FRUITFULL Is a Repressor of Apical Hook Opening in Arabidopsis thaliana

 , , , , , and
, , , , , and
Abstract
:1. Introduction
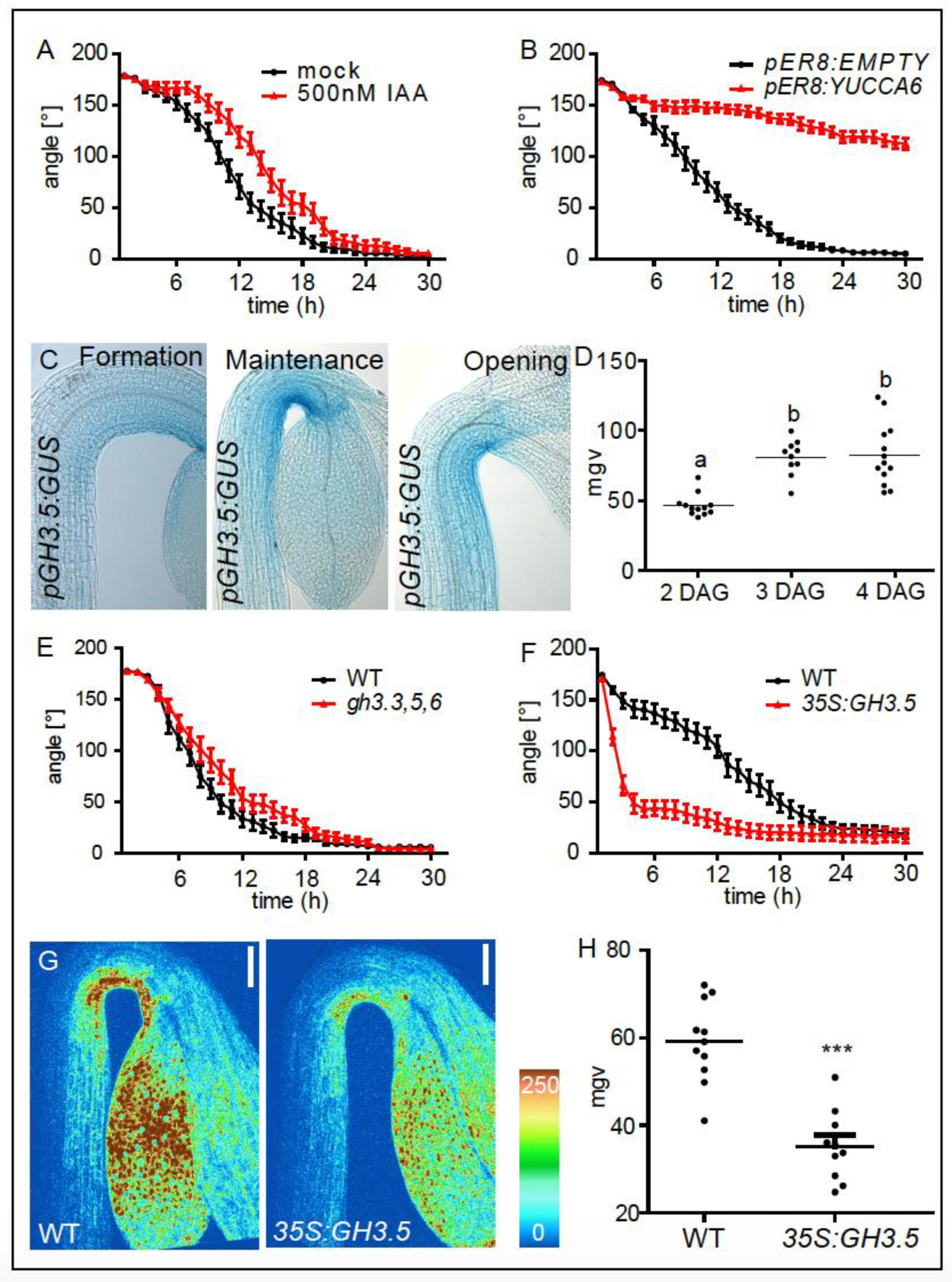
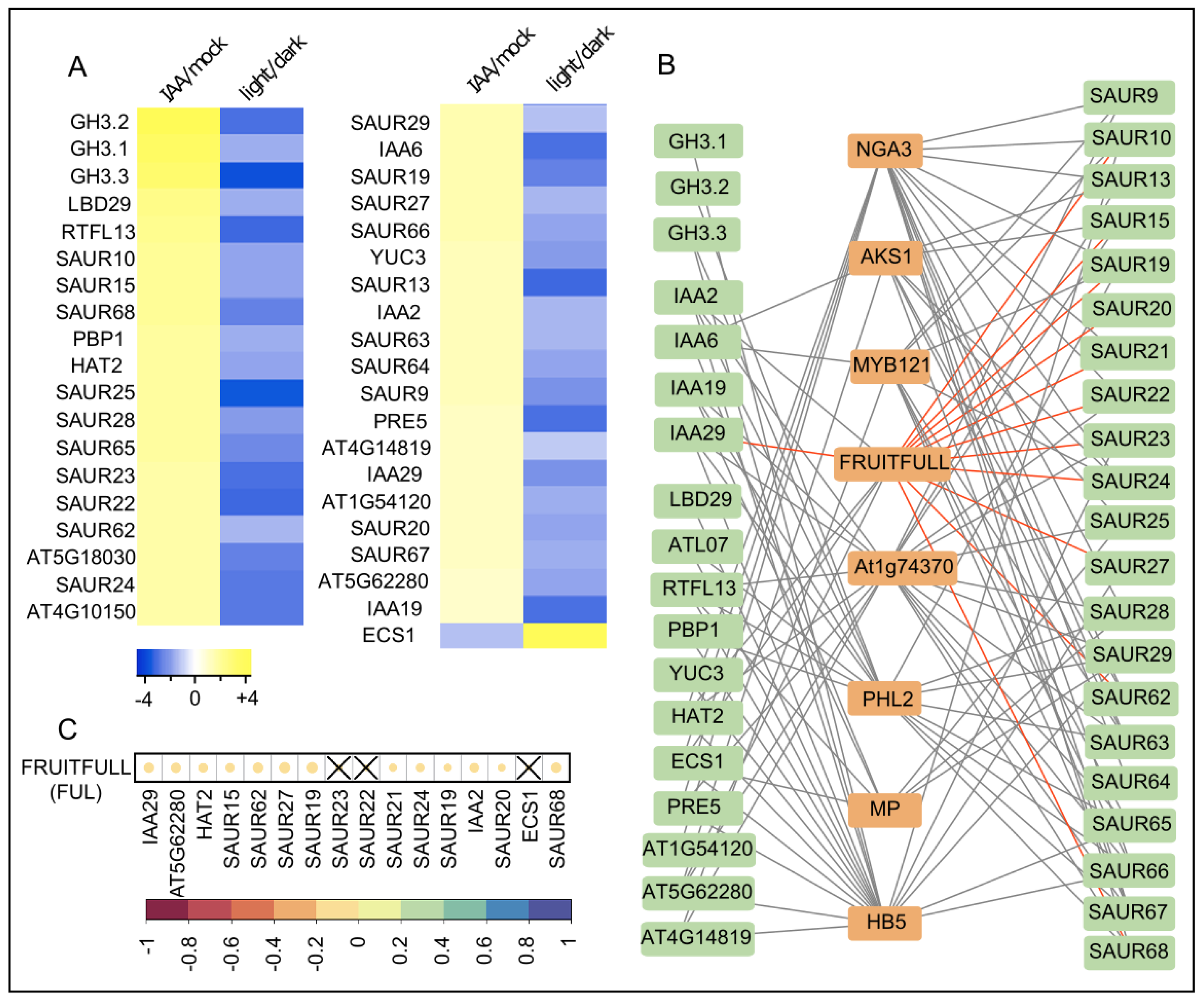
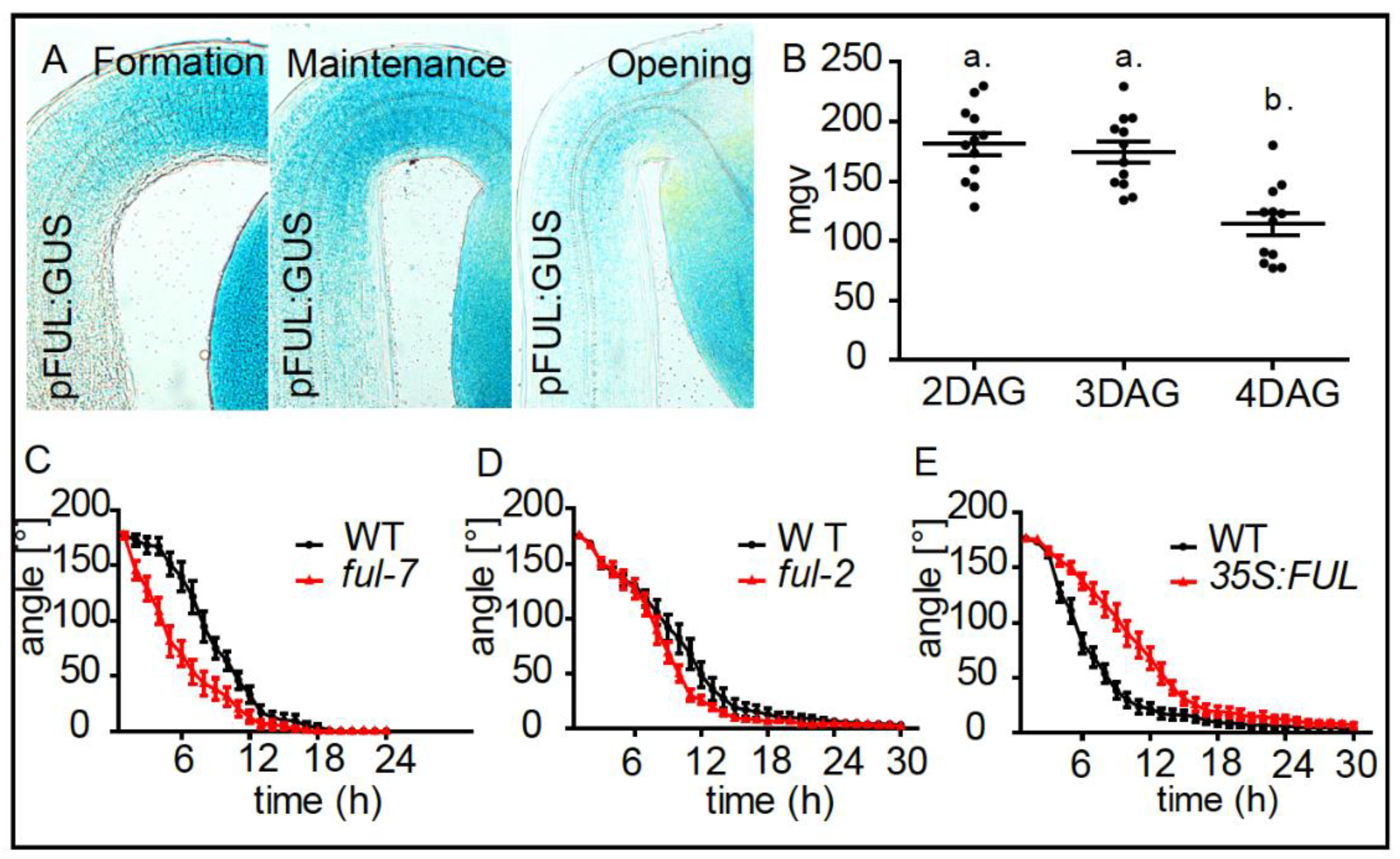
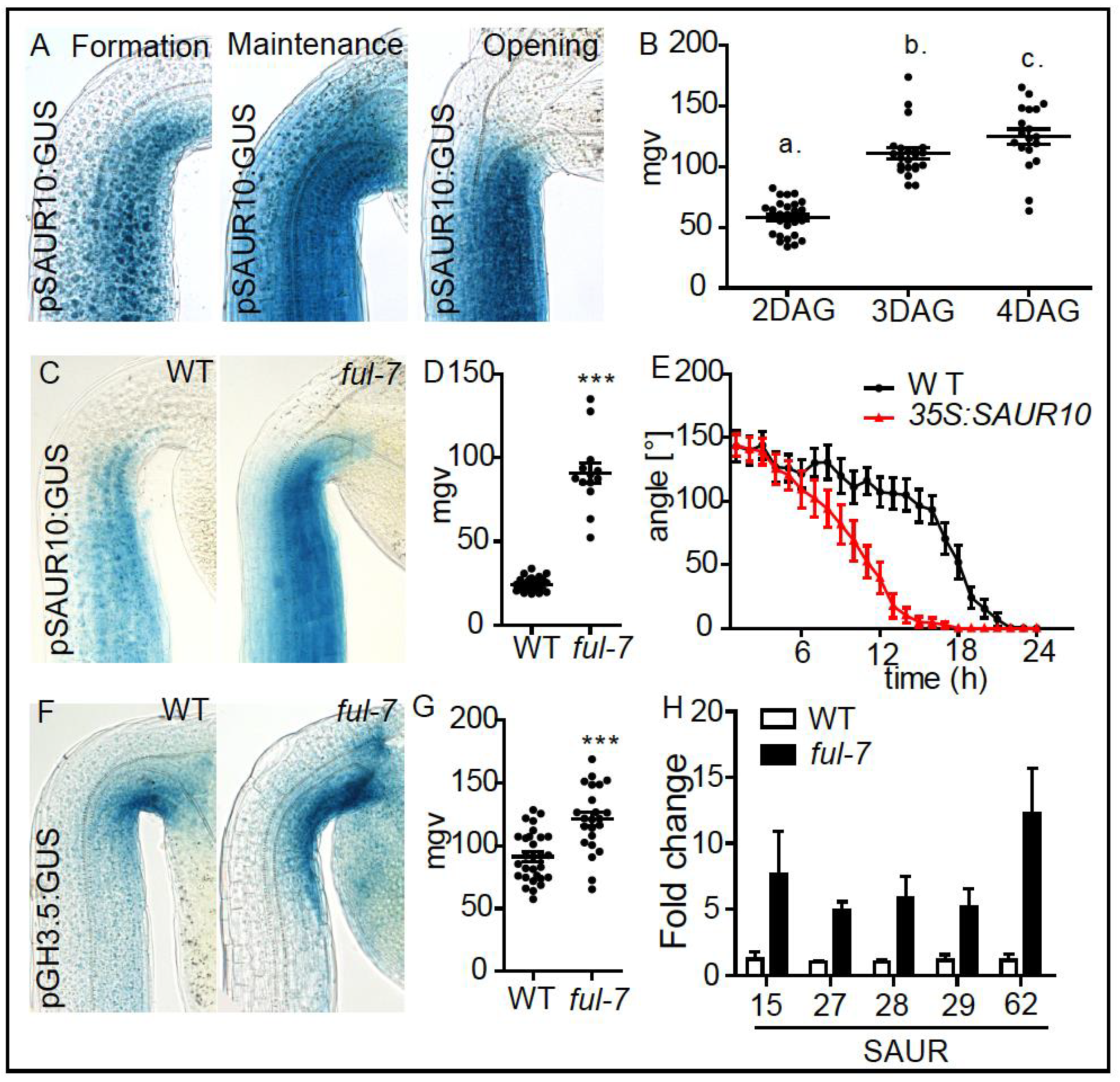
2. Results and Discussion
3. Material and Methods
3.1. Plant Material
3.2. Growth Conditions
3.3. Expression Analysis via GUS Staining
3.4. Time-Lapse Imaging of Apical Hook Kinetics
3.5. Confocal Microscopy
3.6. RNA Sequencing
3.7. qRT-PCR
3.8. In Silico Analysis
3.9. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AGL | AGAMOUS-LIKE |
| FUL | FRUITFULL |
| IAA | indole-3-acetic-acid |
| DAG | days after germination |
| mgv | mean gray value |
| FC | fold change |
| DEG | differentially expressed genes |
| 3 DAG | 3 days after germination |
References
- Abbas, M.; Alabadi, D.; Blázquez, M.A. Differential growth at the apical hook: All roads lead to auxin. Front. Plant Sci. 2013, 4, 441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Béziat, C.; Barbez, E.; Feraru, M.I.; Lucyshyn, D.; Kleine-Vehn, J. Light triggers PILS-dependent reduction in nuclear auxin signaling for growth transition. Nat. Plants 2017, 3, 17105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzella, M.A.; Casal, J.J.; Muschietti, J.; Fox, A.R. Hormonal networks involved in apical hook development in darkness and their response to light. Front. Plant Sci. 2014, 5, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raz, V.; Ecker, J.R. Regulation of differential growth in the apical hook of Arabidopsis. Development 1999, 126, 3661–3668. [Google Scholar] [PubMed]
- Boutté, Y.; Crosnier, M.-T.; Carraro, N.; Traas, J.; Satiat-Jeunemaitre, B. The plasma membrane recycling pathway and cell polarity in plants: Studies on PIN proteins. J. Cell Sci. 2006, 119, 1255–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandenbussche, F.; Petrasek, J.; Žádníková, P.; Hoyerová, K.; Pešek, B.; Raz, V.; Swarup, R.; Bennett, M.; Zažímalová, E.; Benkova, E.; et al. The auxin influx carriers AUX1 and LAX3 are involved in auxin-ethylene interactions during apical hook development in Arabidopsis thaliana seedlings. Development 2010, 137, 597–606. [Google Scholar] [CrossRef] [Green Version]
- Zadnikova, P.; Petrasek, J.; Marhavý, P.; Raz, V.; Vandenbussche, F.; Ding, Z.; Schwarzerová, K.; Morita, M.T.; Tasaka, M.; Hejatko, J.; et al. Role of PIN-mediated auxin efflux in apical hook development of Arabidopsis thaliana. Development 2010, 137, 607–617. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.; Gallemi, M.; Pospisil, J.; Zadnikova, P.; Strnad, M.; Benkova, E. Root gravity response module guides differential growth determining both root bending and apical hook formation in Arabidopsis. Development 2019, 146, dev175919. [Google Scholar] [CrossRef] [Green Version]
- Jonsson, K.; Boutté, Y.; Singh, R.K.; Gendre, D.; Bhalerao, R.P. Ethylene Regulates Differential Growth via BIG ARF-GEF-Dependent Post-Golgi Secretory Trafficking in Arabidopsis. Plant Cell 2017, 29, 1039–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Béziat, C.; Kleine-Vehn, J. The Road to Auxin-Dependent Growth Repression and Promotion in Apical Hooks. Curr. Biol. 2018, 28, 519–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, M.; Chen, R.; Li, P.; Yu, Y.; Zheng, R.; Ge, D.; Zheng, W.; Wang, X.; Gu, Y.; Gelová, Z.; et al. TMK1-mediated auxin signaling regulates differential growth of the apical hook. Nature 2019, 568, 240–243. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-E.; Seo, P.J.; Lee, A.-K.; Jung, J.-H.; Kim, Y.-S.; Park, C.-M. An Arabidopsis GH3 Gene, Encoding an Auxin-Conjugating Enzyme, Mediates Phytochrome B-Regulated Light Signals in Hypocotyl Growth. Plant Cell Physiol. 2007, 48, 1236–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.-E.; Park, J.-Y.; Kim, Y.-S.; Staswick, P.E.; Jeon, J.; Yun, J.; Kim, S.-Y.; Kim, J.; Lee, Y.-H.; Park, C.-M. GH3-mediated Auxin Homeostasis Links Growth Regulation with Stress Adaptation Response in Arabidopsis. J. Boil. Chem. 2007, 282, 10036–10046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staswick, P.; Serban, B.; Rowe, M.; Tiryaki, I.; Maldonado, M.T.; Maldonado, M.C.; Suza, W. Characterization of an Arabidopsis Enzyme Family That Conjugates Amino Acids to Indole-3-Acetic Acid. Plant Cell 2005, 17, 616–627. [Google Scholar] [CrossRef] [Green Version]
- Rosquete, M.R.; Barbez, E.; Kleine-Vehn, J. Cellular Auxin Homeostasis: Gatekeeping Is Housekeeping. Mol. Plant 2012, 5, 772–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, M.G.; Walker, P.L.; Pulgar-Vidal, N.C.; Belmonte, M.F. SeqEnrich: A tool to predict transcription factor networks from co-expressed Arabidopsis and Brassica napus gene sets. PLoS ONE 2017, 12, e0178256. [Google Scholar] [CrossRef]
- Kawakatsu, T.; Huang, S.-S.C.; Jupe, F.; Sasaki, E.; Schmitz, R.J.; Urich, M.A.; Castanon, R.; Nery, J.R.; Barragan, C.; He, Y.; et al. Epigenomic diversity in a global collection of Arabidopsis thaliana accessions. Cell 2016, 166, 492–505. [Google Scholar] [CrossRef] [Green Version]
- De Smet, I.; Lau, S.; Ehrismann, J.S.; Axiotis, I.; Kolb, M.; Kientz, M.; Weijers, D.; Jürgens, G. Transcriptional repression of BODENLOS by HD-ZIP transcription factor HB5 in Arabidopsis thaliana. J. Exp. Bot. 2013, 64, 3009–3019. [Google Scholar] [CrossRef] [Green Version]
- Stamm, P.; Topham, A.T.; Mukhtar, N.K.; Jackson, M.D.; Tomé, D.F.; Beynon, J.; Bassel, G.W. The Transcription Factor ATHB5 Affects GA-Mediated Plasticity in Hypocotyl Cell Growth during Seed Germination. Plant Physiol. 2016, 173, 907–917. [Google Scholar] [CrossRef] [Green Version]
- Garceau, D.; Batson, M.K.; Pan, I.L. Variations on a theme in fruit development: The PLE lineage of MADS-box genes in tomato (TAGL1) and other species. Planta 2017, 246, 313–321. [Google Scholar] [CrossRef]
- Theissen, G.; Saedler, H. Plant biology. Floral quartets. Nature 2001, 409, 469–471. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Li, H.; Wen, J.; Mysore, K.S.; Wang, X.-B.; Pei, Y.; Niu, L.; Lin, H. Functional Specialization of Duplicated AGAMOUS Homologs in Regulating Floral Organ Development of Medicago truncatula. Front. Plant Sci. 2018, 9, 854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueiredo, D.D.; Batista, R.A.; Roszak, P.; Köhler, C. Auxin production couples endosperm development to fertilization. Nat. Plants 2015, 1, 15184. [Google Scholar] [CrossRef] [PubMed]
- Balanza, V.; Martínez-Fernández, I.; Sato, S.; Yanofsky, M.F.; Kaufmann, K.; Angenent, G.C.; Bemer, M.; Ferrandiz, C. Genetic control of meristem arrest and life span in Arabidopsis by a FRUITFULL-APETALA2 pathway. Nat. Commun. 2018, 9, 565. [Google Scholar] [CrossRef] [Green Version]
- Bemer, M.; Van Mourik, H.; Muiño, J.M.; Ferrandiz, C.; Kaufmann, K.; Angenent, G.C. FRUITFULL controls SAUR10 expression and regulates Arabidopsis growth and architecture. J. Exp. Bot. 2017, 68, 3391–3403. [Google Scholar] [CrossRef] [Green Version]
- Gu, Q.; Ferrándiz, C.; Yanofsky, M.F.; Martienssen, R. The FRUITFULL MADS-box gene mediates cell differentiation during Arabidopsis fruit development. Development 1998, 125, 1509–1517. [Google Scholar]
- Kaufmann, K.; Muiño, J.M.; Jáuregui, R.; Airoldi, C.A.; Smaczniak, C.; Krajewski, P.; Angenent, G.C. Target Genes of the MADS Transcription Factor SEPALLATA3: Integration of Developmental and Hormonal Pathways in the Arabidopsis Flower. PLoS Boil. 2009, 7, e1000090. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Q.; Zheng, Y.; Ji, H.; Burnie, W.; Perry, S.E. Gene Regulation by the AGL15 Transcription Factor Reveals Hormone Interactions in Somatic Embryogenesis. Plant Physiol. 2016, 172, 2374–2387. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Ren, N.; Wang, H.; Stromberg, A.J.; Perry, S.E. Global Identification of Targets of the Arabidopsis MADS Domain Protein AGAMOUS-Like15. Plant Cell 2009, 21, 2563–2577. [Google Scholar] [CrossRef] [Green Version]
- Shikata, H.; Hanada, K.; Ushijima, T.; Nakashima, M.; Suzuki, Y.; Matsushita, T. Phytochrome controls alternative splicing to mediate light responses in Arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 18781–18786. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, L.; Mongélard, G.; Floková, K.; Pacurar, D.I.; Novak, O.; Staswick, P.; Kowalczyk, M.; Păcurar, M.; Demailly, H.; Geiss, G.; et al. Auxin Controls Arabidopsis Adventitious Root Initiation by Regulating Jasmonic Acid Homeostasis. Plant Cell 2012, 24, 2515–2527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mashiguchi, K.; Tanaka, K.; Sakai, T.; Sugawara, S.; Kawaide, H.; Natsume, M.; Hanada, A.; Yaeno, T.; Shirasu, K.; Yao, H.; et al. The main auxin biosynthesis pathway in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 18512–18517. [Google Scholar] [CrossRef] [Green Version]
- Ferrándiz, C.; Gu, Q.; Martienssen, R.; Yanofsky, M.F. Redundant regulation of meristem identity and plant architecture by FRUITFULL, APETALA1 and CAULIFLOWER. Development 2000, 127, 725–734. [Google Scholar] [PubMed]
- Béziat, C.; Kleine-Vehn, J.; Feraru, E. Histochemical staining of β-glucuronidase and its spatial quantification. Methods Mol. Biol 2017, 1497, 73–80. [Google Scholar]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Boil. 2013, 14, R36. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapnell, C.; Hendrickson, D.G.; Sauvageau, M.; Goff, L.; Rinn, J.L.; Pachter, L. Differential analysis of gene regulation at transcript resolution with RNA-seq. Nat. Biotechnol. 2012, 31, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.-J.; Wu, S.-H.; Wu, J.-F.; Lin, W.-D.; Wu, Y.-C.; Tsai, T.-Y.; Tsai, H.-L.; Wu, S.-H. Translational Landscape of Photomorphogenic Arabidopsis. Plant Cell 2013, 25, 3699–3710. [Google Scholar] [CrossRef] [Green Version]
- Hruz, T.; Laule, O.; Szabo, G.; Wessendorp, F.; Bleuler, S.; Oertle, L.; Widmayer, P.; Gruissem, W.; Zimmermann, P. Genevestigator V3: A Reference Expression Database for the Meta-Analysis of Transcriptomes. Adv. Bioinform. 2008, 2008, 420747. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Julkowska, M.M.; Saade, S.; Agarwal, G.; Gao, G.; Pailles, Y.; Morton, M.; Awlia, M.; Tester, M. MV app-multivariate analysis application for streamlined data analysis and curation. Plant Physiol. 2019, 180, 1261–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DEG | Up-Regulated | Down-Regulated | |
|---|---|---|---|
| WT + mock vs. WT + IAA | 241 | 159 | 82 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Führer, M.; Gaidora, A.; Venhuizen, P.; Dobrogojski, J.; Béziat, C.; Feraru, M.I.; Kleine-Vehn, J.; Kalyna, M.; Barbez, E. FRUITFULL Is a Repressor of Apical Hook Opening in Arabidopsis thaliana. Int. J. Mol. Sci. 2020, 21, 6438. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176438
Führer M, Gaidora A, Venhuizen P, Dobrogojski J, Béziat C, Feraru MI, Kleine-Vehn J, Kalyna M, Barbez E. FRUITFULL Is a Repressor of Apical Hook Opening in Arabidopsis thaliana. International Journal of Molecular Sciences. 2020; 21(17):6438. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176438
Chicago/Turabian StyleFührer, Miriam, Angelika Gaidora, Peter Venhuizen, Jedrzej Dobrogojski, Chloé Béziat, Mugurel I Feraru, Jürgen Kleine-Vehn, Maria Kalyna, and Elke Barbez. 2020. "FRUITFULL Is a Repressor of Apical Hook Opening in Arabidopsis thaliana" International Journal of Molecular Sciences 21, no. 17: 6438. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176438