Zinc Deficiency Disturbs Mucin Expression, O-Glycosylation and Secretion by Intestinal Goblet Cells

 and
and 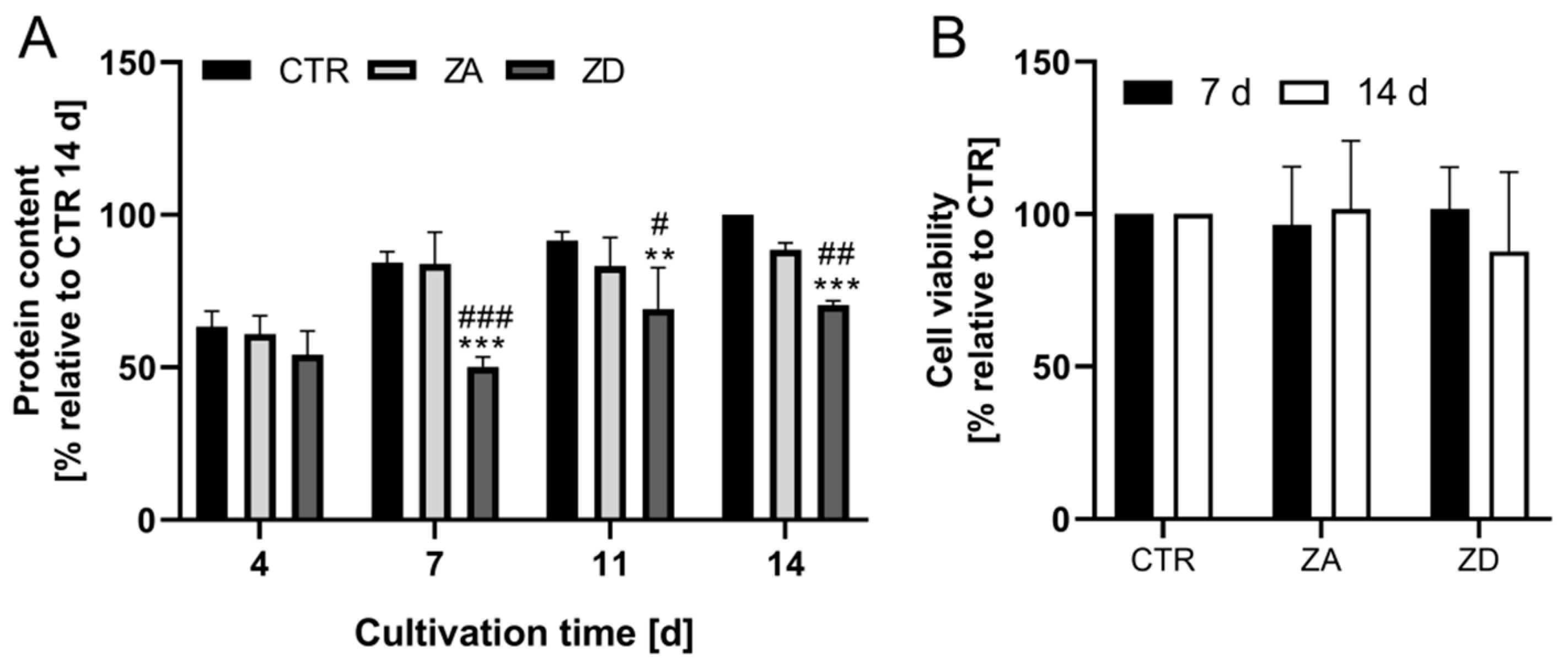{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
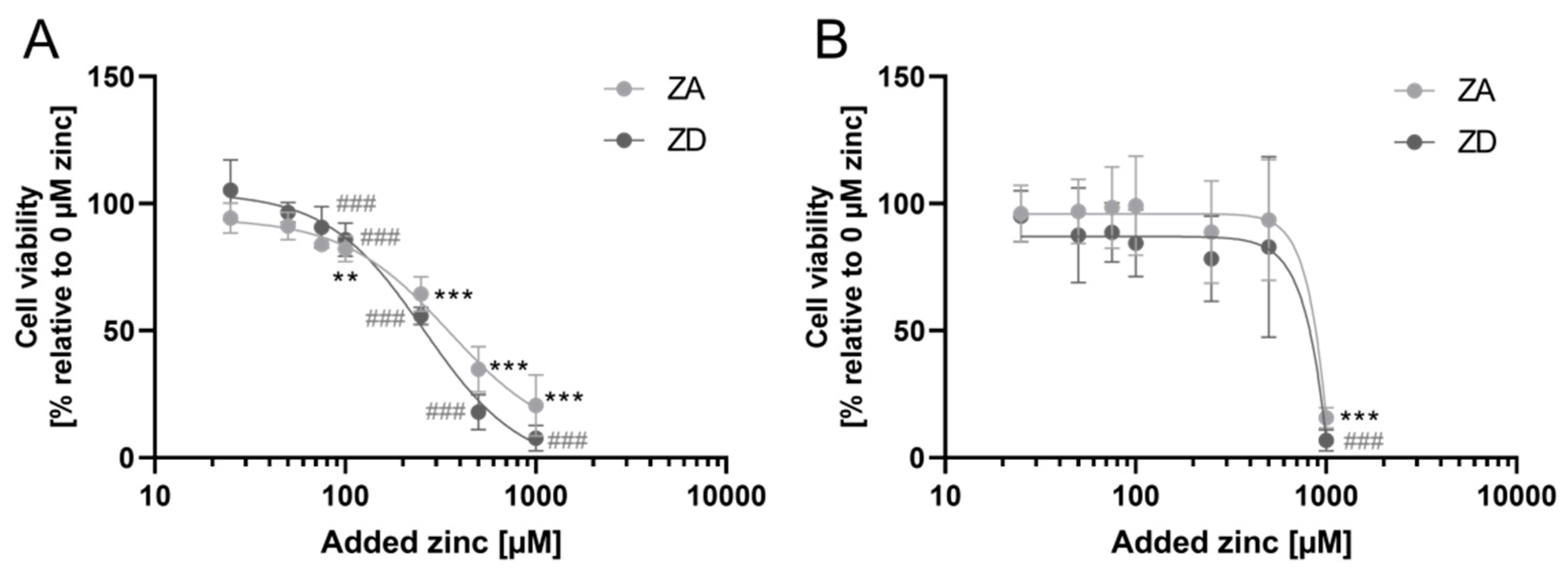
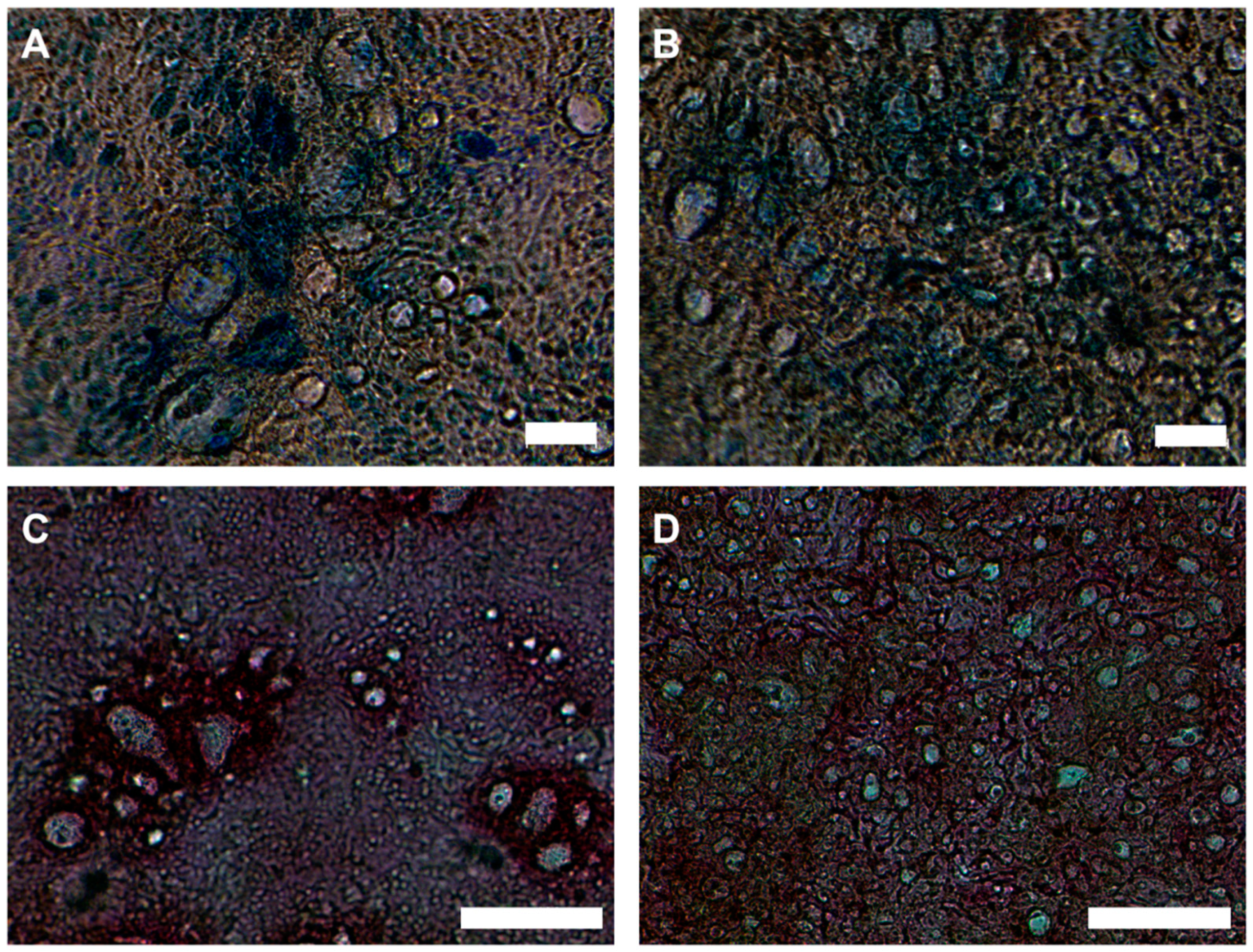
2.1. Characterization of Zinc-Deficient Goblet Cells
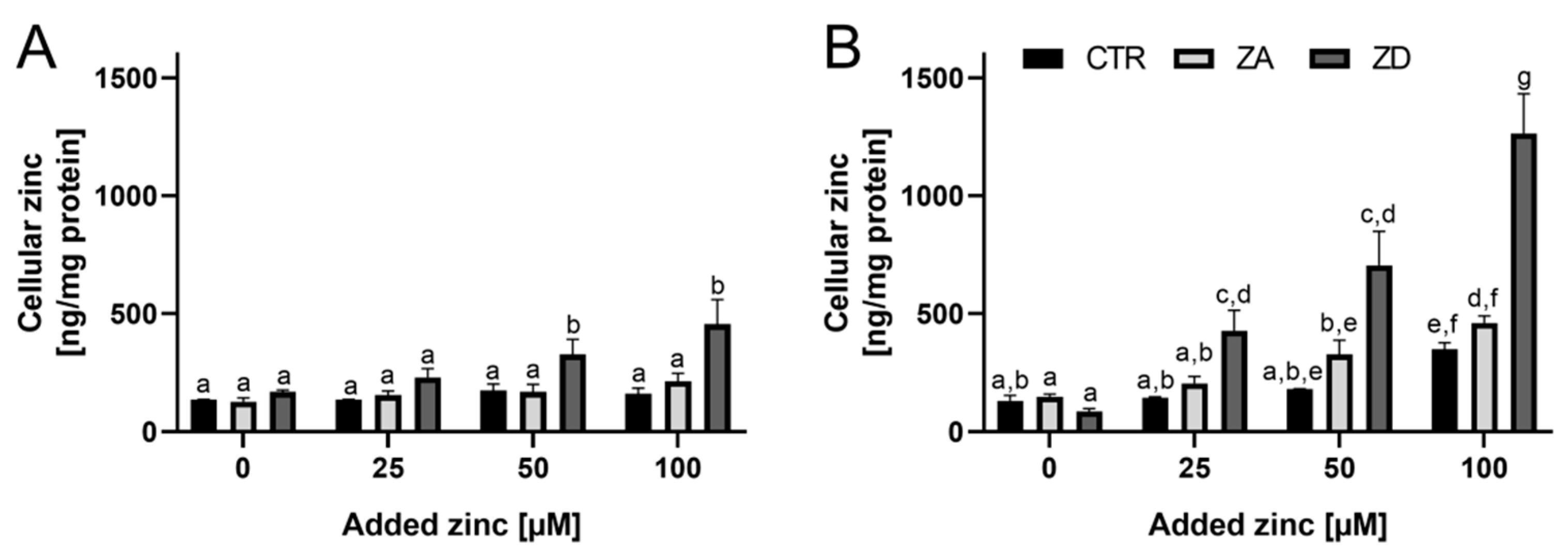
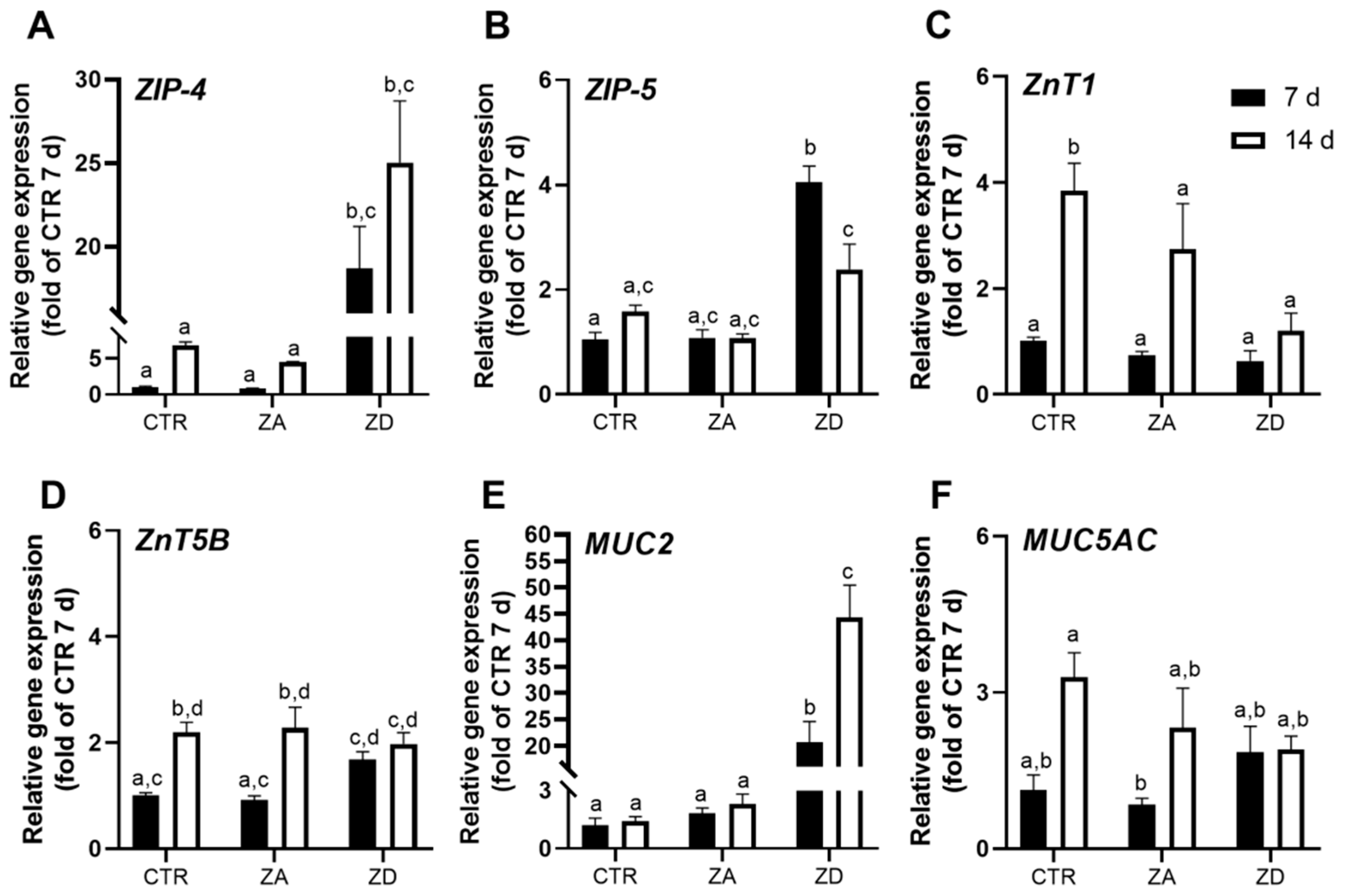
2.2. Zinc Homeostasis of Goblet Cells
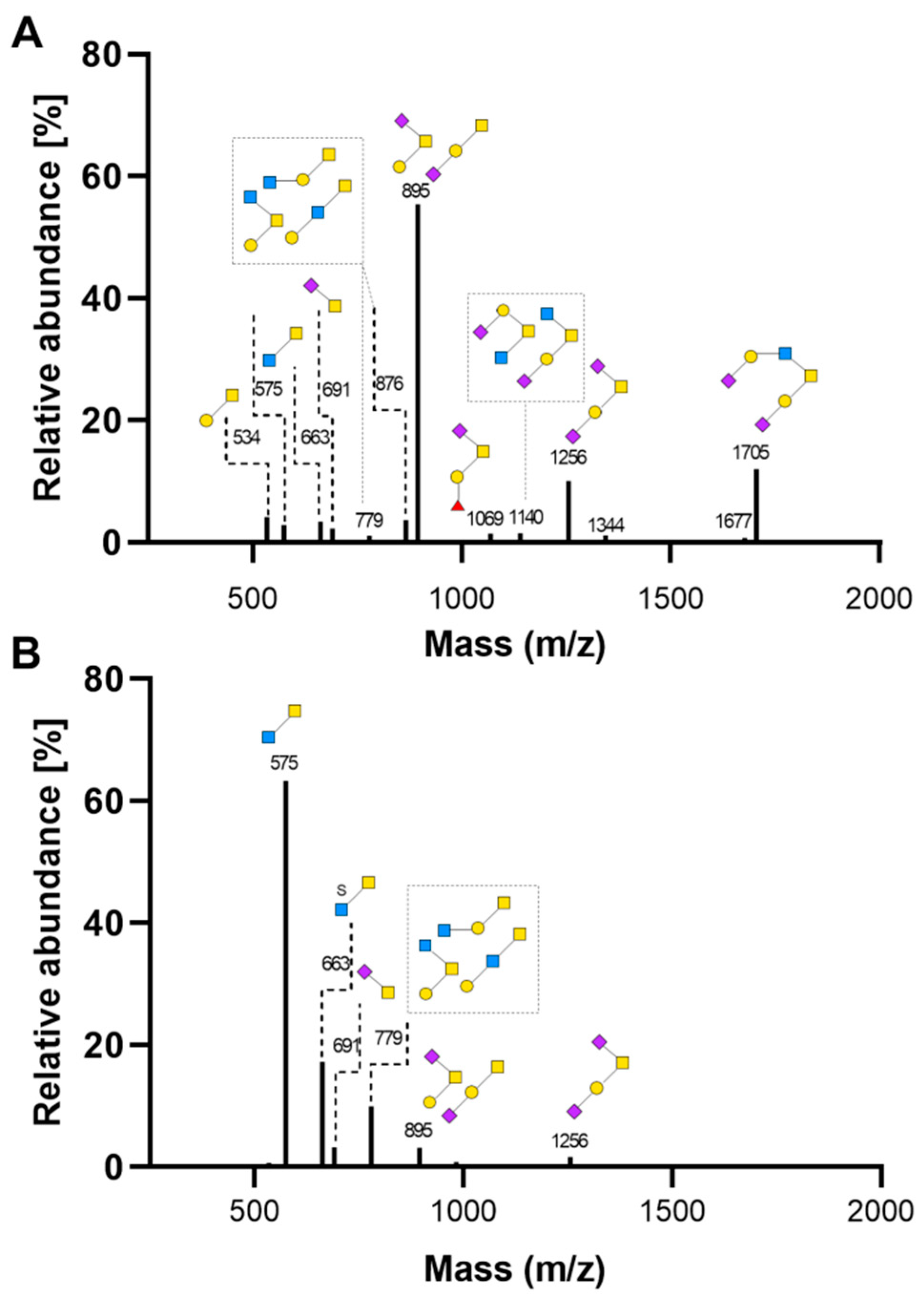
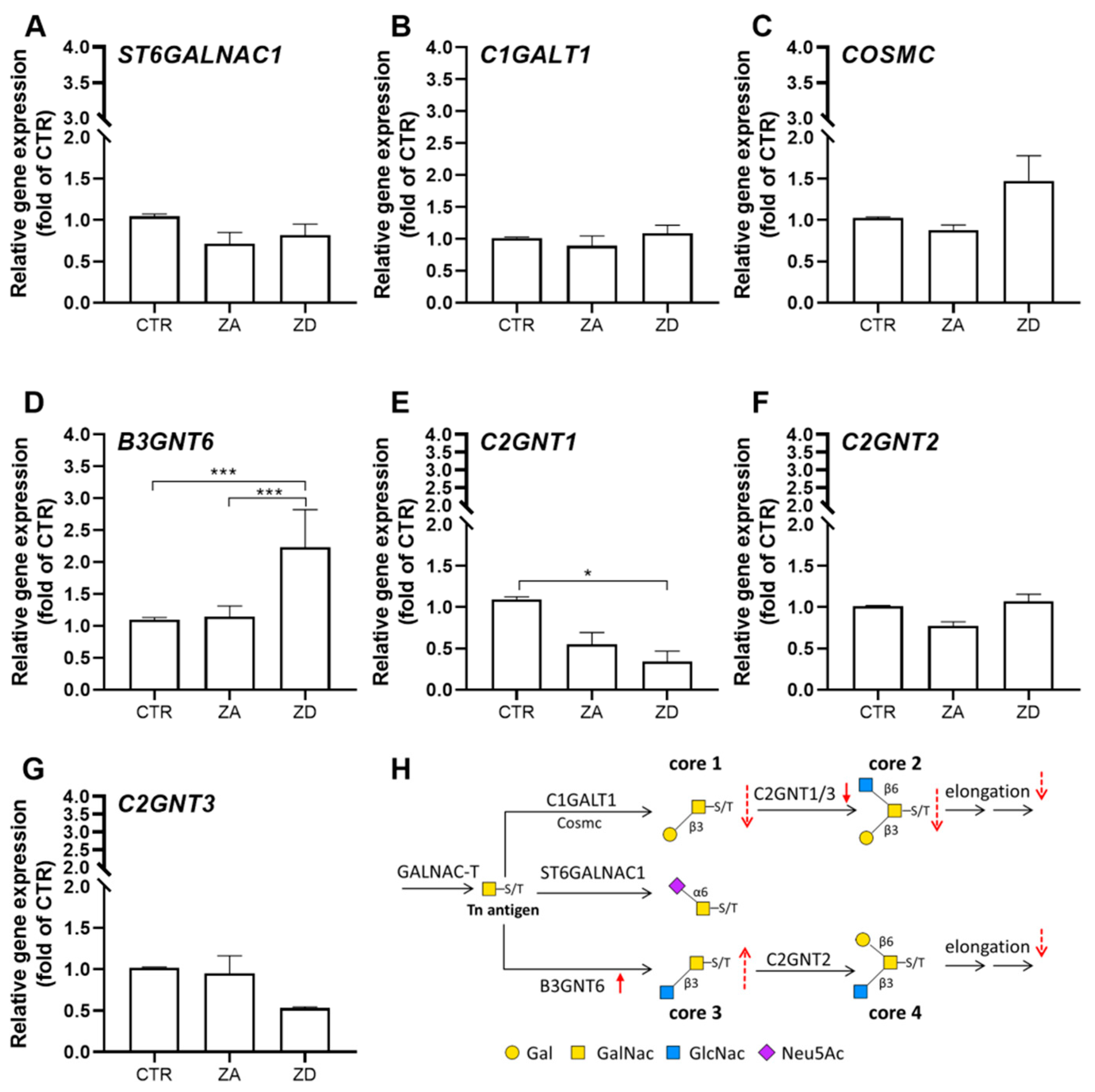
2.3. Intestinal Mucin Synthesis and Secretion during Zinc Deficiency
3. Materials and Methods
3.1. Materials
3.2. Cells and Cell Culture
3.3. Preparation of Cell Culture Medium
3.4. Cellular Protein Content
3.5. Cell Viability
3.6. Cellular Zinc Uptake
3.7. Gene Expression
3.8. Histological Staining of Mucins
3.9. Analysis of O-glycosylation of Secreted Mucins by MALDI-TOF MS
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AB | Alcian blue |
| ANOVA | Analysis of variance |
| BCA | Bicinchoninic acid |
| B3GNT6 | β-1,3-N-acetylglucosaminyltransferase |
| C1GALT1 | Core 1 glycoprotein-N-acetylgalactosamine 3-β-galactosyltransferase |
| CF | Cystic fibrosis |
| COSMC | C1GALT1-specific molecular chaperone |
| DMEM | Dulbecco’s Modified Eagles Medium |
| ECACC | European Collection of Authenticated Cell Culture |
| FAAS | Flame atomic absorption spectrometry |
| FCS | Fetal calf serum |
| GalNAc | N-acetyl-galactosamine |
| GIT | Gastrointestinal tract |
| GNT | β-1,6-N-acetylglucosaminyltransferase |
| MALDI-TOF | Matrix-assisted laser desorption/ionization time-of-flight |
| NeuAc | N-acetylneuraminic acid |
| ICP–MS | Inductively coupled plasma mass spectrometry |
| LOQ | Limit of quantitation |
| NEAA | Non-essential amino acids |
| PAS | Periodic acid Schiff |
| GalNAc-Ts | UDP-GalNAc:polypeptide GalNAc transferases |
| ST6GALNAC1 | ST6 N-Acetylgalactosaminide Alpha-2,6-Sialyltransferase 1 |
| TPEN | N,N,N′,N′-Tetrakis(2-pyridylmethyl)ethylenediamine |
| ZA | Zinc-adequate |
| ZD | Zinc-deficient |
| ZnT | Zinc transporter |
| ZIP | Zrt Irt-like transporter |
References
- Andreini, C.; Banci, L.; Bertini, I.; Rosato, A. Counting the zinc-proteins encoded in the human genome. J. Proteome Res. 2005, 5, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Maares, M.; Haase, H. A guide to human zinc absorption: General overview and recent advances of in vitro intestinal models. Nutrients 2020, 12, 43. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization/Food and Agricultural Organization. Vitamin and Mineral Requirements in Human Nutrition, 2nd ed.; World Health Organization: Geneva, Switzerland, 2004. [Google Scholar]
- Broadley, D.B.K.; Edward, J.M.J.; Ander, E.L.; Michael, J.W.; Scott, D.Y.; Sue, W.; Martin, R. Dietary calcium and zinc deficiency risks are decreasing but remain prevalent. Sci. Rep. 2015, 5, 10974. [Google Scholar]
- Gibson, R.S.; King, J.C.; Lowe, N. A review of dietary zinc recommendations. Food Nutr. Bull. 2016, 37, 443–460. [Google Scholar] [CrossRef] [Green Version]
- Aggett, P.J. Severe zinc deficiency. In Zinc in Human Biology; Mills, C.F., Ed.; Springer: London, UK, 1989; pp. 259–279. [Google Scholar]
- Southon, S.; Gee, J.M.; Johnson, I.T. Hexose transport and mucosal morphology in the small intestine of the zinc-deficient rat. Br. J. Nutr. 1984, 52, 371–380. [Google Scholar] [CrossRef] [Green Version]
- Southon, S.; Gee, J.M.; Bayliss, C.E.; Wyatt, G.M.; Horn, N.; Johnson, I.T. Intestinal microflora, morphology and enzyme activity in zinc-deficient and zn-supplemented rats. Br. J. Nutr. 1986, 55, 603–611. [Google Scholar] [CrossRef]
- Elmes, M.E.; Jones, J.G. Ultrastructural changes in the small intestine of zinc deficient rats. J. Pathol. 1980, 130, 37–43. [Google Scholar] [CrossRef]
- Finamore, A.; Massimi, M.; Conti Devirgiliis, L.; Mengheri, E. Zinc deficiency induces membrane barrier damage and increases neutrophil transmigration in caco-2 cells. J. Nutr. 2008, 138, 1664–1670. [Google Scholar] [CrossRef] [Green Version]
- Southon, S.; Livesey, G.; Gee, J.M.; Johnson, I.T. Intestinal cellular proliferation and protein synthesis in zinc-deficient rats. Br. J. Nutr. 1985, 53, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Grandjean, C.J.; Antonson, D.L.; Vanderhoof, J.A. Effects of short-term isolated zinc deficiency on intestinal growth and activities of several brush border enzymes in weaning rats. Pediatric Res. 1985, 19, 1333–1336. [Google Scholar] [CrossRef] [PubMed]
- Quarterman, J.; Humphries, W.R.; Morrison, J.; Jackson, F.A. The effect of zinc deficiency on intestinal and salivary mucins. Biochem. Soc. Trans. 1973, 1, 101. [Google Scholar] [CrossRef]
- Quarterman, J.; Jackson, F.A.; Morrison, J.N. The effect of zinc deficiency on sheep intestinal mucin. Life Sci. 1976, 19, 979–986. [Google Scholar] [CrossRef]
- Leal, J.; Smyth, H.D.C.; Ghosh, D. Physicochemical properties of mucus and their impact on transmucosal drug delivery. Int. J. Pharm. 2017, 532, 555–572. [Google Scholar] [CrossRef]
- Johansson, M.E.V.; Hansson, G.C. Immunological aspects of intestinal mucus and mucins. Nat. Rev. Immunol. 2016, 16, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Maares, M.; Keil, C.; Koza, J.; Straubing, S.; Schwerdtle, T.; Haase, H. In vitro studies on zinc binding and buffering by intestinal mucins. Int. J. Mol. Sci. 2018, 19, 2662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansson, M.E.; Hansson, G.C. Mucus and the goblet cell. Dig. Dis. 2013, 31, 305–309. [Google Scholar] [CrossRef] [Green Version]
- Tailford, L.; Crost, E.; Kavanaugh, D.; Juge, N. Mucin glycan foraging in the human gut microbiome. Front. Genet. 2015, 6, 81–99. [Google Scholar] [CrossRef] [Green Version]
- Jin, C.; Kenny, D.T.; Skoog, E.C.; Padra, M.; Adamczyk, B.; Vitizeva, V.; Thorell, A.; Venkatakrishnan, V.; Lindén, S.K.; Karlsson, N.G. Structural diversity of human gastric mucin glycans. Mol. Cell. Proteom. 2017, 16, 743–758. [Google Scholar] [CrossRef]
- Hennigar, S.R.; McClung, J.P. Zinc transport in the mammalian intestine. Compr. Physiol. 2018, 9, 59–74. [Google Scholar]
- Mayer, L.S.; Uciechowski, P.; Meyer, S.; Schwerdtle, T.; Rink, L.; Haase, H. Differential impact of zinc deficiency on phagocytosis, oxidative burst, and production of pro-inflammatory cytokines by human monocytes. Metallomics 2014, 6, 1288–1295. [Google Scholar] [CrossRef] [Green Version]
- Messer, H.H.; Murray, E.J.; Goebel, N.K. Removal of trace metals from culture media and sera for in vitro deficiency studies. J. Nutr. 1982, 112, 652–657. [Google Scholar] [CrossRef]
- Hennebicq-Reig, S.; Tetaert, D.; Soudan, B.; Kim, I.; Huet, G.; Briand, G.; Richet, C.; Demeyer, D.; Degand, P. O-glycosylation and cellular differentiation in a subpopulation of mucin-secreting ht-29 cell line. Exp. Cell Res. 1997, 235, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Lesuffleur, T.; Porchet, N.; Aubert, J.P.; Swallow, D.; Gum, J.R.; Kim, Y.S.; Real, F.X.; Zweibaum, A. Differential expression of the human mucin genes muc1 to muc5 in relation to growth and differentiation of different mucus-secreting ht-29 cell subpopulations. J. Cell Sci. 1993, 106, 771–783. [Google Scholar] [PubMed]
- John, S.; Briatka, T.; Rudolf, E. Diverse sensitivity of cells representing various stages of colon carcinogenesis to increased extracellular zinc: Implications for zinc chemoprevention. Oncol. Rep. 2011, 25, 769–780. [Google Scholar]
- Jou, M.Y.; Philipps, A.F.; Kelleher, S.L.; Lonnerdal, B. Effects of zinc exposure on zinc transporter expression in human intestinal cells of varying maturity. J. Pediatric Gastroenterol. Nutr. 2010, 50, 587–595. [Google Scholar] [CrossRef]
- Liuzzi, J.P.; Guo, L.; Chang, S.M.; Cousins, R.J. Kruppel-like factor 4 regulates adaptive expression of the zinc transporter zip4 in mouse small intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G517–G523. [Google Scholar] [CrossRef] [Green Version]
- Liuzzi, J.P.; Bobo, J.A.; Lichten, L.A.; Samuelson, D.A.; Cousins, R.J. Responsive transporter genes within the murine intestinal-pancreatic axis form a basis of zinc homeostasis. Proc. Natl. Acad. Sci. USA 2004, 101, 14355–14360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMahon, R.J.; Cousins, R.J. Regulation of the zinc transporter znt-1 by dietary zinc. Proc. Natl. Acad. Sci. USA 1998, 95, 4841–4846. [Google Scholar] [CrossRef] [Green Version]
- Nishito, Y.; Kambe, T. Zinc transporter 1 (znt1) expression on the cell surface is elaborately controlled by cellular zinc levels. J. Biol. Chem. 2019, 294, 15686–15697. [Google Scholar] [CrossRef]
- Jou, M.Y.; Hall, A.G.; Philipps, A.F.; Kelleher, S.L.; Lonnerdal, B. Tissue-specific alterations in zinc transporter expression in intestine and liver reflect a threshold for homeostatic compensation during dietary zinc deficiency in weanling rats. J. Nutr. 2009, 139, 835–841. [Google Scholar] [CrossRef] [Green Version]
- Robbe, C.; Capon, C.; Coddeville, B.; Michalski, J.-C. Structural diversity and specific distribution of O-glycans in normal human mucins along the intestinal tract. Biochem. J. 2004, 384, 307–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyerholz, D.K.; Rodgers, J.; Castilow, E.M.; Varga, S.M. Alcian blue and pyronine y histochemical stains permit assessment of multiple parameters in pulmonary disease models. Vet. Pathol. 2009, 46, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Leteurtre, E.; Gouyer, V.; Rousseau, K.; Moreau, O.; Barbat, A.; Swallow, D.; Huet, G.; Lesuffleur, T. Differential mucin expression in colon carcinoma ht-29 clones with variable resistance to 5-fluorouracil and methotrexate. Biol. Cell 2004, 96, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Pieper, R.; Rieger, J.; Vahjen, W.; Davin, R.; Plendl, J.; Meyer, W.; Zentek, J. Effect of dietary zinc oxide on morphological characteristics, mucin composition and gene expression in the colon of weaned piglets. PLoS ONE 2014, 9, e91091. [Google Scholar] [CrossRef] [Green Version]
- De Lisle, R.C.; Borowitz, D. The cystic fibrosis intestine. Cold Spring Harb. Perspect. Med. 2013, 3, a009753. [Google Scholar] [CrossRef] [Green Version]
- Monge, M.F.E.; Barrado, E.; Vicente, C.A.; Del Rio, M.P.R.; de Miguelsanz, J.M.M. Zinc nutritional status in patients with cystic fibrosis. Nutrients 2019, 11, 150. [Google Scholar] [CrossRef] [Green Version]
- Kamei, S.; Fujikawa, H.; Nohara, H.; Ueno-Shuto, K.; Maruta, K.; Nakashima, R.; Kawakami, T.; Matsumoto, C.; Sakaguchi, Y.; Ono, T.; et al. Zinc deficiency via a splice switch in zinc importer zip2/slc39a2 causes cystic fibrosis-associated muc5ac hypersecretion in airway epithelial cells. EBioMedicine 2018, 27, 304–316. [Google Scholar] [CrossRef] [Green Version]
- International Zinc Nutrition Consultative Group; Brown, K.H.; Rivera, J.A.; Bhutta, Z.; Gibson, R.S.; King, J.C.; Lonnerdal, B.; Ruel, M.T.; Sandtrom, B.; Wasantwisut, E.; et al. International zinc nutrition consultative group (izincg) technical document #1. Assessment of the risk of zinc deficiency in populations and options for its control. Food Nutr. Bull. 2004, 25, S99–S203. [Google Scholar]
- Bennett, E.P.; Mandel, U.; Clausen, H.; Gerken, T.A.; Fritz, T.A.; Tabak, L.A. Control of mucin-type O-glycosylation: A classification of the polypeptide galnac-transferase gene family. Glycobiology 2012, 22, 736–756. [Google Scholar] [CrossRef] [Green Version]
- Ringot-Destrez, B.; D’Alessandro, Z.; Lacroix, J.M.; Mercier-Bonin, M.; Leonard, R.; Robbe-Masselot, C. A sensitive and rapid method to determin the adhesion capacity of probiotics and pathogenic microorganisms to human gastrointestinal mucins. Microorganisms 2018, 6, 49. [Google Scholar] [CrossRef] [Green Version]
- Huet, G.; Kim, I.; de Bolos, C.; Lo-Guidice, J.M.; Moreau, O.; Hemon, B.; Richet, C.; Delannoy, P.; Real, F.X.; Degand, P. Characterization of mucins and proteoglycans synthesized by a mucin-secreting ht-29 cell subpopulation. J. Cell Sci. 1995, 108 Pt 3, 1275–1285. [Google Scholar]
- Varki, A.; Cummings, R.D.; Aebi, M.; Packer, N.H.; Seeberger, P.H.; Esko, J.D.; Stanley, P.; Hart, G.; Darvill, A.; Kinoshita, T.; et al. Symbol nomenclature for graphical representations of glycans. Glycobiology 2015, 25, 1323–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrow, H.; Tam, B.; Duckworth, C.A.; Rhodes, J.M.; Yu, L.-G. Suppression of core 1 gal-transferase is associated with reduction of tf and reciprocal increase of tn, sialyl-tn and core 3 glycans in human colon cancer cells. PLoS ONE 2013, 8, e59792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ju, T.; Cummings, R.D. A unique molecular chaperone cosmc required for activity of the mammalian core 1 beta 3-galactosyltransferase. Proc. Natl. Acad. Sci. USA 2002, 99, 16613–16618. [Google Scholar] [CrossRef] [Green Version]
- Tran, D.T.; Ten Hagen, K.G. Mucin-type O-glycosylation during development. J. Biol. Chem. 2013, 288, 6921–6929. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.; Leon, F.; Rauth, S.; Batra, S.K.; Ponnusamy, M.P. A systematic review on the implications of O-linked glycan branching and truncating enzymes on cancer progression and metastasis. Cells 2020, 9, 446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomsson, K.A.; Holmen-Larsson, J.M.; Angstrom, J.; Johansson, M.E.; Xia, L.; Hansson, G.C. Detailed O-glycomics of the muc2 mucin from colon of wild-type, core 1- and core 3-transferase-deficient mice highlights differences compared with human muc2. Glycobiology 2012, 22, 1128–1139. [Google Scholar] [CrossRef] [PubMed]
- Neelamegham, S.; Liu, G. Systems glycobiology: Biochemical reaction networks regulating glycan structure and function. Glycobiology 2011, 21, 1541–1553. [Google Scholar] [CrossRef] [PubMed]
- Tu, L.; Banfield, D.K. Localization of golgi-resident glycosyltransferases. Cell. Mol. Life Sci. 2010, 67, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Bergstrom, K.S.B.; Xia, L. Mucin-type O-glycans and their roles in intestinal homeostasis. Glycobiology 2013, 23, 1026–1037. [Google Scholar] [CrossRef] [PubMed]
- McGuckin, M.A.; Linden, S.K.; Sutton, P.; Florin, T.H. Mucin dynamics and enteric pathogens. Nat. Rev. Microbiol. 2011, 9, 265–278. [Google Scholar] [CrossRef] [PubMed]
- Behrens, I.; Stenberg, P.; Artursson, P.; Kissel, T. Transport of lipophilic drug molecules in a new mucus-secreting cell culture model based on ht29-mtx cells. Pharm. Res. 2001, 18, 1138–1145. [Google Scholar] [CrossRef] [PubMed]
- Maares, M.; Duman, A.; Keil, C.; Schwerdtle, T.; Haase, H. The impact of apical and basolateral albumin on intestinal zinc resorption in the caco-2/ht-29-mtx co-culture model. Metallomics 2018, 10, 979–991. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2(-delta delta c(t)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maares, M.; Keil, C.; Straubing, S.; Robbe-Masselot, C.; Haase, H. Zinc Deficiency Disturbs Mucin Expression, O-Glycosylation and Secretion by Intestinal Goblet Cells. Int. J. Mol. Sci. 2020, 21, 6149. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176149
Maares M, Keil C, Straubing S, Robbe-Masselot C, Haase H. Zinc Deficiency Disturbs Mucin Expression, O-Glycosylation and Secretion by Intestinal Goblet Cells. International Journal of Molecular Sciences. 2020; 21(17):6149. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176149
Chicago/Turabian StyleMaares, Maria, Claudia Keil, Sophia Straubing, Catherine Robbe-Masselot, and Hajo Haase. 2020. "Zinc Deficiency Disturbs Mucin Expression, O-Glycosylation and Secretion by Intestinal Goblet Cells" International Journal of Molecular Sciences 21, no. 17: 6149. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176149