Hyperspectral Reflectance of Light-Adapted Leaves Can Predict Both Dark- and Light-Adapted Chl Fluorescence Parameters, and the Effects of Chronic Ozone Exposure on Date Palm (Phoenix dactylifera)

 ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
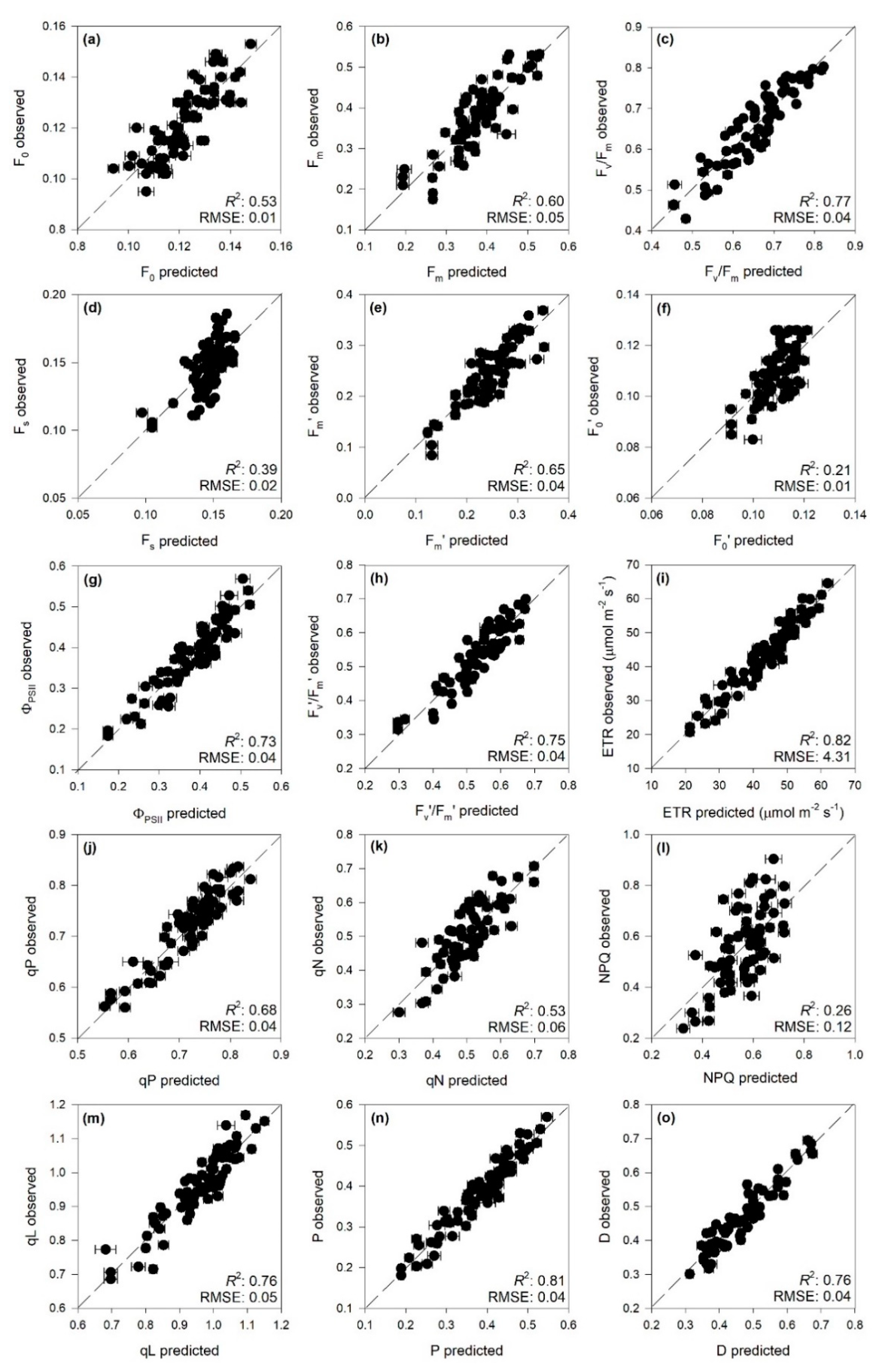
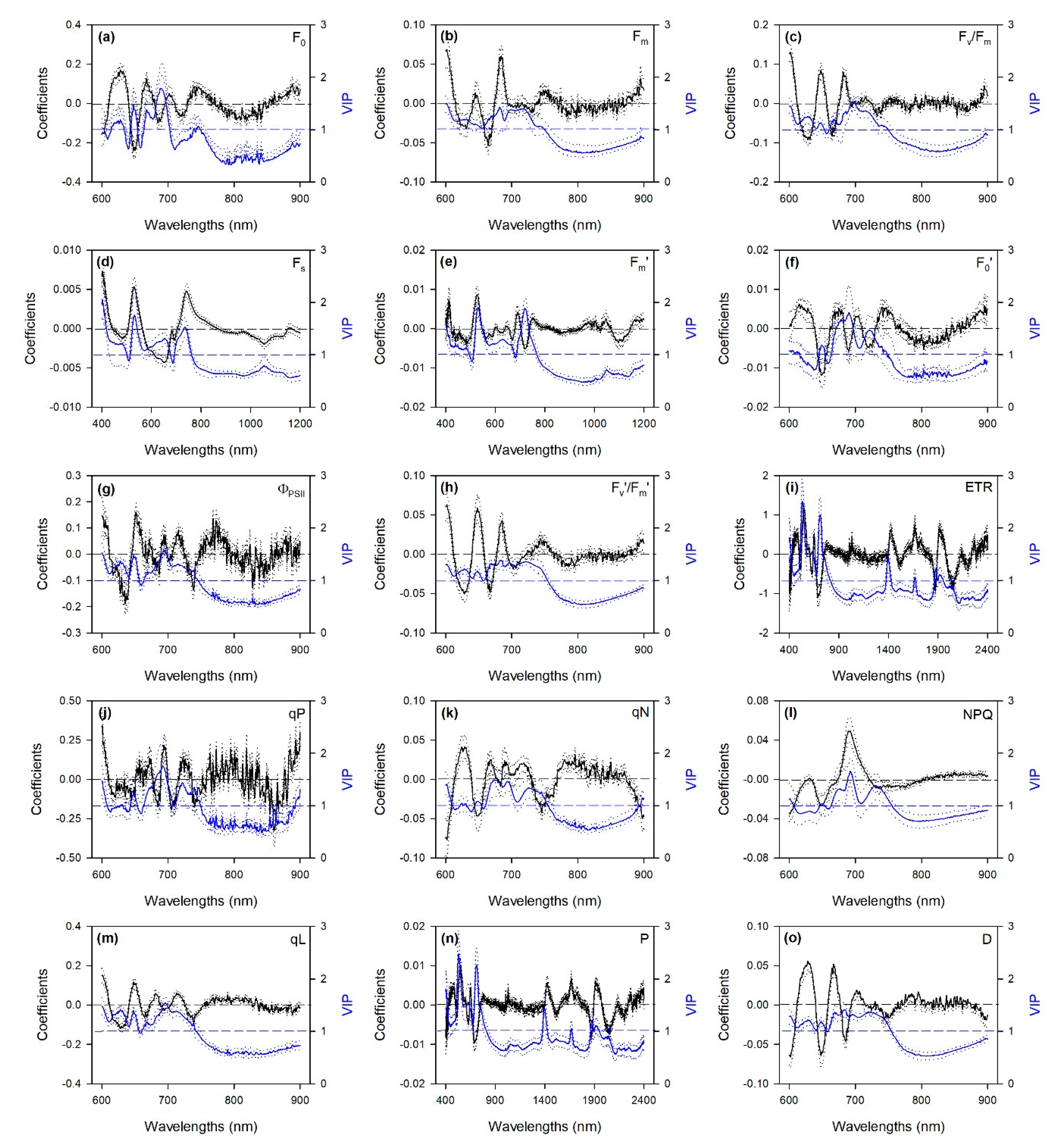
2.1. Predictions of ChlF Parameters
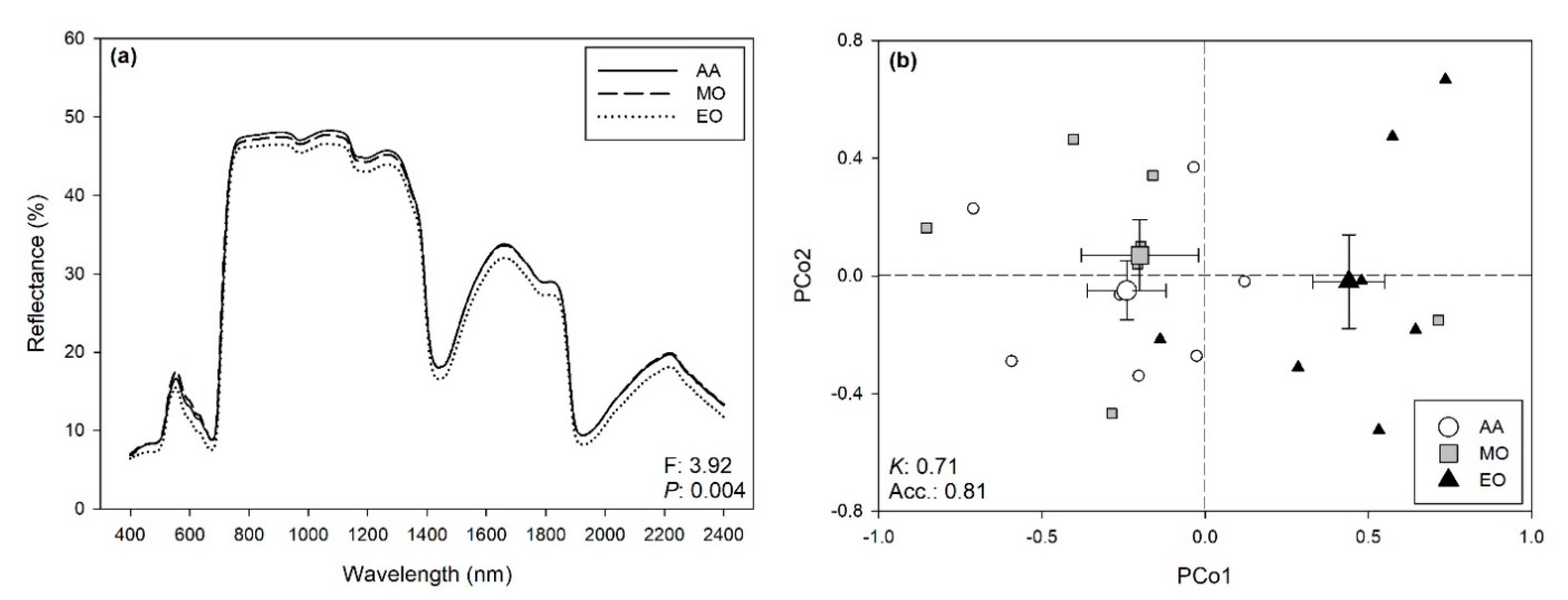
2.2. Hyperspectral Phenotyping
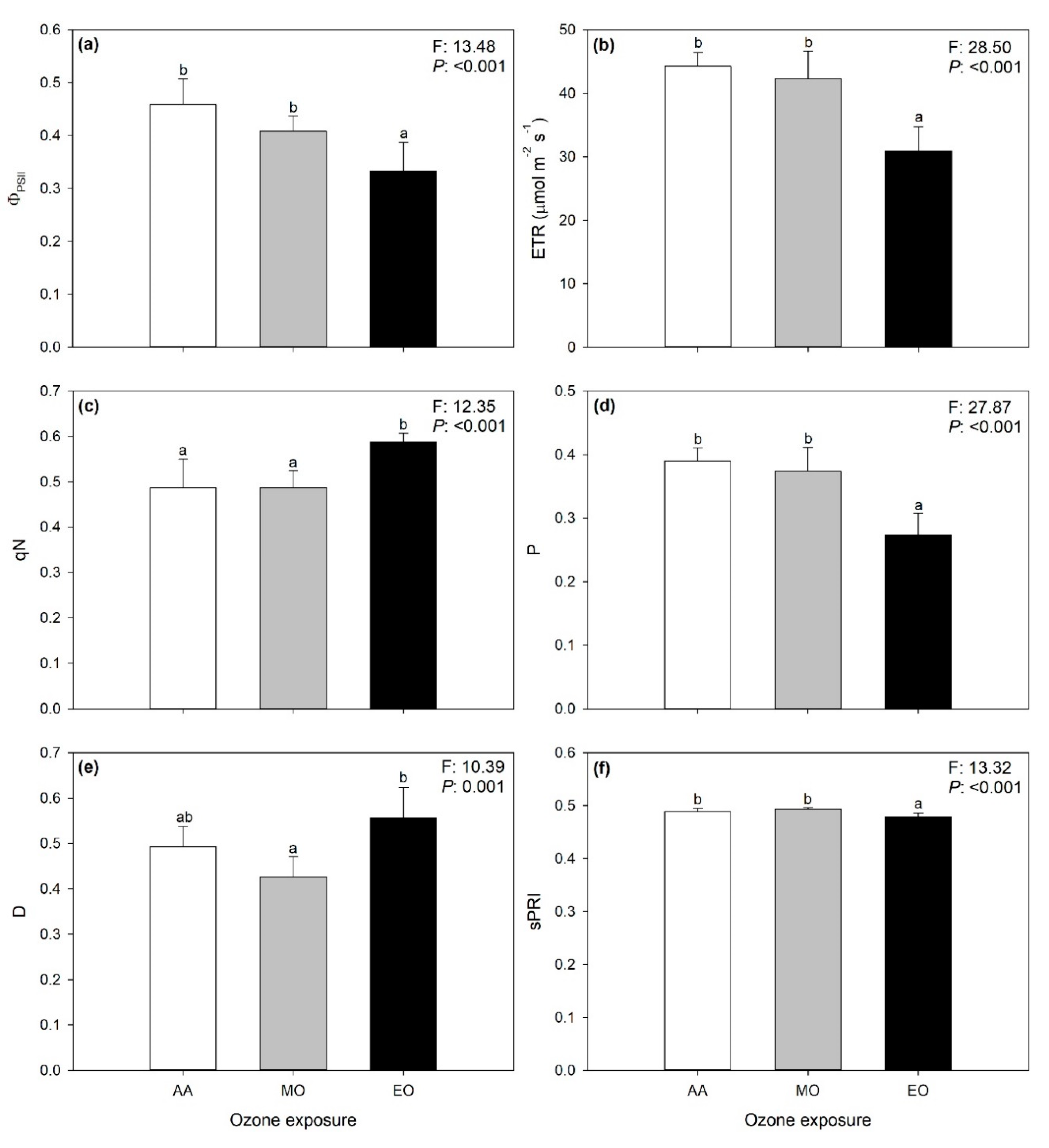
2.3. Variations of Spectra-Estimated ChlF Parameters and VIs
3. Discussion
4. Materials and Methods
4.1. Plant Material and Experimental Design
4.2. Chlorophyll a Fluorescence
4.3. Hyperspectral Reflectance
4.4. PLSR-model Calibration and Validations
4.5. Estimation of Leaf Traits by Vegetation Indices and PLSR Models
4.6. Statistical Analyses for the Effects of Ozone on Leaf Traits Derived from Spectra, and on the Leaf Spectral Signatures (i.e., Hyperspectral Phenotyping)
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | Ambient air |
| ANOVA | Analysis of variance |
| AOT40 | Accumulated exposure over a threshold of 40 ppb |
| ChlF | Chlorophyll a fluorescence |
| CI | Chlorophyll index |
| D | Fraction of light absorbed in PSII antennae that is dissipated thermally |
| EO | Elevated ozone |
| ETR | Electron transport rate |
| F0 | Minimum fluorescence yield in the dark-adapted state |
| F0′ | Minimum fluorescence intensity in the light-adapted state |
| Fm | Maximum fluorescence yield in the dark-adapted state |
| Fm’ | Maximum fluorescence intensity in the light-adapted state |
| Fs | Steady state fluorescence intensity in light-adapted state |
| Fv/Fm | Maximum quantum efficiency of PSII photochemistry |
| Fv’/Fm’ | Maximum efficiency of PSII in light conditions |
| FACE | Free air controlled exposure |
| MO | Moderate ozone |
| NDVI | Normalized difference vegetation index |
| NIR | Near-infrared spectral region |
| NPQ | Non-photochemical quenching calculated as (Fm-Fm’)/Fm’ |
| O3 | Ozone |
| P | Fraction of light absorbed in PSII antennae that is utilized in PSII photochemistry |
| PAM | Pulse-amplitude-modulation |
| PCoA | Principal coordinates analysis |
| PERMANOVA | Permutational analysis of variance |
| PLS-DA | Partial least squares discriminant analysis |
| PLSR | Partial least squares regression |
| PRESS | Predicted residual sum of squares |
| PRI | Photochemical reflectance index |
| PSI | Photosystem I |
| PSII | Photosystem II |
| PSRI | Plant senescence reflectance index |
| qL | Fraction of open PSII centers |
| qN | Non-photochemical quenching calculated as (Fm-Fm’)/(Fm-F0′) |
| qP | Photochemical quenching |
| R2 | Model goodness-of-fit |
| RMSE | Root mean square error |
| sPRI | Scaled photochemical reflectance index |
| sPSRI | Scaled plant senescence reflectance index |
| SWIR | Short-wave infrared spectral region |
| VIs | Vegetation indices |
| VIP | Variable importance to the projection |
| VIS | Visible spectral region |
| ΦPSII | PSII operating efficiency in light conditions |
| %RMSE | Percent root mean square error of the data range |
References
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence – A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schreiber, U. Pulse-Amplitude-Modulation (PAM) fluorometry and saturation pulse method: An overview. In Chlorophyll a Fluorescence: A Signature of Photosynthesis; Papageorgiou, G.C., Govindjee, Eds.; Springer: Amsterdam, The Netherlands, 2004; pp. 279–319. [Google Scholar]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porcar-Castell, A.; Pfuendel, E.; Korhonen, J.F.J.; Juurola, E. A new monitoring PAM fluorometer (MONI-PAM) to study the short and long-term acclimation of photosystem II in field conditions. Photosynth. Res. 2008, 96, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Guidi, L.E.; Degl’Innocenti, E. Imaging of Chlorophyll a fluorescence: A tool to Study Abiotic Stress in Plants. In Abiotic Stress in Plants – Mechanisms and Adaptations; Shanker, A., Venkateswarlu, B., Eds.; IntechOpen: Rijeka, Croatia, 2011; pp. 3–20. [Google Scholar]
- Magney, T.S.; Frankenberg, C.; Fisher, J.B.; Sun, Y.; North, G.B.; Davis, T.S.; Kornfeld, A.; Siebke, K. Connecting active to passive fluorescence with photosynthesis: A method for evaluating remote sensing measurements of Chl fluorescence. New Phytol. 2017, 215, 1594–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotrozzi, L.; Couture, J.J. Hyperspectral assessment of plant responses to multi-stress environments: Prospects for managing protected agrosystems. Plants People Planet 2020, 2, 244–258. [Google Scholar] [CrossRef]
- Cotrozzi, L.; Townsend, P.A.; Pellegrini, E.; Nali, C.; Couture, J.J. Reflectance spectroscopy: A novel approach to better understand and monitor the impact of air pollution on Mediterranean plants. Environ. Sci. Pollut. Res. 2018, 25, 8249–8267. [Google Scholar] [CrossRef]
- Gamon, J.A.; Field, C.B.; Goulden, M.L.; Griffin, K.L.; Hartley, A.E.; Joel, G.; Peñuelas, J.; Valentini, R. Relationships between NDVI, canopy structure, and photosynthesis in three Californian vegetation types. Ecol. Appl. 1995, 5, 28–41. [Google Scholar] [CrossRef] [Green Version]
- Gitelson, A.A.; Merzlyak, M.N. Quantitative estimation of chlorophyll a using reflectance spectra—Experiments with autumn chestnut and maple leaves. J. Photochem. Photobiol. 1994, 22, 247–252. [Google Scholar] [CrossRef]
- Wold, S.; Sjöström, M.; Eriksson, L. PLS-regression: A basic tool of chemometrics. Chemom. Intell. Lab. Syst. 2001, 58, 109–130. [Google Scholar] [CrossRef]
- Serbin, S.P.; Singh, A.; Desai, A.R.; Dubois, S.G.; Jablosnki, A.D.; Kingdon, C.C.; Kruger, E.L.; Townsend, P.A. Remotely estimating photosynthetic capacity, and its response to temperature, in vegetation canopies using imaging spectroscopy. Remote Sens. Environ. 2015, 167, 78–87. [Google Scholar] [CrossRef]
- Couture, J.J.; Singh, A.; Rubert-Nason, K.F.; Serbin, S.P.; Lindroth, R.L.; Townsend, P.A. Spectroscopic determination of ecologically relevant plant secondary metabolites. Methods Ecol. Evol. 2016, 7, 1402–1412. [Google Scholar] [CrossRef] [Green Version]
- Cotrozzi, L.; Couture, J.J.; Cavender-Bares, J.; Kingdon, C.C.; Fallon, B.; Pilz, G.; Pellegrini, E.; Nali, C.; Townsend, P.A. Using foliar spectral properties to assess the effects of drought on plant water potential. Tree Physiol. 2017, 37, 1582–1591. [Google Scholar] [CrossRef] [PubMed]
- Ely, K.S.; Burnett, A.C.; Lieberman-Cribbin, W.; Serbin, S.P.; Rogers, A. Spectroscopy can predict key leaf traits associated with source-sink balance and carbon-nitrogen status. J. Exp. Bot. 2019, 70, 1789–1799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchica, A.; Loré, S.; Cotrozzi, L.; Lorenzini, G.; Nali, C.; Pellegrini, E.; Remorini, D. Early detection of sage (Salvia officinalis L.) responses to ozone using reflectance spectroscopy. Plants 2019, 8, 346. [Google Scholar] [CrossRef] [Green Version]
- Couture, J.J.; Lindroth, R.L. Atmospheric change alters performance of an invasive forest insect. Glob. Chang. Biol. 2012, 18, 3543–3557. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Miller, J.R.; Mohammed, G.H.; Noland, T.L.; Sampson, P.H. Estimation of chlorophyll fluorescence under natural illumination from hyperspectral data. Int. J. Appl. Earth Obs. Geoinf. 2001, 3, 321–327. [Google Scholar] [CrossRef] [Green Version]
- Jia, M.; Li, D.; Colombo, R.; Wang, Y.; Wang, X.; Cheng, T.; Zhu, Y.; Yao, X.; Xu, C.; Ouer, G.; et al. Quantifying chlorophyll fluorescence parameters from hyperspectral reflectance at the leaf scale under various nitrogen treatment regimes in winter wheat. Remote Sens. 2019, 11, 2838. [Google Scholar] [CrossRef] [Green Version]
- Zohari, D.; Hopf, M.; Weiss, E. Domestication of Plants in the Old World: The Origin and Spread of Domesticated Plants in South-West Asia, Europe and the Mediterranean Basin, 4th ed.; Oxford University Press: Oxford, UK, 2012; p. 264. [Google Scholar]
- Hassan, I.; Cotrozzi, L.; Haiba, N.S.; Basahi, J.; Ismail, I.; Almeelbi, T.; Hammam, E. Trace elements in the fruits of date palm (Phoenix dactylifera L.) in Jeddah City, Saudi Arabia. Agrochimica 2017, 61, 75–93. [Google Scholar]
- Arab, L.; Kreuzwieser, J.; Kruse, J.; Zimmer, I.; Ache, P.; Alfarraj, S.; Al-Rasheid, K.A.; Schnitzler, J.P.; Hedrich, R.; Rennenberg, H. Acclimation to heat and drought–lessons to learn from the date palm (Phoenix dactylifera). Environ. Exp. Bot. 2016, 125, 20–30. [Google Scholar] [CrossRef] [Green Version]
- Safronov, O.; Kreuzweiser, J.; Haberer, G.; Alyousif, M.S.; Schulze, W.; Al-Harbi, N.; Arab, L.; Ache, P.; Stempfl, T.; Kruse, J.; et al. Detecting early signs of heat and drought stress in Phoenix dactylifera (date palm). PLoS ONE 2017, 12, e0177883. [Google Scholar] [CrossRef] [PubMed]
- Kruse, J.; Adams, M.; Winkler, B.; Ghirardo, A.; Alfarraj, S.; Kreuzweiser, J.; Hedrich, R.; Schnitzler, J.-P.; Rennenberg, H. Optimization of photosynthesis and stomatal conductance in the date palm Phoenix dactylifera during acclimation to heat and drought. New Phytol. 2019, 223, 1973–1988. [Google Scholar] [CrossRef] [PubMed]
- Du, B.; Kruse, J.; Winkler, J.B.; Schnitzler, J.P.; Ache, P.; Hedrich, R.; Rennenberg, H. Climate and development modulate the metabolome and antioxidative system of date palm leaves. J. Exp. Bot. 2019, 70, 5959–5969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, B.; Kreuzweiser, J.; Winkler, J.B.; Ghirardo, A.; Schnitzler, J.P.; Ache, P.; Alfarraj, S.; Hedrich, R.; White, P.; Rennenberg, H. Physiological responses of date palm (Phoenix dactylifera) seedlings to acute ozone exposure at high temperature. Environ. Pollut. 2018, 242, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Paoletti, E.; Hoshika, Y.; Arab, L.; Martini, S.; Cotrozzi, L.; Weber, D.; Ache, P.; Neri, L.; Baraldi, R.; Pellegrini, E.; et al. Date palm responses to realistic ozone exposure in a FACE experiment. Manuscript in preparation.
- Ainsworth, E.A. Understanding and improving global crop response to ozone pollution. Plant J. 2017, 90, 886–897. [Google Scholar] [CrossRef] [PubMed]
- Bhuiyan, M.A.; Khan, H.U.R.; Zaman, K.; Hishan, S.S. Measuring the impact of global tropospheric ozone, carbon dioxide and sulfur dioxide concentrations on biodiversity loss. Environ. Res. 2018, 160, 398–411. [Google Scholar] [CrossRef]
- Nuvolone, D.; Petri, D.; Voller, F. The effects of ozone on human health. Environ. Sci. Pollut. Res. 2018, 25, 8074–8088. [Google Scholar] [CrossRef]
- Lefohn, A.S.; Malley, C.S.; Smith, L.; Wells, B.; Hazucha, M.; Simon, H.; Naik, V.; Mills, G.; Schultz, M.G.; Paoletti, E. Tropospheric ozone assessment report: Global ozone metrics for climate change, human health, and crop/ecosystem research. Elementa 2018, 6, 28. [Google Scholar] [CrossRef] [Green Version]
- Ochoa-Hueso, R.; Munzi, S.; Alonso, R.; Arróniz-Crespo, M.; Avila, A.; Bermejo, V.; Bobbink, R.; Branquinho, C.; Concostrina-Zubiri, L.; Cruz, C. Ecological impacts of atmospheric pollution and interactions with climate change in terrestrial ecosystems of the Mediterranean Basin: Current research and future directions. Environ. Pollut. 2017, 227, 194–206. [Google Scholar] [CrossRef] [Green Version]
- Sicard, P.; De Marco, A.; Troussier, F.; Renou, C.; Vas, N.; Paoletti, E. Decrease in surface ozone concentrations at Mediterranean remote sites and increase in the cities. Atmos. Environ. 2013, 79, 705–715. [Google Scholar] [CrossRef]
- Feng, Z.; De Marco, A.; Anav, A.; Gualtieri, M.; Sicard, P.; Tian, H.; Fornasier, F.; Tao, F.; Guo, A.; Paoletti, E. Economic losses due to ozone impacts on human health, forest productivity and crop yield across China. Environ. Internat. 2019, 131, 104966. [Google Scholar] [CrossRef]
- Guidi, L.; Nali, C.; Ciompi, S.; Lorenzini, G.; Soldatini, G.F. The use of chlorophyll fluorescence and leaf gas exchange as methods for studying the different responses to ozone of two bean cultivars. J. Exp. Bot. 1997, 48, 173–179. [Google Scholar] [CrossRef] [Green Version]
- Cotrozzi, L.; Remorini, D.; Pellegrini, E.; Guidi, L.; Nali, C.; Lorenzini, G.; Massai, R.; Landi, M. Living in a Mediterranean city in 2050: Broadleaf or evergreen ‘citizens’? Environ. Sci. Pollut. Res. 2018, 25, 8161–8173. [Google Scholar] [CrossRef] [PubMed]
- Landi, M.; Cotrozzi, L.; Pellegrini, E.; Remorini, D.; Tonelli, M.; Trivellini, A.; Nali, C.; Guidi, L.; Massai, R.; Vernieri, P.; et al. When “thirsty” means “less able to activate the signalling wave trigged by a pulse of ozone”: A case of study in two Mediterranean deciduous oak species with different drought sensitivity. Sci. Total Environ. 2019, 657, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Gamon, J.A.; Serrano, L.; Surfus, J.S. The photochemical reflectance index: An optical indicator of photosynthetic radiation use efficiency across species, functional types, and nutrient levels. Oecologia 1997, 112, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J.; Garbulsky, M.F.; Filella, I. Photochemical reflectance index (PRI) and remote sensing of plant CO2 uptake. New Phytol. 2011, 191, 596–599. [Google Scholar]
- Serbin, S.P.; Dillaway, D.N.; Kruger, E.L.; Townsend, P.A. Leaf optical properties reflect variation in photosynthetic metabolism and its sensitivity to temperature. J. Exp. Bot. 2012, 63, 489–502. [Google Scholar] [CrossRef] [Green Version]
- Heckmann, D.; Schlüter, U.; Weber, A.P.M. Machine learning techniques for predicting crop photosynthetic capacity from leaf reflectance spectra. Mol. Plant 2017, 10, 878–890. [Google Scholar] [CrossRef]
- Yendrek, C.R.; Tomaz, T.; Montes, C.M.; Cao, Y.; Morse, A.M.; Brown, P.J.; McIntyre, L.M.; Leakey, A.D.B.; Ainsworth, E.A. High-throughput phenotyping maize leaf physiological and biochemical traits using hyperspectral reflectance. Plant Physiol. 2017, 173, 614–626. [Google Scholar] [CrossRef]
- Fu, P.; Meacham-Hensold, K.; Guan, K.; Wu, J.; Bernacchi, C. Estimating photosynthesis traits from reflectance spectra: A synthesis of spectral indices, numerical inversion, and partial least square regression. Plant Cell Environ. 2020, 43, 1241–1258. [Google Scholar] [CrossRef]
- Merzlyak, M.N.; Gitelson, A.A.; Chivkunova, O.B.; Solovchenko, A.E.; Pogosyan, S.I. Application of reflectance spectroscopy for analysis of higher plant pigments. Russ. J. Plant Physiol. 2003, 50, 704–710. [Google Scholar] [CrossRef]
- Pedrós, R.; Moya, I.; Goulas, Y.; Jacquemond, S. Chlorophyll fluorescence emission spectrum inside a leaf. Photochem. Photobiol. Sci. 2008, 7, 498–502. [Google Scholar]
- Mutanga, O.; Skidmore, A.K. Red edge shift and biochemical content in grass canopies. ISPRS J. Photogramm. Remote Sens. 2007, 62, 34–42. [Google Scholar] [CrossRef]
- Smith, K.L.; Steven, M.D.; Colls, J.J. Use of hyperspectral derivative tools in red-edge region to identify plant stress response to gas leaks. Remote Sens. Environ. 2004, 92, 207–212. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Miller, J.R.; Morales, A.; Berjón, A.; Agüera, J. Hyperspectral indices and model simulation for chlorophyll estimation in open-canopy tree crops. Remote Sens. Environ. 2004, 9, 463–476. [Google Scholar] [CrossRef]
- Curran, P.J. Remote sensing of foliar chemistry. Remote Sens. Environ. 1989, 30, 271–278. [Google Scholar] [CrossRef]
- Campos-Medina, V.A.; Cotrozzi, L.; Stuart, J.J.; Couture, J.J. Spectral characterization of wheat functional trait responses to Hessian fly: Mechanisms for trait-based resistance. PLoS ONE 2019, 14, e0219431. [Google Scholar] [CrossRef] [Green Version]
- Gongora-Canul, C.; Salgado, J.D.; Singh, D.; Cruz, A.P.; Cotrozzi, L.; Couture, J.; Rivadeneira, M.G.; Cruppe, G.; Valent, B.; Todd, T.; et al. Temporal dynamics of wheat blast epidemics and disease measurements using multispectral imagery. Phytopathology 2020, 110, 393–405. [Google Scholar] [CrossRef]
- Meacham-Hensold, K.; Montes, C.M.; Wu, J.; Guan, K.; Fu, P.; Ainsworth, E.A.; Pederson, T.; Moore, C.E.; Brown, K.L.; Raines, C.; et al. High-throughput field phenotyping using hyperspectral reflectance and partial least squares regression (PLSR) reveals genetic modifications to photosynthetic capacity. Remote Sens. Environ. 2019, 231, 111176. [Google Scholar] [CrossRef]
- Paoletti, E.; Materassi, A.; Fasano, G.; Hoshika, Y.; Carriero, G.; Silaghi, D.; Badea, O. A new-generation 3D ozone Face (Free Air Controlled Exposure). Sci. Total Environ. 2017, 575, 1407–1414. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.-M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Adama, W.W., III; Barker, D.H.; Logan, B.A.; Bowling, D.R.; Verhoeven, A.S. Using chlorophyll fluorescence to assess the fraction of absorbed light allocated to thermal dissipation of excess excitation. Physiol. Plant. 1996, 98, 253–264. [Google Scholar] [CrossRef]
- Björkman, O.; Demmig, B. Photon yield of O2-evolution and chloroplast fluorescence characteristics at 77 K among vascular plants of diverse origins. Planta 1987, 170, 489–504. [Google Scholar]
- Schreiber, U.; Bilger, W.; Neubauer, C. Chlorophyll fluorescence as a non-intrusive indicator for a rapid assessment of in vivo photosynthesis. In Ecophysiology of Photosynthesis. Series Ecological Studies; Schulze, E.-D., Caldwell, M.M., Eds.; Springer-Verlag: Berlin, Germany, 1994; Volume 100, pp. 49–70. [Google Scholar]
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New fluorescence parameters for the determination of QA redox state and excitation energy fluxes. Photosynth. Res. 2004, 79, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Atzberger, C.; Guerif, M.; Baret, F.; Werner, W. Comparative analysis of three chemometric techniques for the spectrometric assessment of canopy chlorophyll content in winter wheat. Comput. Electron. Agric. 2010, 73, 165–173. [Google Scholar] [CrossRef]
- Grossman, Y.L.; Ustin, S.L.; Jacquemond, S.; Sanderson, E.W.; Schmuck, G.; Verdebout, J. Critique of stepwise multiple linear regression for the extraction of leaf biochemistry information from leaf reflectance data. Remote Sens. Environ. 1996, 56, 182–193. [Google Scholar] [CrossRef]
- Chen, S.; Hong, X.; Harris, C.J.; Sharkey, P.M. Sparse modeling using orthogonal forest regression with PRESS statistic and regularization. IEEE T. Syst. Man Cy. B 2004, 34, 898–911. [Google Scholar] [CrossRef]
- Chong, I.-G.; Jun, C.-H. Performance of some variable selection methods when multicollinearity is present. Chemom. Intell. Lab. Syst. 2005, 28, 103–112. [Google Scholar] [CrossRef]
- Mevik, B.H.; Wehrrens, R.; Liland, K.H. pls: Partial Least Squares and Principal Component Regression. R package version 2.6-0. 2016. Available online: https://CRAN.R-project.org/package=pls (accessed on 31 July 2020).
- Merzlyak, M.N.; Gitelson, A.A.; Chivkunova, O.B.; Rakitin, V.Y. Non-destructive optical detection of pigment changes during leaf senescence and fruit ripening. Physiol. Plant. 1999, 106, 135–141. [Google Scholar] [CrossRef] [Green Version]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar]
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Chevallier, S.; Bertand, D.; Kohler, A.; Courcoux, P. Application of PLS-DA in multivariate image analysis. J. Chemom. 2006, 20, 221–229. [Google Scholar] [CrossRef]
- Kuhn, M. Building predictive models in R using the caret package. J. Stat. Softw. 2008, 28, 5. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Range (nm) | Com | Cal | Val | ||||
|---|---|---|---|---|---|---|---|---|
| R2 | RMSE | %RMSE | R2 | RMSE | %RMSE | |||
| F0 | 600–900 | 8 | 0.69 ± 0.03 | 0.01 ± 0.00 | 13 | 0.53 ± 0.14 | 0.01 ± 0.00 | 17 |
| Fm | 600–900 | 7 | 0.74 ± 0.03 | 0.04 ± 0.00 | 12 | 0.60 ± 0.15 | 0.05 ± 0.01 | 15 |
| Fv/Fm | 600–900 | 8 | 0.85 ± 0.02 | 0.04 ± 0.00 | 11 | 0.77 ± 0.08 | 0.04 ± 0.01 | 11 |
| Fs | 400–1200 | 4 | 0.46 ± 0.04 | 0.01 ± 0.00 | 17 | 0.39 ± 0.18 | 0.02 ± 0.00 | 18 |
| Fm’ | 400–1200 | 8 | 0.80 ± 0.03 | 0.03 ± 0.00 | 9 | 0.65 ± 0.14 | 0.04 ± 0.01 | 12 |
| F0′ | 600–900 | 7 | 0.47 ± 0.05 | 0.01 ± 0.00 | 17 | 0.21 ± 0.19 | 0.01 ± 0.00 | 21 |
| ΦPSII | 600–900 | 10 | 0.87 ± 0.03 | 0.03 ± 0.00 | 8 | 0.73 ± 0.12 | 0.04 ± 0.01 | 11 |
| Fv’/Fm’ | 600–900 | 7 | 0.84 ± 0.02 | 0.04 ± 0.00 | 9 | 0.75 ± 0.10 | 0.04 ± 0.01 | 11 |
| ETR | 400–2400 | 12 | 0.93 ± 0.01 | 2.63 ± 0.15 | 6 | 0.82 ± 0.08 | 4.31 ± 0.68 | 10 |
| qP | 600–900 | 11 | 0.87 ± 0.02 | 0.02 ± 0.00 | 9 | 0.68 ± 0.14 | 0.04 ± 0.01 | 14 |
| qN | 600–900 | 7 | 0.70 ± 0.04 | 0.05 ± 0.00 | 12 | 0.53 ± 0.17 | 0.06 ± 0.01 | 15 |
| NPQ | 600–900 | 6 | 0.40 ± 0.05 | 0.11 ± 0.00 | 16 | 0.26 ± 0.19 | 0.12 ± 0.02 | 18 |
| qL | 600–900 | 8 | 0.86 ± 0.02 | 0.04 ± 0.00 | 8 | 0.76 ± 0.12 | 0.05 ± 0.01 | 11 |
| P | 400–2400 | 12 | 0.93 ± 0.01 | 0.02 ± 0.00 | 6 | 0.81 ± 0.08 | 0.04 ± 0.01 | 10 |
| D | 600–900 | 7 | 0.84 ± 0.02 | 0.04 ± 0.00 | 7 | 0.76 ± 0.11 | 0.04 ± 0.01 | 10 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cotrozzi, L.; Lorenzini, G.; Nali, C.; Pellegrini, E.; Saponaro, V.; Hoshika, Y.; Arab, L.; Rennenberg, H.; Paoletti, E. Hyperspectral Reflectance of Light-Adapted Leaves Can Predict Both Dark- and Light-Adapted Chl Fluorescence Parameters, and the Effects of Chronic Ozone Exposure on Date Palm (Phoenix dactylifera). Int. J. Mol. Sci. 2020, 21, 6441. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176441
Cotrozzi L, Lorenzini G, Nali C, Pellegrini E, Saponaro V, Hoshika Y, Arab L, Rennenberg H, Paoletti E. Hyperspectral Reflectance of Light-Adapted Leaves Can Predict Both Dark- and Light-Adapted Chl Fluorescence Parameters, and the Effects of Chronic Ozone Exposure on Date Palm (Phoenix dactylifera). International Journal of Molecular Sciences. 2020; 21(17):6441. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176441
Chicago/Turabian StyleCotrozzi, Lorenzo, Giacomo Lorenzini, Cristina Nali, Elisa Pellegrini, Vincenzo Saponaro, Yasutomo Hoshika, Leila Arab, Heinz Rennenberg, and Elena Paoletti. 2020. "Hyperspectral Reflectance of Light-Adapted Leaves Can Predict Both Dark- and Light-Adapted Chl Fluorescence Parameters, and the Effects of Chronic Ozone Exposure on Date Palm (Phoenix dactylifera)" International Journal of Molecular Sciences 21, no. 17: 6441. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176441