Biosurfactants Induce Antimicrobial Peptide Production through the Activation of TmSpatzles in Tenebrio molitor

 , , ,
, , ,
Abstract
:
1. Introduction
2. Results
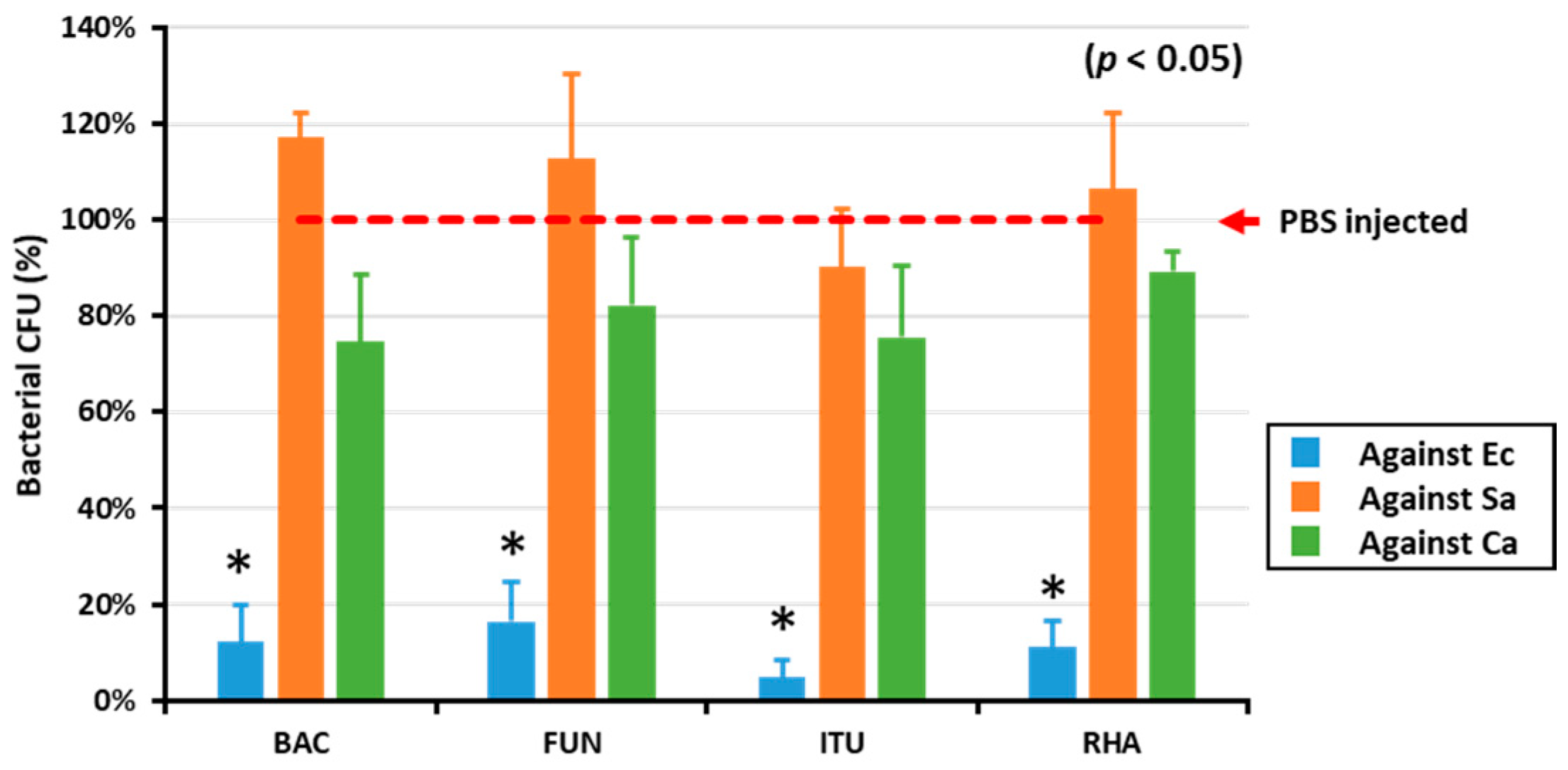
2.1. AMP Activity in the Hemolymph of Biosurfactant-Injected T. molitor Larvae
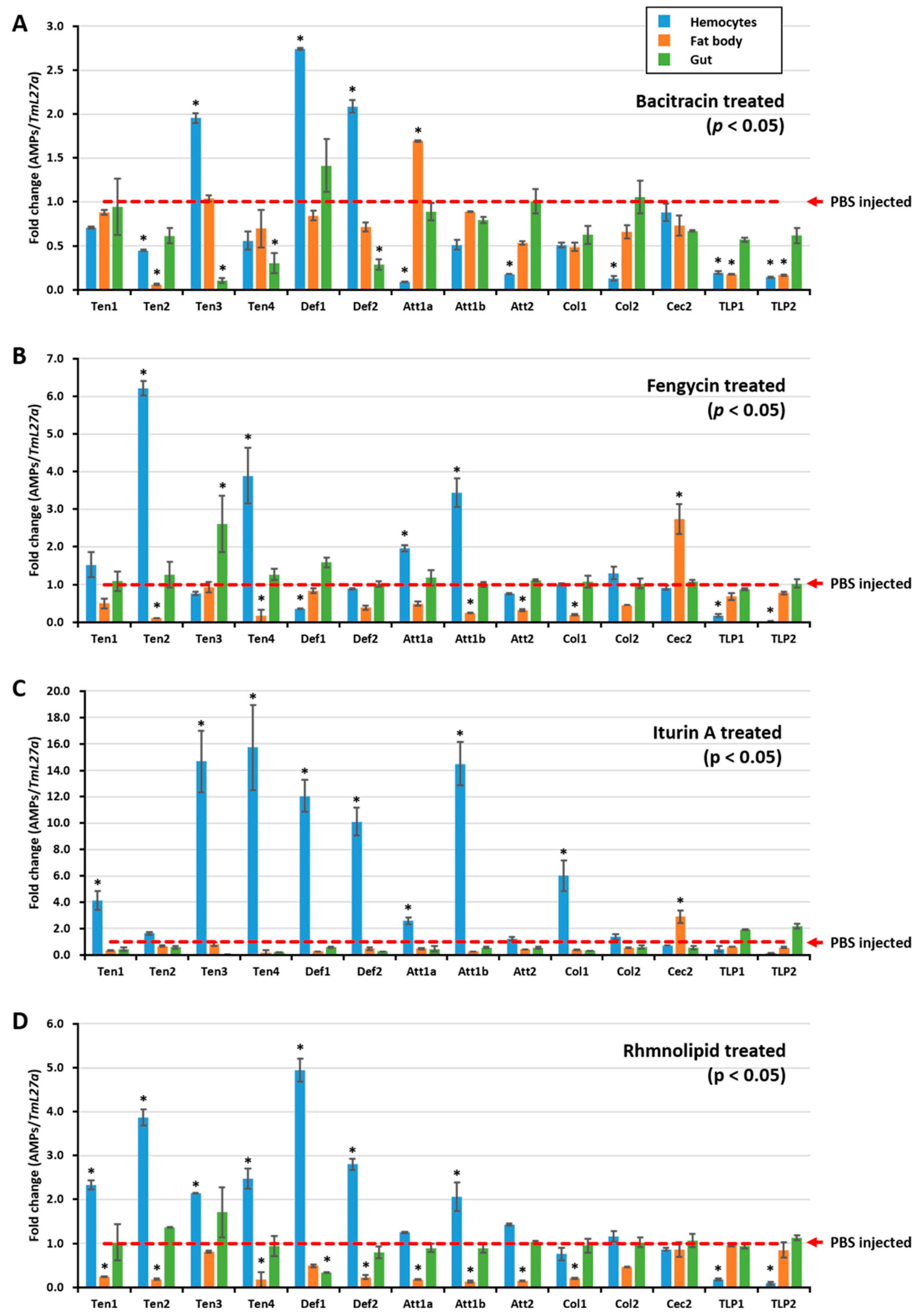
2.2. Biosurfactants Induced AMP Expression in T. molitor Hemocytes
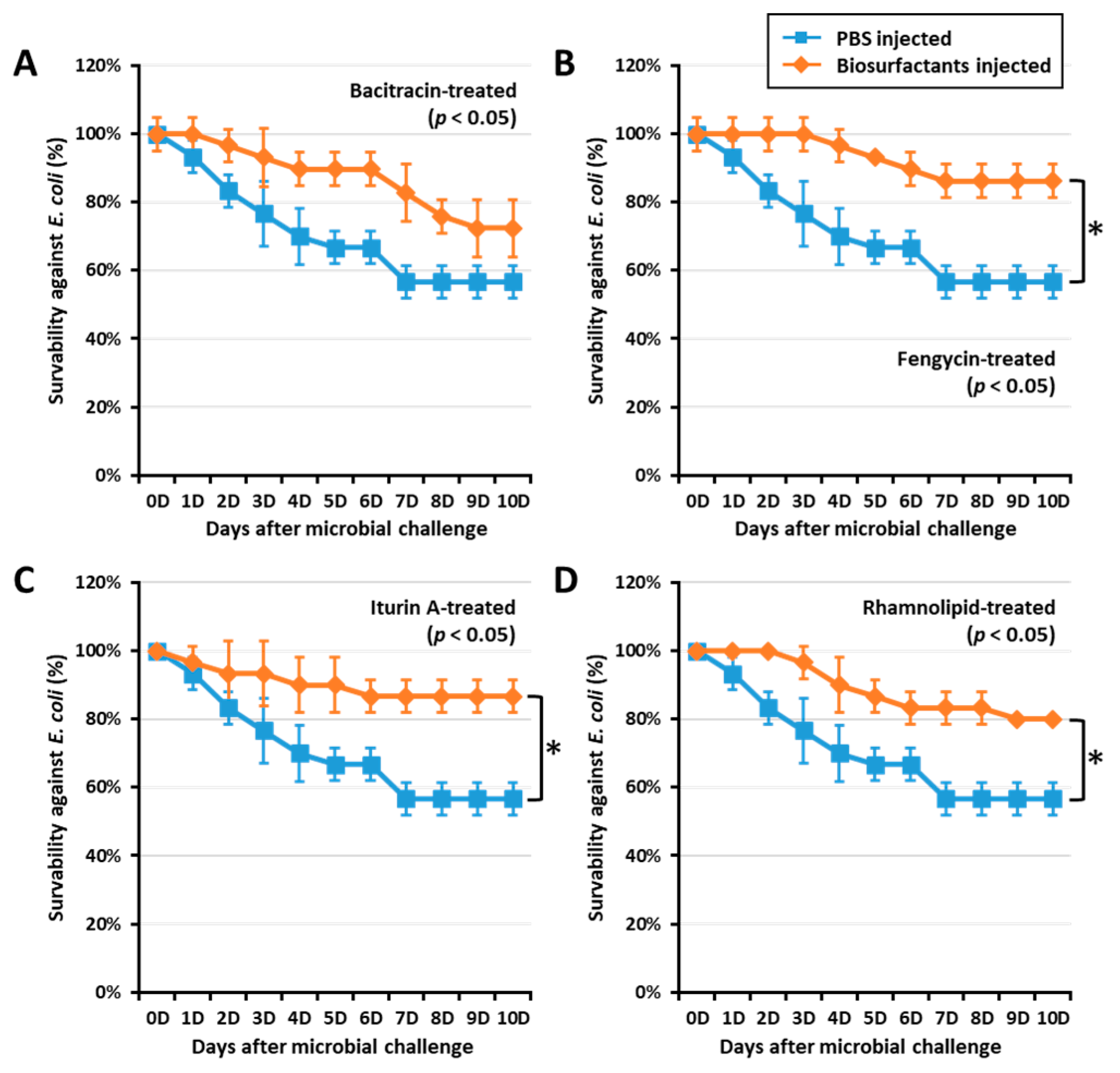
2.3. Biosurfactants Increased T. molitor Larvae Survivability against E. coli
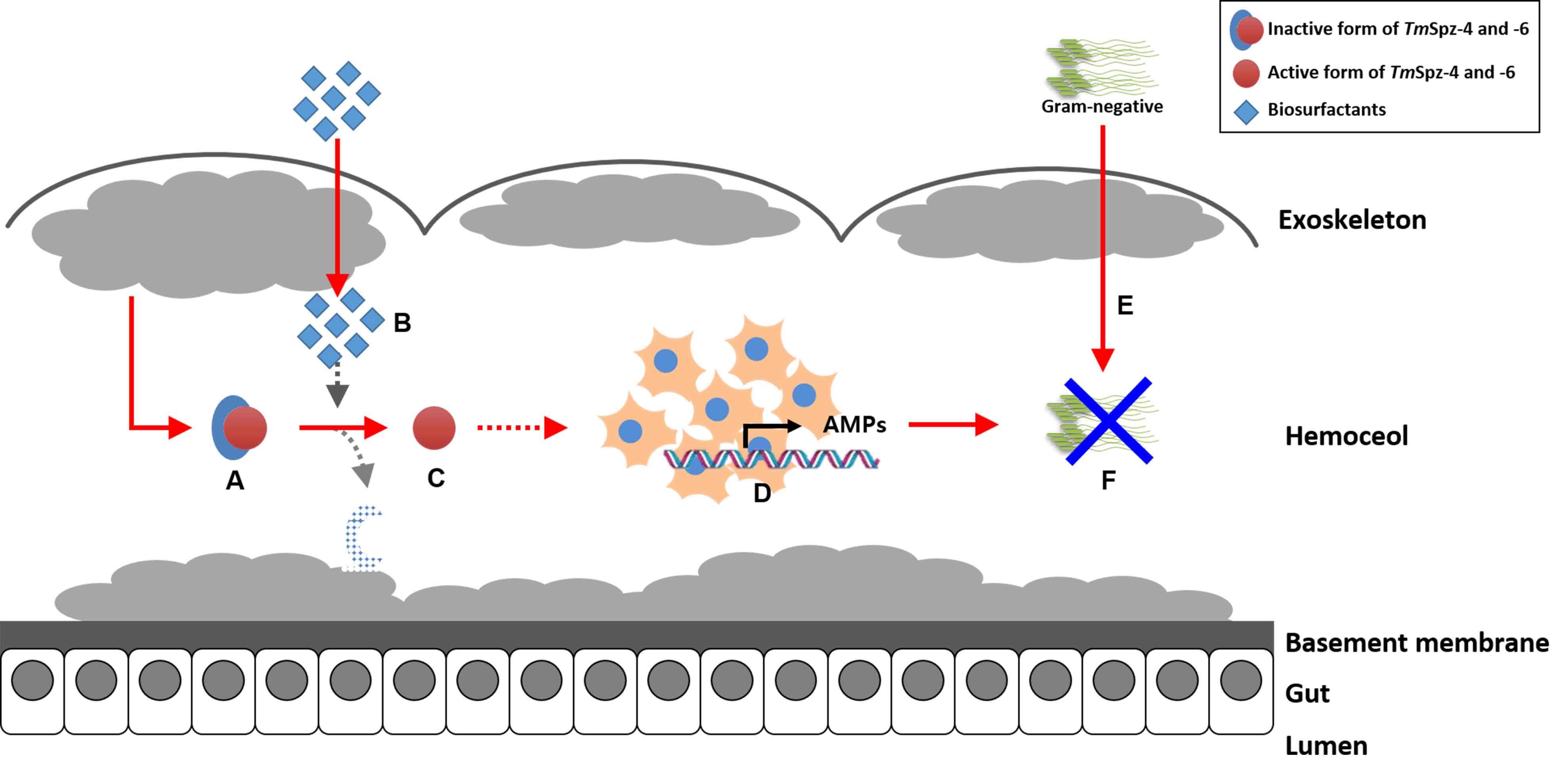
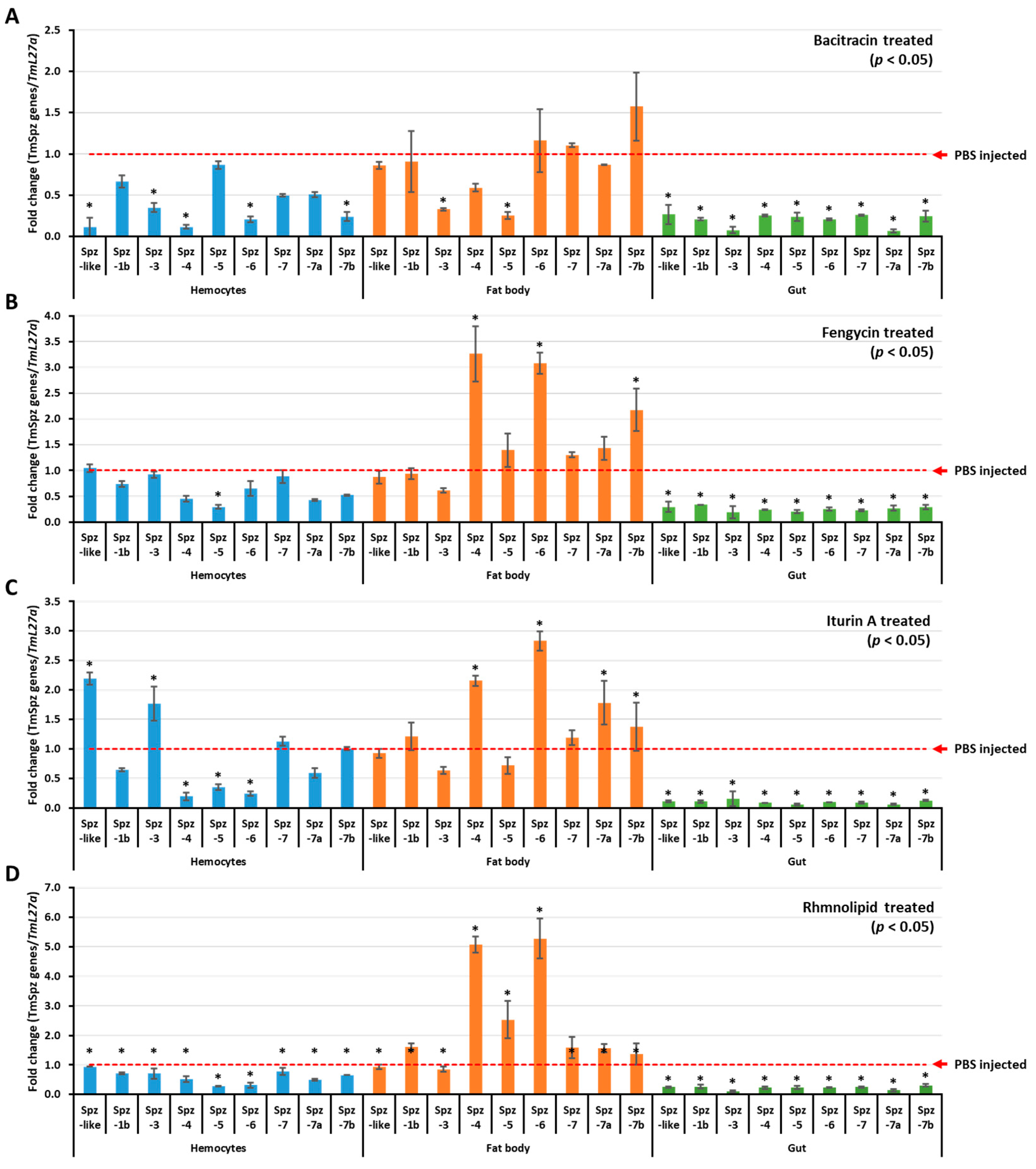
2.4. Biosurfactants Activated the Expression of Spätzle Genes in T. molitor
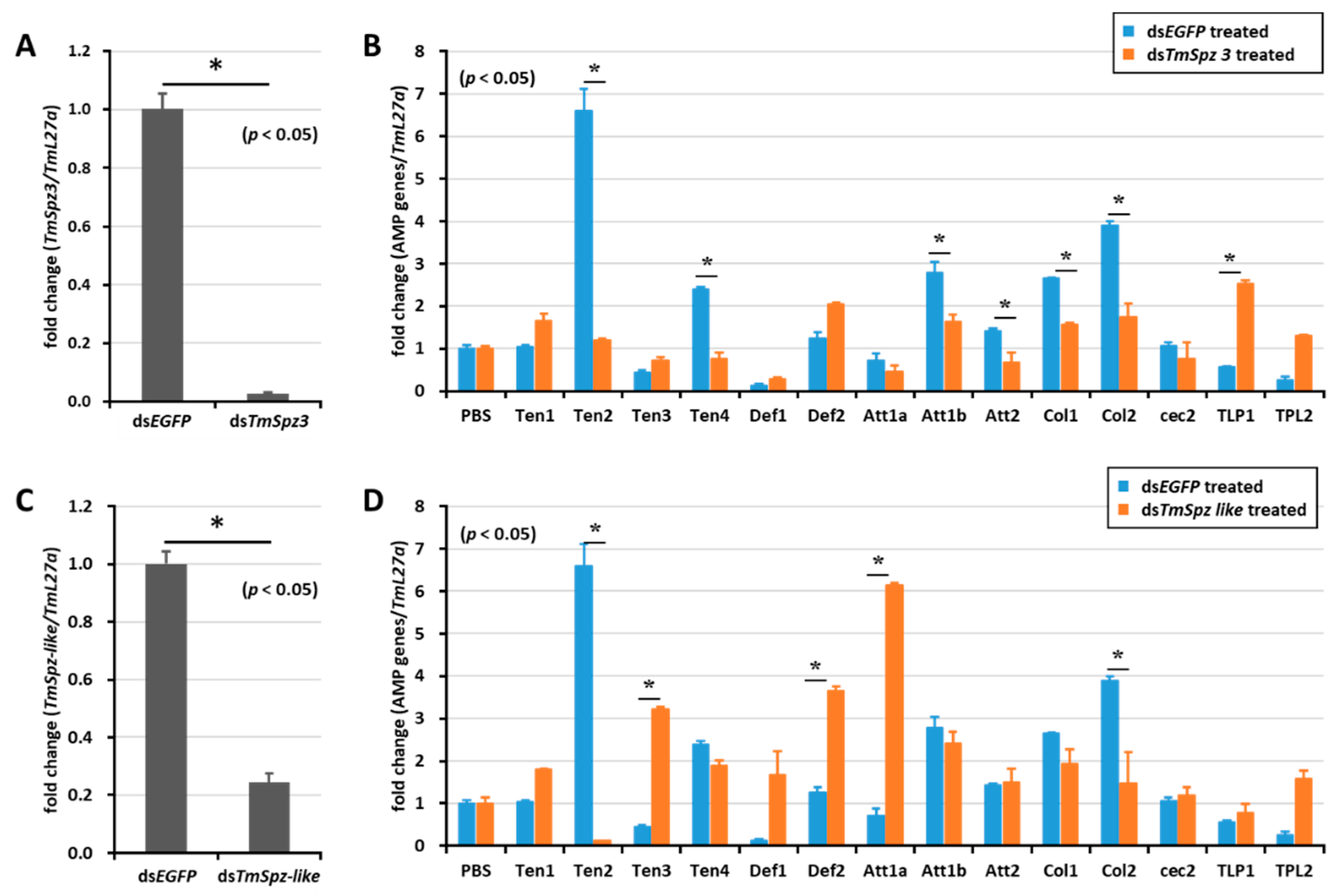
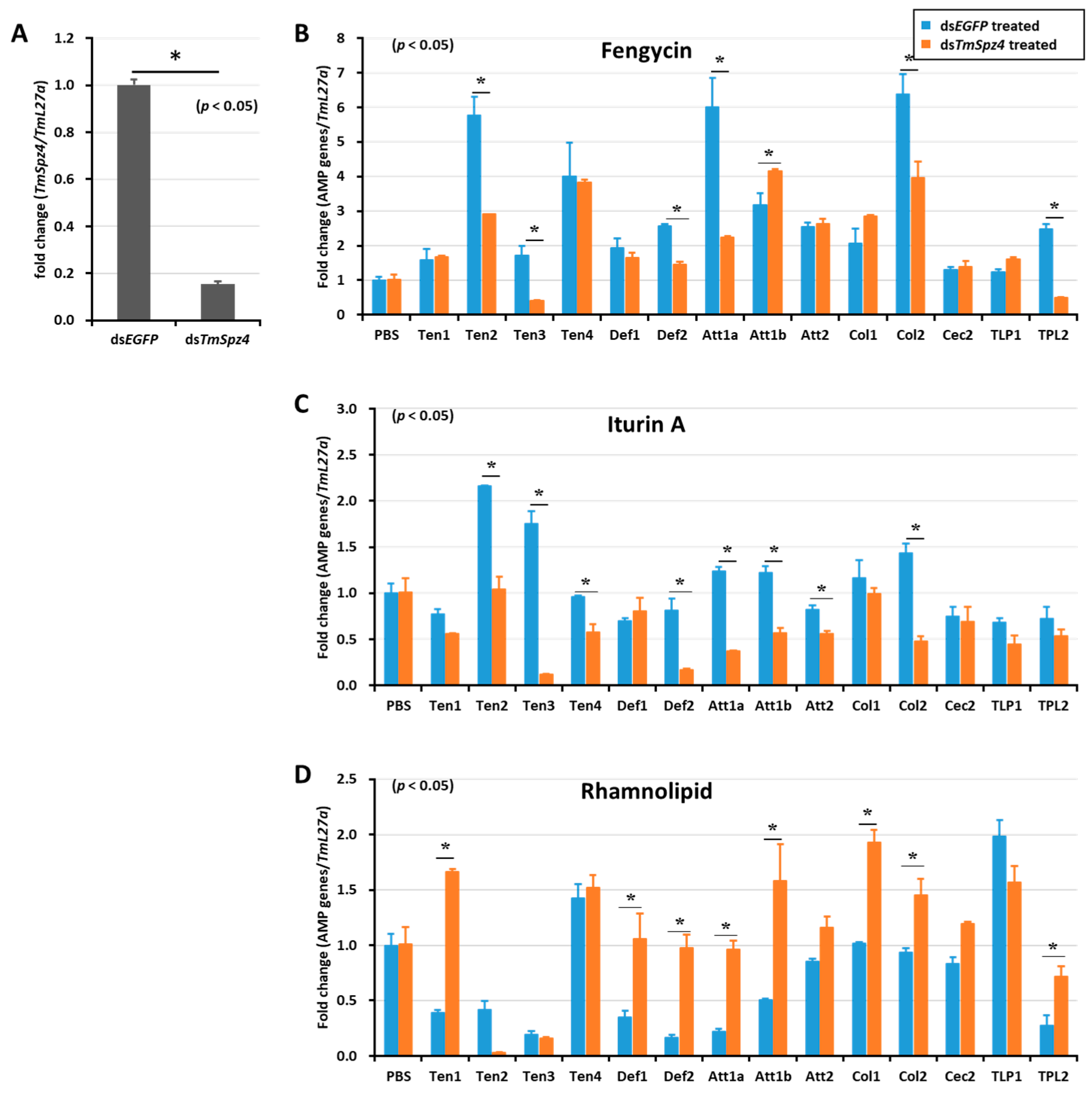
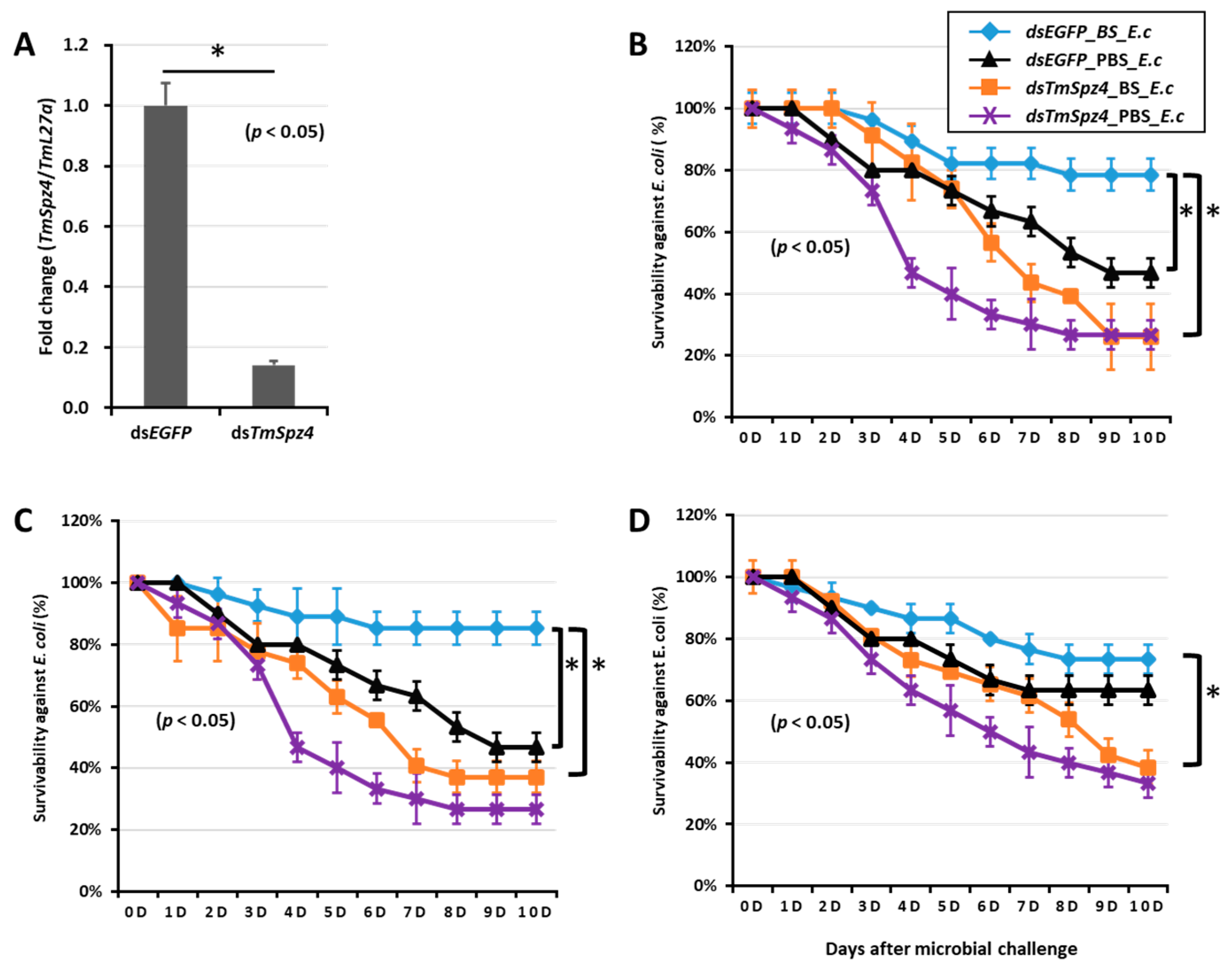
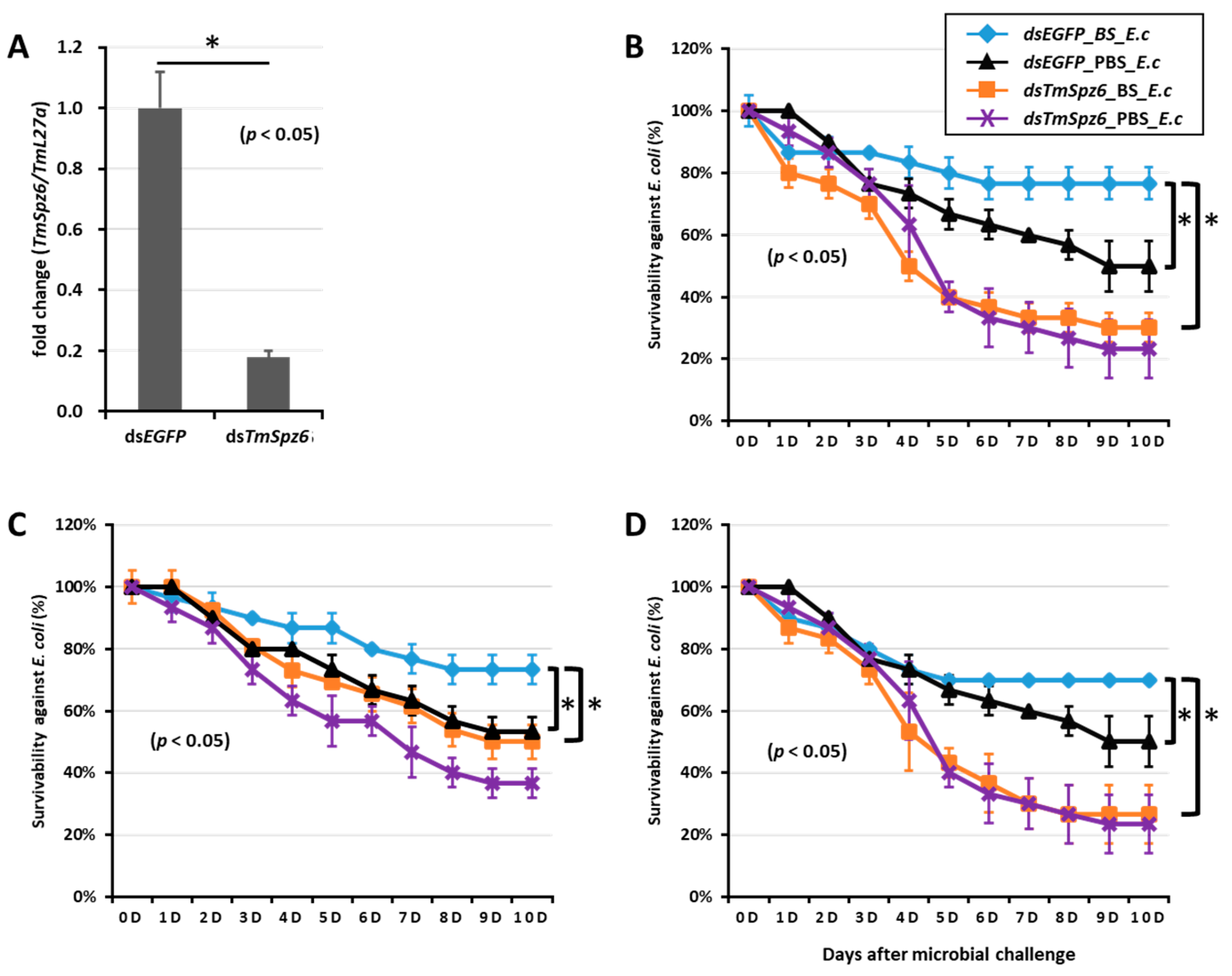
2.5. Effect of TmSpz Gene Silencing on the Expression of AMPs in T. molitor
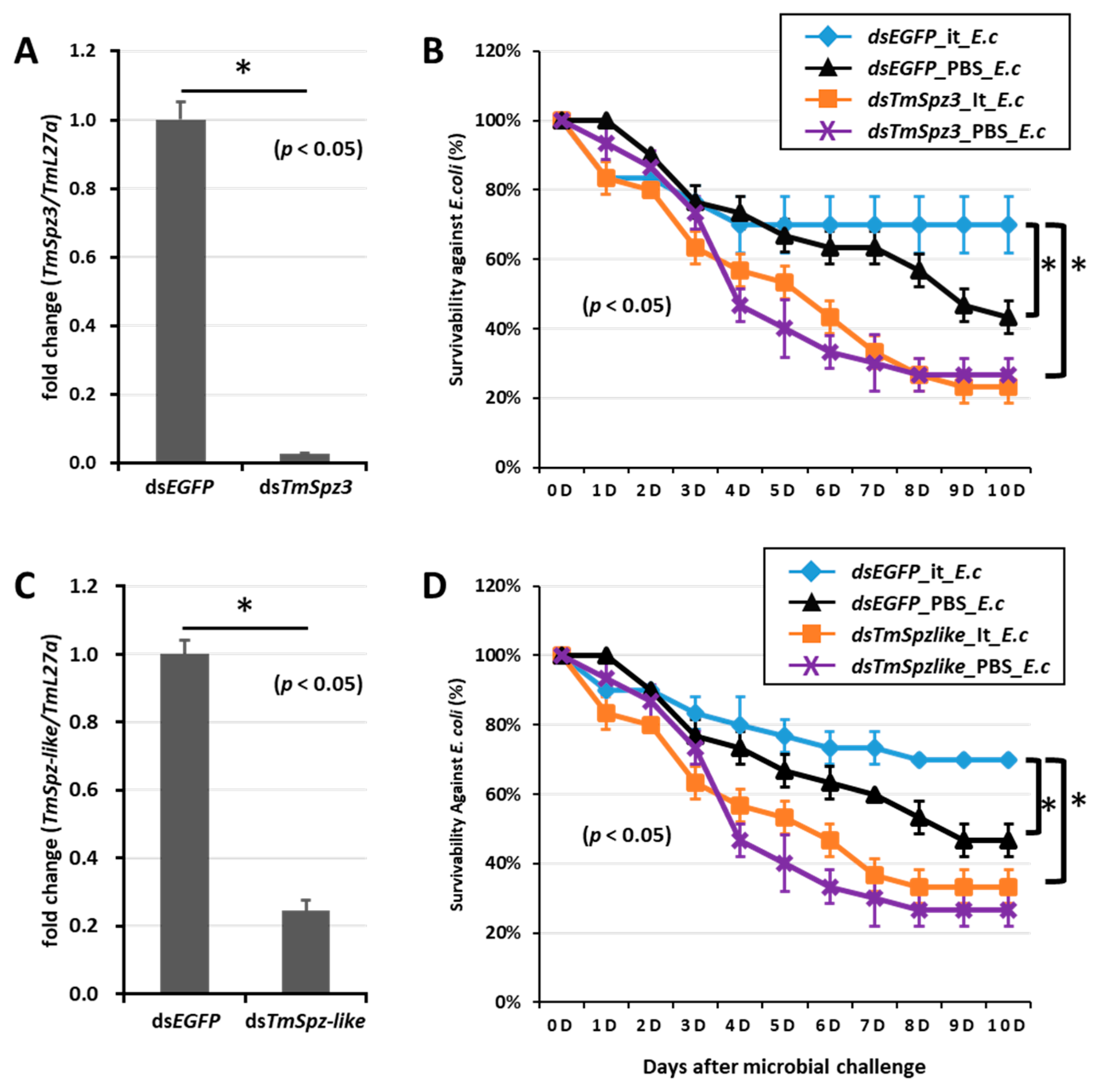
2.6. T. molitor Larvae Survivability Study
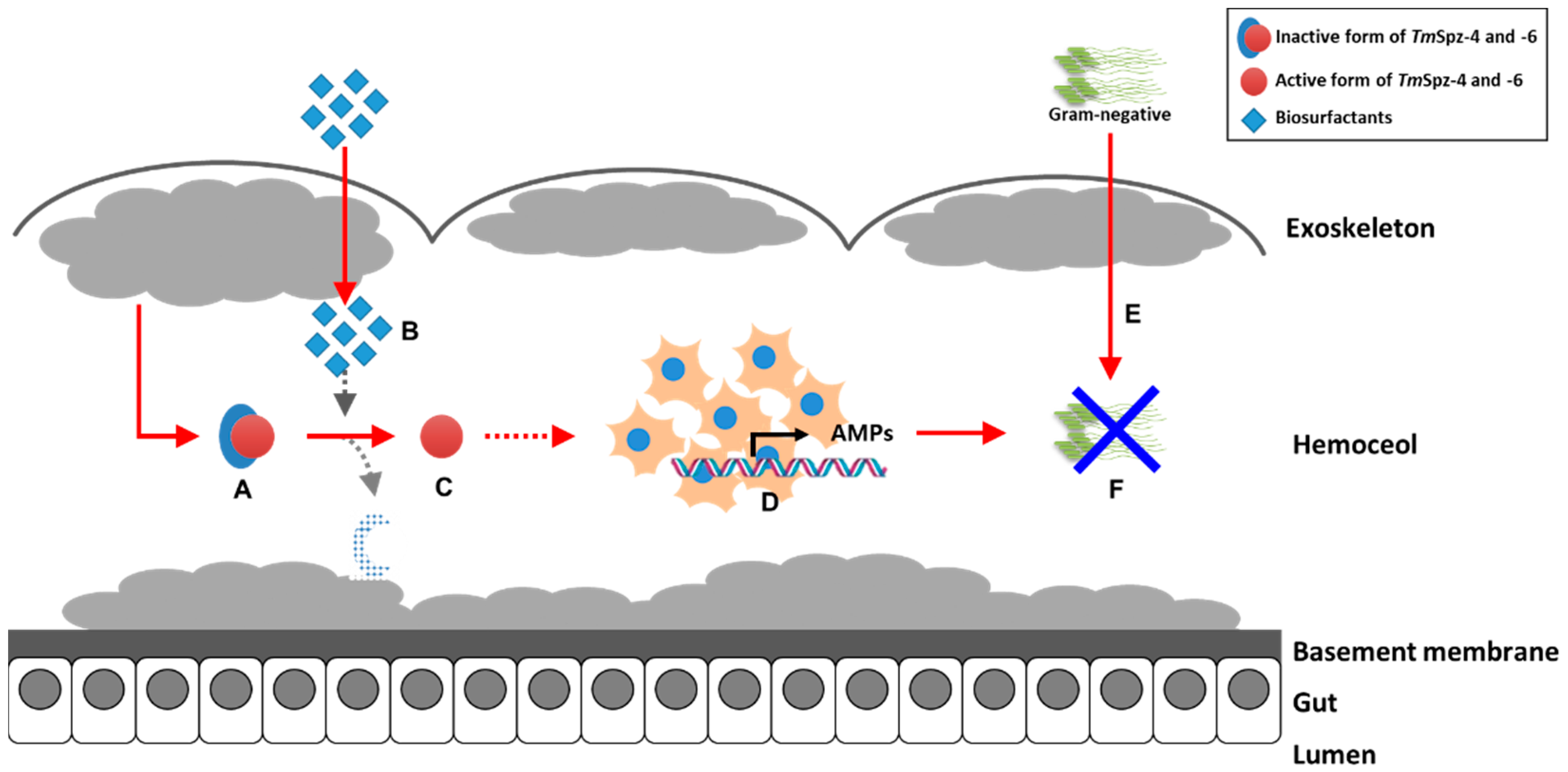
3. Discussion
4. Materials and Methods
4.1. Insect Culture
4.2. Source of Biosurfactants
4.3. Preparation of Microorganisms
4.4. AMPs Activity Study
4.5. RNA Extraction, cDNA Synthesis, and Expression of AMPs
4.6. Biosurfactant-Triggered Immune Survivability of T. molitor
4.7. Screening of Biosurfactant-Activated T. molitor Spatzle Genes
4.8. Effect of Tm Spätzle Gene Silencing on the Expression of AMPs
4.9. Effect of TmSpz Genes Silencing on T. molitor Survivability
4.10. Data Analyses
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Shekhar, S.; Sundaramanickam, A.; Balasubramanian, T. Biosurfactant producing microbes and their potential applications: A review. Crit. Rev. Environ. Sci. Technol. 2015, 45, 1522–1554. [Google Scholar] [CrossRef]
- Vijayakuma, S.; Varatharajan, S. Biosurfactants-types, sources and applications. Res. J. Microbiol. 2015, 10, 181–192. [Google Scholar]
- Edwards, J.R.; Hayashi, J.A. Structure of a rhamnolipid from pseudomonas aeruginosa. Arch. Biochem. Biophys. 1965, 111, 415–421. [Google Scholar] [CrossRef]
- Asselineau, C.; Asselineau, J. Trehalose-containing glycolipids. Prog. Chem. Fats Other Lipids 1978, 16, 59–99. [Google Scholar] [CrossRef]
- Gautam, K.K.; Tyagi, V.K. Microbial surfactants: A review. J.Oleo Sci. 2006, 55, 155–166. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarti, S. Bacterial Biosurfactant: Characterization, Antimicrobial and Metal Remediation Properties. Master’s Thesis, National Institute of Technology, Rourkela, India, 2012. [Google Scholar]
- Cavalero, D.A.; Cooper, D.G. The effect of medium composition on the structure and physical state of sophorolipids produced by candida bombicola atcc 22214. J. Biotechnol. 2003, 103, 31–41. [Google Scholar] [CrossRef]
- Mohan, P.K.; Nakhla, G.; Yanful, E.K. Biokinetics of biodegradation of surfactants under aerobic, anoxic and anaerobic conditions. Water Res. 2006, 40, 533–540. [Google Scholar] [CrossRef]
- Nitschke, M.; Costa, S.G.V.A.O. Biosurfactants in food industry. Trends Food Sci. Technol. 2007, 18, 252–259. [Google Scholar] [CrossRef]
- Singh, P.; Cameotra, S.S. Potential applications of microbial surfactants in biomedical sciences. Trends Biotechnol. 2004, 22, 142–146. [Google Scholar] [CrossRef]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 16, 115–125. [Google Scholar] [CrossRef]
- Gordillo, A.; Maldonado, M.C. Purification of peptides from bacillus strains with biological activity. Chromatogr. Appl. 2012, 11, 201–225. [Google Scholar]
- Gamalero, E.; Glick, B. Bacteria in Agrobiology: Plant Nutrient Management; Springer: New York, NY, USA, 2011; pp. 17–46. [Google Scholar]
- Velho, R.; Medina, L.; Segalin, J.; Brandelli, A. Production of lipopeptides among bacillus strains showing growth inhibition of phytopathogenic fungi. Folia Microbiol. 2011, 56, 297. [Google Scholar] [CrossRef] [PubMed]
- Haddad, N.I.; Wang, J.; Mu, B. Isolation and characterization of a biosurfactant producing strain, brevibacilis brevis hob1. J. Ind. Microbiol. Biotechnol. 2008, 35, 1597–1604. [Google Scholar] [CrossRef] [PubMed]
- Vatsa, P.; Sanchez, L.; Clement, C.; Baillieul, F.; Dorey, S. Rhamnolipid biosurfactants as new players in animal and plant defense against microbes. Int. J. Mol. Sci. 2010, 11, 5095–5108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, L.; Banat, I.M.; Teixeira, J.; Oliveira, R. Biosurfactants: Potential applications in medicine. J. Antimicrob. Chemother. 2006, 57, 609–618. [Google Scholar] [CrossRef]
- Gudiña, E.J.; Rangarajan, V.; Sen, R.; Rodrigues, L.R. Potential therapeutic applications of biosurfactants. Trends Pharmacol. Sci. 2013, 34, 667–675. [Google Scholar] [CrossRef] [Green Version]
- Gharaei-Fathabad, E. Biosurfactants in pharmaceutical industry: A mini-review. Am. J. Drug Discov. Dev. 2011, 1, 58–69. [Google Scholar]
- Das, P.; Mukherjee, S.; Sen, R. Antimicrobial potential of a lipopeptide biosurfactant derived from a marine bacillus circulans. J. Appl. Microbiol. 2008, 104, 1675–1684. [Google Scholar] [CrossRef]
- Díaz De Rienzo, M.A.; Stevenson, P.; Marchant, R.; Banat, I.M. Antibacterial properties of biosurfactants against selected gram-positive and -negative bacteria. FEMS Microbiol. Lett. 2015, 363, fnv224. [Google Scholar] [CrossRef] [Green Version]
- Ahimou, F.; Jacques, P.; Deleu, M. Surfactin and iturin a effects on bacillus subtilis surface hydrophobicity. Enzym. Microb. Technol. 2000, 27, 749–754. [Google Scholar] [CrossRef]
- Tsuge, K.; Akiyama, T.; Shoda, M. Cloning, sequencing, and characterization of the iturin a operon. J. Bacteriol. 2001, 183, 6265–6273. [Google Scholar] [CrossRef] [Green Version]
- Akpa, E.; Jacques, P.; Wathelet, B.; Paquot, M.; Fuchs, R.; Budzikiewicz, H.; Thonart, P. Influence of culture conditions on lipopeptide production by bacillus subtilis. Appl. Biochem. Biotechnol. 2001, 91, 551–561. [Google Scholar] [CrossRef]
- Deleu, M.; Paquot, M.; Nylander, T. Effect of fengycin, a lipopeptide produced by bacillus subtilis, on model biomembranes. Biophys. J. 2008, 94, 2667–2679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadeq Al-wazni, W. Immunomodulator activity of biosurfactant extract from serratia marcescens. Int. J. Microbiol. Res. 2016, 7, 36–42. [Google Scholar]
- Gan, P.; Gao, Z.; Zhao, X.; Qi, G. Surfactin inducing mitochondria-dependent ros to activate mapks, nf-κb and inflammasomes in macrophages for adjuvant activity. Sci. Rep. 2016, 6, 39303. [Google Scholar] [CrossRef] [PubMed]
- Iatsenko, I.; Yim, J.J.; Schroeder, F.C.; Sommer, R.J.B. Subtilis gs67 protects c. Elegans from gram-positive pathogens via fengycin-mediated microbial antagonism. Curr. Biol. 2014, 24, 2720–2727. [Google Scholar] [CrossRef] [Green Version]
- Van Wees, S.C.; Van der Ent, S.; Pieterse, C.M. Plant immune responses triggered by beneficial microbes. Curr. Opin. Plant Biol. 2008, 11, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Sil, J.; Dandapat, P.; Das, S. Health care applications of different biosurfactants. Int. J. Sci. Res. 2017, 6, 41–50. [Google Scholar]
- Takeda, K.; Kaisho, T.; Akira, S. Toll-like receptors. Annu. Rev. Immunol. 2003, 21, 335–376. [Google Scholar] [CrossRef]
- Medzhitov, R.; Janeway, C.A. Decoding the patterns of self and nonself by the innate immune system. Science 2002, 296, 298–300. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Weber, A.N.; Atilano, M.L.; Filipe, S.R.; Gay, N.J.; Ligoxygakis, P. Sensing of gram-positive bacteria in drosophila: Gnbp1 is needed to process and present peptidoglycan to pgrp-sa. EMBO J. 2006, 25, 5005–5014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gobert, V.; Gottar, M.; Matskevich, A.A.; Rutschmann, S.; Royet, J.; Belvin, M.; Hoffmann, J.A.; Ferrandon, D. Dual activation of the drosophila toll pathway by two pattern recognition receptors. Science 2003, 302, 2126–2130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roh, K.-B.; Kim, C.-H.; Lee, H.; Kwon, H.-M.; Park, J.-W.; Ryu, J.-H.; Kurokawa, K.; Ha, N.-C.; Lee, W.-J.; Lemaitre, B. Proteolytic cascade for the activation of the insect toll pathway induced by the fungal cell wall component. J. Biol. Chem. 2009, 284, 19474–19481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, I.-H.; Chosa, N.; Kim, S.-H.; Nam, H.-J.; Lemaitre, B.; Ochiai, M.; Kambris, Z.; Brun, S.; Hashimoto, C.; Ashida, M. A spätzle-processing enzyme required for toll signaling activation in drosophila innate immunity. Dev. Cell 2006, 10, 45–55. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.-H.; Kim, S.-J.; Kan, H.; Kwon, H.-M.; Roh, K.-B.; Jiang, R.; Yang, Y.; Park, J.-W.; Lee, H.-H.; Ha, N.-C. A three-step proteolytic cascade mediates the activation of the peptidoglycan-induced toll pathway in an insect. J. Biol. Chem. 2008, 283, 7599–7607. [Google Scholar] [CrossRef] [Green Version]
- McDonald, C.; Inohara, N.; Nuñez, G. Peptidoglycan signaling in innate immunity and inflammatory disease. J. Biol. Chem. 2005, 280, 20177–20180. [Google Scholar] [CrossRef] [Green Version]
- Sasabe, J.; Suzuki, M. Emerging role of d-amino acid metabolism in the innate defense. Front. Microbiol. 2018, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Liu, S.A.; Mou, H.; Ma, Y.; Li, M.; Hu, X. Characterization of lipopeptide biosurfactants produced by bacillus licheniformis mb01 from marine sediments. Front. Microbiol. 2017, 8, 871. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Mawgoud, A.M.; Lépine, F.; Déziel, E. Rhamnolipids: Diversity of structures, microbial origins and roles. Appl. Microbiol. Biotechnol. 2010, 86, 1323–1336. [Google Scholar] [CrossRef] [Green Version]
- Ndlovu, T.; Rautenbach, M.; Vosloo, J.A.; Khan, S.; Khan, W. Characterisation and antimicrobial activity of biosurfactant extracts produced by bacillus amyloliquefaciens and pseudomonas aeruginosa isolated from a wastewater treatment plant. AMB Express 2017, 7, 108. [Google Scholar] [CrossRef] [Green Version]
- Abalos, A.; Pinazo, A.; Infante, M.; Casals, M.; Garcia, F.; Manresa, A. Physicochemical and antimicrobial properties of new rhamnolipids produced by pseudomonas a eruginosa at10 from soybean oil refinery wastes. Langmuir 2001, 17, 1367–1371. [Google Scholar] [CrossRef]
- Chen, W.-C.; Juang, R.-S.; Wei, Y.-H. Applications of a lipopeptide biosurfactant, surfactin, produced by microorganisms. Biochem. Eng. J. 2015, 103, 158–169. [Google Scholar] [CrossRef]
- Altincicek, B.; Vilcinskas, A. Metamorphosis and collagen-iv-fragments stimulate innate immune response in the greater wax moth, galleria mellonella. Dev. Comp. Immunol. 2006, 30, 1108–1118. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.-S.; Lee, J.; Kim, Y.-J.; Bang, H.-S.; Yun, E.-Y.; Kim, S.-R.; Suh, H.-J.; Kang, B.-R.; Nam, S.-H.; Jeon, J.-P. Isolation and characterization of a defensin-like peptide (coprisin) from the dung beetle, copris tripartitus. Int. J. Pept. 2009, 2009, 136284. [Google Scholar] [CrossRef]
- Jozefiak, A.; Engberg, R.M. Insect proteins as a potential source of antimicrobial peptides in livestock production. A review. J. Anim. Feed Sci. 2017, 26, 87–99. [Google Scholar] [CrossRef]
- Chae, J.-H.; Kurokawa, K.; So, Y.-I.; Hwang, H.O.; Kim, M.-S.; Park, J.-W.; Jo, Y.-H.; Lee, Y.S.; Lee, B.L. Purification and characterization of tenecin 4, a new anti-gram-negative bacterial peptide, from the beetle tenebrio molitor. Dev. Comp. Immunol. 2012, 36, 540–546. [Google Scholar] [CrossRef]
- Kim, D.-H.; Lee, Y.T.; Lee, Y.J.; Chung, J.H.; Lee, B.L.; Choi, B.S.; Lee, Y. Bacterial expression of tenecin 3, an insect antifungal protein isolated from tenebrio molitor, and its efficient purification. Mol. Cells (Springer Sci. Bus. Media BV) 1998, 8, 8. [Google Scholar]
- Lee, K.H.; Hong, S.Y.; Oh, J.E.; Kwon, M.y.; Yoon, J.H.; Lee, J.h.; Lee, B.L.; Moon, H.M. Identification and characterization of the antimicrobial peptide corresponding to c-terminal beta-sheet domain of tenecin 1, an antibacterial protein of larvae of tenebrio molitor. Biochem. J. 1998, 334, 99. [Google Scholar] [CrossRef]
- Dobson, A.J.; Johnston, P.R.; Vilcinskas, A.; Rolff, J. Identification of immunological expressed sequence tags in the mealworm beetle tenebrio molitor. J. Insect Physiol. 2012, 58, 1556–1561. [Google Scholar] [CrossRef]
- Zhu, S. Discovery of six families of fungal defensin-like peptides provides insights into origin and evolution of the csαβ defensins. Mol. Immunol. 2008, 45, 828–838. [Google Scholar] [CrossRef]
- Zhu, J.-Y.; Wu, G.-X.; Zhang, Z. Upregulation of coleoptericin transcription in tenebrio molitor parasitized by scleroderma guani. J. Asia Pac. Entomol. 2014, 17, 339–342. [Google Scholar] [CrossRef]
- Cheng, T.; Zhao, P.; Liu, C.; Xu, P.; Gao, Z.; Xia, Q.; Xiang, Z. Structures, regulatory regions, and inductive expression patterns of antimicrobial peptide genes in the silkworm bombyx mori. Genomics 2006, 87, 356–365. [Google Scholar] [CrossRef] [PubMed]
- Rozgonyi, F.; Szabo, D.; Kocsis, B.; Ostorhazi, E.; Abbadessa, G.; Cassone, M.; Wade, J.; Otvos, L., Jr. The antibacterial effect of a proline-rich antibacterial peptide a3-apo. Curr. Med. Chem. 2009, 16, 3996–4002. [Google Scholar] [CrossRef] [PubMed]
- Ongey, E.L.; Pflugmacher, S.; Neubauer, P. Bioinspired designs, molecular premise and tools for evaluating the ecological importance of antimicrobial peptides. Pharmaceuticals 2018, 11, 68. [Google Scholar] [CrossRef] [Green Version]
- Patocka, J.; Nepovimova, E.; Klimova, B.; Wu, Q.; Kuca, K. Antimicrobial peptides: Amphibian host defense peptides. Curr. Med. Chem. 2019, 26, 5924–5946. [Google Scholar] [CrossRef]
- Giri, S.S.; Sen, S.S.; Jun, J.W.; Sukumaran, V.; Park, S.C. Role of bacillus licheniformis vs16-derived biosurfactant in mediating immune responses in carp rohu and its application to the food industry. Front. Microbiol. 2017, 8, 514. [Google Scholar] [CrossRef] [Green Version]
- Revathi, J.A. Immunostimulation by rhamnolipid biosurfactant from pseudomonas putida in labeo rohita. Int. J. Adv. Res. Biol. Sci. 2016, 3, 179–183. [Google Scholar]
- Bauer, J.; Brandenburg, K.; Zähringer, U.; Rademann, J. Chemical synthesis of a glycolipid library by a solid-phase strategy allows elucidation of the structural specificity of immunostimulation by rhamnolipids. Chem. Eur. J. 2006, 12, 7116–7124. [Google Scholar] [CrossRef]
- Li, H.; Anuwongcharoen, N.; Malik, A.A.; Prachayasittikul, V.; Wikberg, J.E.S.; Nantasenamat, C. Roles of d-amino acids on the bioactivity of host defense peptides. Int. J. Mol. Sci. 2016, 17, 1023. [Google Scholar] [CrossRef] [Green Version]
- Mondol, M.; Shin, H.; Islam, M. Diversity of secondary metabolites from marine bacillus species: Chemistry and biological activity. Mar. Drugs 2013, 11, 2846–2872. [Google Scholar] [CrossRef] [Green Version]
- Ali Mohammadie Kojour, M.; Han, Y.S.; Jo, Y.H. An overview of insect innate immunity. Entomol. Res. 2020, 50, 282–291. [Google Scholar] [CrossRef]
- Nakata, T.; Yasuda, M.; Fujita, M.; Kataoka, H.; Kiura, K.; Sano, H.; Shibata, K. Cd14 directly binds to triacylated lipopeptides and facilitates recognition of the lipopeptides by the receptor complex of toll-like receptors 2 and 1 without binding to the complex. Cell. Microbiol. 2006, 8, 1899–1909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buwitt-Beckmann, U.; Heine, H.; Wiesmüller, K.-H.; Jung, G.; Brock, R.; Akira, S.; Ulmer, A.J. Tlr1-and tlr6-independent recognition of bacterial lipopeptides. J. Biol. Chem. 2006, 281, 9049–9057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.Y.; Nan, X.; Jin, M.S.; Youn, S.-J.; Ryu, Y.H.; Mah, S.; Han, S.H.; Lee, H.; Paik, S.-G.; Lee, J.-O. Recognition of lipopeptide patterns by toll-like receptor 2-toll-like receptor 6 heterodimer. Immunity 2009, 31, 873–884. [Google Scholar] [CrossRef] [Green Version]
- Wright, S.D.; Ramos, R.A.; Tobias, P.S.; Ulevitch, R.J.; Mathison, J.C. Cd14, a receptor for complexes of lipopolysaccharide (lps) and lps binding protein. Science 1990, 249, 1431–1433. [Google Scholar] [CrossRef]
- Zaman, M.; Toth, I. Immunostimulation by synthetic lipopeptide-based vaccine candidates: Structure-activity relationships. Front. Immunol. 2013, 4, 318. [Google Scholar] [CrossRef] [Green Version]
- Weber, A.N.; Gangloff, M.; Moncrieffe, M.C.; Hyvert, Y.; Imler, J.-L.; Gay, N.J. Role of the spätzle pro-domain in the generation of an active toll receptor ligand. J. Biol. Chem. 2007, 282, 13522–13531. [Google Scholar] [CrossRef] [Green Version]
- Weber, A.N.; Tauszig-Delamasure, S.; Hoffmann, J.A.; Lelièvre, E.; Gascan, H.; Ray, K.P.; Morse, M.A.; Imler, J.-L.; Gay, N.J. Binding of the drosophila cytokine spätzle to toll is direct and establishes signaling. Nat. Immunol. 2003, 4, 794. [Google Scholar] [CrossRef]
- Yu, Y.; Park, J.W.; Kwon, H.M.; Hwang, H.O.; Jang, I.H.; Masuda, A.; Kurokawa, K.; Nakayama, H.; Lee, W.J.; Dohmae, N.; et al. Diversity of innate immune recognition mechanism for bacterial polymeric meso-diaminopimelic acid-type peptidoglycan in insects. J. Biol. Chem. 2010, 285, 32937–32945. [Google Scholar] [CrossRef] [Green Version]
- Jo, Y.H.; Park, S.; Park, K.B.; Noh, M.Y.; Cho, J.H.; Ko, H.J.; Kim, C.E.; Patnaik, B.B.; Kim, J.; Won, R. In silico identification, characterization and expression analysis of attacin gene family in response to bacterial and fungal pathogens in tenebrio molitor. Entomol. Res. 2018, 48, 45–54. [Google Scholar] [CrossRef]
- Casteels, P.; Ampe, C.; Jacobs, F.; Tempst, P. Functional and chemical characterization of hymenoptaecin, an antibacterial polypeptide that is infection-inducible in the honeybee (apis mellifera). J. Biol. Chem. 1993, 268, 7044–7054. [Google Scholar] [PubMed]
- Carlsson, A.; Engström, P.; Palva, E.T.; Bennich, H. Attacin, an antibacterial protein from hyalophora cecropia, inhibits synthesis of outer membrane proteins in escherichia coli by interfering with omp gene transcription. Infect. Immun. 1991, 59, 3040–3045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edosa, T.T.; Jo, Y.H.; Keshavarz, M.; Bae, Y.M.; Kim, D.H.; Lee, Y.S.; Han, Y.S. Tmspz6 is essential for regulating the immune response to escherichia coli and staphylococcus aureus infection in tenebrio molitor. Insects 2020, 11, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edosa, T.T.; Jo, Y.H.; Keshavarz, M.; Park, K.B.; Cho, J.H.; Bae, Y.M.; Kim, B.; Lee, Y.S.; Han, Y.S. Tmatg6 plays an important role in anti-microbial defense against listeria monocytogenes in the mealworm, tenebrio molitor. Int. J. Mol. Sci. 2020, 21, 1232. [Google Scholar] [CrossRef] [Green Version]
- Keshavarz, M.; Jo, Y.H.; Park, K.B.; Ko, H.J.; Edosa, T.T.; Lee, Y.S.; Han, Y.S. Tm dorx2 positively regulates antimicrobial peptides in tenebrio molitor gut, fat body, and hemocytes in response to bacterial and fungal infection. Sci. Rep. 2019, 9, 1–19. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biosurfactants. | Type | Biological Source | Type | Purity |
|---|---|---|---|---|
| Bacitracin | cyclic lipopeptide | Bacillus licheniformis | Gram-positive | 90% |
| Fengycin | lipopeptide complex | Bacillus subtilis | Gram-positive | >90% |
| Iturin A | cyclic lipopeptide | Bacillus subtilis | Gram-positive | >95% |
| Rhamnolipid | Glycolipids | Pseudomonas aeruginosa | Gram-negative | >95% |
| Name | Primer Sequences |
|---|---|
| TmTencin-1_Fw | 5′-CAGCTGAAGAAATCGAACAAGG-3′ |
| TmTencin-1_Rv | 5′-CAGACCCTCTTTCCGTTACAGT-3′ |
| TmTencin-2_Fw | 5′-CAGCAAAACGGAGGATGGTC-3′ |
| TmTencin-2_Rv | 5′-CGTTGAAATCGTGATCTTGTCC-3′ |
| TmTencin-3_Fw | 5′-GATTTGCTTGATTCTGGTGGTC-3′ |
| TmTencin-3_Rv | 5′-CTGATGGCCTCCTAAATGTCC-3′ |
| TmTencin-4_Fw | 5′-GGACATTGAAGATCCAGGAAAG-3′ |
| TmTencin-4_Rv | 5′-CGGTGTTCCTTATGTAGAGCTG-3′ |
| TmDefensin-1_Fw | 5′-AAATCGAACAAGGCCAACAC-3′ |
| TmDefencin-1_Rv | 5′-GCAAATGCAGACCCTCTTTC-3′ |
| TmDefencin-2_Fw | 5′-GGGATGCCTCATGAAGATGTAG-3′ |
| TmDefencin-2_Rv | 5′-CCAATGCAAACACATTCGTC-3′ |
| TmColoptericin-1_Fw | 5′-GGACAGAATGGTGGATGGTC-3′ |
| TmColoptericin-1_Rv | 5′-CTCCAACATTCCAGGTAGGC-3 |
| TmColoptericin-2_Fw | 5′-GGACGGTTCTGATCTTCTTGAT-3′ |
| TmColoptericin-2_Rv | 5′-CAGCTGTTTGTTTGTTCTCGTC-3′ |
| TmAttacin-1a_Fw | 5′-GAAACGAAATGGAAGGTGGA-3′ |
| TmAttacin-1a_Rv | 5′-TGCTTCGGCAGACAATACAG-3′ |
| TmAttacin-1b_Fw | 5′-GAGCTGTGAATGCAGGACAA-3′ |
| TmAttacin-1b_Rv | 5′-CCCTCTGATGAAACCTCCAA-3′ |
| TmAttacin-2_Fw | 5′-AACTGGGATATTCGCACGTC-3′ |
| TmAttacin-2_Rv | 5′-CCCTCCGAAATGTCTGTTGT-3 |
| TmCecropin-2_Fw | 5′-TACTAGCAGCGCCAAAACCT-3′ |
| TmCecropin-2_Rv | 5′-CTGGAACATTAGGCGGAGAA-3′ |
| TmThaumatin-like protein-1_Fw | 5′-CTCAAAGGACACGCAGGACT-3′ |
| TmThaumatin-like protein-1_Rv | 5′-ACTTTGAGCTTCTCGGGACA-3′ |
| TmThaumatin-like protein-2_Fw | 5′-CCGTCTGGCTAGGAGTTCTG-3′ |
| TmThaumatin-like protein-2_Rv | 5′-ACTCCTCCAGCTCCGTTACA-3′ |
| TmL27a_qPCR_Fw | 5′-TCATCCTGAAGGCAAAGCTCCAGT-3′ |
| TmL27a_qPCR_Rv | 5′-AGGTTGGTTAGGCAGGCACCTTTA-3′ |
| TmSpz4-qPCR-Fw | 5′-GGCGATGCTCTTCCAGGAC-3′ |
| TmSpz4-qPCR-Rv | 5′-CGCGTTCACTCCTTTCATTTGG-3′ |
| TmSpz4-T7-Fw | 5′-TAATACGACTCACTATAGGGT CCAGATGTACTGTCGCGATG-3′ |
| TmSpz4-T7-Rv | 5′-TAATACGACTCACTATAGGGT TTCCTTCTGTACCAGTCGGG-3′ |
| TmSpz6-qPCR-Fw | 5′-GCACAACTCCAAGACGACCT-3′ |
| TmSpz6-qPCR-Rv | 5′-TCTCTTCACCCGATCGTTGC-3′ |
| TmSpz6-T7-Fw | 5′-TAATACGACTCACTATAGGGT ACCGCGCAAGAGAGTAAAAA-3′ |
| TmSpz6-T7-Rv | 5′-TAATACGACTCACTATAGGGT ACGTATCTCCACACCCCTTG-3′ |
| TmSpz3-qPCR-Fw | 5′-TCTCAACAACGGGACCTTCG-3′ |
| TmSpz3-qPCR-Rv | 5′-GGGACGCCCCGTATGTATTC-3′ |
| TmSPZ3-T7-270bp-Fw | 5′-TAATACGACTCACTATAGGGT CGAGAACAAGGCACTGATGA-3′ |
| TmSPZ3-T7-270bp-Rv | 5′-TAATACGACTCACTATAGGGT GCGGTGCCATTTGTACTTCT-3′ |
| TmSPZ-like-qPCR-Fw | 5′-CAGTTGAGGGTGCCTGTTCA-3′ |
| TmSPZ-like-qPCR-Rv | 5′-TTGTTGGCATCGTCCCTTGA-3′ |
| TmSpz-like-T7-Fw | 5′-TAATACGACTCACTATAGGGT ATGTTCCCAAAATCAACGGA-3′ |
| TmSpz-like-T7-Rv | 5′-TAATACGACTCACTATAGGGT AATCACACGCAGATCCTTCC-3′ |
| dsEGFP_Fw | 5′-TAATACGACTCACTATAGGGT CGTAAACGGCCACAAGTTC-3′ |
| dsEGFP_Rv | 5′-TAATACGACTCACTATAGGGT TGCTCAGGTAGTGTTGTCG-3′ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Edosa, T.T.; Jo, Y.H.; Keshavarz, M.; Kim, I.S.; Han, Y.S. Biosurfactants Induce Antimicrobial Peptide Production through the Activation of TmSpatzles in Tenebrio molitor. Int. J. Mol. Sci. 2020, 21, 6090. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176090
Edosa TT, Jo YH, Keshavarz M, Kim IS, Han YS. Biosurfactants Induce Antimicrobial Peptide Production through the Activation of TmSpatzles in Tenebrio molitor. International Journal of Molecular Sciences. 2020; 21(17):6090. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176090
Chicago/Turabian StyleEdosa, Tariku Tesfaye, Yong Hun Jo, Maryam Keshavarz, In Seon Kim, and Yeon Soo Han. 2020. "Biosurfactants Induce Antimicrobial Peptide Production through the Activation of TmSpatzles in Tenebrio molitor" International Journal of Molecular Sciences 21, no. 17: 6090. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176090