Modulatory Roles of ATP and Adenosine in Cholinergic Neuromuscular Transmission


Abstract
:1. Introduction
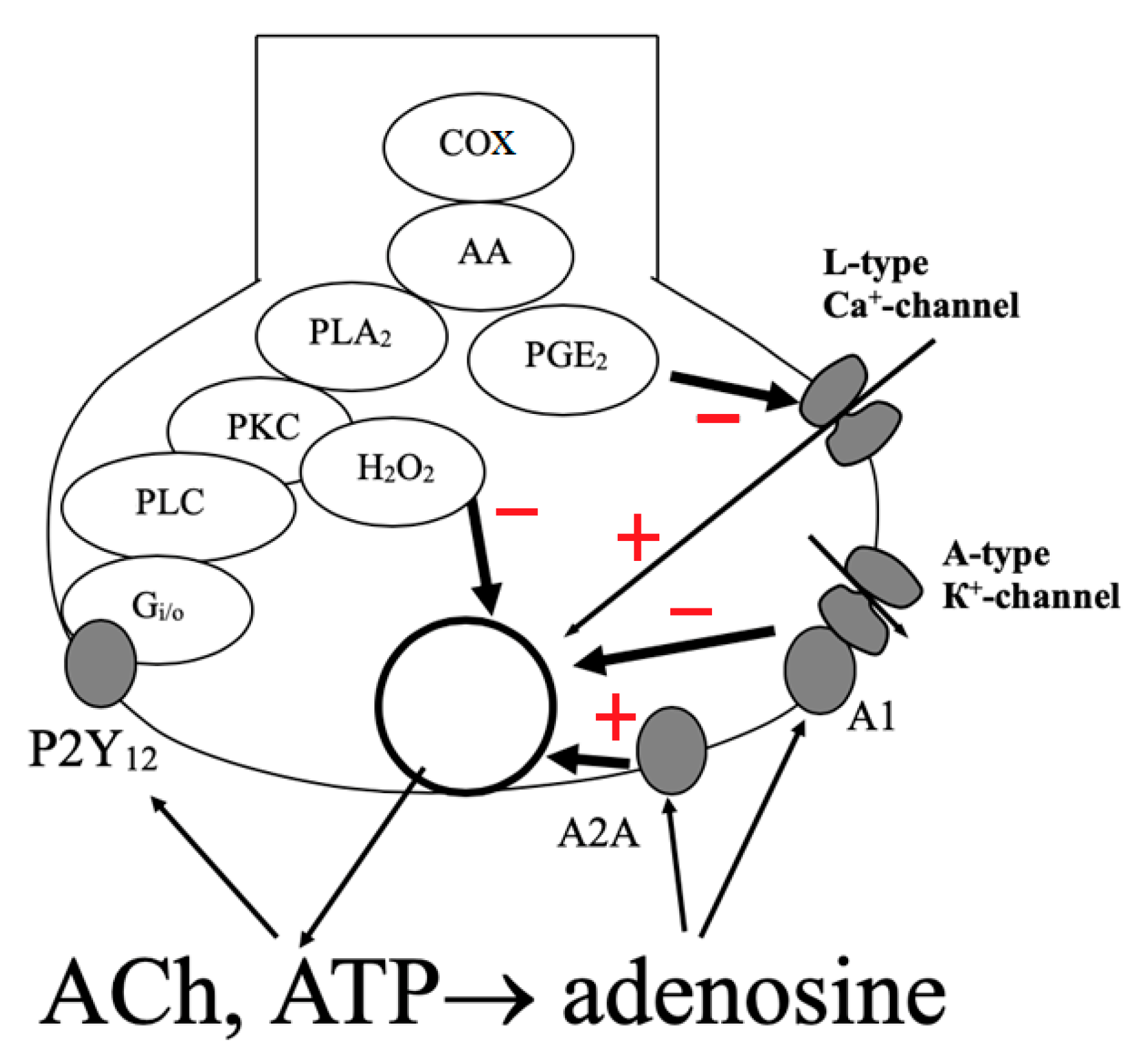
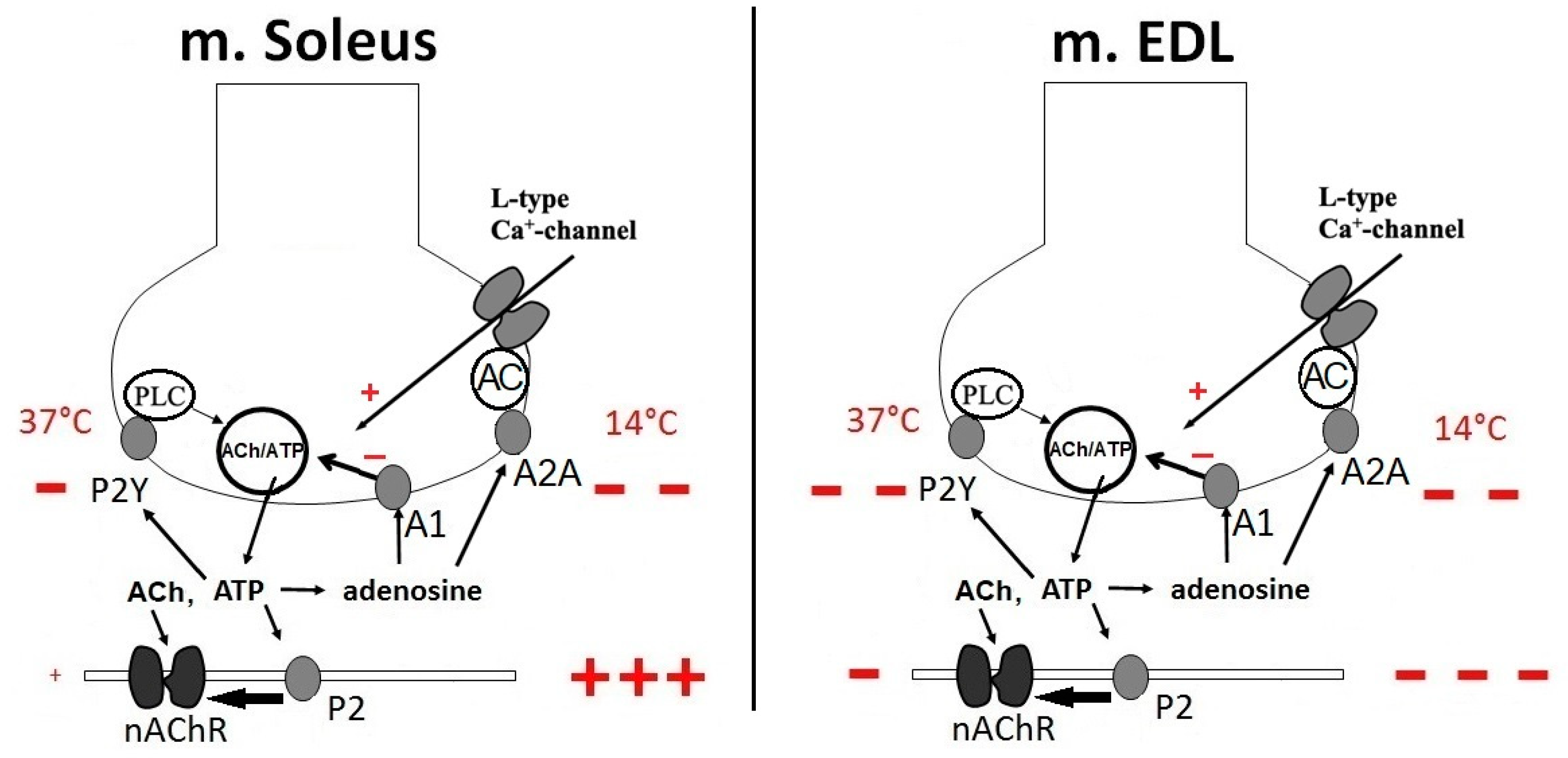
2. Effects of ATP and Adenosine on Presynaptic Currents
3. Effects of ATP and Adenosine on the Secretion of Acetylcholine
3.1. Effects of ATP and Adenosine on Induced Quantal Secretion
3.2. Effects of ATP and Adenosine on Spontaneous Quantal Secretion
3.3. Effects of ATP and Adenosine on Non-Quantal Secretion
4. Post-Synaptic Effects of ATP and Adenosine
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ACh | Acetylcholine |
| ADP | Adenosine 5′-diphosphate |
| AMP | Adenosine 5′-monophosphate |
| ATP | Adenosine 5′-triphosphate |
| CNS | Central nervous system |
| EDL | Extensor digitorum longus |
| GABA | γ-amino butyric acid |
| IDP | Inosine-5′-diphospate |
| PPADS | Pyridoxalphosphate-6-azophenyl-2′,4′-disulfonic acid |
| UTP | Uridine 5′-triphosphate |
| 2-MeSADP | 2-methylthio-adenosine 5′-diphosphate |
References
- Jimsheleishvili, S.; Marwaha, K.; Sherman, A.I. Physiology, Neuromuscular Transmission; StatPearls Publishing: Treasure Island, FL, USA, 2019. [Google Scholar]
- Mukund, K.; Subramaniam, S. Skeletal muscle: A review of molecular structure and function, in health and disease. Wiley Interdiscip. Rev. Syst. Biol. Med. 2020, 12, 1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasnoor, M.; Dimachkie, M.M. Approach to Muscle and Neuromuscular Junction Disorders. Continuum 2019, 25, 1536–1563. [Google Scholar] [CrossRef] [PubMed]
- Giniatullin, R.A.; Sokolova, E.M. The modulatory role of ATP at the neuromuscular synapse. Ross. Fiziol. Zhurnal Imeni IM Sechenovac 1998, 84, 1132–1138. (In Russian) [Google Scholar]
- Apergis-Schoute, J.; Burnstock, G.; Nusbaum, M.P.; Parker, D.; Morales, M.A.; Trudeau, L.E.; Svensson, E. Editorial: Neuronal Ca-transmission. Front. Neural Circuits 2019, 13, 19. [Google Scholar] [CrossRef] [PubMed]
- Svensson, E.; Apergis-Schoute, J.; Burnstock, G.; Nusbaum, M.P.; Parker, D.; Schiöth, H.B. General principles of neuronal co-transmission: Insights from multiple model systems. Front. Neural Circuits 2019, 21, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burnstock, G.; Dale, N. Purinergic signalling during development and ageing. Purinergic Signal. 2015, 11, 277–305. [Google Scholar] [CrossRef] [Green Version]
- Fulle, S. Purinergic signalling during myogenesis: A role for adenosine and its receptors. Acta Physiol. Oxf. 2015, 214, 436–439. [Google Scholar] [CrossRef]
- Burnstock, G. Introduction to Purinergic Signaling. Methods Mol. Biol. 2020, 2041, 1–15. [Google Scholar] [CrossRef]
- Burnstock, G. Purinergic nerves. Pharmacol. Rev. 1972, 24, 509–581. [Google Scholar]
- Burnstock, G. Purinergic receptors. J. Theor. Biol. 1976, 62, 491–503. [Google Scholar] [CrossRef]
- Alexander, S.P.; Christopoulos, A.; Davenport, A.P.; Kelly, E.; Mathie, A.; Peters, J.A.; Veale, E.L.; Armstrong, J.F.; Faccenda, E.; Harding, S.D. The concise guide to pharmacology 2019/20: G protein-coupled receptors. Br. J. Pharmacol. 2019, 176, S21–S141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, S.P.; Mathie, A.; Peters, J.A.; Veale, E.L.; Striessnig, J.; Kelly, E.; Armstrong, J.F.; Faccenda, E.; Harding, S.D.; Pawson, A.J. The concise guide to pharmacology 2019/20: Ion Channels. Br. J. Pharm. 2019, 176, S142–S228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redman, R.S.; Silinsky, E.M. ATP released together with acetylcholine as the mediator of neuromuscular depression at frog motor nerve endings. J. Physiol. 1994, 477, 117–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silinsky, E.M.; Hunt, J.M.; Solsona, C.S.; Hirsh, J.K. Prejunctional adenosine and ATP receptors. Ann. N. Y. Acad. Sci. 1990, 603, 324–333. [Google Scholar] [CrossRef]
- Silinsky, E.M.; Hirsh, J.K.; Searl, T.J.; Redman, R.S.; Watanabe, M. Quantal ATP release from motor nerve endings and its role in neurally mediated depression. Prog. Brain Res. 1999, 120, 145–158. [Google Scholar] [CrossRef]
- Cunha, R.A.; Sebastião, A.M. Extracellular metabolism of adenine nucleotides and adenosine in the innervated skeletal muscle of the frog. Eur. J. Pharm. 1991, 197, 83–92. [Google Scholar] [CrossRef]
- Edwards, F.A.; Gibb, A.J.; Colquhoun, D. ATP receptor-mediated synaptic currents in the central nervous system. Nature 1992, 359, 144–147. [Google Scholar] [CrossRef]
- Fu, W.M. Potentiation by ATP of the postsynaptic acetylcholine response at developing neuromuscular synapses in Xenopus cell cultures. J. Physiol. 1994, 477, 449–458. [Google Scholar] [CrossRef]
- Mozrzymas, J.W.; Ruzzier, F. ATP activates junctional and extrajunctional acetylcholine receptor channels in isolated adult rat muscle fibres. Neurosci. Lett. 1992, 139, 217–220. [Google Scholar] [CrossRef]
- Giniatullin, R.A.; Sokolova, E.M. ATP and adenosine inhibit transmitter release at the frog neuromuscular junction through distinct presynaptic receptors. Br. J. Pharmacol. 1998, 124, 839–844. [Google Scholar] [CrossRef] [Green Version]
- Salgado, A.I.; Cunha, R.A.; Ribeiro, J.A. Facilitation by P (2) receptor activation of acetylcholine release from rat motor nerve terminals: Interaction with presynaptic nicotinic receptors. Brain Res. 2000, 877, 245–250. [Google Scholar] [CrossRef]
- Galkin, A.V.; Giniatullin, R.A.; Mukhtarov, M.R.; Svandova, I.; Grishin, S.N.; Vyskocil, F. ATP but not adenosine inhibits nonquantal acetylcholine release at the mouse neuromuscular junction. Eur. J. Neurosci. 2001, 13, 2047–2053. [Google Scholar] [CrossRef]
- Sokolova, E.M.; Grishin, S.N.; Shakirzyanova, A.V.; Talantova, M.V.; Giniatullin, R.A. Distinct receptors and different transduction mechanisms for ATP and adenosine at the frog motor nerve endings. Eur. J. Neurosci. 2003, 18, 1254–1264. [Google Scholar] [CrossRef] [PubMed]
- De Lorenzo, S.; Veggetti, M.; Muchnik, S.; Losavio, A. Presynaptic inhibition of spontaneous acetylcholine release mediated by P2Y receptors at the mouse neuromuscular junction. Neuroscience 2006, 142, 71–85. [Google Scholar] [CrossRef]
- Malomouzh, A.; Ilyin, V.; Nikolsky, E. Components of the GABAergic signaling in the peripheral cholinergic synapses of vertebrates: A review. Amino Acids 2019, 51, 1093–1102. [Google Scholar] [CrossRef]
- Burnstock, G. Introduction to Purinergic Signalling in the Brain. Adv. Exp. Med. Biol. 2020, 1202, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Gundersen, V.; Storm-Mathisen, J.; Bergersen, L.H. Neuroglial Transmission. Physiol. Rev. 2015, 95, 695–726. [Google Scholar] [CrossRef] [Green Version]
- Magalhães-Cardoso, M.T.; Pereira, M.F.; Oliveira, L.; Ribeiro, J.A.; Cunha, R.A.; Correia-de-Sá, P. Ecto-AMP deaminase blunts the ATP-derived adenosine A2A receptor facilitation of acetylcholine release at rat motor nerve endings. J. Physiol. 2003, 549, 399–408. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, B.R.; Smith, D.O. Autoreceptor-mediated purinergic and cholinergic inhibition of motor nerve terminal calcium currents in the rat. J. Physiol. 1991, 432, 327–341. [Google Scholar] [CrossRef]
- Robitaille, R. Purinergic receptors and their activation by endogenous purines at perisynaptic glial cells of the frog neuromuscular junction. J. Neurosci. 1995, 15, 7121–7131. [Google Scholar] [CrossRef]
- Noronha-Matos, J.B.; Oliveira, L.; Peixoto, A.R.; Almeida, L.; Castellão-Santana, L.M.; Ambiel, C.R.; Alves-do-Prado, W.; Correia-de-Sá, P. Nicotinic α7 receptor-induced adenosine release from perisynaptic Schwann cells controls acetylcholine spillover from motor endplates. J. Neurochem. 2020, 154, 263–283. [Google Scholar] [CrossRef] [PubMed]
- Grishin, S.; Shakirzyanova, A.; Giniatullin, A.; Afzalov, R.; Giniatullin, R. Mechanisms of ATP action on motor nerve terminals at the frog neuromuscular junction. Eur. J. Neurosci. 2005, 21, 1271–1279. [Google Scholar] [CrossRef] [PubMed]
- Grishin, S.N. Neuromuscular transmission in Ca2+-free extracellular solution. Biochem. Mosc. Suppl. Ser. A 2016, 10, 99–108. [Google Scholar] [CrossRef]
- Khaziev, E.F.; Samigullin, D.V.; Tsentsevitsky, A.N.; Bukharaeva, E.A.; Nikolsky, E.E. ATP reduces the entry of calcium ions into the nerve ending by blocking L-type calcium channels. Acta Nat. 2018, 10, 93–96. [Google Scholar] [CrossRef] [Green Version]
- Correia-de-Sá, P.; Timóteo, M.A.; Ribeiro, J.A. A (2A) Adenosine receptor facilitation of neuromuscular transmission: Influence of stimulus paradigm on calcium mobilization. J. Neurochem. 2000, 74, 2462–2469. [Google Scholar] [CrossRef] [PubMed]
- Zuber, B.; Lučić, V. Molecular architecture of the presynaptic terminal. Curr. Opin. Struct. Biol. 2019, 54, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Slater, C.R. The functional organization of motor nerve terminals. Prog. Neurobiol. 2015, 134, 55–103. [Google Scholar] [CrossRef]
- Hounsgaard, J. Motor Neurons. Compr. Physiol. 2017, 7, 463–484. [Google Scholar] [CrossRef]
- Fatt, P.; Katz, B. Spontaneous subthreshold activity at motor nerve endings. J. Physiol. 1952, 117, 109–128. [Google Scholar]
- Del Castillo, J.; Katz, B. Local activity at a depolarized nerve-muscle junction. J. Physiol. 1955, 128, 396–411. [Google Scholar] [CrossRef]
- Andreae, L.C.; Burrone, J. The role of spontaneous neurotransmission in synapse and circuit development. J. Neurosci. Res. 2018, 96, 354–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nassenstein, C.; Wiegand, S.; Lips, K.S.; Li, G.; Klein, J.; Kummer, W. Cholinergic activation of the murine trachealis muscle via non-vesicular acetylcholine release involving low-affinity choline transporters. Int. Immunopharmacol. 2015, 29, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, J.A.; Walker, J. Action of adenosine triphosphate on endplate potentials recorded from muscle fibres of the rat-diaphragm and frog sartorius. Br. J. Pharmacol. 1973, 49, 724–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, J.A.; Dominguez, M.L. Mechanisms of depression of neuromuscular transmission by ATP and adenosine. J. Physiol. 1978, 74, 491–496. [Google Scholar]
- Grishin, S.N.; Ziganshin, A.U. Modulatory role of purines in neuromuscular transmission. Biochem. Mosc. Suppl. Ser. A 2013, 7, 183–191. [Google Scholar] [CrossRef]
- Giniatullin, A.R.; Grishin, S.N.; Sharifullina, E.R.; Petrov, A.M.; Zefirov, A.L.; Giniatullin, R.A. Reactive oxygen species contribute to the presynaptic action of extracellular ATP at the frog neuromuscular junction. J. Physiol. 2005, 15, 229–242. [Google Scholar] [CrossRef]
- Arkhipova, O.V.; Grishin, S.N.; Sitdikova, G.F.; Zefirov, A.L. The presynaptic effects of arachidonic acid and prostaglandin E2 at the frog neuromuscular junction. Neurosci. Behav. Physiol. 2006, 36, 307–312. [Google Scholar] [CrossRef]
- Cunha, R.A. How does adenosine control neuronal dysfunction and neurodegeneration? J. Neurochem. 2016, 139, 1019–1055. [Google Scholar] [CrossRef]
- Correia-De-Sá, P.; Sebastião, A.M.; Ribeiro, J.A. Inhibitory and excitatory effects of adenosine receptor agonists on evoked transmitter release from phrenic nerve endings of the rat. Br. J. Pharmacol. 1991, 103, 1614–1620. [Google Scholar] [CrossRef]
- Ribeiro, J.A.; Cunha, R.A.; Correia-de-Sá, P.; Sebastião, A.M. Purinergic regulation of acetylcholine release. Prog. Brain Res. 1996, 109, 231–241. [Google Scholar]
- Grishin, S.N.; Galkin, A.V.; Priazhnikov, E.G.; Sokolova, E.M.; Giniatullin, R.A. Role of adenosine receptors in modulation of synaptic transmission. Neurophysiology 2002, 34, 155–157. [Google Scholar] [CrossRef]
- Oliveira, L.; Correia, A.; Costa, A.C.; Guerra-Gomes, S.; Ferreirinha, F.; Magalhães-Cardoso, M.T.; Vilanova, M.; Correia-de-Sá, P. Deficits in endogenous adenosine formation by ecto-5′-nucleotidase/CD73 impair neuromuscular transmission and immune competence in experimental autoimmune Myasthenia gravis. Mediat. Inflamm. 2015, 2015, 460610. [Google Scholar] [CrossRef] [PubMed]
- Cacciari, B.; Spalluto, G.; Federico, S. A2A Adenosine receptor antagonists as therapeutic candidates: Are they still an interesting challenge? Mini Rev. Med. Chem. 2018, 18, 1168–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grishin, S.N.; Ziganshin, A.U. Co-Transmitter Modulation of Neuromuscular Transmission in Different Types of Motor Units; Kazan University: Kazan, Russia, 2015; p. 240. [Google Scholar]
- Oliveira, L.; Timóteo, M.A.; Correia-de-Sá, P. Modulation by adenosine of both muscarinic M1-facilitation and M2-inhibition of [3H]-acetylcholine release from the rat motor nerve terminals. Eur. J. Neurosci. 2002, 15, 1728–1736. [Google Scholar] [CrossRef]
- Tomàs, J.; Santafé, M.M.; Garcia, N.; Lanuza, M.A.; Tomàs, M.; Besalduch, N.; Obis, T.; Priego, M.; Hurtado, E. Presynaptic membrane receptors in acetylcholine release modulation in the neuromuscular synapse. J. Neurosci. Res. 2014, 92, 543–554. [Google Scholar] [CrossRef]
- Tomàs, J.; Garcia, N.; Lanuza, M.A.; Santafé, M.M.; Tomàs, M.; Nadal, L.; Hurtado, E.; Simó-Ollé, A.; Cilleros-Mañé, V.; Just-Borràs, L. Adenosine receptors in developing and adult mouse neuromuscular junctions and functional links with other metabotropic receptor pathways. Front. Pharm. 2018, 9, 397. [Google Scholar] [CrossRef] [Green Version]
- Bernareggi, A.; Sciancalepore, M.; Lorenzon, P. Interplay between cholinergic and adenosinergic systems in skeletal muscle. Neuroscience 2019, 19, 30336–30337. [Google Scholar] [CrossRef]
- Giniatullin, A.R.; Grishin, S.N.; Giniatullin, R.A. Effect of hydrocortisone on purine actions in the nerve-muscle preparation. Ross. Fiziol. Zhurnal Imeni IM Sechenovac 2000, 86, 1293–1299. [Google Scholar]
- Grishin, S.N.; Gabdrakhmanov, A.I.; Khairullin, A.E.; Ziganshin, A.U. The Influence of glucocorticoids and catecholamines on the neuromuscular transmission. Biochem. Mosc. Suppl. Ser. A 2017, 11, 253–260. [Google Scholar] [CrossRef]
- Ziganshin, A.U.; Kamaliev, R.R.; Gabdrakhmanov, A.I.; Khairullin, A.E.; Grishin, S.N. Foot-shock stimulation decreases the inhibitory action of ATP on contractility and end-plate current of frog sartorius muscle. Int. J. Pharmacol. 2018, 14, 1198–1202. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, L.; Costa, A.C.; Noronha-Matos, J.B.; Silva, I.; Cavalcante, W.L.; Timóteo, M.A.; Corrado, A.P.; Dal Belo, C.A.; Ambiel, C.R.; Alves-do-Prado, W.; et al. Amplification of neuromuscular transmission by methylprednisolone involves activation of presynaptic facilitatory adenosine A2A receptors and redistribution of synaptic vesicles. Neuropharmacology 2015, 89, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Giniatullin, A.; Petrov, A.; Giniatullin, R. The involvement of P2Y12 receptors, NADPH oxidase, and lipid rafts in the action of extracellular ATP on synaptic transmission at the frog neuromuscular junction. Neuroscience 2015, 285, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Guarracino, J.F.; Cinalli, A.R.; Fernández, V.; Roquel, L.I.; Losavio, A.S. P2Y13 receptors mediate presynaptic inhibition of acetylcholine release induced by adenine nucleotides at the mouse neuromuscular junction. Neuroscience 2016, 326, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Guarracino, J.F.; Cinalli, A.R.; Veggetti, M.I.; Losavio, A.S. Endogenous purines modulate K+-evoked ACh secretion at the mouse neuromuscular junction. J. Neurosci. Res. 2018, 96, 1066–1079. [Google Scholar] [CrossRef]
- Lindgren, C.A.; Smith, D.O. Extracellular ATP modulates calcium uptake and transmitter release at the neuromuscular junction. J. Neurosci. 1987, 7, 1567–1573. [Google Scholar] [CrossRef] [Green Version]
- Celenza, K.M.; Shugert, E.; Vélez, S.J. Depressing effect of caffeine at crayfish neuromuscular synapses II. Initial search for possible sites of action. Cell. Mol. Neurobiol. 2007, 27, 381–393. [Google Scholar] [CrossRef]
- Ribeiro, J.A.; Sebastião, A.M. On the role, inactivation and origin of endogenous adenosine at the frog neuromuscular junction. J. Physiol. 1987, 384, 571–585. [Google Scholar] [CrossRef]
- Supinski, G.S.; Deal, E.C., Jr.; Kelsen, S.G. Comparative effects of theophylline and adenosine on respiratory skeletal and smooth muscle. Am. Rev. Respir. Dis. 1986, 133, 809–813. [Google Scholar]
- Ziganshin, A.U.; Khairullin, A.E.; Teplov, A.Y.; Gabdrakhmanov, A.I.; Ziganshina, L.E.; Hoyle, C.H.V.; Ziganshin, B.A.; Grishin, S.N. The effects of ATP on the contractions of rat and mouse fast skeletal muscle. Muscle Nerve 2019, 59, 509–516. [Google Scholar] [CrossRef]
- Khairullin, A.E.; Teplov, A.Y.; Grishin, S.N.; Farkhutdinov, A.M.; Ziganshin, A.U. The thermal sensitivity of purinergic modulation of contractile activity of locomotor and respiratory muscles in mice. Biophysics 2019, 64, 812–817. [Google Scholar] [CrossRef]
- Buckle, P.J.; Spence, I. The actions of adenosine and some analogues on evoked and potassium stimulated release at skeletal and autonomic neuromuscular junctions. Naunyn Schmiedebergs Arch. Pharmacol. 1982, 319, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Khairullin, A.E.; Ziganshin, A.U.; Grishin, S.N. Effect of hypothermia on purinergic synaptic modulation in the rat diaphragm. Biophysics 2020, accepted. [Google Scholar]
- Chiou, L.C.; Hong, S.J.; Chang, C.C. Does endogenous adenosine modulate the release of acetylcholine from motor nerve during single and repetitive stimulations in the mouse diaphragm? JPN J. Pharmacol. 1987, 44, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Ziganshin, A.U.; Khairullin, A.E.; Zobov, V.V.; Ziganshina, L.E.; Gabdrakhmanov, A.I.; Ziganshin, B.A.; Grishin, S.N. Effects of ATP and adenosine on contraction amplitude of rat soleus muscle at different temperatures. Muscle Nerve 2017, 55, 417–423. [Google Scholar] [CrossRef]
- Sebastião, A.M.; Ribeiro, J.A. On the adenosine receptor and adenosine inactivation at the rat diaphragm neuromuscular junction. Br. J. Pharmacol. 1988, 94, 109–120. [Google Scholar] [CrossRef]
- Elnozahi, N.A.; AlQot, H.E.; Mohy El-Din, M.M.; Bistawroos, A.E.; Abou Zeit-Har, M.S. Modulation of dopamine-mediated facilitation at the neuromuscular junction of Wistar rats: A role for adenosine A1/A2A receptors and P2 purinoceptors. Neuroscience 2016, 326, 45–55. [Google Scholar] [CrossRef]
- Voss, A.A. Extracellular ATP inhibits chloride channels in mature mammalian skeletal muscle by activating P2Y1 receptors. J. Physiol. 2009, 587, 5739–5752. [Google Scholar] [CrossRef]
- Shakirzyanova, A.V.; Nikol’skii, E.E.; Giniatullin, R.A. Effects of carbachol and adenosine on neurotransmitter secretion induced by potassium chloride, ionomycin, and sucrose. Bull. Exp. Biol. Med. 2005, 139, 193–195. [Google Scholar] [CrossRef]
- Shakirzyanova, A.V.; Bukharaeva, E.A.; Nikolsky, E.E.; Giniatullin, R.A. Negative cross-talk between presynaptic adenosine and acetylcholine receptors. Eur. J. Neurosci. 2006, 24, 105–115. [Google Scholar] [CrossRef]
- Vyskocil, F.; Illes, P. Non-quantal release of transmitter at mouse neuromuscular junction and its dependence on the activity of Na+-K+ ATP-ase. Pflug. Arch. 1977, 370, 295–297. [Google Scholar] [CrossRef]
- Malomouzh, A.I.; Nikolsky, E.E.; Vyskočil, F. Purine P2Y receptors in ATP-mediated regulation of non-quantal acetylcholine release from motor nerve endings of rat diaphragm. Neurosci. Res. 2011, 71, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Katz, B.; Miledi, R. Transmitter Leakage from Motor Nerve Endings. Proc. R. Soc. B Biol. Sci. 1977, 196, 59–72. [Google Scholar] [CrossRef]
- Xu, M.L.; Bi, C.W.; Cheng, L.K.; Mak, S.; Yao, P.; Luk, W.K.; Lau, K.K.; Cheng, A.W.; Tsim, K.W. Reduced Expression of P2Y2 Receptor and Acetylcholinesterase at Neuromuscular Junction of P2Y1 Receptor Knock-out Mice. J. Mol. Neurosci. 2015, 57, 446–451. [Google Scholar] [CrossRef] [PubMed]
- Trang, A.; Khandhar, P.B. Physiology, Acetylcholinesterase; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Khairullin, A.E.; Ziganshin, A.U.; Grishin, S.N. Motor units at various temperatures. Biochem. Mosc. Suppl. Ser. A 2017, 11, 1–7. [Google Scholar] [CrossRef]
- Ziganshin, A.U.; Grishin, S.N. Temperature-dependent effects of ATP on smooth and skeletal muscles. In Adenosine Triphosphate in Health and Disease; Mozsik, G., Ed.; IntechOpen: London, UK, 2019; pp. 11–23. [Google Scholar] [CrossRef] [Green Version]
- Burnstock, G.; Arnett, T.R.; Orriss, I.R. Purinergic signalling in the musculoskeletal system. Purinergic Signal. 2013, 9, 541–572. [Google Scholar] [CrossRef] [Green Version]
- Bernareggi, A.; Ren, E.; Giniatullin, A.; Luin, E.; Sciancalepore, M.; Giniatullin, R.; Lorenzon, P. Adenosine promotes endplate nAChR channel activity in adult mouse skeletal muscle fibers via low affinity P1 receptors. Neuroscience 2018, 383, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Fu, W.M. Regulatory role of ATP at developing neuromuscular junctions. Prog. Neurobiol. 1995, 47, 31–44. [Google Scholar] [CrossRef]
- Igusa, Y. Adenosine 5′-triphosphate activates acetylcholine receptor channels in cultured Xenopus myotomal muscle cells. J. Physiol. 1988, 405, 169–185. [Google Scholar] [CrossRef]
- Carlson, B.J.; Raftery, M.A. Specific binding of ATP to extracellular sites on Torpedo acetylcholine receptor. Biochemistry 1993, 32, 7329–7333. [Google Scholar] [CrossRef]
- Giniatullin, R.A.; Khamitov, K.S.; Khazipov, R.N.; Magazanik, L.G.; Nikolsky, E.E.; Sneikov, V.A.; Vyskocil, F. Development of desensitization during repetitive end-plate activity and single end-plate currents in frog muscle. J. Physiol. 1989, 412, 113–122. [Google Scholar] [CrossRef]
- Magazanik, L.G.; Nikolsky, E.E.; Giniatullin, R.A. End-plate currents evoked by paired stimuli in frog muscle. Pflug. Arch. 1984, 401, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Smith, D.O. Adenosine 5′-triphosphate increases acetylcholine channel-opening frequency in rat skeletal muscle. J. Physiol. 1991, 436, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Sandona, D.; Danieli-Betto, D.; Germinario, E.; Biral, D.; Martinello, T.; Lioy, A.; Tarricone, E.; Gastaldello, S.; Betto, R. The T-tubule membrane ATP-operated P2X4 receptor influences contractility of skeletal muscle. FASEB J. 2005, 19, 1184–1186. [Google Scholar] [CrossRef] [PubMed]
- Deuchars, S.A.; Atkinson, L.; Brooke, R.E.; Musa, H.; Milligan, C.J.; Batten, T.F.; Buckley, N.J.; Parson, S.H.; Deuchars, J. Neuronal P2X7 receptors are targeted to presynaptic terminals in the central and peripheral nervous systems. J. Neurosci. 2001, 21, 7143–7152. [Google Scholar] [CrossRef] [Green Version]
- Moores, T.S.; Hasdemir, B.; Vega-Riveroll, L.; Deuchars, J.; Parson, S.H. Properties of presynaptic P2X7-like receptors at the neuromuscular junction. Brain Res. 2005, 1034, 40–50. [Google Scholar] [CrossRef]
- Koles, L.; Furst, S.; Illes, P. Purine ionotropic (P2X) receptors. Curr. Pharm. Des. 2007, 13, 2368–2384. [Google Scholar] [CrossRef]
- Koles, L.; Gerevich, Z.; Oliveira, J.F.; Zadori, Z.S.; Wirkner, K.; Illes, P. Interaction of P2 purinergic receptors with cellular macromolecules. Naunyn Schmiedeberg’s. Arch. Pharmacol. 2008, 377, 1–33. [Google Scholar] [CrossRef]
- Ferrari, D.; Pizzirani, C.; Adinolfi, E.; Lemoli, R.M.; Curti, A.; Idzko, M.; Panther, E.; Virgilio, F.D. The P2X7 receptor: A key player in IL-1 processing and release. J. Immunol. 2006, 176, 3877–3883. [Google Scholar] [CrossRef] [Green Version]
- Wood, S.J.; Slater, C. Safety factor at the neuromuscular junction. Prog. Neurobiol. 2001, 64, 393–429. [Google Scholar] [CrossRef]
- Grishin, S.N.; Ziganshin, A.U. Synaptic organization of tonic motor units in vertebrates. Biochem. Mosc. Suppl. Ser. A 2015, 9, 13–20. [Google Scholar] [CrossRef]
- Pousinha, P.; Ribeiro, J.A. Regulation of synaptic transmission by adenosine at the neuromuscular junction. In Adenosine Receptors in Neurodegenerative Disease; Blum, D., Lopes, L.V., Eds.; Academic Press: London, UK, 2017; pp. 77–96. [Google Scholar]



{kind=link}
{kind=link}
| Animal | Muscle Name | Muscle Type | Synaptic Level | Frequency of Electrical Stimulation (Hz) | Agent | References | |
|---|---|---|---|---|---|---|---|
| Adenosine | ATP | ||||||
| Crayfish | Walking leg | - | Pre-synaptic | 0.5–2 | NE | ↓ | [67] |
| - | NE | NT | [68] | ||||
| Frog | Sartorius | - | Pre-synaptic | 0.05 | NT | ↓ | [14] |
| 0.03 | ↓ | ↓ | [21] | ||||
| 0.5 | NT | ↓ | [44] | ||||
| 0.03 | NT | ↓ | [64] | ||||
| 0.5 | ↓ | NT | [69] | ||||
| Guinea pig | Diaphragm | Mixed | - | NE | NT | [70] | |
| Mouse | EDL | Fast | Pre-synaptic | 0.1 | ↓ | ↑ | [71] |
| Post-synaptic | - | NE | ↓ | [71] | |||
| Soleus | Slow | Pre-synaptic | 0.1 | NT | ↑ | [72] | |
| Post-synaptic | - | NT | NE | [72] | |||
| Diaphragm | Mixed | Pre-synaptic | 0.5 | NT | ↓ | [65] | |
| 0.5 | ↓ | NT | [73] | ||||
| 0.1 | NT | ↑ | [74] | ||||
| 1 | NE | ↓ | [75] | ||||
| Post-synaptic | - | NT | ↑ | [74] | |||
| Rat | EDL | Fast | Pre-synaptic | 0.1 | ↓ | ↓ | [72] |
| Post-synaptic | - | NE | ↓ | [72] | |||
| Soleus | Slow | Pre-synaptic | 0.1 | ↓ | ↓ | [76] | |
| Post-synaptic | - | NE | NE | [76] | |||
| Diaphragm | Mixed | Pre-synaptic | 0.5 | NT | ↓ | [44] | |
| 0.5 | ↓ | NT | [77] | ||||
| 0.1 | ↓ or ↑ * | ↓ | [78] | ||||
| 0.1 | NT | ↑ | [74] | ||||
| Post-synaptic | - | NT | ↑ | [74] | |||
| LAL | Fast | Pre-synaptic | 2 | NT | ↑ | [79] | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ziganshin, A.U.; Khairullin, A.E.; Hoyle, C.H.V.; Grishin, S.N. Modulatory Roles of ATP and Adenosine in Cholinergic Neuromuscular Transmission. Int. J. Mol. Sci. 2020, 21, 6423. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176423
Ziganshin AU, Khairullin AE, Hoyle CHV, Grishin SN. Modulatory Roles of ATP and Adenosine in Cholinergic Neuromuscular Transmission. International Journal of Molecular Sciences. 2020; 21(17):6423. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176423
Chicago/Turabian StyleZiganshin, Ayrat U., Adel E. Khairullin, Charles H. V. Hoyle, and Sergey N. Grishin. 2020. "Modulatory Roles of ATP and Adenosine in Cholinergic Neuromuscular Transmission" International Journal of Molecular Sciences 21, no. 17: 6423. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176423