Y.; Zhang, J.; Bai, L.; Jiao, S. Level of plasmacytoid dendritic cells is increased in non-small cell lung carcinoma. Tumor. Biol. 2014, 35, 2247–2252. [Google Scholar] [CrossRef] [PubMed]
- Talmadge, J.E.; Gabrilovich, D.I. History of myeloid-derived suppressor cells. Nat. Rev. Cancer 2013, 13, 739–752. [Google Scholar] [CrossRef] [PubMed]
- Ostrand-Rosenberg, S.; Horn, L.A.; Haile, S.T. The Programmed Death-1 Immune-Suppressive Pathway: Barrier to Antitumor Immunity. J. Immunol. 2014, 193, 3835–3841. [Google Scholar] [CrossRef] [PubMed]
- Escors, D.; Talmadge, J.E.; Breckpot, K.; Van Ginderachter, J.A.; Kochan, G. Myeloid-Deriv Suppressor Cells and Cancer; Springer: Berlin, Germany, 2016. [Google Scholar] [CrossRef]
- Morales, J.K.; Kmieciak, M.; Knutson, K.L.; Bear, H.D.; Manjili, M.H. GM-CSF is one of the main breast tumor-derived soluble factors involved in the differentiation of CD11b-Gr1- bone marrow progenitor cells into myeloid-derived suppressor cells. Breast Cancer Res. Treat. 2010, 123, 39–49. [Google Scholar] [CrossRef] [Green Version]
- Bronte, V.; Brandau, S.; Chen, S.-H.; Colombo, M.; Frey, A.; Greten, T.; Mandruzzato, S.; Murray, P.; Ochoa, A.; Ostrand-Rosenberg, S.; et al. Recommendations for myeloid-derived suppressor cell nomenclature and characterization standards. Nat. Commun. 2016, 7, 12150. [Google Scholar] [CrossRef] [Green Version]
- Liechtenstein, T.; Perez-Janices, N.; Gato, M.; Caliendo, F.; Kochan, G.; Blanco-Luquin, I.; Van der Jeught, K.; Arce, F.; Guerrero-Setas, D.; Fernandez-Irigoyen, J.; et al. A highly efficient tumor-infiltrating MDSC differentiation system for discovery of anti-neoplastic targets, which circumvents the need for tumor establishment in mice. Oncotarget 2014, 5, 7843–7857. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Gebhardt, C.; Umansky, L.; Beckhove, P.; Schulze, T.; Utikal, J.; Umansky, V. Elevated chronic inflammatory factors and myeloid-derived suppressor cells indicate poor prognosis in advanced melanoma patients. Int. J. Cancer 2015, 136, 2352–2360. [Google Scholar] [CrossRef]
- Sun, H.-L.; Zhou, X.; Xue, Y.-F.; Wang, K.; Shen, Y.; Mao, J.; Guo, H.; Miao, Z. Increased frequency and clinical significance of myeloid-derived suppressor cells in human colorectal carcinoma. World J. Gastroenterol. 2012, 18, 3303–3309. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Ma, X.; Zhu, C.; Liu, L.; Wang, G.; Yuan, X. The Role of Myeloid-Derived Suppressor Cells in Patients with Solid Tumors: A Meta-Analysis. PLoS ONE 2016, 11, e0164514. [Google Scholar] [CrossRef] [PubMed]
- Sade-Feldman, M.; Kanterman, J.; Klieger, Y.; Ish-Shalom, E.; Olga, M.; Saragovi, A.; Shtainberg, H.; Lotem, M.; Baniyash, M. Clinical significance of circulating CD33+ CD11bHLA-DR myeloid cells in patients with stage IV melanoma treated with ipilimumab. Clin. Cancer Res. 2016, 22, 5661–5672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimura, T.; Takahashi, M. IFN-y-mediated survival enables human neutrophils to produce MCP-1/CCL2 in response to activation by TLR ligands. J. Immunol. 2007, 179, 1942–1949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishalian, I.; Granot, Z.; Fridlender, Z.G. The diversity of circulating neutrophils in cancer. Immunobiology 2017, 222, 82–88. [Google Scholar] [CrossRef]
- Mantovani, A.; Cassatella, M.A.; Costantini, C.; Jaillon, S. Neutrophils in the activation and regulation of innate and adaptive immunity. Nat. Rev. Immunol. 2011, 11, 519–531. [Google Scholar] [CrossRef]
- Sconocchia, G.; Zlobec, I.; Lugli, A.; Calabrese, D.; Iezzi, G.; Karamitopoulou, E.; Patsouris, E.; Peros, G.; Horcic, M.; Tornillo, L.; et al. Tumor infiltration by FcγRIII (CD16)+ myeloid cells is associated with improved survival in patients with colorectal carcinoma. Int. J. Cancer 2011, 128, 2663–2672. [Google Scholar] [CrossRef] [Green Version]
- Kargl, J.; Zhu, X.; Zhang, H.; Yang, G.; Friesen, T.; Shipley, M.; Maeda, D.; Zebala, J.; McKay-Fleisch, J.; Meredith, G.; et al. Neutrophil content predicts lymphocyte depletion and anti-PD-1 treatment failure in NSCLC. JCI Insight 2019, 4, e130850. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Li, H.; Deng, Y.; Tai, Y.; Zeng, K.; Zhang, Y.; Liu, W.; Zhang, Q.; Yang, Y. Cancer-associated fibroblasts induce PD-L1+ neutrophils through the IL6-STAT3 pathway that foster immune suppression in hepatocellular carcinoma. Cell Death Dis. 2018, 9, 422. [Google Scholar] [CrossRef]
- Castell, S.D.; Harman, M.F.; Morón, G.; Maletto, B.A.; Pistoresi-Palencia, M.C. Neutrophils Which Migrate to Lymph Nodes Modulate CD4+ T Cell Response by a PD-L1 Dependent Mechanism. Front. Immunol. 2019, 10, 105. [Google Scholar] [CrossRef]
- Faria, S.S.; Fernandes, P.C.; Silva, M.J.B.; Lima, V.; Fontes, W.; Freitas-Junior, R.; Eterovic, A.; Forget, P. The neutrophil-to-lymphocyte ratio: A narrative review. Ecancermedicalscience 2016, 10, 702. [Google Scholar] [CrossRef] [Green Version]
- Bowen, R.C.; Little, N.A.B.; Harmer, J.; Ma, J.; Mirabelli, L.; Roller, K.; Breivik, A.; Signor, E.; Miller, A.; Khong, H. Neutrophil-to-Lymphocyte Ratio as Prognostic Indicator in Gastrointestinal Cancers: A Systematic Review and Meta-Analysis. Oncotarget 2017, 8, 32171–32189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diskin, B.; Adam, S.; Cassini, M.F.; Sanchez, G.; Liria, M.; Aykut, B.; Buttar, C.; Li, E.; Sundberg, B.; Salas, R.; et al. PD-L1 engagement on T cells promotes self-tolerance and suppression of neighboring macrophages and effector T cells in cancer. Nat. Immunol. 2020, 21, 442–454. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.M.G.; Wen, T.; Dong, H. Bidirectional signals of PD-L1 in T cells that fraternize with cancer cells. Nat. Immunol. 2020, 21, 365–366. [Google Scholar] [CrossRef] [PubMed]
- Brochez, L.; Meireson, A.; Chevolet, I.; Sundahl, N.; Ost, P.; Kruse, V. Challenging PD-L1 expressing cytotoxic T cells as a predictor for response to immunotherapy in melanoma. Nat. Commun. 2018, 9, 1–3. [Google Scholar] [CrossRef] [Green Version]


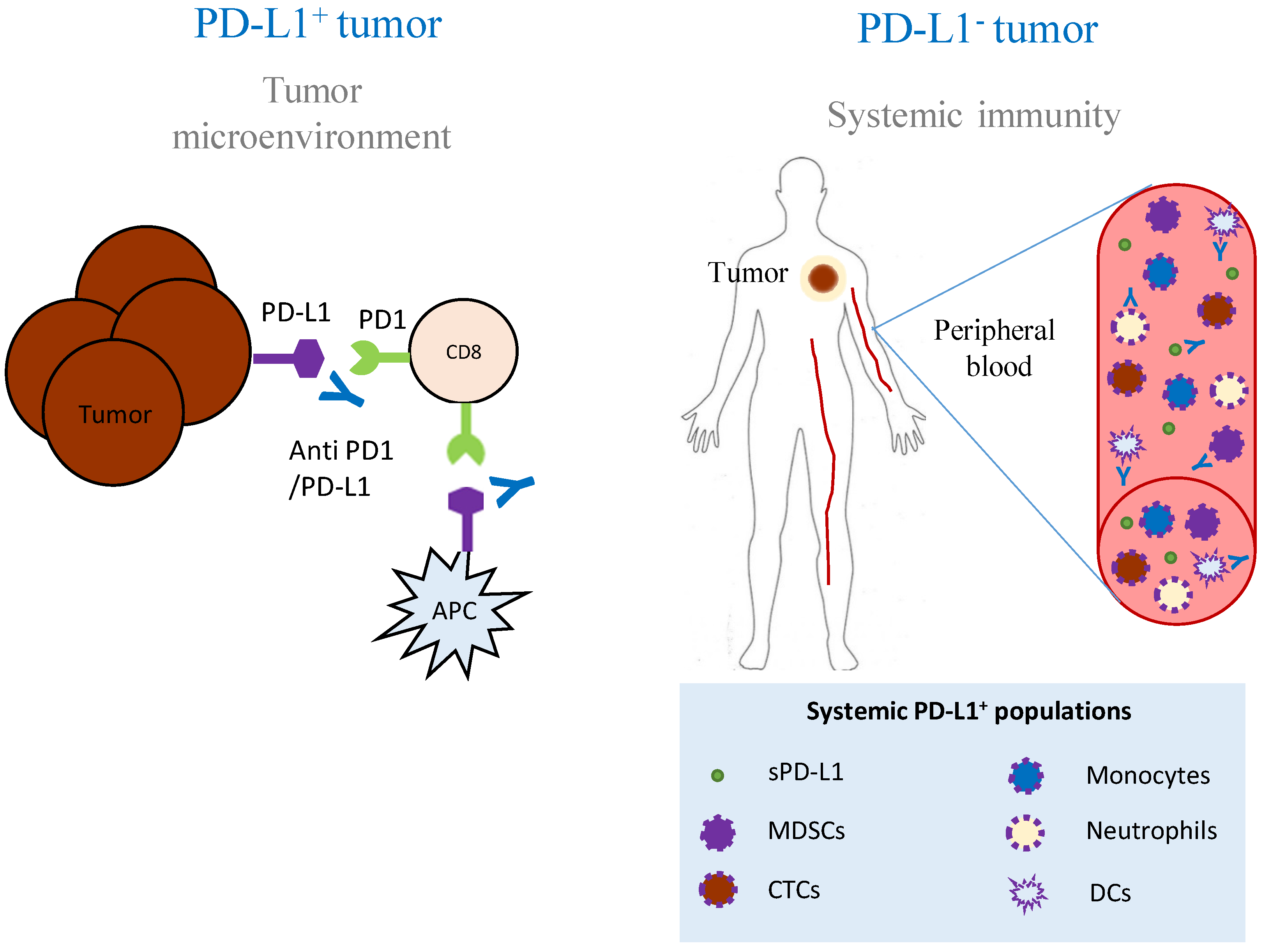{kind=link}
| Cancer Type | PD-L1 Source | Number of Patients/Treatment | Main Results | References |
|---|---|---|---|---|
| NSCLC | Soluble PD-L1 | 39 patients/nivolumab | Elevated baseline levels of sPD-L1 correlate with progressive disease | Okuma et al. [26] |
| NSCLC | Soluble PD-L1 | 51 patients/nivolumab | High baseline levels of sPD-1 and sPD-L1 render shorter PFS and OS | Meyo et al. [27] |
| NSCLC | Soluble PD-L1 | 43 patients/nivolumab | Increased sPD-L1 in non-responders at first tumor evaluation | Costantini et al. [28] |
| NSCLC/gastric cancer | Soluble PD-L1 | 21 patients/nivolumab, pembrolizumab | Reduced sPD-L1 levels after treatment correlates with tumor regression | Ando et al. [29] |
| mesothelioma | Soluble PD-L1 | 40 patients/durvalumab + tremelimumab | Low baseline levels of sPD-L1 render longer OS | Chiarucci et al. [30] |
| Melanoma | Soluble PD-L1 | 100 patients/ipilimumab, bevacizumab, pembrolizumab | Increased sPD-L1 after treatment correlates with partial responses | Zhou et al. [31] |
| NSCLC | Circulating tumor cells | 96 patients/nivolumab | High baseline CTC numbers associate with progression | Guibert et al. [32] |
| NSCLC | Circulating tumor cells | 24 patients/nivolumab | Presence of PD-L1+ CTCs both at baseline and after treatment correlates with progression and worsened outcome | Nicolazzo et al. [33] |
| NSCLC | CD11b+ cells | 32 patients/nivolumab, pembrolizumab, atezolizumab | Circulating PD-L1+ myeloid populations correlate with response to anti PD-L1/anti PD-1 treatment in NSCLC patients, independently of tumor PD-L1 expression | Bocanegra et al. [21] |
| Melanoma | Peripheral T cells | 190 patients/ipilimumab, nivolumab | PD-L1 expression on peripheral T cells is a prognostic biomarker of OS and PFS | Jacquelot et al. [34] |
| Population | Main Results | References |
|---|---|---|
| Monocytes | Monocytes acquire PD-L1+ phenotype via tumor-derived extracellular vesicles and exert pro-tumorigenic functions | Haderk et al. [56] |
| Non-classical monocytes (NCM) | PD-L1 is a marker of NCMunder inflammatory conditions and promotes T cell survival | Bianchini et al. [57] |
| Monocytes | PD-L1+ circulating monocytes promote exhaustion of PD-1high natural killer cells | Vari et al. [58] |
| Metastasis associated macrophages (MAMs) | Despite PD-L1 expression, they suppress T cell activity by a ROS-dependent but checkpoint-independent mechanism | Kitamura et al. [59] |
| Dendritic cells (DCs) | The immunotherapy-driven blockade of the cis interaction of PD-L1 with CD80 on DCs enables the interaction CD80-CD28, thus reinvigorating cytotoxic CD8 T cell responses | Sigiura et al. [60], Mayoux et al. [61] |
| Dendritic cells (DCs) | PD-L1 blockade reverses natural killer cells suppression lead by DCs | Ray et al. [62] |
| Dendritic cells (DCs) | DCs induce the expansion of Treg in a PD-L1 dependent mechanism | Liu et al. [63] |
| Dendritic cells (DCs) | PD-1 blockade induces proliferation and cytotoxic capacity of cytokine-induced killer cells co-cultured with DCs in a liver cancel model in vitro and in vivo, rendering enhanced clinical benefits | Zhang et al. [64] |
| Myeloid-derived suppressor cells (MDSCs) | High numbers of MDSCs were associated with poor survival in ipilimumab-refractory melanoma patients treated with nivolumab | Weber et al. [65] |
| Myeloid-derived suppressor cells (MDSCs) | PD-L1+ MDSCs are less frequent in peripheral blood as compared to tumor tissues. pSTAT1-IRF1 axis regulates PD-L1 expression in MDSCs | Lu et al. [66] |
| Myeloid-derived suppressor cells (MDSCs) | MDSC inhibition augments general and tumor-specific immunity in head and neck squamous cell carcinoma (HNSCC)patients | Califano et al. [67] |
| Neutrophils | IFNγ-induced expression of PD-L1 on circulating neutrophils suppress lymphocyte proliferation | De Kleijn et al. [68] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bocanegra, A.; Blanco, E.; Fernandez-Hinojal, G.; Arasanz, H.; Chocarro, L.; Zuazo, M.; Morente, P.; Vera, R.; Escors, D.; Kochan, G. PD-L1 in Systemic Immunity: Unraveling Its Contribution to PD-1/PD-L1 Blockade Immunotherapy. Int. J. Mol. Sci. 2020, 21, 5918. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165918
Bocanegra A, Blanco E, Fernandez-Hinojal G, Arasanz H, Chocarro L, Zuazo M, Morente P, Vera R, Escors D, Kochan G. PD-L1 in Systemic Immunity: Unraveling Its Contribution to PD-1/PD-L1 Blockade Immunotherapy. International Journal of Molecular Sciences. 2020; 21(16):5918. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165918
Chicago/Turabian StyleBocanegra, Ana, Ester Blanco, Gonzalo Fernandez-Hinojal, Hugo Arasanz, Luisa Chocarro, Miren Zuazo, Pilar Morente, Ruth Vera, David Escors, and Grazyna Kochan. 2020. "PD-L1 in Systemic Immunity: Unraveling Its Contribution to PD-1/PD-L1 Blockade Immunotherapy" International Journal of Molecular Sciences 21, no. 16: 5918. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165918