Characteristics of Malignant Pleural Effusion Resident CD8+ T Cells from a Heterogeneous Collection of Tumors


Abstract
:1. Introduction
2. Results
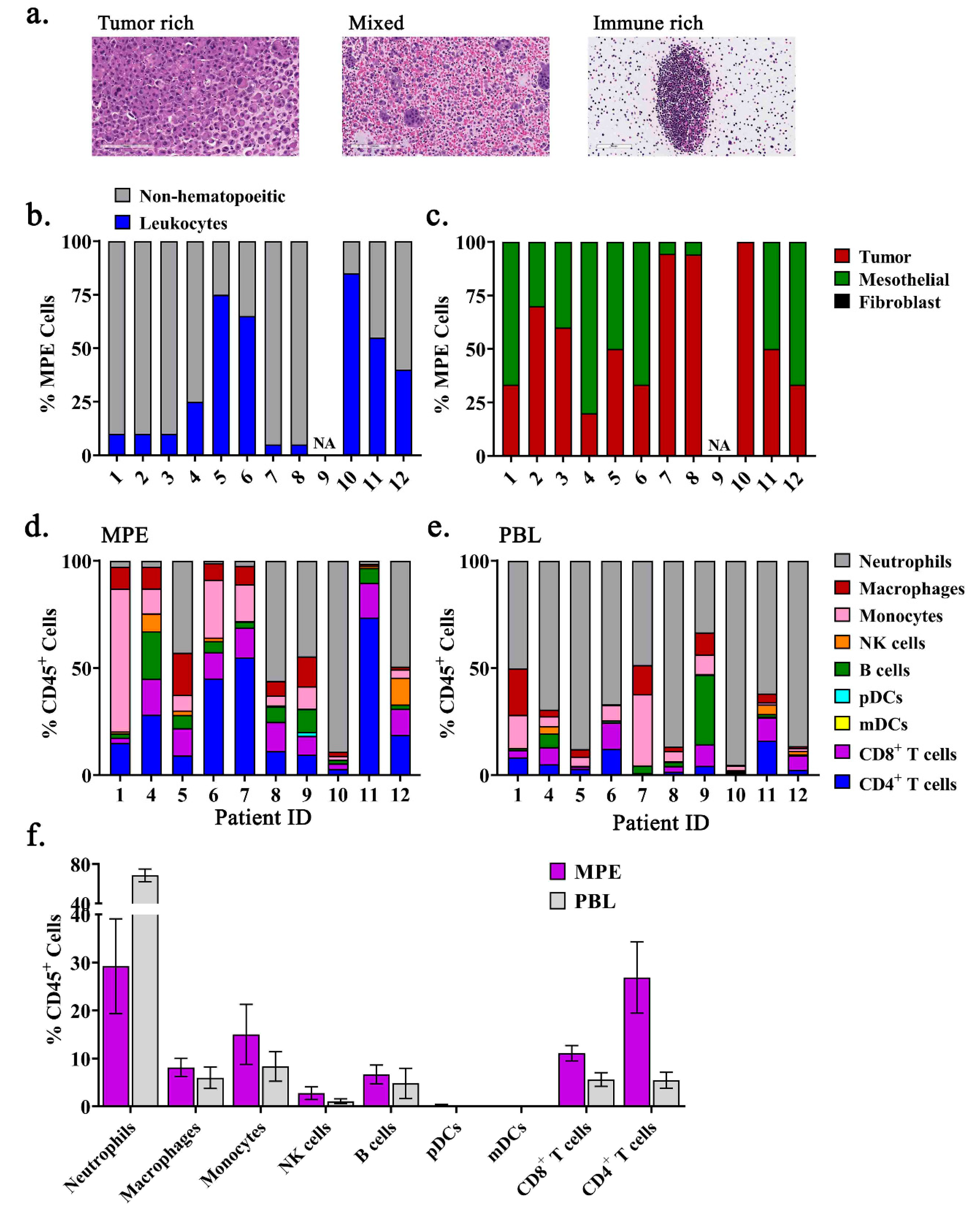
2.1. The Immune Composition of MPEs is Heterogeneous and Unique from Peripheral Blood
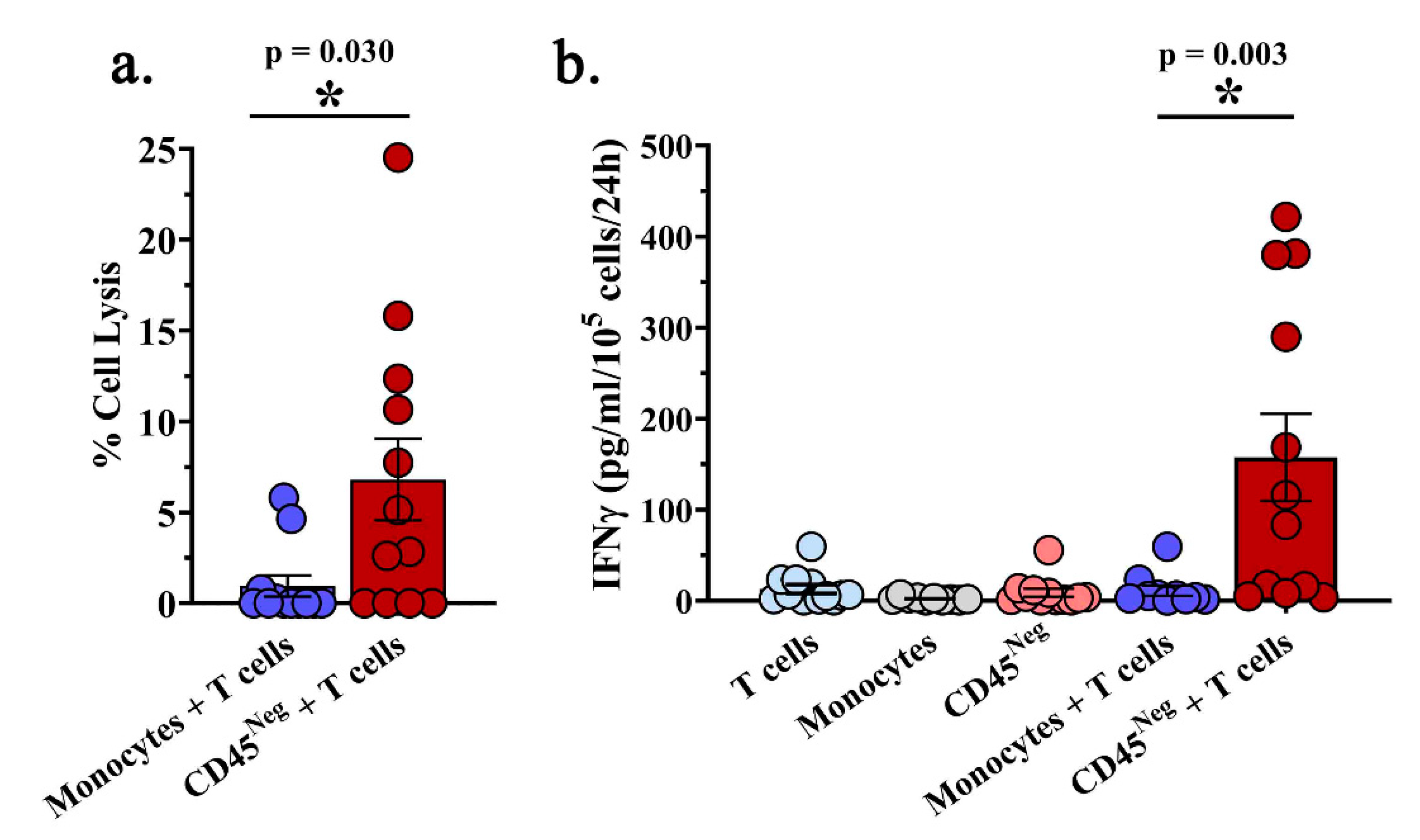
2.2. A Subset of MPE-Resident CD8+ T Cells Possess Tumor-Reactive Functionality Ex Vivo
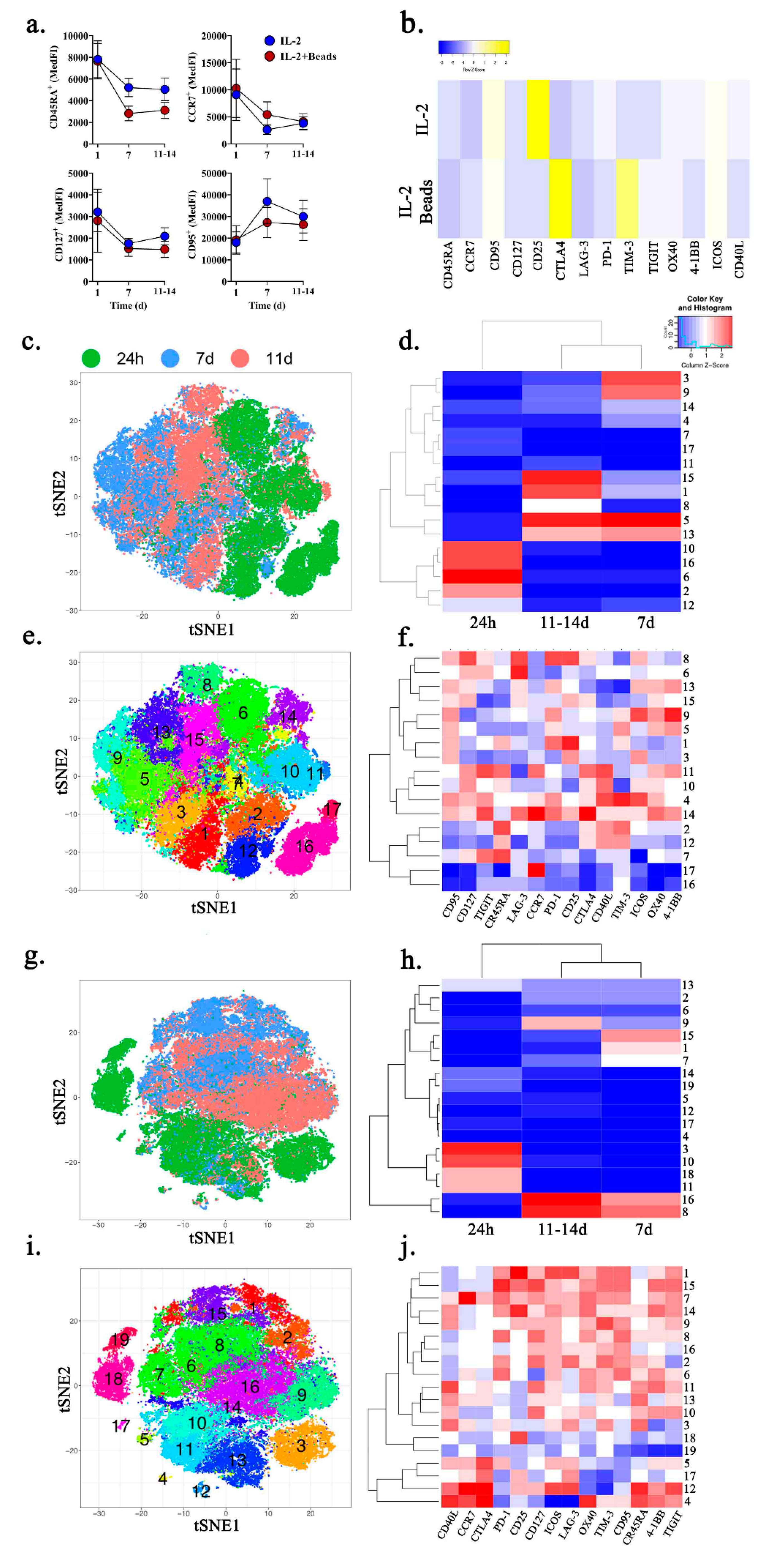
2.3. Ex Vivo Expansion of MPE-Resident CD8+ T Cells Promotes an Exhausted Phenotype
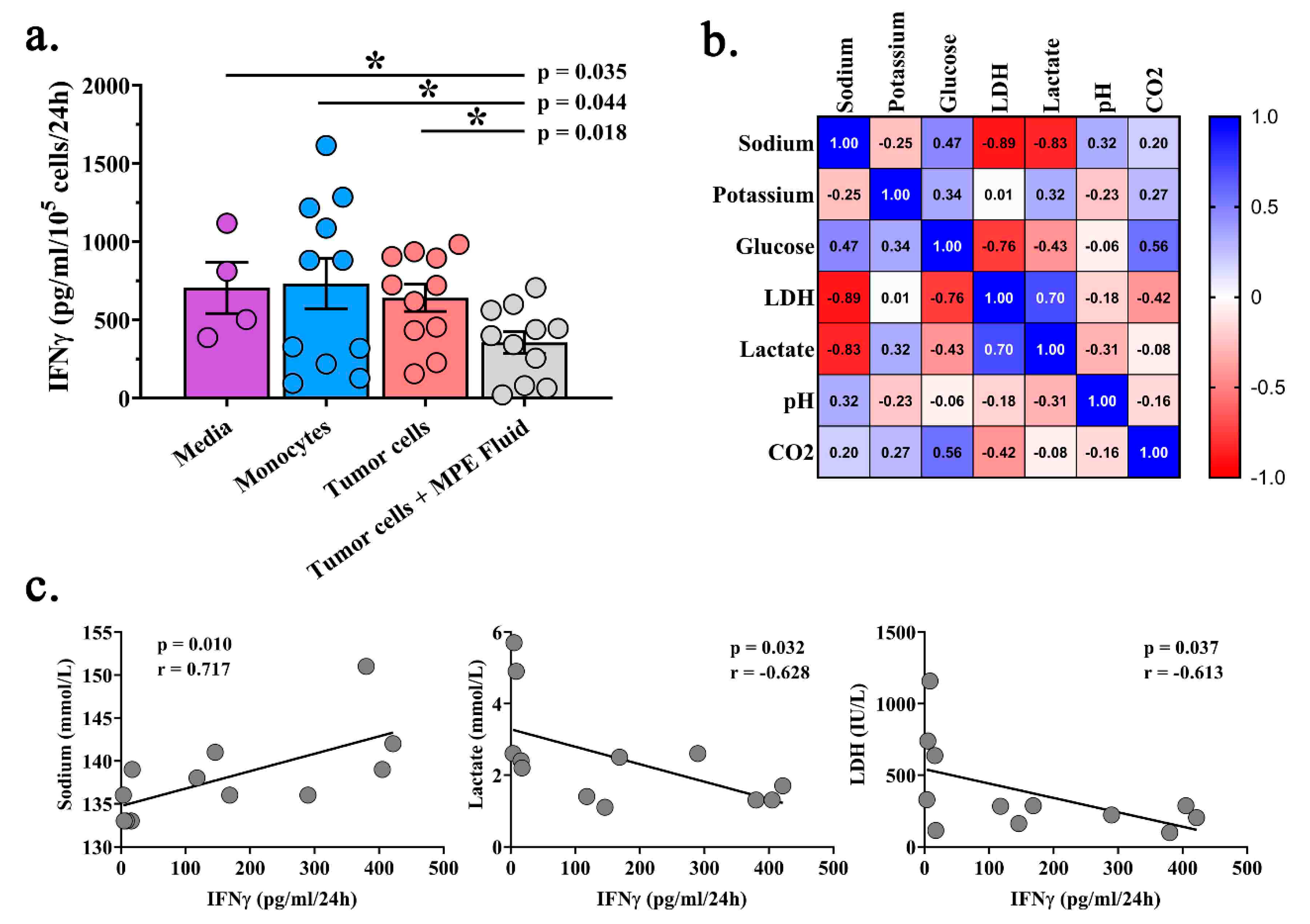
2.4. Acellular MPE Fluid Inhibits T Cell IFNγ Production and May Influence T Cell Phenotype
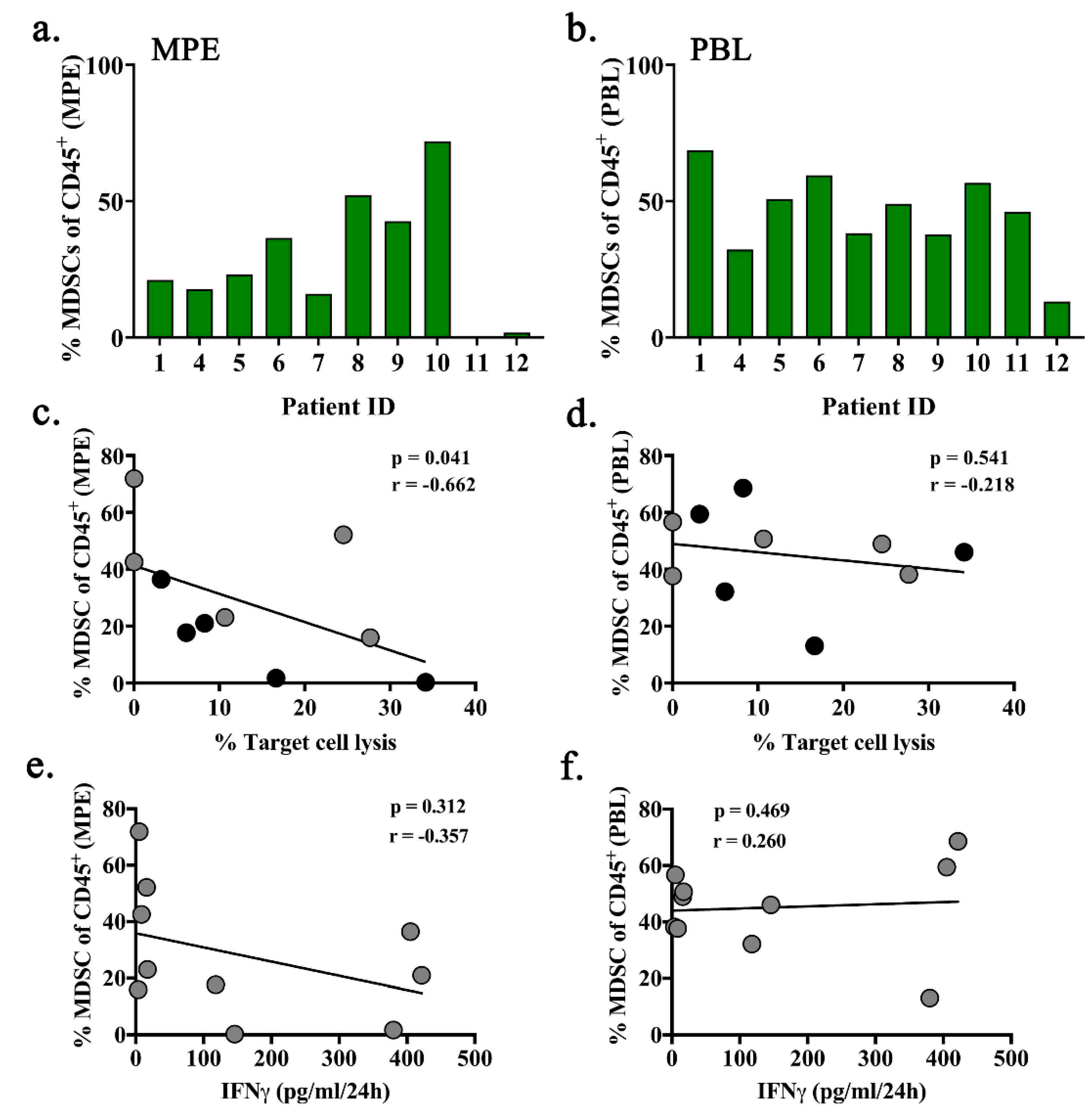
2.5. Intra-Pleural MDSCs are Associated with Decreased Cytotoxic Function of MPE-Resident T Cells
3. Discussion
4. Materials and Methods
4.1. Collection of Specimens
4.2. T Cell Expansion and Co-Cultures
4.3. LDH Cytotoxicity Assay
4.4. IFNγ Elisa
4.5. Flow Cytometry
4.6. Clinical Chemistries
4.7. Statistical Analysis and Scientific Rigor
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| MPE | Malignant pleural effusion |
| ACT | Adoptive cell transfer |
| TAM | Tumor associated macrophage |
| LDH | Lactose dehydrogenase |
| IFNγ | Interferon gamma |
| FCS | Flow cytometry standard |
| t-SNE | t-distributed stochastic neighbor embedding |
| SEM | Standard error of the mean |
| Tregs | Regulatory T cells |
| TIL | Tumor infiltrating lymphocytes |
References
- Scharping, N.; Menk, A.V.; Whetstone, R.D.; Zeng, X.; Delgoffe, G.M. Efficacy of PD-1 Blockade Is Potentiated by Metformin-Induced Reduction of Tumor Hypoxia. Cancer Immunol. Res. 2016, 5, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Neumann, A.K.; Yang, J.; Biju, M.P.; Joseph, S.K.; Johnson, R.S.; Haase, V.H.; Freedman, B.D.; Turka, L.A. Hypoxia inducible factor 1 regulates T cell receptor signal transduction. Proc. Natl. Acad. Sci. USA 2005, 102, 17071–17076. [Google Scholar] [CrossRef] [Green Version]
- Prado-García, H.; Romero-Garcia, S.; Castro-Flores, D.A.; Rumbo-Nava, U. Deficient glucose uptake is linked to impaired Glut1 expression upon CD3/CD28 stimulation in memory T cells from pleural effusions secondary to lung cancer. Scand. J. Immunol. 2019, 90, e12802. [Google Scholar] [CrossRef]
- Chang, C.; Qiu, J.; O’Sullivan, D.; Buck, M.D.; Noguchi, T.; Curtis, J.D.; Chen, Q.; Gindin, M.; Gubin, M.M.; Van Der Windt, G.J.; et al. Metabolic Competition in the Tumor Microenvironment Is a Driver of Cancer Progression. Cell 2015, 162, 1229–1241. [Google Scholar] [CrossRef] [Green Version]
- Ho, P.-C.; Bihuniak, J.D.; MacIntyre, A.; Staron, M.; Liu, X.; Amezquita, R.; Tsui, Y.-C.; Cui, G.; Micevic, G.; Perales, J.C.; et al. Phosphoenolpyruvate Is a Metabolic Checkpoint of Anti-tumor T Cell Responses. Cell 2015, 162, 1217–1228. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Yang, L.; Yue, D.; Cao, L.; Li, L.; Wang, D.; Ping, Y.; Shen, Z.; Zheng, Y.; Wang, L.; et al. Macrophage-derived CCL22 promotes an immunosuppressive tumor microenvironment via IL-8 in malignant pleural effusion. Cancer Lett. 2019, 452, 244–253. [Google Scholar] [CrossRef]
- Li, L.; Yang, L.; Wang, L.; Zhang, Y.; Yue, D.; Chen, X.; Ping, Y.; Huang, L. Impaired T cell function in malignant pleural effusion is caused by TGF-β derived predominantly from macrophages. Int. J. Cancer 2016, 139, 2261–2269. [Google Scholar] [CrossRef]
- Romero-Garcia, S.; Moreno-Altamirano, M.M.B.; Prado-García, H.; Sánchez-García, F.J. Lactate Contribution to the Tumor Microenvironment: Mechanisms, Effects on Immune Cells and Therapeutic Relevance. Front. Immunol. 2016, 7, 52. [Google Scholar] [CrossRef] [Green Version]
- Haas, R.; Smith, J.; Rocher-Ros, V.; Nadkarni, S.; Montero-Melendez, T.; D’Acquisto, F.; Bland, E.J.; Bombardieri, M.; Pitzalis, C.; Perretti, M.; et al. Lactate Regulates Metabolic and Pro-inflammatory Circuits in Control of T Cell Migration and Effector Functions. PLoS Boil. 2015, 13, e1002202. [Google Scholar] [CrossRef]
- Ohue, Y.; Nishikawa, H. Regulatory T (Treg) cells in cancer: Can Treg cells be a new therapeutic target? Cancer Sci. 2019, 110, 2080–2089. [Google Scholar] [CrossRef]
- Kumar, V.; Patel, S.; Tcyganov, E.; Gabrilovich, D.I. The Nature of Myeloid-Derived Suppressor Cells in the Tumor Microenvironment. Trends Immunol. 2016, 37, 208–220. [Google Scholar] [CrossRef] [PubMed]
- Pathria, P.; Louis, T.L.; Varner, J.A. Targeting Tumor-Associated Macrophages in Cancer. Trends Immunol. 2019, 40, 310–327. [Google Scholar] [CrossRef] [PubMed]
- Antony, P.A.; Piccirillo, C.A.; Akpinarli, A.; Finkelstein, S.E.; Speiss, P.J.; Surman, D.R.; Palmer, D.C.; Chan, C.-C.; Klebanoff, C.A.; Overwijk, W.W.; et al. CD8+ T Cell Immunity Against a Tumor/Self-Antigen Is Augmented by CD4+ T Helper Cells and Hindered by Naturally Occurring T Regulatory Cells. J. Immunol. 2005, 174, 2591–2601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salaroglio, I.C.; Kopecka, J.; Napoli, F.; Pradotto, M.; Maletta, F.; Costardi, L.; Gagliasso, M.; Milosevic, V.; Ananthanarayanan, P.; Bironzo, P.; et al. Potential Diagnostic and Prognostic Role of Microenvironment in Malignant Pleural Mesothelioma. J. Thorac. Oncol. 2019, 14, 1458–1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murthy, P.; Ekeke, C.N.; Russell, K.L.; Butler, S.C.; Wang, Y.; Luketich, J.D.; Soloff, A.C.; Dhupar, R.; Lotze, M.T. Making cold malignant pleural effusions hot: Driving novel immunotherapies. OncoImmunology 2019, 8, e1554969. [Google Scholar] [CrossRef] [Green Version]
- Donnenberg, A.D.; Luketich, J.D.; Dhupar, R.; Donnenberg, V.S. Treatment of malignant pleural effusions: The case for localized immunotherapy. J. Immunother. Cancer 2019, 7, 110. [Google Scholar] [CrossRef]
- Society, A.T. Management of Malignant Pleural Effusions. Am. J. Respir. Crit. Care Med. 2000, 162, 1987–2001. [Google Scholar]
- Chen, Y.-M.; Tsai, C.-M.; Whang-Peng, J.; Perng, R.-P. Double Signal Stimulation was Required for Full Recovery of the Autologous Tumor-Killing Effect of Effusion-Associated Lymphocytes. Chest 2002, 122, 1421–1427. [Google Scholar] [CrossRef] [Green Version]
- Tseng, Y.-H.; Ho, H.-L.; Lai, C.-R.; Luo, Y.-H.; Tseng, Y.-C.; Whang-Peng, J.; Lin, Y.-H.; Chou, T.-Y.; Chen, Y.-M. PD-L1 Expression of Tumor Cells, Macrophages, and Immune Cells in Non–Small Cell Lung Cancer Patients with Malignant Pleural Effusion. J. Thorac. Oncol. 2018, 13, 447–453. [Google Scholar] [CrossRef] [Green Version]
- Prado-García, H.; Romero-Garcia, S.; Puerto-Aquino, A.; Rumbo-Nava, U. The PD-L1/PD-1 pathway promotes dysfunction, but not “exhaustion”, in tumor-responding T cells from pleural effusions in lung cancer patients. Cancer Immunol. Immunother. 2017, 65, 87–776. [Google Scholar] [CrossRef]
- Scherpereel, A.; Grigoriu, B.; Noppen, M.; Gey, T.; Chahine, B.; Baldacci, S.; Trauet, J.; Copin, M.-C.; Dessaint, J.-P.; Porte, H.; et al. Defect in recruiting effector memory CD8+T-cells in malignant pleural effusions compared to normal pleural fluid. BMC Cancer 2013, 13, 324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atanackovic, D.; Block, A.; de Weerth, A.; Faltz, C.; Hossfeld, D.K.; Hegewisch-Becker, S. Characterization of Effusion-Infiltrating T Cells: Benign versus Malignant Effusions. Clin. Cancer Res. 2004, 10, 2600–2608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khanna, S.; Thomas, A.; Abate-Daga, D.; Zhang, J.; Morrow, B.; Steinberg, S.M.; Orlandi, A.; Ferroni, P.; Schlom, J.; Ferroni, P.; et al. Malignant Mesothelioma Effusions Are Infiltrated by CD3+ T Cells Highly Expressing PD-L1 and the PD-L1+ Tumor Cells within These Effusions Are Susceptible to ADCC by the Anti-PD-L1 Antibody Avelumab. J. Thorac. Oncol. 2016, 11, 1993–2005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prado-García, H.; Aguilar-Cazares, D.; Flores-Vergara, H.; Mandoki, J.J.; López-González, J.S. Effector, memory and naïve CD8+ T cells in peripheral blood and pleural effusion from lung adenocarcinoma patients. Lung Cancer 2005, 47, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Lau, M.C.; Wong, M.T.; Newell, E.W.; Poidinger, M.; Chen, J. Cytofkit: A Bioconductor Package for an Integrated Mass Cytometry Data Analysis Pipeline. PLoS Comput. Boil. 2016, 12, e1005112. [Google Scholar] [CrossRef]
- Poch, M.; Hall, M.; Joerger, A.; Kodumudi, K.; Beatty, M.; Innamarato, P.P.; Bunch, B.L.; Fishman, M.N.; Zhang, J.; Sexton, W.J.; et al. Expansion of tumor infiltrating lymphocytes (TIL) from bladder cancer. OncoImmunology 2018, 7, e1476816. [Google Scholar] [CrossRef]
- Chandran, S.S.; Somerville, R.P.T.; Yang, J.C.; Sherry, R.M.; Klebanoff, C.A.; Goff, S.L.; Wunderlich, J.R.; Danforth, D.N.; Zlott, D.; Paria, B.C.; et al. Treatment of metastatic uveal melanoma with adoptive transfer of tumour-infiltrating lymphocytes: A single-centre, two-stage, single-arm, phase 2 study. Lancet Oncol. 2017, 18, 792–802. [Google Scholar] [CrossRef]
- Ben-Avi, R.; Farhi, R.; Ben-Nun, A.; Gorodner, M.; Greenberg, E.; Markel, G.; Schachter, J.; Itzhaki, O.; Besser, M.J. Establishment of adoptive cell therapy with tumor infiltrating lymphocytes for non-small cell lung cancer patients. Cancer Immunol. Immunother. 2018, 67, 1221–1230. [Google Scholar] [CrossRef]
- Friedman, K.M.; Prieto, P.A.; DeVillier, L.E.; Gross, C.A.; Yang, J.C.; Wunderlich, J.R.; Rosenberg, S.A.; Dudley, M.E. Tumor-specific CD4+ Melanoma Tumor-infiltrating Lymphocytes. J. Immunother. 2012, 35, 400–408. [Google Scholar] [CrossRef]
- Pasetto, A.; Gros, A.; Robbins, P.F.; Deniger, D.C.; Prickett, T.D.; Matus-Nicodemos, R.; Douek, D.C.; Howie, B.; Robins, H.; Parkhurst, M.R.; et al. Tumor- and Neoantigen-Reactive T-cell Receptors Can Be Identified Based on Their Frequency in Fresh Tumor. Cancer Immunol. Res. 2016, 4, 734–743. [Google Scholar] [CrossRef] [Green Version]
- Gros, A.; Robbins, P.F.; Yao, X.; Li, Y.F.; Turcotte, S.; Tran, E.; Wunderlich, J.R.; Mixon, A.; Farid, S.; Dudley, M.E.; et al. PD-1 identifies the patient-specific CD8⁺ tumor-reactive repertoire infiltrating human tumors. J. Clin. Investig. 2014, 124, 2246–2259. [Google Scholar] [CrossRef] [PubMed]
- Delong, P.; Carroll, R.G.; Henry, A.C.; Tanaka, T.; Ahmad, S.; Leibowitz, M.S.; Sterman, D.H.; June, C.H.; Albelda, S.M.; Vonderheide, R.H. Regulatory T cells and cytokines in malignant pleural effusions secondary to mesothelioma and carcinoma. Cancer Boil. Ther. 2005, 4, 342–346. [Google Scholar] [CrossRef] [Green Version]
- Prado-García, H.; Romero-Garcia, S.; Rumbo-Nava, U.; Lopez-Gonzalez, J.S. Predominance of Th17 over regulatory T-cells in pleural effusions of patients with lung cancer implicates a proinflammatory profile. Anticancer. Res. 2015, 35, 1529–1535. [Google Scholar] [PubMed]
- Rivadeneira, D.B.; Delgoffe, G.M. Antitumor T-cell Reconditioning: Improving Metabolic Fitness for Optimal Cancer Immunotherapy. Clin. Cancer Res. 2018, 24, 2473–2481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, J.; Karp, J.E.; Emadi, A. Elevated lactate dehydrogenase (LDH) can be a marker of immune suppression in cancer: Interplay between hematologic and solid neoplastic clones and their microenvironments. Cancer Biomark. 2017, 19, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Husain, Z.; Huang, Y.; Seth, P.; Sukhatme, V.P. Tumor-Derived Lactate Modifies Antitumor Immune Response: Effect on Myeloid-Derived Suppressor Cells and NK Cells. J. Immunol. 2013, 191, 1486–1495. [Google Scholar] [CrossRef]
- Fischer, K.; Hoffmann, P.; Voelkl, S.; Meidenbauer, N.; Ammer, J.; Edinger, M.; Gottfried, E.; Schwarz, S.; Rothe, G.; Hoves, S.; et al. Inhibitory effect of tumor cell–derived lactic acid on human T cells. Blood 2007, 109, 3812–3819. [Google Scholar] [CrossRef]
- Hernandez, A.L.; Kitz, A.; Wu, C.; Lowther, D.E.; Rodriguez, D.M.; Vudattu, N.; Deng, S.; Herold, K.C.; Kuchroo, V.K.; Kleinewietfeld, M.; et al. Sodium chloride inhibits the suppressive function of FOXP3+ regulatory T cells. J. Clin. Investig. 2015, 125, 4212–4222. [Google Scholar] [CrossRef] [Green Version]
- Willebrand, R.; Kleinewietfeld, M. The role of salt for immune cell function and disease. Immunology 2018, 154, 346–353. [Google Scholar] [CrossRef]
- Dudley, M.E.; Gross, C.A.; Langhan, M.M.; Garcia, M.R.; Sherry, R.M.; Yang, J.C.; Phan, G.Q.; Kammula, U.S.; Hughes, M.S.; Citrin, D.E.; et al. CD8+ enriched “young” tumor infiltrating lymphocytes can mediate regression of metastatic melanoma. Clin. Cancer Res. 2010, 16, 6122–6131. [Google Scholar] [CrossRef] [Green Version]
- Dudley, M.E.; Wunderlich, J.R.; Shelton, T.E.; Even, J.; Rosenberg, S.A. Generation of Tumor-Infiltrating Lymphocyte Cultures for Use in Adoptive Transfer Therapy for Melanoma Patients. J. Immunother. 2003, 26, 332–342. [Google Scholar] [CrossRef]
- Fernandez-Poma, S.M.; Salas, D.; Lozano, T.; Casares, N.; Riezu-Boj, J.I.; Mancheño, U.; Elizalde, E.; Alignani, D.; Zubeldia, N.; Otano, I.; et al. Expansion of Tumor-Infiltrating CD8+T cells Expressing PD-1 Improves the Efficacy of Adoptive T-cell Therapy. Cancer Res. 2017, 77, 3672–3684. [Google Scholar] [CrossRef] [Green Version]
- Hall, M.; Liu, H.; Malafa, M.P.; Centeno, B.A.; Hodul, P.J.; Pimiento, J.M.; Pilon-Thomas, S.; Sarnaik, A. Expansion of tumor-infiltrating lymphocytes (TIL) from human pancreatic tumors. J. Immunother. Cancer 2016, 4, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurtado, M.O.; Wolbert, J.; Fisher, J.; Flutter, B.; Stafford, S.; Barton, J.; Jain, N.; Barone, G.; Majani, Y.; Anderson, J. Tumor infiltrating lymphocytes expanded from pediatric neuroblastoma display heterogeneity of phenotype and function. PLoS ONE 2019, 14, e0216373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poschke, I.C.; Hassel, J.C.; Ehrenfried, A.R.; Lindner, K.A.; Heras-Murillo, I.; Appel, L.M.; Lehmann, J.; Lövgren, T.; Wickström, S.L.; Lauenstein, C.; et al. The Outcome of Ex Vivo TIL Expansion Is Highly Influenced by Spatial Heterogeneity of the Tumor T-Cell Repertoire and Differences in Intrinsic In Vitro Growth Capacity between T-Cell Clones. Clin. Cancer Res. 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, K.Q.; Zhou, J.; Durflinger, K.H.; Langhan, M.M.; Shelton, T.E.; Wunderlich, J.R.; Robbins, P.F.; Rosenberg, S.A.; Dudley, M.E. Minimally Cultured Tumor-infiltrating Lymphocytes Display Optimal Characteristics for Adoptive Cell Therapy. J. Immunother. 2008, 31, 742–751. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.; Zhou, J.; Hathcock, K.S.; Robbins, P.; Powell, D.J.; Rosenberg, S.A.; Hodes, R.J. Persistence of Tumor Infiltrating Lymphocytes in Adoptive Immunotherapy Correlates With Telomere Length. J. Immunother. 2007, 30, 123–129. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | Tumor Origin | Patient Age (years) | Total Volume Collected (L)/Cellularity (cells/L) | Treatment at Drainage | Prior Treatments | Survival from Initial Drainage (days) |
|---|---|---|---|---|---|---|
| 1 | Ovarian | 68 | 1.0/8.00 × 108 | olaparib | doxorubicin, taxol | 231 |
| 2 | Pancreatic * | 70 | 0.7/5.63 × 108 | TRX518/gemcitabine | FOLFIRINOX; gemcitabine/nab-paclitaxel | 42 |
| 3 | Pancreatic * | 70 | 0.6/5.29 × 108 | TRX518/gemcitabine | FOLFIRINOX; gemcitabine/nab-paclitaxel | 42 |
| 4 | Lung | 62 | 0.5/2.32 × 108 | dabrafenib/trametinib | none | 198 |
| 5 | Breast | 62 | 0.6/8.68 × 108 | nab-paclitaxel/herceptin | pertuzumab; trastuzumab; paclitaxel; anastrozole; letrozole; tamoxifen; exemestane | 185 |
| 6 | Lung (SCLC) | 68 | 0.5/1.36 × 109 | ipilimumab/nivolumab | AEB1102; rovalpituzumab teserine; nivolumab; carboplatin/etoposide; topotecan | 45 |
| 7 | Salivary | 74 | 0.3/6.67 × 1010 | cisplatin/cyclophosphomide/doxorubicin | carboplatin/taxol; lupron; pembrolizumab | 7 |
| 8 | Gastric | 49 | 0.7/2.32 × 108 | paclitaxel/ramucirumab | FOLFOX; XELIRI and pembrolizumab | 13 |
| 9 | Lung (SCC) | 77 | 0.5/1.85 × 108 | pembrolizumab/nab-paclitaxel | none | 10 |
| 10 | Lung (AC) | 46 | 1.0/1.02 × 108 | alcetinib | none | 117 |
| 11 | Lung (NEC) | 61 | 0.4/2.2 × 109 | carboplatin/etoposide/atezolizumab | lanreotide; carboplatin/etoposide | 107 |
| 12 | Lung (AC) | 91 | 0.7/2.03 × 108 | none | none | 8 |
| Population | MPE (mean±SEM) | PBL (mean±SEM) | Spearman r | p-Value |
|---|---|---|---|---|
| Neutrophils | 29.24 ± 9.891 | 68.66 ± 6.462 | 0.5758 | 0.0883 |
| Macrophages | 8.11 ± 1.874 | 5.97 ± 2.232 | 0.4667 | 0.1786 |
| Monocytes | 15.00 ± 6.267 | 8.33 ± 3.092 | 0.8545 | 0.0029 |
| NK cells | 2.77 ± 1.336 | 1.06 ± 0.503 | 0.1520 | 0.6738 |
| B cells | 6.66 ± 1.947 | 4.82 ± 3.141 | 0.7576 | 0.0149 |
| pDCs | 0.209 ± 0.176 | 0.024 ± 0.012 | −0.2330 | 0.5240 |
| mDCs | 0.044 ± 0.013 | 0.071 ± 0.031 | −0.6123 | 0.0649 |
| CD8+ T cells | 11.09 ± 1.600 | 5.598 ± 1.425 | 0.1879 | 0.6073 |
| CD4+ T cells | 26.87 ± 7.413 | 5.445 ± 1.654 | 0.4061 | 0.2475 |
| Patient | Tumor Origin | Sodium (mEq/L) | Potassium (mEq/L) | Glucose (mg/dL) | LDH (IU/L) | Lactate (mEq/L) | pH | CO2 (mEq/L) |
|---|---|---|---|---|---|---|---|---|
| 1 | Ovarian | 142 | 3.3 | 101 | 202 | 1.7 | 8.3 | 20 |
| 2 | Pancreatic * | 136 | 4.3 | 126 | 286 | 2.5 | 7.9 | 26 |
| 3 | Pancreatic * | 136 | 4.3 | 125 | 221 | 2.6 | 8.0 | 26 |
| 4 | Lung | 138 | 4.1 | 106 | 282 | 1.4 | 8.1 | 25 |
| 5 | Breast | 139 | 4.5 | 164 | 113 | 2.2 | 8.1 | 26 |
| 6 | Lung (SCLC) | 139 | 3.9 | 101 | 286 | 1.3 | 8.1 | 22 |
| 7 | Salivary | 136 | 3.8 | 80 | 328 | 2.6 | 8.0 | 33 |
| 8 | Gastric | 133 | 3.7 | 102 | 636 | 2.4 | 8.2 | 24 |
| 9 | Lung (SCC) | 133 | 4.4 | 55 | 1158 | 4.9 | 8.3 | 24 |
| 10 | Lung (AC) | 133 | 4.0 | 85 | 739 | 5.7 | 7.9 | 21 |
| 11 | Lung (NEC) | 141 | 3.9 | 105 | 161 | 1.1 | 8.0 | 25 |
| 12 | Lung (AC) | 151 | 3.9 | 134 | 98 | 1.3 | 8.3 | 38 |
| MPE Values (mean ± SEM | 138.1 ± 1.5 | 4.0 ± 0.1 | 107 ± 8.1 | 375.8 ± 90.7 | 2.5 ± 0.42 | 8.1 ± 0.04 | 25.8 ± 1.5 | |
| Normal serum reference | 136–146 | 3.5–5.0 | 70–99 | <171 | 0.5–1.6 | 7.35–7.45 | 35–45 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dhupar, R.; Okusanya, O.T.; Eisenberg, S.H.; Monaco, S.E.; Ruffin, A.T.; Liu, D.; Luketich, J.D.; Kammula, U.S.; Bruno, T.C.; Lotze, M.T.; et al. Characteristics of Malignant Pleural Effusion Resident CD8+ T Cells from a Heterogeneous Collection of Tumors. Int. J. Mol. Sci. 2020, 21, 6178. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176178
Dhupar R, Okusanya OT, Eisenberg SH, Monaco SE, Ruffin AT, Liu D, Luketich JD, Kammula US, Bruno TC, Lotze MT, et al. Characteristics of Malignant Pleural Effusion Resident CD8+ T Cells from a Heterogeneous Collection of Tumors. International Journal of Molecular Sciences. 2020; 21(17):6178. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176178
Chicago/Turabian StyleDhupar, Rajeev, Olugbenga T. Okusanya, Seth H. Eisenberg, Sara E. Monaco, Ayana T. Ruffin, Dongyan Liu, James D. Luketich, Udai S. Kammula, Tullia C. Bruno, Michael T. Lotze, and et al. 2020. "Characteristics of Malignant Pleural Effusion Resident CD8+ T Cells from a Heterogeneous Collection of Tumors" International Journal of Molecular Sciences 21, no. 17: 6178. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176178