Characterization of the Role of SPL9 in Drought Stress Tolerance in Medicago sativa



Abstract
:1. Introduction
2. Results
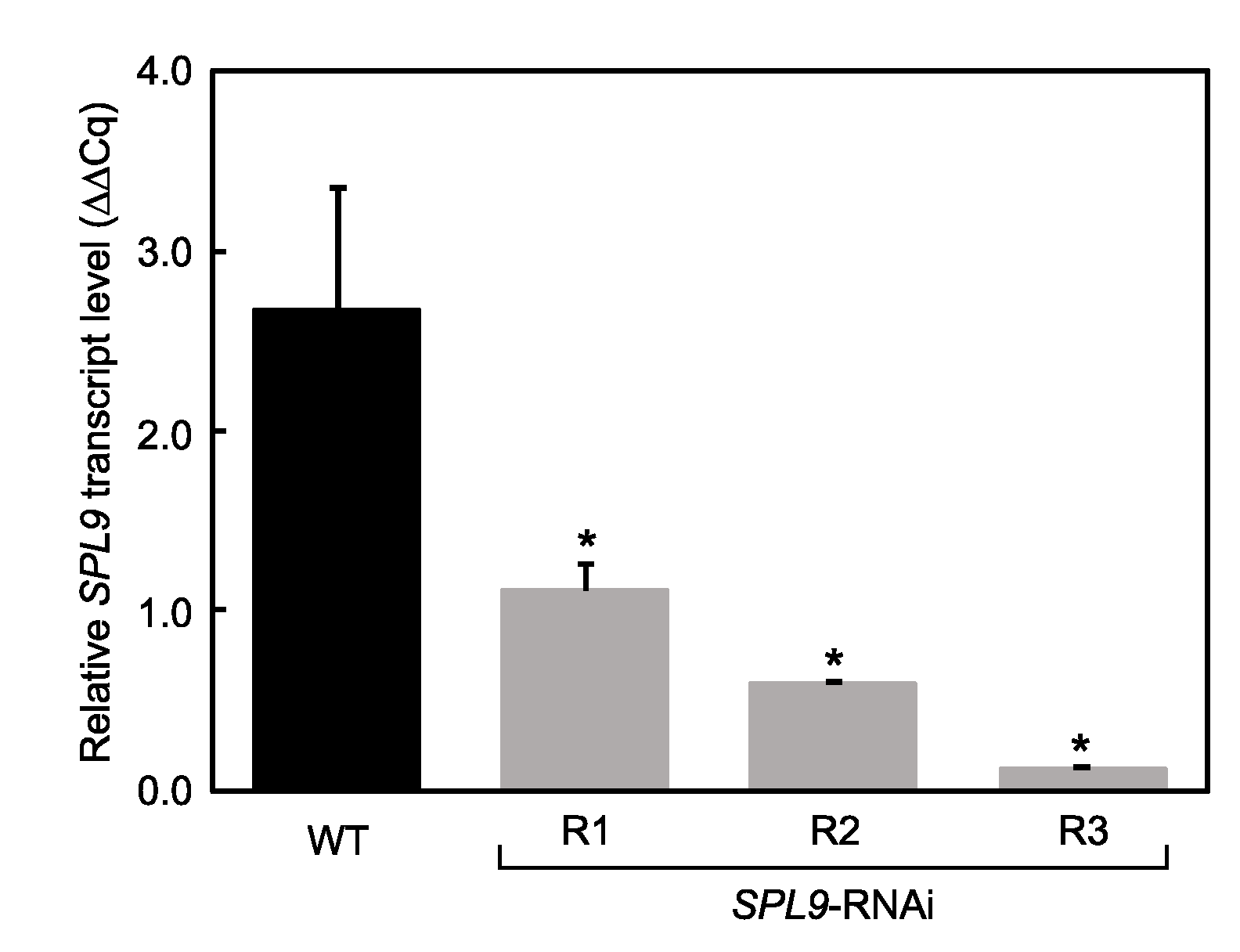
2.1. Phenotypic Characterization of SPL9-RNAi Plants
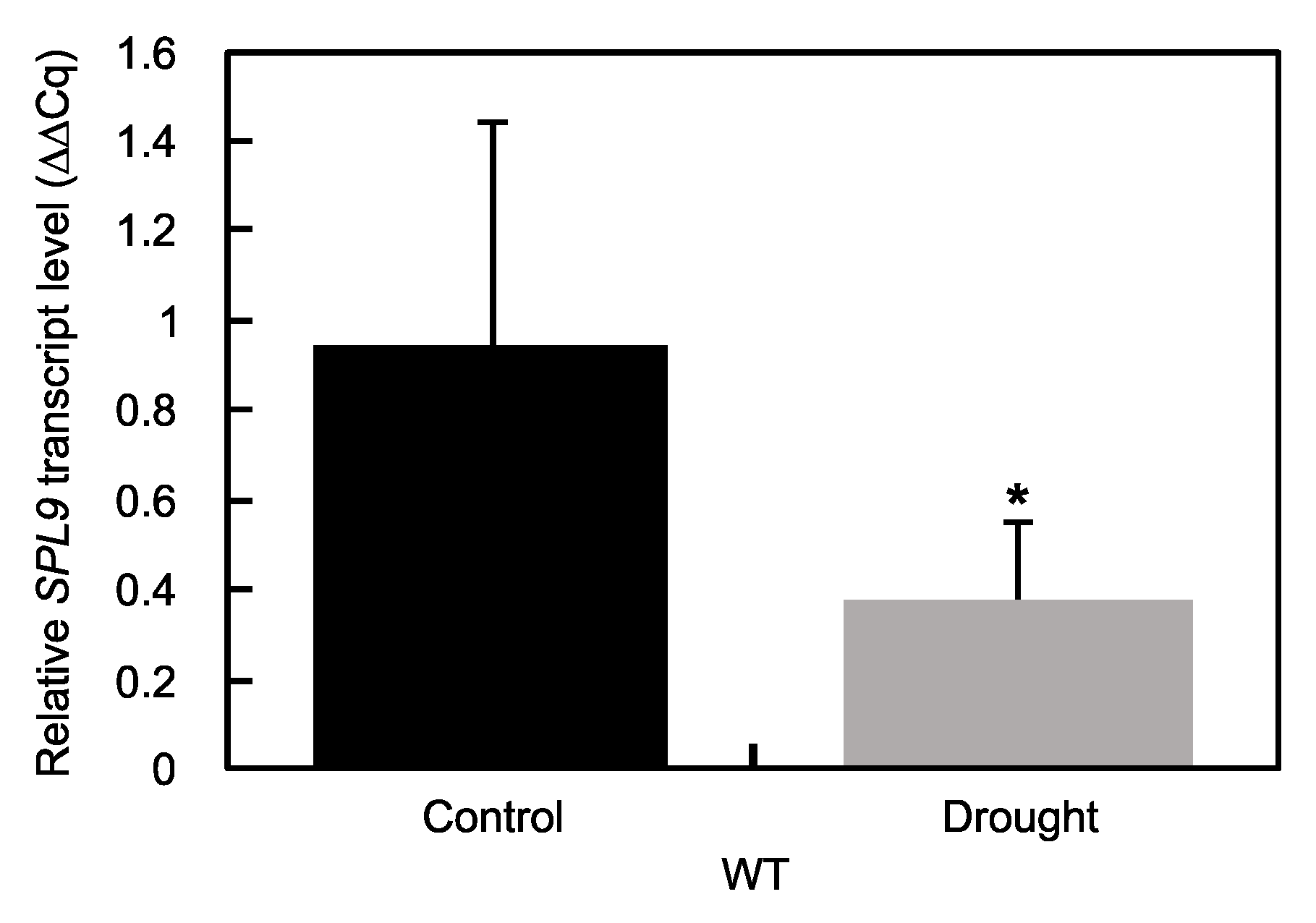
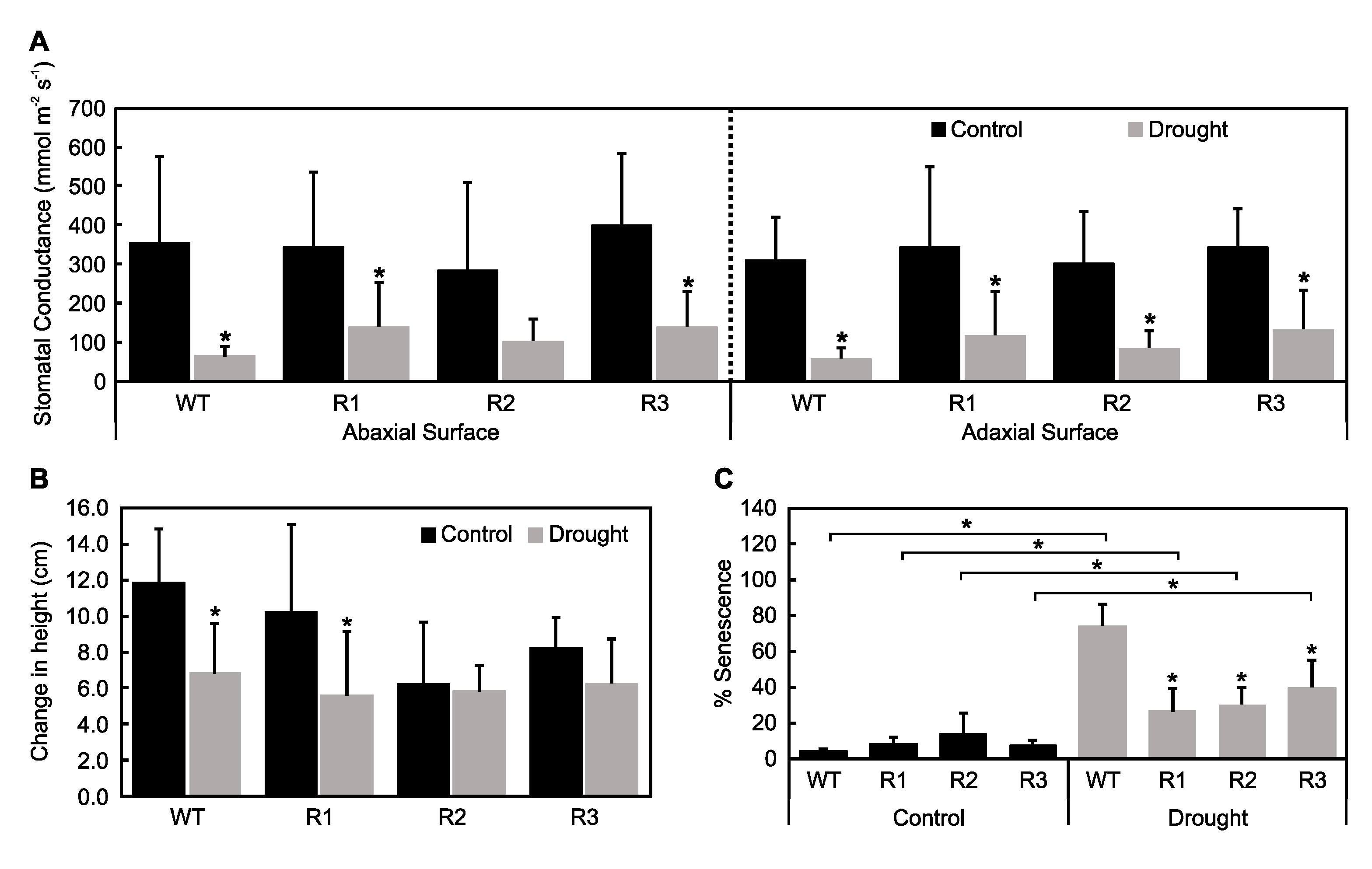
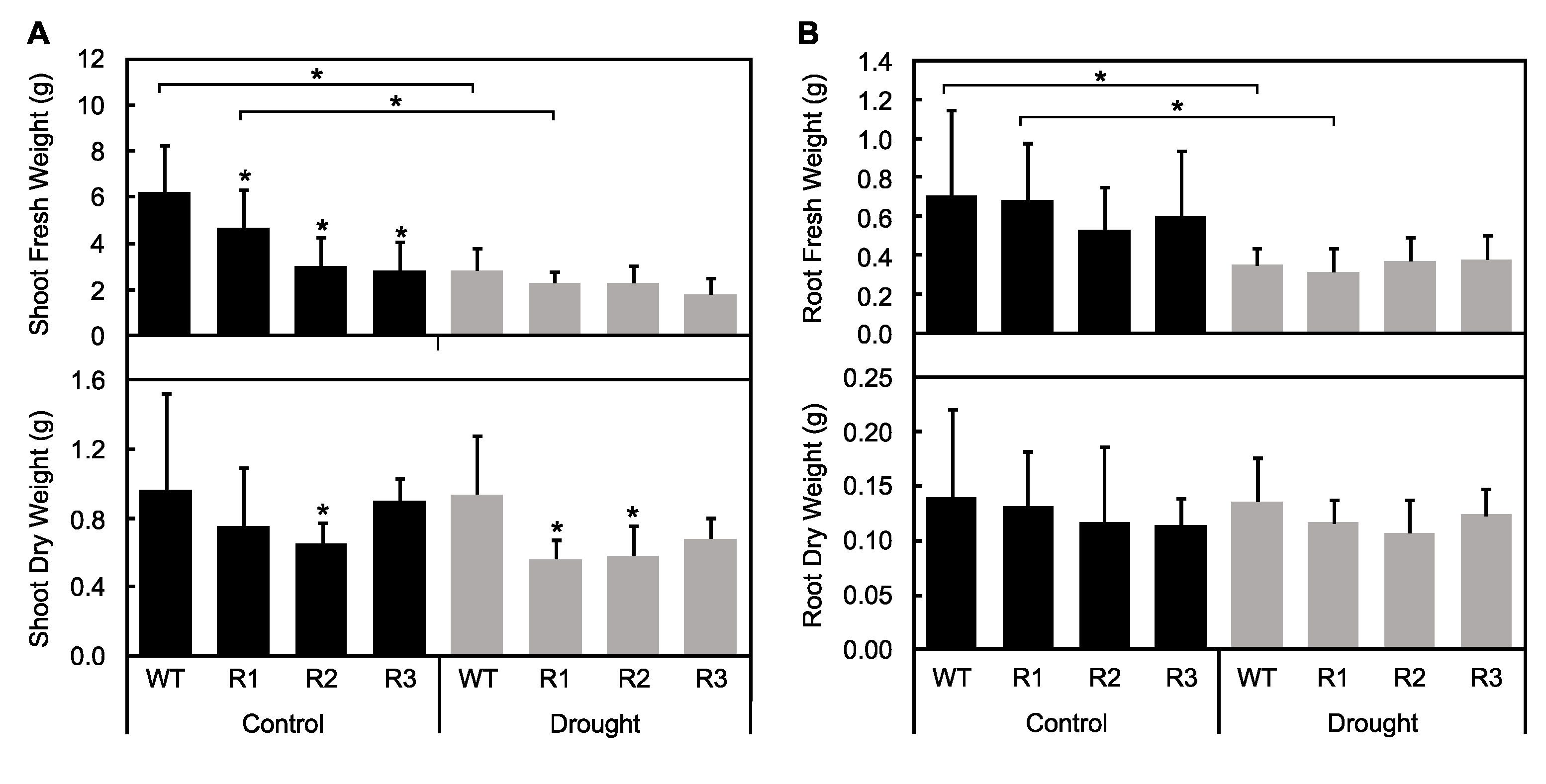
2.2. Response to Drought Stress in SPL9-RNAi Plants
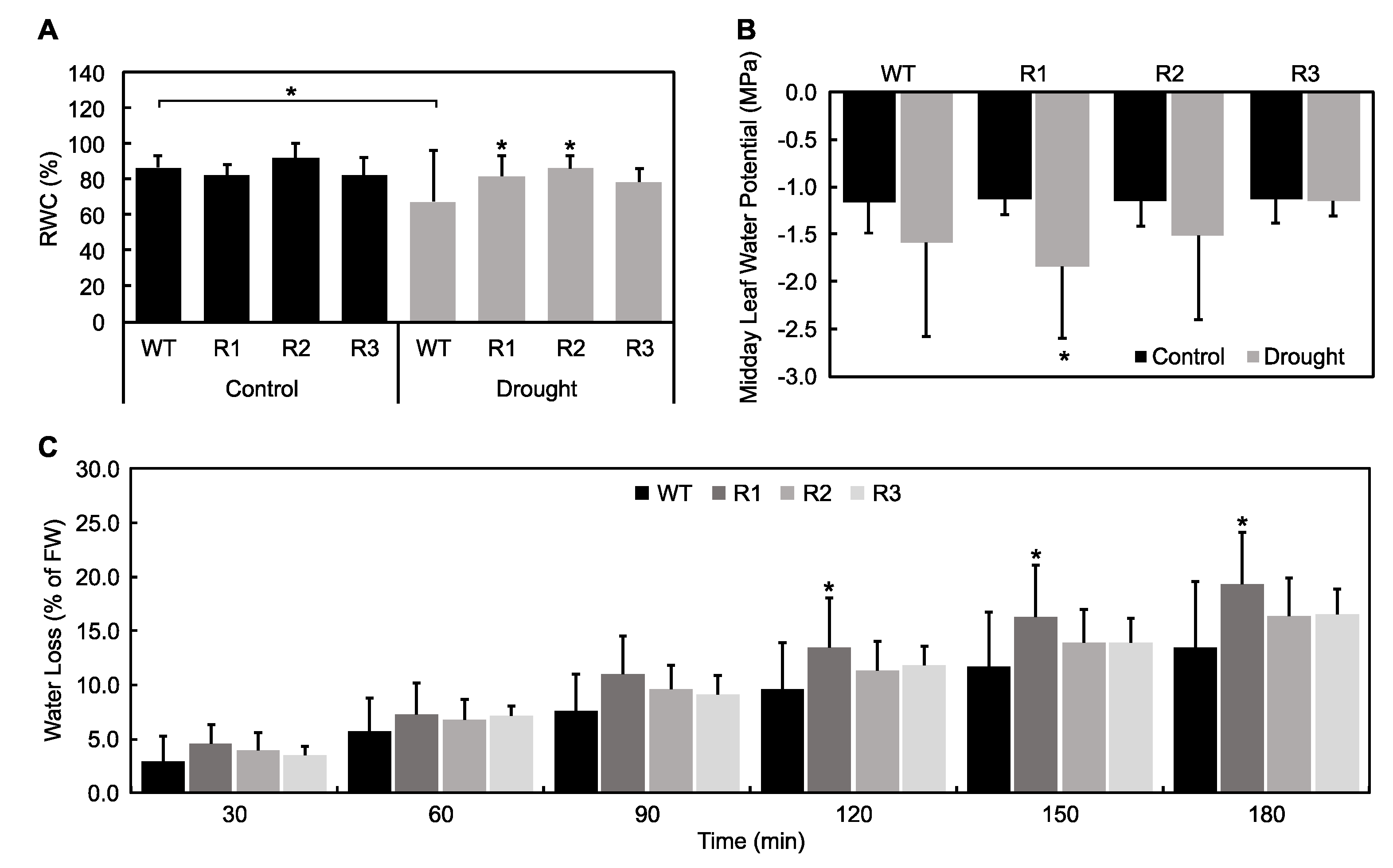
2.3. Evaluation of Plant Water Status under Drought Stress
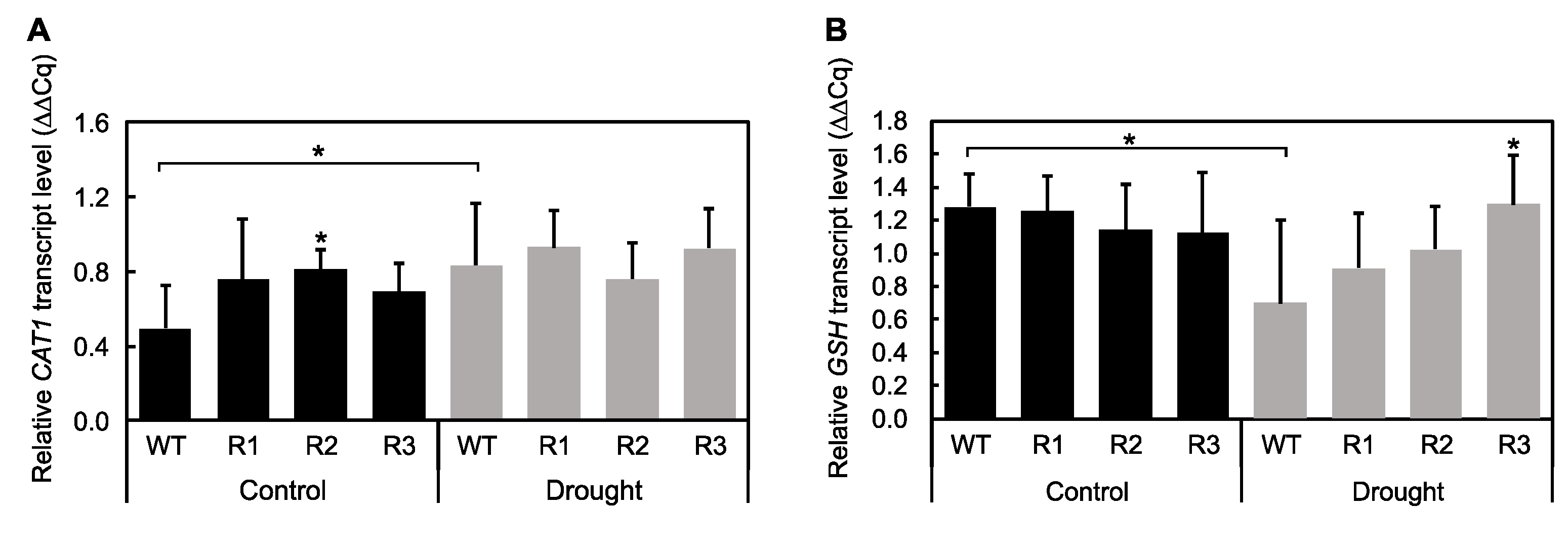
2.4. Effect of Drought on the Antioxidant-Mediated ROS Scavenging Capabilities of SPL9-RNAi Alfalfa
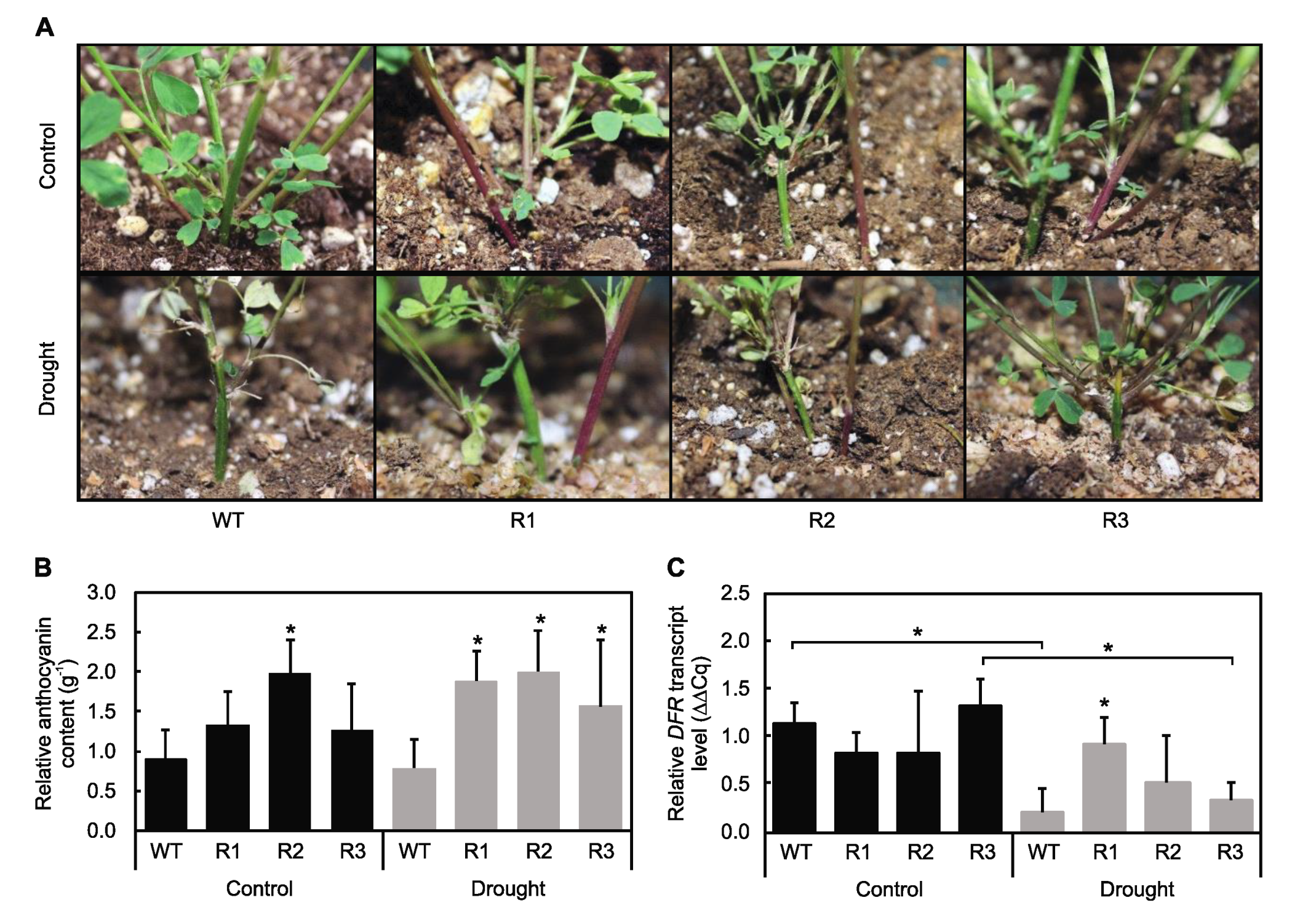
2.5. MsSPL9 Regulates Anthocyanin Biosynthesis
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. SPL9-RNAi Design and Construction
4.3. SPL9-RNAi Growing Conditions, Propagation, and Characterization
4.4. Evaluation of Drought Tolerance
4.5. Evaluation of Water Status
4.6. Stomatal Conductance
4.7. Extraction and Analysis of Anthocyanin and Antioxidants
4.8. cDNA Synthesis and qRT-PCR Analysis
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CAT1 | Catalase 1 |
| DFR | Dihydroflavonol 4-reductase |
| DW | Dry weight |
| FW | Fresh weight |
| GSH | Glutathione synthase |
| OE | Overexpression |
| RNAi | RNA interference |
| ROS | Reactive oxygen species |
| RWC | Relative water content |
| SBP | Squamosal promoter binding domain |
| SPL | Squamosa promoter binding protein-like |
| WT | Wild type |
References
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobell, D.B.; Schlenker, W.; Costa-Roberts, J. Climate trends and global crop production since 1980. Science 2011, 333, 616–620. [Google Scholar] [CrossRef] [Green Version]
- Schindler, D.W.; Donahue, W.F. An impending water crisis in Canada’s western prairie provinces. Proc. Natl. Acad. Sci. USA 2006, 103, 7210–7216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daryanto, S.; Wang, L.; Jacinthe, P.-A. Global synthesis of drought effects on maize and wheat production. PLoS ONE 2016, 11, e0156362. [Google Scholar] [CrossRef] [PubMed]
- Statistics Canada 2016. Hay and Field Crops. Available online: https://www150.statcan.gc.ca/t1/tbl1/en/tv.action?pid=3210041601&pickMembers%5B0%5D=1.1&pickMembers%5B1%5D=2.22 (accessed on 21 February 2020).
- Marković, J.; Radović, J.; Lugić, Z.; Sokolovic, D. The effect of development stage on chemical composition of alfalfa leaf and stem. Biotechnol. Anim. Husb. 2007, 23, 383–388. [Google Scholar] [CrossRef] [Green Version]
- Radović, J.; Sokolović, D.; Marković, J. Alfalfa-most important perennial forage legume in animal husbandry. Biotechnol. Anim. Husb. 2009, 25, 465–475. [Google Scholar] [CrossRef]
- Attram, J.; Acharya, S.N.; Woods, S.A.; Smith, E.; Thomas, J.E. Yield and net return from alfalfa cultivars under irrigation in Southern Alberta. Can. J. Plant Sci. 2016, 96, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Lamaoui, M.; Jemo, M.; Datla, R.; Bekkaoui, F. Heat and drought stresses in crops and approaches for their mitigation. Front. Chem. 2018, 6, 26. [Google Scholar] [CrossRef]
- Rivero, R.M.; Kojima, M.; Gepstein, A.; Sakakibara, H.; Mittler, R.; Gepstein, S.; Blumwald, E. Delayed leaf senescence induces extreme drought tolerance in a flowering plant. Proc. Natl. Acad. Sci. USA 2007, 104, 19631–19636. [Google Scholar] [CrossRef] [Green Version]
- Sicher, R.C.; Timlin, D.; Bailey, B. Responses of growth and primary metabolism of water-stressed barley roots to rehydration. J. Plant. Physiol. 2012, 169, 686–695. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Jiang, J.-G. Osmotic adjustment and plant adaptation to environmental changes related to drought and salinity. Environ. Rev. 2010, 18, 309–319. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhury, S.; Panda, P.; Sahoo, L.; Panda, S.K. Reactive oxygen species signaling in plants under abiotic stress. Plant Signal Behav. 2013, 8, e23681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarker, U.; Oba, S. Catalase, superoxide dismutase and ascorbate-glutathione cycle enzymes confer drought tolerance of Amaranthus tricolor. Sci. Rep. 2018, 8, 16496. [Google Scholar] [CrossRef] [Green Version]
- Rüegsegger, A.; Schmutz, D.; Brunold, C. Regulation of glutathione synthesis by cadmium in Pisum sativum L. Plant Physiol. 1990, 93, 1579–1584. [Google Scholar] [CrossRef] [Green Version]
- Arshad, M.; Feyissa, B.A.; Amyot, L.; Aung, B.; Hannoufa, A. MicroRNA156 improves drought stress tolerance in alfalfa (Medicago sativa) by silencing SPL13. Plant Sci. 2017, 258, 122–136. [Google Scholar] [CrossRef]
- Aung, B.; Gruber, M.Y.; Amyot, L.; Omari, K.; Bertrand, A.; Hannoufa, A. MicroRNA156 as a promising tool for alfalfa improvement. Plant Biotechnol. J. 2015, 13, 779–790. [Google Scholar] [CrossRef]
- Arshad, M.; Gruber, M.Y.; Wall, K.; Hannoufa, A. An insight into microRNA156 role in salinity stress responses of alfalfa. Front. Plant Sci. 2017, 8, 356. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.-G.; Shan, J.-X.; Shi, M.; Gao, J.-P.; Lin, H.-X. The miR156-SPL9-DFR pathway coordinates the relationship between development and abiotic stress tolerance in plants. Plant J. 2014, 80, 1108–1117. [Google Scholar] [CrossRef]
- Feyissa, B.A.; Arshad, M.; Gruber, M.Y.; Kohalmi, S.E.; Hannoufa, A. The interplay between miR156/SPL13 and DFR/WD40-1 regulate drought tolerance in alfalfa. BMC Plant Biol. 2019, 19, 434–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthews, C.; Arshad, M.; Hannoufa, A. Alfalfa response to heat stress is modulated by microRNA156. Physiol. Plant 2019, 165, 830–842. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, S.; Grande, A.V.; Bujdoso, N.; Saedler, H.; Huijser, P. The microRNA regulated SBP-box genes SPL9 and SPL15 control shoot maturation in Arabidopsis. Plant Mol. Biol. 2008, 67, 183–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, N.; Niu, Q.-W.; Ng, K.-H.; Chua, N.-H. The role of miR156/SPLs modules in Arabidopsis lateral root development. Plant J. 2015, 83, 673–685. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Park, M.Y.; Conway, S.R.; Wang, J.-W.; Weigel, D.; Poethig, R.S. The sequential action of miR156 and miR172 regulates developmental timing in Arabidopsis. Cell 2009, 138, 750–759. [Google Scholar] [CrossRef] [Green Version]
- Li, R.-J.; Li, L.M.; Liu, X.L.; Kim, J.C.; Jenks, M.A.; Lu, S. Diurnal regulation of plant epidermal wax synthesis through antagonistic roles of the transcription factors SPL9 and DEWAX. Plant Cell 2019, 31, 2711–2733. [Google Scholar] [CrossRef]
- Gao, R.; Austin, R.S.; Amyot, L.; Hannoufa, A. Comparative transcriptome investigation of global gene expression changes caused by miR156 overexpression in Medicago sativa. BMC Genom. 2016, 17, 658–672. [Google Scholar] [CrossRef] [Green Version]
- Gou, J.-Y.; Felippes, F.F.; Liu, C.-J.; Weigel, D.; Wang, J.-W. Negative regulation of anthocyanin biosynthesis in Arabidopsis by a miR156-targeted SPL transcription factor. Plant Cell 2011, 23, 1512–1522. [Google Scholar] [CrossRef] [Green Version]
- Aerts, R.J.; Barry, T.N.; McNabb, W.C. Polyphenols and agriculture: Beneficial effects of proanthocyanidins in forages. Agric. Ecosyst. Environ. 1999, 75, 1–12. [Google Scholar] [CrossRef]
- Dixon, R.A.; Liu, C.; Jun, J.H. Metabolic engineering of anthocyanins and condensed tannins in plants. Curr. Opin. Biotechnol. 2013, 24, 329–335. [Google Scholar] [CrossRef]
- Jones, W.T.; Mangan, J.L. Complexes of the condensed tannins of sainfoin (Onobrychis viciifolia scop.) with fraction 1 leaf protein and with submaxillary mucoprotein, and their reversal by polyethylene glycol and pH. J. Sci. Food Agric. 1977, 28, 126–136. [Google Scholar] [CrossRef]
- Waghorn, G.C.; Ulyatt, M.J.; John, A.; Fisher, M.T. The effect of condensed tannins on the site of digestion of amino acids and other nutrients in sheep fed on Lotus corniculatus L. Br. J. Nutr. 1987, 57, 115–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakatsuka, T.; Abe, Y.; Kakizaki, Y.; Yamamura, S.; Nishihara, M. Production of red-flowered plants by genetic engineering of multiple flavonoid biosynthetic genes. Plant Cell Rep. 2007, 26, 1951–1959. [Google Scholar] [CrossRef] [PubMed]
- Azuma, K.; Ohyama, A.; Ippoushi, K.; Ichiyanagi, T.; Takeuchi, A.; Saito, T.; Fukuoka, H. Structures and antioxidant activity of anthocyanins in many accessions of eggplant and its related species. J. Agric. Food Chem. 2008, 56, 10154–10159. [Google Scholar] [CrossRef]
- Kähkönen, M.P.; Heinonen, M. Antioxidant activity of anthocyanins and their aglycons. J. Agric. Food Chem. 2003, 51, 628–633. [Google Scholar] [CrossRef]
- Badhan, A.; Jin, L.; Wang, Y.; Han, S.; Kowalczys, K.; Brown, D.C.; Ayala, C.J.; Latoszek-Green, M.; Miki, B.; Tsang, A.; et al. Expression of a fungal ferulic acid esterase in alfalfa modifies cell wall digestibility. Biotechnol. Biofuels 2014, 7, 39–53. [Google Scholar] [CrossRef] [Green Version]
- Tian, L.; Wang, H.; Wu, K.; Latoszek-Green, M.; Hu, M.; Miki, B.; Brown, D. Efficient recovery of transgenic plants through organogenesis and embryogenesis using a cryptic promoter to drive marker gene expression. Plant Cell Rep. 2002, 20, 1181–1187. [Google Scholar] [CrossRef]
- Anderson, J.E.; McNaughton, S.J. Effects of low soil temperature on transpiration, photosynthesis, leaf relative water content, and growth among elevationally diverse plant populations. Ecology 1973, 54, 1220–1233. [Google Scholar] [CrossRef]
- Weatherley, P.E. Studies in the water relations of the cotton plant I. The field measurement of water deficits in leaves. New Phytol. 1950, 49, 81–97. [Google Scholar] [CrossRef]
- Turner, N.C. Measurement of plant water status by the pressure chamber technique. Irrig. Sci. 1988, 9, 289–308. [Google Scholar] [CrossRef]
- Neff, M.M.; Chory, J. Genetic interactions between phytochrome A, phytochrome B, and cryptochrome 1 during Arabidopsis development. Plant Physiol. 1998, 118, 27–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | 2-Month-Old Alfalfa | |||
|---|---|---|---|---|
| WT | R1 | R2 | R3 | |
| # Main branches | 2 ± 0.82 | 2 ± 0.67 | 2 ± 0.47 | 3 ± 1.03 |
| # Lateral branches | 7 ± 4.19 | 9 ± 5.43 | 7 ± 4.86 | 11 ± 3.68 |
| Total branching | 9 ± 4.57 | 11 ± 5.87 | 9 ± 4.89 | 14 ± 3.74 |
| # of nodes | 10 ± 2.80 | 10 ± 3.05 | 10 ± 1.69 | 11 ± 2.92 |
| Plant height (cm) | 32.7 ± 12.79 | 32.4 ± 7.84 | 33.3 ± 5.76 | 36.5 ± 6.56 |
| Average internode length (cm) | 3.5 ± 1.61 | 3.3 ± 1.01 | 3.5 ± 0.64 | 3.5 ± 0.95 |
| Stem thickness (cm) | 1.53 ± 0.14 | 1.25 ± 0.16 * | 1.23 ± 0.13 * | 1.27 ± 0.10 * |
| Fresh weight (g) | 3.61 ± 2.60 | 4.09 ± 2.98 | 3.23 ± 2.67 | 4.07 ± 2.48 |
| Dry weight (g) | 0.94 ± 0.60 | 0.99 ± 0.69 | 0.75 ± 0.56 | 1.11 ± 0.64 |
| Time to flower (days) | NF | NF | NF | NF |
| Root length (cm) | 37.9 ± 3.84 | 41.4 ± 9.35 | 38.8 ± 7.89 | 40.5 ± 5.01 |
| Root fresh weight (g) | 4.78 ± 3.20 | 6.03 ± 4.58 | 4.69 ± 3.69 | 5.94 ± 3.87 |
| Root dry weight (g) | 0.97 ± 0.31 | 0.96 ± 0.40 | 0.88 ± 0.27 | 1.30 ± 0.36 |
| Genotype | 6-Month-Old Alfalfa | |||
|---|---|---|---|---|
| WT | R1 | R2 | R3 | |
| # Main branches | 27 ± 5.72 | 24 ± 9.52 | 15 ± 6.79 * | 31 ± 7.78 |
| # Lateral branches | 104 ± 39.04 | 151 ± 31.20 * | 113 ± 22.80 | 182 ± 34.08 * |
| Total branching | 131 ± 40.37 | 175 ± 36.32 * | 128 ± 20.06 | 213 ± 34.61 * |
| # of nodes | 19 ± 1.48 | 19 ± 1.55 | 17 ± 0.00 * | 18 ± 0.76 |
| Plant height (cm) | 86.7 ± 12.17 | 70.4 ± 2.75 * | 71.8 ± 7.12 * | 63.6 ± 3.92 * |
| Average internode length (cm) | 4.6 ± 0.56 | 3.7 ± 0.22 * | 4.2 ± 0.37 | 3.5 ± 0.18 * |
| Fresh weight (g) | 130.62 ± 22.02 | 115.41 ± 22.92 | 101.02 ± 26.08 | 125.58 ± 21.71 |
| Dry weight (g) | 33.46 ± 5.61 | 28.55 ± 6.30 | 24.65 ± 6.69 | 33.79 ± 5.87 |
| Time to flower (days) | 114 ± 8.77 | 113 ± 6.18 | 146 ± 25.56 * | 122 ± 15.55 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hanly, A.; Karagiannis, J.; Lu, Q.S.M.; Tian, L.; Hannoufa, A. Characterization of the Role of SPL9 in Drought Stress Tolerance in Medicago sativa. Int. J. Mol. Sci. 2020, 21, 6003. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176003
Hanly A, Karagiannis J, Lu QSM, Tian L, Hannoufa A. Characterization of the Role of SPL9 in Drought Stress Tolerance in Medicago sativa. International Journal of Molecular Sciences. 2020; 21(17):6003. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176003
Chicago/Turabian StyleHanly, Alexandria, Jim Karagiannis, Qing Shi Mimmie Lu, Lining Tian, and Abdelali Hannoufa. 2020. "Characterization of the Role of SPL9 in Drought Stress Tolerance in Medicago sativa" International Journal of Molecular Sciences 21, no. 17: 6003. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176003