Transcriptome Analysis of Testis from HFD-Induced Obese Rats (Rattus norvigicus) Indicated Predisposition for Male Infertility

 , ,
, ,  ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Quality of Obtained Data
2.1.1. Distribution of Detected Genes Counts
2.1.2. Distribution of Detected Gene Expression Level
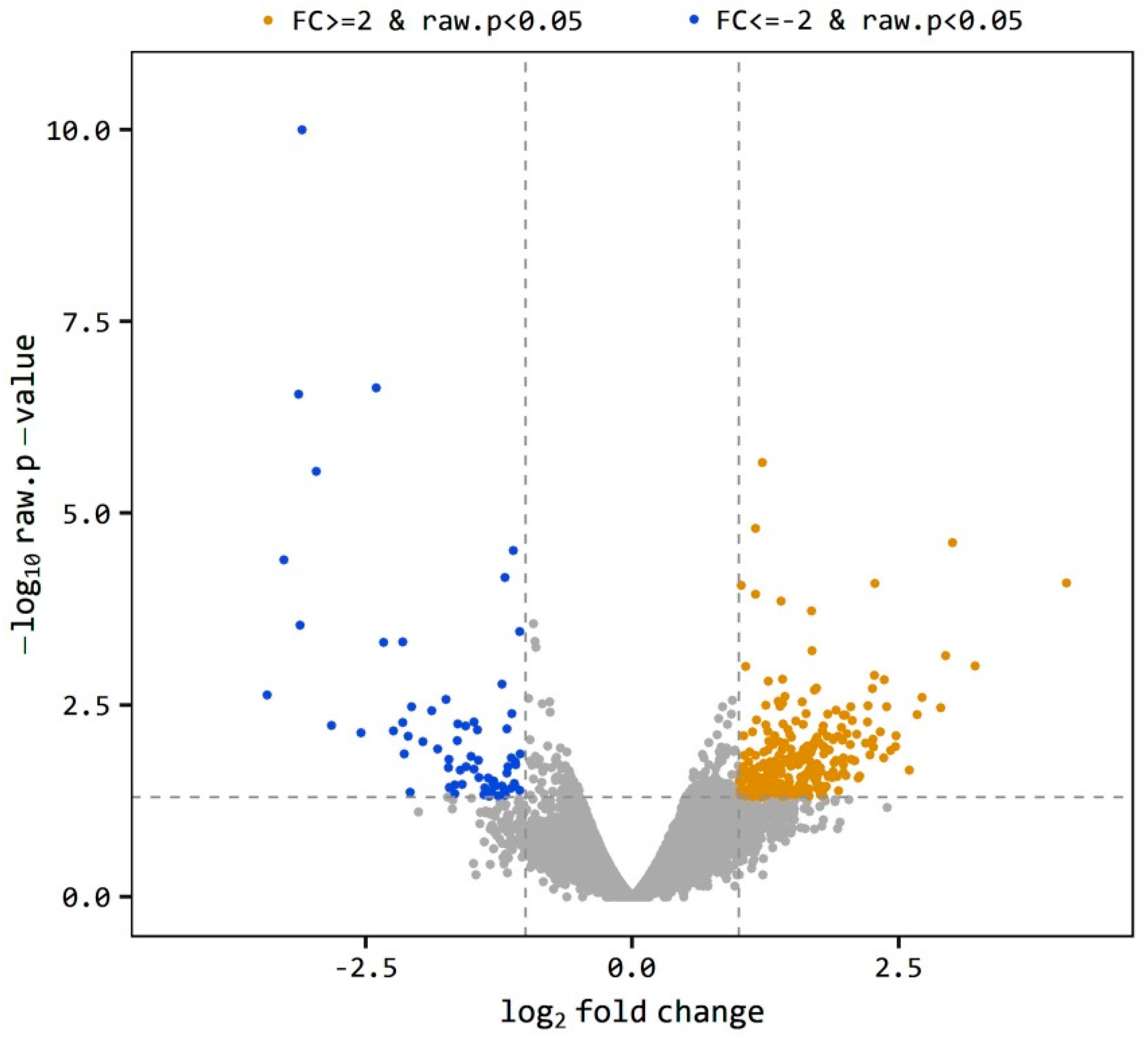
2.1.3. Differentially Expressed Genes (Degs)
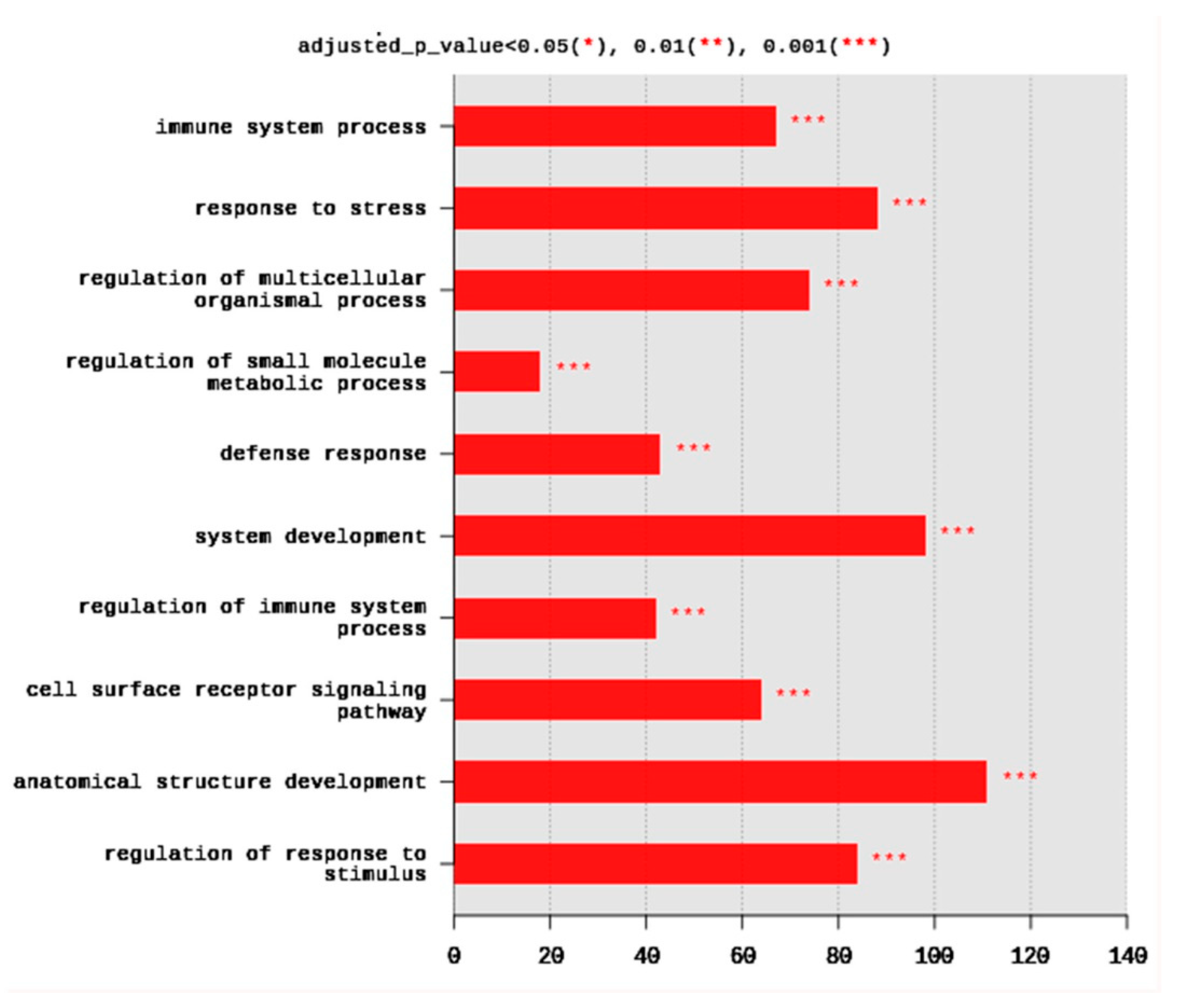
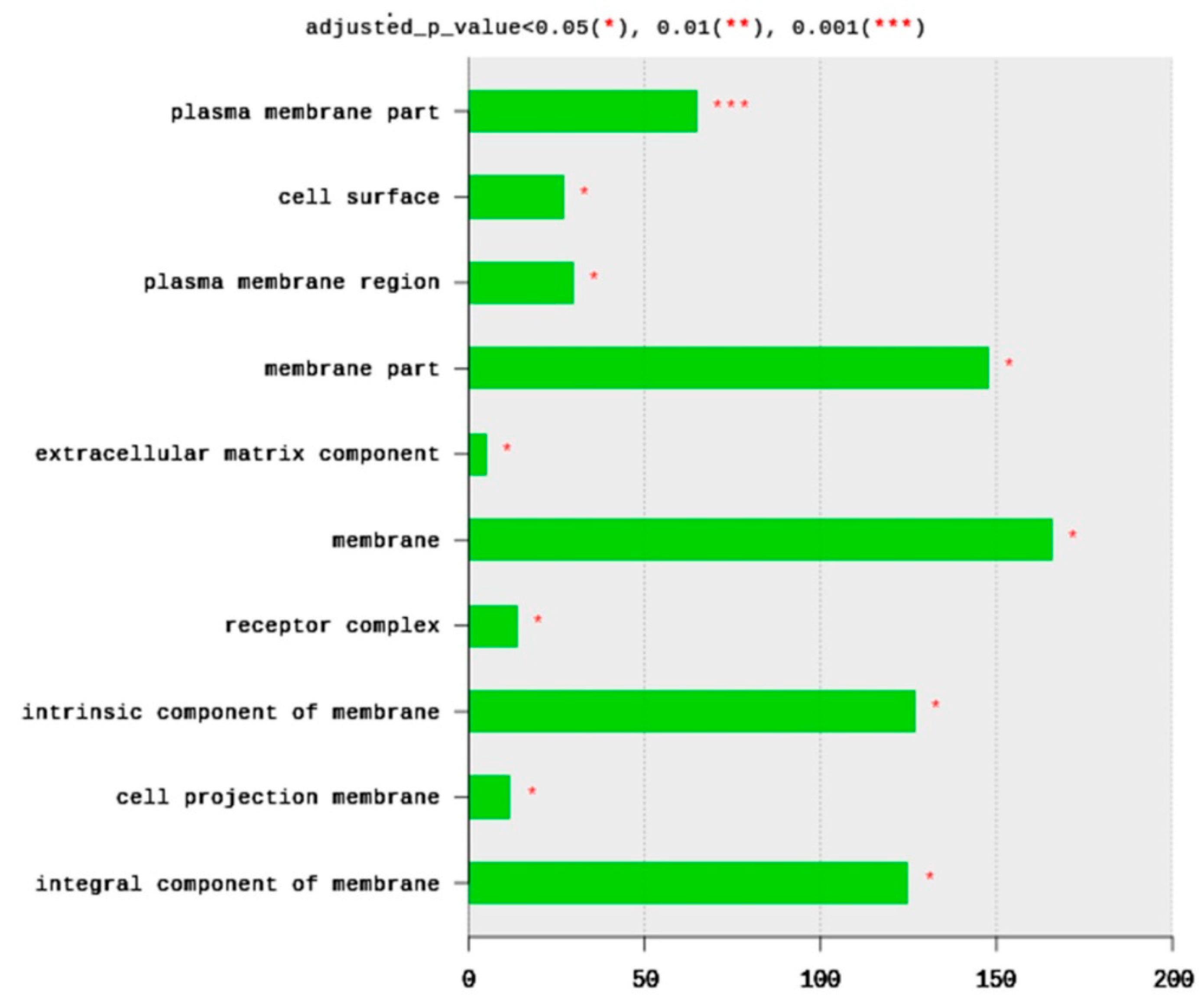
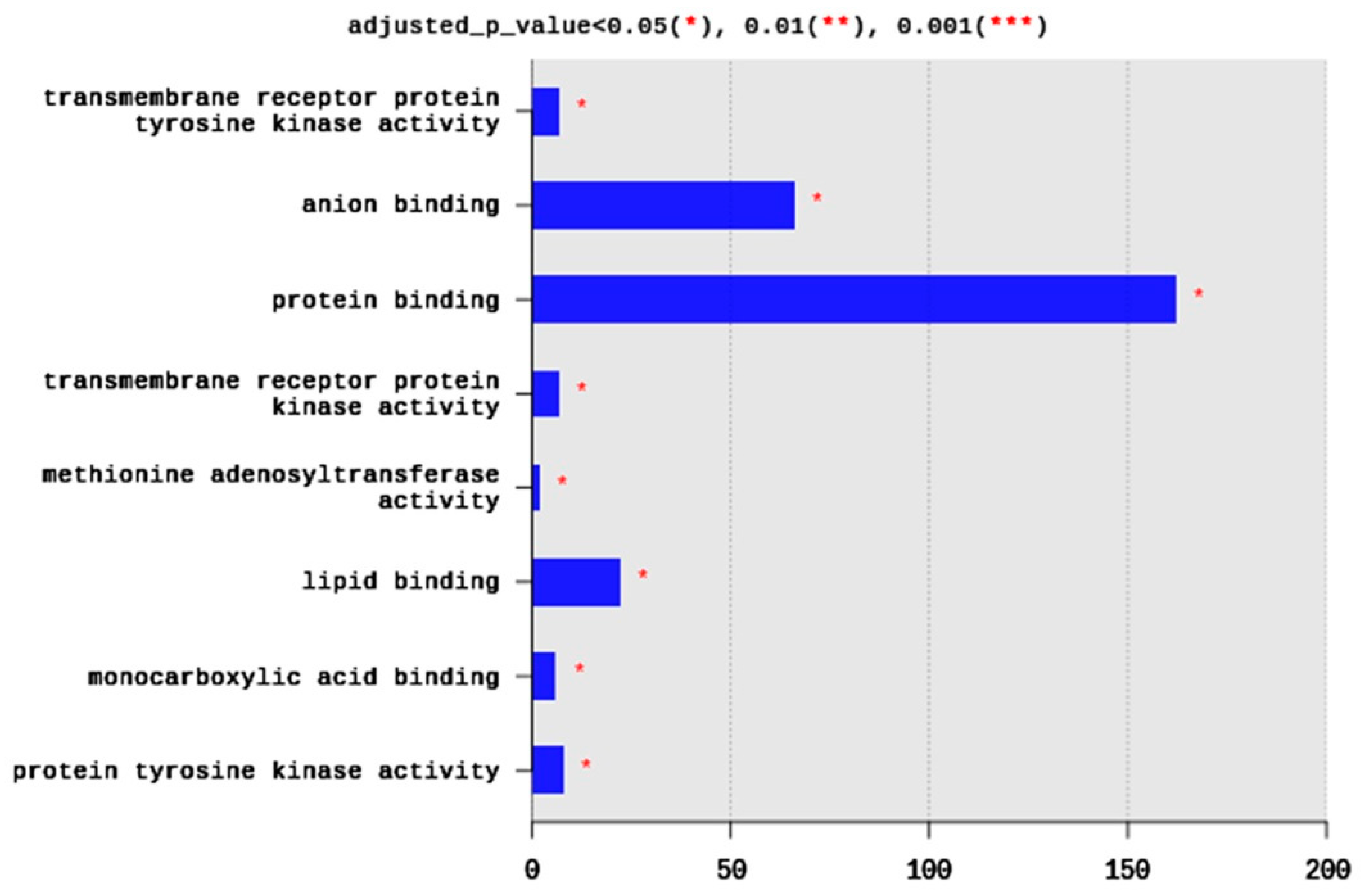
2.1.4. Gene Ontology (GO)
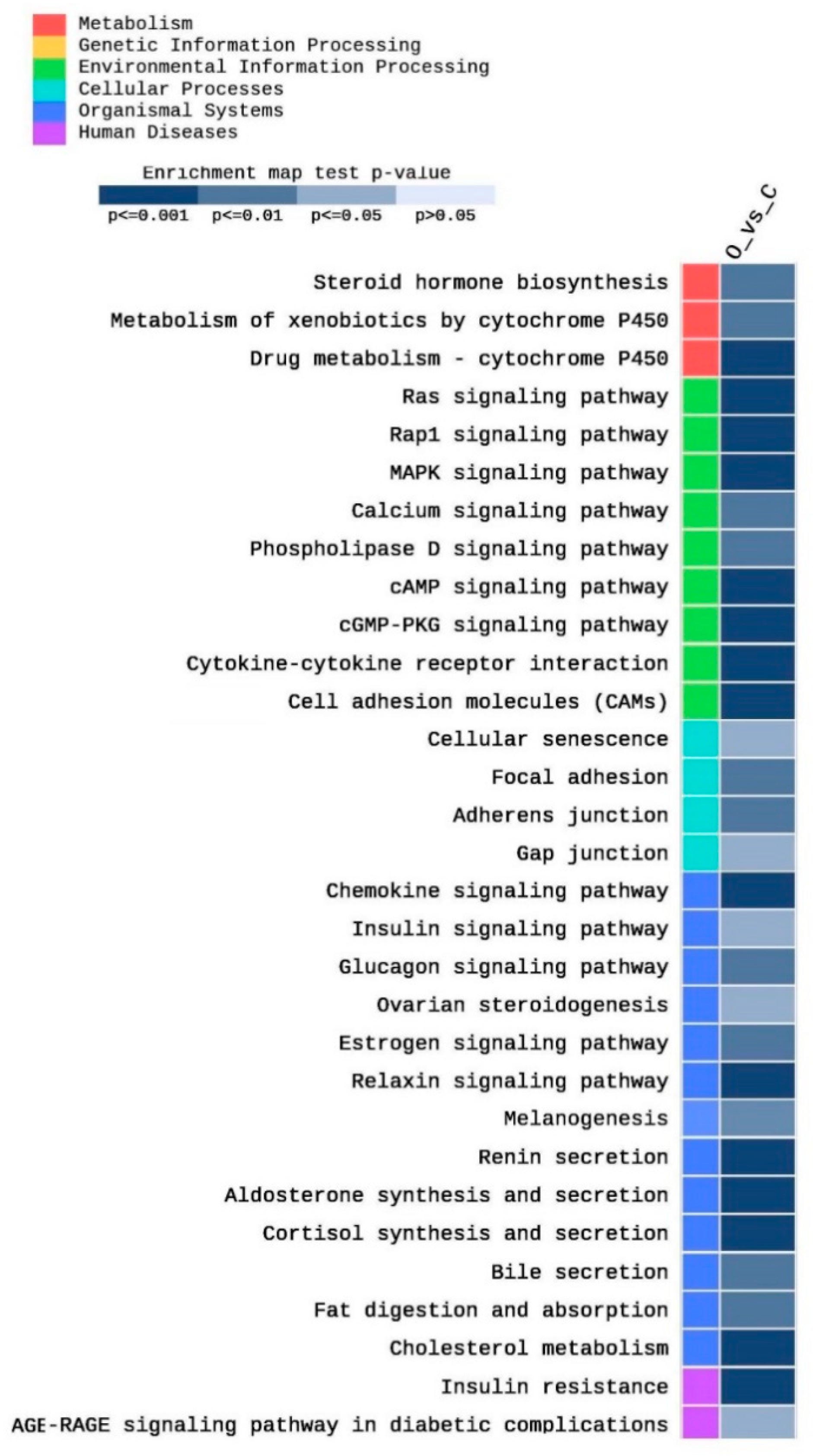
2.2. KEGG Analysis
2.3. Gene Expression
2.3.1. Obesity and Lipid Metabolism
2.3.2. Obesity Marker Genes (OM)
2.3.3. Spermatogenesis, Reproduction, Fertilization
2.3.4. Cytochrome P450 (CYPs)
2.3.5. Cell Adhesion
2.3.6. Immune Response
2.3.7. Olfaction
3. Discussion
3.1. KEGG Pathways
3.2. Gene Expression Analysis
3.2.1. Lipid Metabolism and Control of Obesity
3.2.2. Spermatogenesis and Fertilization
3.2.3. Cell Adhesion and Fertility
3.2.4. Olfaction and Olfactory Receptors
3.2.5. CYPs expression
3.2.6. Immune Response
4. Materials and Methods
4.1. Animals and Experimental Design
4.2. Sampling
4.3. RNA Isolation and cDNA Library Construction
4.4. Transcriptome Sequencing and Assembly
4.5. Transcriptome Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Fan, W.; Xu, Y.; Liu, Y.; Zhang, Z.; Lu, L.; Ding, Z. Obesity or Overweight, a Chronic Inflammatory Status in Male Reproductive System, Leads to Mice and Human Subfertility. Front. Physiol. 2018, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F.; et al. Global, regional, and national prevalence of overweight and obesity in children and adults during 1980–2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2014, 384, 776–781. [Google Scholar] [CrossRef] [Green Version]
- World, Health, Organization. WHO Fact Sheet No 311. Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 1 April 2020).
- Kyrou, I.; Randeva, H.S.; Tsigos, C.; Kaltsas, G.; Weickert, M.O. Clinical Problems Caused by Obesity; Feingold, K.R., Anawalt, B., Boyce, A., Eds.; Endotext MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Bakos, H.W.; Mitchell, M.; Setchell, B.P.; Lane, M. The effect of paternal diet-induced obesity on sperm function and fertilization in a mouse model. Int. J. Androl. 2010, 34, 402–410. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.; Gaffen, S.L. IL-17 in obesity and adipogenesis. Cytokine Growth Factor Rev. 2010, 21, 449–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, R.; Ji, M.; Zhang, S. Global warming and obesity: A systematic review. Obes. Rev. 2017, 19, 150–163. [Google Scholar] [CrossRef]
- Yang, X.-Y.; Gu, Y.-J.; An, T.; Liu, J.-X.; Pan, Y.-Y.; Mo, F.-F.; Miao, J.-N.; Zhao, D.-D.; Zhang, D.; Gao, S.-H.; et al. Proteomics Analysis of Testis of Rats Fed a High-Fat Diet. Cell. Physiol. Biochem. 2018, 47, 378–389. [Google Scholar] [CrossRef]
- Tilg, H.; Moschen, A.R. Adipocytokines: Mediators linking adipose tissue, inflammation and immunity. Nat. Rev. Immunol. 2006, 6, 772–783. [Google Scholar] [CrossRef]
- Moraes, J.C.; Coope, A.; Morari, J.; Cintra, D.E.C.; Roman, E.A.; Pauli, J.R.; Romanatto, T.; Carvalheira, J.B.; Oliveira, A.L.R.; Saad, M.J.; et al. High-Fat Diet Induces Apoptosis of Hypothalamic Neurons. PLoS ONE 2009, 4, e5045. [Google Scholar] [CrossRef]
- Buffone, M.G.; Verstraeten, S.V.; Calamera, J.C.; Doncel, G.F. High Cholesterol Content and Decreased Membrane Fluidity in Human Spermatozoa Are Associated With Protein Tyrosine Phosphorylation and Functional Deficiencies. J. Androl. 2009, 30, 552–558. [Google Scholar] [CrossRef]
- Chambers, T.J.G.; Anderson, R.A. The impact of obesity on male fertility. Hormones 2015, 14, 563–568. [Google Scholar] [CrossRef]
- Bieniek, J.M.; Kashanian, J.A.; Deibert, C.M.; Grober, E.D.; Lo, K.C.; Brannigan, R.E.; Sandlow, J.I.; Jarvi, K.A. Influence of increasing body mass index on semen and reproductive hormonal parameters in a multi-institutional cohort of subfertile men. Fertil. Steril. 2016, 106, 1070–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du Plessis, S.S.; Cabler, S.; McAlister, D.A.; Sabanegh, E.; Agarwal, A. The effect of obesity on sperm disorders and male infertility. Nat. Rev. Urol. 2010, 7, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Palmer, N.O.; Bakos, H.W.; Fullston, T.; Lane, M. Impact of obesity on male fertility, sperm function and molecular composition. Spermatogenesis 2012, 2, 253–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, C.D.; Bellentani, F.F.; Fernandes, G.S.; Perobelli, J.E.; Favareto, A.P.A.; Nascimento, A.F.; Cicogna, A.C.; Kempinas, W.D.G. Diet-induced obesity in rats leads to a decrease in sperm motility. Reprod. Biol. Endocrinol. 2011, 9, 32. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, G.S.A.; Arena, A.C.; De Campos, K.E.; Volpato, G.T.; Anselmo-Franci, J.A.; Damasceno, D.C.; Kempinas, W.D.G. Glutamate-induced obesity leads to decreased sperm reserves and acceleration of transit time in the epididymis of adult male rats. Reprod. Biol. Endocrinol. 2012, 10, 105. [Google Scholar] [CrossRef] [Green Version]
- Ghanayem, B.I.; Bai, R.; Kissling, G.E.; Travlos, G.; Hoffler, U. Diet-Induced Obesity in Male Mice Is Associated with Reduced Fertility and Potentiation of Acrylamide-Induced Reproductive Toxicity1. Biol. Reprod. 2010, 82, 96–104. [Google Scholar] [CrossRef] [Green Version]
- La Vignera, S.; Condorelli, R.; Vicari, E.; D’Agata, R.; Calogero, A.E. Diabetes Mellitus and Sperm Parameters. J. Androl. 2011, 33, 145–153. [Google Scholar] [CrossRef]
- Li, L.; Law, C.; Lo Conte, R.; Power, C. A Intergenerational influences on childhood body mass index: The effect of parental body mass index trajectories. Am. J. Clin. Nutr. 2009, 89, 551–557. [Google Scholar] [CrossRef]
- Chavarro, J.E.; Toth, T.L.; Wright, D.L.; Meeker, J.D.; Hauser, R. Body mass index in relation to semen quality, sperm DNA integrity, and serum reproductive hormone levels among men attending an infertility clinic. Fertil. Steril. 2010, 93, 2222–2231. [Google Scholar] [CrossRef] [Green Version]
- Wake, D.J.; Strand, M.; Rask, E.; Westerbacka, J.; Livingstone, D.E.W.; Soderberg, S.; Andrew, R.; Yki-Jarvinen, H.; Olsson, T.; Walker, B.R. Intra-adipose sex steroid metabolism and body fat distribution in idiopathic human obesity. Clin. Endocrinol. 2007, 66, 440–446. [Google Scholar] [CrossRef]
- Meinhardt, U.; Mullis, P.E. The Essential Role of the Aromatase/P450arom. Semin. Reprod. Med. 2002, 20, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Caprio, M.; Isidori, A.M.; Carta, A.R.; Moretti, C.; Dufau, M.L.; Fabbri, A. Expression of functional leptin receptors in rodent Leydig cells. Endocrinology 1999, 140, 4939–4947. [Google Scholar] [CrossRef] [PubMed]
- Klok, M.D.; Jakobsdottir, S.; Drent, M.L. The role of leptin and ghrelin in the regulation of food intake and body weight in humans: A review. Obes. Rev. 2007, 8, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Martini, A.C.; Molina, R.I.; Tissera, A.; Ruiz, R.D.; De Cuneo, M.F. The impact of obesity on male reproduction: Its biological significance. Expert Rev. Endocrinol. Metab. 2013, 8, 139–148. [Google Scholar] [CrossRef]
- Craig, J.R.; Jenkins, T.G.; Carrell, D.T.; Hotaling, J.M. Obesity, male infertility, and the sperm epigenome. Fertil. Steril. 2017, 107, 848–859. [Google Scholar] [CrossRef] [Green Version]
- Palmer, N.O.; Bakos, H.W.; Owens, J.A.; Setchell, B.P.; Lane, M. Diet and exercise in an obese mouse fed a high-fat diet improve metabolic health and reverse perturbed sperm function. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E768–E780. [Google Scholar] [CrossRef]
- Samavat, J.; Natali, I.; Degl’Innocenti, S.; Filimberti, E.; Cantini, G.; Di Franco, A.; Danza, G.; Seghieri, G.; Lucchese, M.; Baldi, E.; et al. Acrosome reaction is impaired in spermatozoa of obese men: A preliminary study. Fertil. Steril. 2014, 102, 1274–1281.e2. [Google Scholar] [CrossRef]
- Zhao, J.; Zhai, L.; Liu, Z.; Wu, S.; Xu, L. Leptin Level and Oxidative Stress Contribute to Obesity-Induced Low Testosterone in Murine Testicular Tissue. Oxidative Med. Cell. Longev. 2014, 2014, 1–14. [Google Scholar] [CrossRef]
- Duale, N.; Steffensen, I.-L.; Andersen, J.; Brevik, A.; Brunborg, G.; Lindeman, B. Impaired sperm chromatin integrity in obese mice. Andrology 2014, 2, 234–243. [Google Scholar] [CrossRef]
- Zhang, E.; Xu, F.; Liang, H.; Yan, J.; Xu, H.; Li, Z.; Wen, X.; Weng, J. GLP-1 Receptor Agonist Exenatide Attenuates the Detrimental Effects of Obesity on Inflammatory Profile in Testis and Sperm Quality in Mice. Am. J. Reprod. Immunol. 2015, 74, 457–466. [Google Scholar] [CrossRef]
- Chojnacka, K.; Bilińska, B.; Mruk, D.D. Interleukin 1alpha-induced disruption of the Sertoli cell cytoskeleton affects gap junctional communication. Cell Signal. 2016, 28, 469–480. [Google Scholar] [CrossRef]
- El-Shazly, S.A.; Ahmed, M.M.; Al-Harbi, M.S.; Alkafafy, M.E.; El-Sawy, H.B.; Amer, S.A. Physiological and molecular study on the anti-obesity effects of pineapple (Ananas comosus) juice in male Wistar rat. Food Sci. Biotechnol. 2018, 27, 1429–1438. [Google Scholar] [CrossRef]
- Guilherme, A.; Virbasius, J.V.; Puri, V.; Czech, M.P. Adipocyte dysfunctions linking obesity to insulin resistance and type 2 diabetes. Nat. Rev. Mol. Cell Biol. 2008, 9, 367–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, C.; E Murphy, K.; Fernandez, M.L. Impact of Obesity and Metabolic Syndrome on Immunity. Adv. Nutr. 2016, 7, 66–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanneganti, T.-D.; Dixit, V.D. Immunological complications of obesity. Nat. Immunol. 2012, 13, 707–712. [Google Scholar] [CrossRef] [PubMed]
- Wiernsperger, N.F. Membrane physiology as a basis for the cellular effects of metformin in insulin resistance and diabetes. Diabetes Metab. 1999, 25, 110–127. [Google Scholar]
- Perona, J.S. Membrane lipid alterations in the metabolic syndrome and the role of dietary oils. Biochim. Biophys. Acta Biomembr. 2017, 1859, 1690–1703. [Google Scholar] [CrossRef]
- Nandipati, K.C.; Subramanian, S.; Agrawal, D.K. Protein kinases: Mechanisms and downstream targets in inflammation-mediated obesity and insulin resistance. Mol. Cell. Biochem. 2016, 426, 27–45. [Google Scholar] [CrossRef] [Green Version]
- Morrison, D.K. MAP Kinase Pathways. Cold Spring Harb. Perspect. Biol. 2012, 4, a011254. [Google Scholar] [CrossRef]
- Li, M.W.M.; Mruk, D.D.; Cheng, C.Y. Mitogen-activated protein kinases in male reproductive function. Trends Mol. Med. 2009, 15, 159–168. [Google Scholar] [CrossRef] [Green Version]
- Aivatiadou, E.; Mattei, E.; Ceriani, M.; Tilia, L.; Berruti, G. Impaired Fertility and Spermiogenetic Disorders with Loss of Cell Adhesion in Male Mice Expressing an Interfering Rap1 Mutant. Mol. Biol. Cell 2007, 18, 1530–1542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viera, J.T.; El-Merahbi, R.; Nieswandt, B.; Stegner, D.; Sumara, G. Phospholipases D1 and D2 Suppress Appetite and Protect against Overweight. PLoS ONE 2016, 11, e0157607. [Google Scholar] [CrossRef]
- Song, Z.; Wang, Y.; Zhang, F.; Yao, F.; Chao, S. Calcium Signaling Pathways: Key Pathways in the Regulation of Obesity. Int. J. Mol. Sci. 2019, 20, 2768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tengholm, A.; Gylfe, E. cAMP signalling in insulin and glucagon secretion. Diabetes Obes. Metab. 2017, 19 (Suppl. 1), 42–53. [Google Scholar] [CrossRef] [Green Version]
- Sanyal, A.; Naumann, J.; Hoffmann, L.S.; Chabowska-Kita, A.; Ehrlund, A.; Schlitzer, A.; Arner, P.; Blüher, M.; Pfeifer, A. Interplay between Obesity-Induced Inflammation and cGMP Signaling in White Adipose Tissue. Cell Rep. 2017, 18, 225–236. [Google Scholar] [CrossRef] [Green Version]
- Pietrobon, M.; Zamparo, I.; Maritan, M.; Franchi, S.A.; Pozzan, T.; Lodovichi, C. Interplay among cGMP, cAMP, and Ca2+ in Living Olfactory Sensory Neurons In Vitro and In Vivo. J. Neurosci. 2011, 31, 8395–8405. [Google Scholar] [CrossRef] [Green Version]
- Akalestou, E.; Genser, L.; Rutter, G. Glucocorticoid Metabolism in Obesity and Following Weight Loss. Front. Endocrinol. 2020, 11. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Liu, Y.; Xue, K.; Gu, G.; Fan, W.; Xu, Y.; Ding, Z. Diet-Induced Obesity in Male C57BL/6 Mice Decreases Fertility as a Consequence of Disrupted Blood-Testis Barrier. PLoS ONE 2015, 10, e0120775. [Google Scholar] [CrossRef]
- Munro, J.; Skrobot, O.; Sanyoura, M.; Kay, V.; Susce, M.T.; Glaser, P.E.; De Leon, J.; Blakemore, A.I.; Arranz, M.J. Relaxin polymorphisms associated with metabolic disturbance in patients treated with antipsychotics. J. Psychopharmacol. 2011, 26, 374–379. [Google Scholar] [CrossRef]
- Randhawa, M.; Huff, T.; Valencia, J.C.; Younossi, Z.; Chandhoke, V.; Hearing, V.J.; Baranova, A. Evidence for the ectopic synthesis of melanin in human adipose tissue. FASEB J. 2008, 23, 835–843. [Google Scholar] [CrossRef] [Green Version]
- Kawarazaki, W.; Fujita, T. The Role of Aldosterone in Obesity-Related Hypertension. Am. J. Hypertens. 2016, 29, 415–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozcan, L.; Tabas, I. Calcium signalling and ER stress in insulin resistance and atherosclerosis. J. Intern. Med. 2016, 280, 457–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kay, A.M.; Simpson, C.L.; Stewart, J.A., Jr. The Role of AGE/RAGE Signaling in Diabetes-Mediated Vascular Calcification. J. Diabetes Res. 2016, 2016, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohtomo, T.; Hoshino, A.; Yajima, M.; Tsuchiya, A.; Momose, A.; Tanonaka, K.; Toyoda, H.; Kato, T.; Yamada, J. Expression and distribution of acyl-CoA thioesterases in the white adipose tissue of rats. Histochem. Cell Biol. 2013, 140, 223–232. [Google Scholar] [CrossRef]
- Ellis, J.M.; Bowman, C.E.; Wolfgang, M.J. Metabolic and Tissue-Specific Regulation of Acyl-CoA Metabolism. PLoS ONE 2015, 10, e0116587. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Hulver, M.W.; McMillan, R.P.; Cline, M.A.; Gilbert, E.R. The pivotal role of pyruvate dehydrogenase kinases in metabolic flexibility. Nutr. Metab. 2014, 11, 10. [Google Scholar] [CrossRef] [Green Version]
- Newhardt, M.; Batushansky, A.; Matsuzaki, S.; Young, Z.T.; West, M.; Chin, N.C.; Szweda, L.I.; Kinter, M.; Humphries, K.M. Enhancing cardiac glycolysis causes an increase in PDK4 content in response to short-term high-fat diet. J. Biol. Chem. 2019, 294, 16831–16845. [Google Scholar] [CrossRef] [PubMed]
- Ali, F.; Ismail, A.; Esa, N.M.; Pei, C.P.; Kersten, S. Hepatic genome-wide expression of lipid metabolism in diet-induced obesity rats treated with cocoa polyphenols. J. Funct. Foods 2015, 17, 969–978. [Google Scholar] [CrossRef]
- Gilham, D.; Labonté, E.D.; Rojas, J.C.; Jandacek, R.J.; Howles, P.N.; Hui, D.Y. Carboxyl Ester Lipase Deficiency Exacerbates Dietary Lipid Absorption Abnormalities and Resistance to Diet-induced Obesity in Pancreatic Triglyceride Lipase Knockout Mice. J. Biol. Chem. 2007, 282, 24642–24649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norata, G.D.; Tsimikas, S.; Pirillo, A.; Catapano, A.L. Apolipoprotein C-III: From Pathophysiology to Pharmacology. Trends Pharmacol. Sci. 2015, 36, 675–687. [Google Scholar] [CrossRef]
- Duivenvoorden, I.; Teusink, B.; Rensen, P.C.; Romijn, J.A.; Havekes, L.M.; Voshol, P.J. Apolipoprotein C3 Deficiency Results in Diet-Induced Obesity and Aggravated Insulin Resistance in Mice. Diabetes 2005, 54, 664–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohan, A.B. Apolipoprotein C-III: A potent modulator of hypertriglyceridemia and cardiovascular disease. Curr. Opin. Endocrinol. Diabetes Obes. 2015, 22, 119–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thuahnai, S.T.; Lund-Katz, S.; Anantharamaiah, G.M.; Williams, D.L.; Phillips, M.C. A quantitative analysis of apolipoprotein binding to SR-BI. J. Lipid Res. 2003, 44, 1132–1142. [Google Scholar] [CrossRef] [Green Version]
- Calderón, B.; Huerta, L.; Casado, M.E.; González-Casbas, J.M.; Botella-Carretero, J.; Martin, A. Morbid obesity–related changes in the expression of lipid receptors, transporters, and HSL in human sperm. J. Assist. Reprod. Genet. 2019, 36, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Shulga, Y.V.; Topham, M.K.; Epand, R.M. Regulation and Functions of Diacylglycerol Kinases. Chem. Rev. 2011, 111, 6186–6208. [Google Scholar] [CrossRef]
- Nakano, T.; Seino, K.; Wakabayashi, I.; Stafforini, D.M.; Topham, M.K.; Goto, K. Deletion of diacylglycerol kinase Ƹ confers susceptibility to obesity via reduced lipolytic activity in murine adipocytes. FASEB J. 2018, 32, 4121–4413. [Google Scholar] [CrossRef] [Green Version]
- Ochocka, A.; Pawelczyk, T. Isozymes delta of phosphoinositide-specific phospholipase Cand their role in signal transduction in the cell. Acta Biochemica Polonica 2003, 50, 4. [Google Scholar]
- Jarak, I.; Almeida, S.; Carvalho, E.; Sousa, M.; Barros, A.; Alves, M.G.; Oliveira, P. Senescence and declining reproductive potential: Insight into molecular mechanisms through testicular metabolomics. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 3388–3396. [Google Scholar] [CrossRef]
- Park, E.; Kumar, S.; Lee, B.; Kim, K.-J.; Seo, J.-E.; Choi, H.-S.; Lee, K. Estrogen receptor-related receptor γ regulates testicular steroidogenesis through direct and indirect regulation of steroidogenic gene expression. Mol. Cell. Endocrinol. 2017, 452, 15–24. [Google Scholar] [CrossRef]
- Entrez Gene2. SPAG11 Sperm Associated Antigen 11. 2019. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/gene?Db=gene&Cmd=ShowDetailView&TermToSearch=10407) (accessed on 18 July 2020).
- Ellerman, D.A.; Myles, D.G.; Primàkoff, P. A Role for Sperm Surface Protein Disulfide Isomerase Activity in Gamete Fusion: Evidence for the Participation of ERp57. Dev. Cell 2006, 10, 831–837. [Google Scholar] [CrossRef] [Green Version]
- Schorr-Lenz, A.M.; Alves, J.; Henckes, N.; Seibel, P.M.; Benham, A.M.; Bustamante-Filho, I.C.; Bustamante-Filho, I.C. GnRH immunization alters the expression and distribution of protein disulfide isomerases in the epididymis. Andrology 2016, 4, 957–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Guo, Y.; Song, N.; Fan, Y.; Li, K.; Teng, X.; Guo, Q.; Ding, Z. Proteomic pattern changes associated with obesity-induced asthenozoospermia. Andrology 2014, 3, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Crisóstomo, L.D.; Rato, L.P.; Jarak, I.; Silva, B.M.; Raposo, J.F.; Batterham, R.L.; Oliveira, P.F.; Alves, M.G. A switch from high-fat to normal diet does not restore sperm quality but prevents metabolic syndrome. Reproduction 2019, 158, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Entrez Gene1. SPATS2L Spermatogenesis Associated Serine Rich 2 Like. 2019. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/gene?Db=gene&Cmd=DetailsSearch&Term=26010 (accessed on 18 July 2020).
- Bourbia, N.; Chandler, P.; Codner, G.F.; Banks, G.T.; Nolan, P.M. The guanine nucleotide exchange factor, Spata13, influences social behaviour and nocturnal activity. Mamm. Genome 2019, 30, 54–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macdonald, A.; Stewart, A.; Farquhar, C. Body mass index in relation to semen quality and reproductive hormones in New Zealand men: A cross-sectional study in fertility clinics. Hum. Reprod. 2013, 28, 3178–3187. [Google Scholar] [CrossRef] [Green Version]
- Davidson, L.M.; Millar, K.; Jones, C.; Fatum, M.; Coward, K. Deleterious effects of obesity upon the hormonal and molecular mechanisms controlling spermatogenesis and male fertility. Hum. Fertil. 2015, 18, 184–193. [Google Scholar] [CrossRef]
- Yan, H.H.N.; Mruk, D.D.; Lee, W.M.; Cheng, C.Y. Blood-testis barrier dynamics are regulated by testosterone and cytokines via their differential effects on the kinetics of protein endocytosis and recycling in Sertoli cells. FASEB J. 2008, 22, 1945–1959. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.Y.; Mruk, L.D. The blood-testis barrier and its implications for male contraception. Pharmacol. Rev. 2011, 64, 16–64. [Google Scholar] [CrossRef] [Green Version]
- Lie, P.P.Y.; Mruk, L.D.; Mok, K.W.; Su, L.; Lee, W.M.; Cheng, C.Y. Focal adhesion kinase-Tyr407 and -Tyr397 exhibit antagonistic effects on blood-testis barrier dynamics in the rat. Proc. Natl. Acad. Sci. USA 2012, 109, 12562–12567. [Google Scholar] [CrossRef] [Green Version]
- Lui, W.-Y.; Mruk, D.D.; Lee, W.M.; Cheng, C.Y. Adherens junction dynamics in the testis and spermatogenesis. J. Androl. 2003, 24, 1–14. [Google Scholar]
- Goossens, S.; Van Roy, F. Cadherin-mediated cell-cell adhesion in the testis. Front. Biosci. 2005, 10, 398–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinrich, G.; Muturi, H.T.; Rezaei, K.; Al-Share, Q.Y.; DeAngelis, A.M.; Bowman, T.A.; Ghadieh, H.E.; Ghanem, S.S.; Zhang, D.; Garofalo, R.S.; et al. Reduced Hepatic Carcinoembryonic Antigen-Related Cell Adhesion Molecule 1 Level in Obesity. Front. Endocrinol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Gebauer, F.; Wicklein, D.; Horst, J.; Sundermann, P.; Maar, H.; Streichert, T.; Tachezy, M.; Izbicki, J.R.; Bockhorn, M.; Schumacher, U. Carcinoembryonic Antigen-Related Cell Adhesion Molecules (CEACAM) 1, 5 and 6 as Biomarkers in Pancreatic Cancer. PLoS ONE 2014, 9, e113023. [Google Scholar] [CrossRef] [Green Version]
- Serizawa, S.; Ishii, T.; Nakatani, H.; Tsuboi, A.; Nagawa, F.; Asano, M.; Sudo, K.; Sakagami, J.; Sakano, H.; Ijiri, T.; et al. Mutually exclusive expression of odorant receptor transgenes. Nat. Neurosci. 2000, 3, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Durzyński, Ł.; Gaudin, J.; Myga, M.; Szydłowski, J.; Goździcka-Józefiak, A.; Haertle, T. Olfactory-like receptor cDNAs are present in human lingual cDNA libraries. Biochem. Biophys. Res. Commun. 2005, 333, 264–272. [Google Scholar] [CrossRef]
- Xu, L.L.; Stackhouse, B.G.; Florence, K.; Zhang, W.; Shanmugam, N.; Sesterhenn, I.A.; Zou, Z.; Srikantan, V.; Augustus, M.; Roschke, V.; et al. PSGR, a novel prostate-specific gene with homology to a G protein-coupled receptor, is overexpressed in prostate cancer. Cancer Res. 2000, 60, 6568–6572. [Google Scholar] [PubMed]
- Flegel, C.; Vogel, F.; Hofreuter, A.; Schreiner, B.S.P.; Osthold, S.; Veitinger, S.; Becker, C.; Brockmeyer, N.H.; Muschol, M.; Wennemuth, G.; et al. Characterization of the Olfactory Receptors Expressed in Human Spermatozoa. Front. Mol. Biosci. 2016, 2. [Google Scholar] [CrossRef] [Green Version]
- Suleiman, J.B.; Abu Bakar, A.B.; Mohamed, M. Review on effects of obesity on male reproductive system and the role of natural products. J. Appl. Pharm. Sci. 2019, 9, 131–141. [Google Scholar]
- Spehr, M.; Gisselmann, G.; Poplawski, A.; Riffell, J.A.; Wetzel, C.H.; Zimmer, R.K.; Hatt, H. Identification of a Testicular Odorant Receptor Mediating Human Sperm Chemotaxis. Science 2003, 299, 2054–2058. [Google Scholar] [CrossRef]
- Goto, T.; Salpekar, A.; Monk, M. Expression of a testis-specific member of the olfactory receptor gene family in human primordial germ cells. Mol. Hum. Reprod. 2001, 7, 553–558. [Google Scholar] [CrossRef]
- Fukuda, N.; Yomogida, K.; Okabe, M.; Touhara, K. Functional characterization of a mouse testicular olfactory receptor and its role in chemosensing and in regulation of sperm motility. J. Cell Sci. 2004, 117, 5835–5845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiebaud, N.; Johnson, M.C.; Butler, J.L.; Bell, G.A.; Ferguson, K.L.; Fadool, A.R.; Fadool, J.C.; Gale, A.M.; Gale, D.S.; Fadool, D.A. Hyperlipidemic Diet Causes Loss of Olfactory Sensory Neurons, Reduces Olfactory Discrimination, and Disrupts Odor-Reversal Learning. J. Neurosci. 2014, 34, 6970–6984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, J.; Kim, D.-I.; Han, G.-Y.; Kwon, H.W. The Effects of High Fat Diet-Induced Stress on Olfactory Sensitivity, Behaviors, and Transcriptional Profiling in Drosophila melanogaster. Int. J. Mol. Sci. 2018, 19, 2855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanger, U.M.; Schwab, M. Cytochrome P450 enzymes in drug metabolism: Regulation of gene expression, enzyme activities, and impact of genetic variation. Pharmacol. Ther. 2013, 138, 103–141. [Google Scholar] [CrossRef] [PubMed]
- Lynch, T.; Price, A. The effect of cytochrome P450 metabolism on drug response, interactions, and adverse effects. Am. Fam. Physician 2007, 76, 391–396. [Google Scholar]
- Carreau, S. Estrogens--male hormones? Folia Histochemica Cytobiologica 2003, 41, 107–111. [Google Scholar]
- Ramasamy, R.; Schulster, M.; Bernie, A.M. The role of estradiol in male reproductive function. Asian J. Androl. 2016, 18, 435–440. [Google Scholar] [CrossRef]
- UniProtKB—P11711 CP2A1_RAT. Available online: https://www.uniprot.org/uniprot/P11711 (accessed on 20 July 2020).
- UniProtKB—P33274 (CP4F1_RAT). Available online: https://www.uniprot.org/uniprot/P33274 (accessed on 20 July 2020).
- UniProtKB—P12938 (CP2D3_RAT). Available online: https://www.uniprot.org/uniprot/P12938 (accessed on 20 July 2020).
- Arnoldi, A.; Crimella, C.; Tenderini, E.; Martinuzzi, A.; D’Angelo, M.G.; Musumeci, O.; Toscano, A.; Scarlato, M.; Fantin, M.; Bresolin, N.; et al. Clinical phenotype variability in patients with hereditary spastic paraplegia type 5 associated with CYP7B1 mutations. Clin. Genet. 2011, 81, 150–157. [Google Scholar] [CrossRef]
- Turner, M.D.; Nedjai, B.; Hurst, T.; Pennington, D.J. Cytokines and chemokines: At the crossroads of cell signalling and inflammatory disease. Biochim. Biophys. Acta Bioenerg. 2014, 1843, 2563–2582. [Google Scholar] [CrossRef] [Green Version]
- Ota, T. Chemokine Systems Link Obesity to Insulin Resistance. Diabetes Metab. J. 2013, 37, 165–172. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Kitade, H.; Ni, Y.; Ota, T. Roles of Chemokines and Chemokine Receptors in Obesity-Associated Insulin Resistance and Nonalcoholic Fatty Liver Disease. Biomolecules 2015, 5, 1563–1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, F.M.; Weschenfelder, J.; Sander, C.; Minkwitz, J.; Thormann, J.; Chittka, T.; Mergl, R.; Kirkby, K.C.; Faßhauer, M.; Stumvoll, M.; et al. Inflammatory Cytokines in General and Central Obesity and Modulating Effects of Physical Activity. PLoS ONE 2015, 10, e0121971. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.-C.; Lee, J. Cellular and molecular players in adipose tissue inflammation in the development of obesity-induced insulin resistance. Biochim. Biophys. Acta Bioenerg. 2013, 1842, 446–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujisaka, S.; Usui, I.; Bukhari, A.; Ikutani, M.; Oya, T.; Kanatani, Y.; Tsuneyama, K.; Nagai, Y.; Takatsu, K.; Urakaze, M.; et al. Regulatorymechanisms for adipose tissue M1andM2macrophages in diet-induced obesemice. Diabetes 2009, 58, 2574–2582. [Google Scholar] [CrossRef] [Green Version]
- Blaschitz, C.; Raffatellu, M. Th17 Cytokines and the Gut Mucosal Barrier. J. Clin. Immunol. 2010, 30, 196–203. [Google Scholar] [CrossRef] [Green Version]
- Qu, N.; Xu, M.; Mizoguchi, I.; Furusawa, J.-I.; Kaneko, K.; Watanabe, K.; Mizuguchi, J.; Itoh, M.; Kawakami, Y.; Yoshimoto, T. Pivotal Roles of T-Helper 17-Related Cytokines, IL-17, IL-22, and IL-23, in Inflammatory Diseases. Clin. Dev. Immunol. 2013, 2013, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudakov, J.A.; Hanash, A.M.; Brink, M.R.V.D. Interleukin-22: Immunobiology and pathology. Annu. Rev. Immunol. 2015, 33, 747–785. [Google Scholar] [CrossRef] [Green Version]
- Liang, S.C.; Nickerson-Nutter, C.; Pittman, D.D.; Carrier, Y.; Goodwin, D.G.; Shields, K.M.; Lambert, A.J.; Schelling, S.H.; Medley, Q.G.; Ma, H.L.; et al. IL-22 induces an acute-phase response. J. Immunol. 2010, 185, 5531–5538. [Google Scholar] [CrossRef] [Green Version]
- Morris, D.L.; Oatmen, K.E.; Mergian, T.A.; Cho, K.W.; DelProposto, J.L.; Singer, K.; Evans-Molina, C.; O’Rourke, R.W.; Lumeng, C.N. CD40 promotes MHC class II expression on adipose tissue macrophages and regulates adipose tissue CD4+ T cells with obesity. J. Leukoc. Biol. 2015, 99, 1107–1119. [Google Scholar] [CrossRef] [Green Version]
- Zou, J.; Lai, B.; Zheng, M.; Chen, Q.; Jiang, S.; Song, A.; Huang, Z.; Shi, P.; Tu, X.; Wang, D.; et al. CD4+ T cells memorize obesity and promote weight regain. Cell. Mol. Immunol. 2017, 15, 630–639. [Google Scholar] [CrossRef] [Green Version]
- Woods, S.C.; Seeley, R.J.; Rushing, P.A.; D’Alessio, D.; Tso, P. A Controlled High-Fat Diet Induces an Obese Syndrome in Rats. J. Nutr. 2003, 133, 1081–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. g:Profiler: A web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2009, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Transcript ID | Gene Symbol | Description | Function | O/C FC |
|---|---|---|---|---|---|
| Obesity and lipid metabolism | |||||
| 681337 | NM_001109440 | Acot4 | acyl-CoA thioesterase 4 | OM, fatty acid elongation, biosynthesis of unsaturated fatty acids | −10.75 |
| 24654 | NM_001077641 | Plcb1 | phospholipase C beta 1 | Protection against HFD-induced obesity, Aldosterone synthesis and secretion, regulation of acrosome reaction, positive regulation of fertilization, interleukin-12 production | −8.55 |
| 24254 | NM_016997 | Cel | carboxyl ester lipase | OM, Steroid biosynthesis, Fat digestion and absorption | −3.9 |
| 296138 | NM_001106508 | Acoxl | acyl-CoA oxidase-like | OM, fatty acid beta-oxidation | −4.72 |
| 690388 | NM_001109587 | Pla2g2f | phospholipase A2, group IIF | protection against HFD-induced obesity | 3.0 |
| 24207 | NM_001271053, NM_012501 | Apoc3 | apolipoprotein C3 | OM, reduce obesity, negative regulation of lipid metabolism and transport, negative regulation of lipase activity | 7.66 |
| 362540 | NM_001108669 | Hacd4 | 3-hydroxyacyl-CoA dehydratase 4 | Fatty acid elongation, fatty acid metabolism | 4.81 |
| 89813 | NM_053551 | Pdk4 | pyruvate dehydrogenase kinase 4 | OM, response to stress, cellular response to fatty acid, regulation of fatty acid oxidation, cellular response to stress | 4.69 |
| 300438 | NM_175762 | Ldlr | Low-density lipoprotein receptor | Ovarian steroidogenesis, Aldosterone synthesis and secretion, Cortisol synthesis and secretion, cholesterol transport, intestinal cholesterol absorption, intestinal lipid | 3.81 |
| Spermatogenesis, reproduction, fertilization | |||||
| 305938 | NM_001191686 | Spata13 | spermatogenesis associated 13 | Regulation of Rho and Ras signal transduction pathways, regulation of actin cytoskeleton | 7.42 |
| 316426 | NM_001014102 | Spats2l | spermatogenesis associated, serine-rich 2-like | spermatogenesis | 2.91 |
| 100362580 | NM_001304424 | Spag11bl | sperm associated antigen 11b-like | Sperm maturation | −3.18 |
| 360896 | NM_203336 | Esrrg | estrogen-related receptor gamma | spermatogenesis, regulation of transcription | −9.63 |
| 298961 | NM_001106725 | Agr2 | anterior gradient 2, protein disulphide isomerase family member | Sperm maturation and fertilization, cell adhesion, response to stress | −7.8 |
| Cytochrome P450 (CYPs) | |||||
| 24303 | NM_173093 | Cyp2d3 | Cytochrome P450, family 2, subfamily d, polypeptide 3 | OM, Steroid hormone biosynthesis, Serotonergic synapse | −3.07 |
| 24894 | NM_012692 | Cyp2a1 | Cytochrome P450, family 2, subfamily a, polypeptide 1 | OM, Retinol metabolism, steroid hydroxylase activity, aromatase activity | 2.51 |
| 56266 | NM_019623 | Cyp4f1 | Cytochrome P450, family 4, subfamily f, polypeptide 1 | OM, arachidonic acid metabolism, monooxygenase activity, | 2.75 |
| 25429 | NM_019138 | Cyp7b1 | Cytochrome P450, family 7, subfamily b, polypeptide 1 | OM, Primary bile acid biosynthesis, localization of cell, monooxygenase activity, | 3.49 |
| 25147 | NM_017085 | Cyp19a1 | Cytochrome P450, family 19, subfamily a, polypeptide 1 | aromatase | 2.1 |
| Cell adhesion | |||||
| 364844 | NM_001012215 | Pcdhgb7 | protocadherin gamma subfamily B, 7 | OM | −2.96 |
| 81613 | NM_001033860, NM_001033861, NM_001033862, NM_031755 | Ceacam1 | carcinoembryonic antigen-related cell adhesion molecule 1 | protection from obesity | 3.07 |
| 116711 | NM_053919 | Ceacam9 | carcinoembryonic antigen-related cell adhesion molecule 9 | protection from obesity | −2.74 |
| Immune response | |||||
| 288593 | NM_001013045 | Ccl24 | C-C motif chemokine ligand 24 | OM | 3.57 |
| 362629 | NM_001191869 | Il22ra1 | interleukin 22 receptor subunit alpha 1 | OM, Cytokine-cytokine receptor interaction, Jak-STAT signaling pathway | 5.53 |
| 24932 | NM_012705 | Cd4 | Cd4 molecule | OM, Cytokine-cytokine receptor interaction | 6.37 |
| 362417 | NM_001004091 | Il17re | interleukin 17 receptor E | OM | 3.30 |
| 287910 | NM_001004202 | Ccl6 | chemokine (C-C motif) ligand 6 | OM | 2.75 |
| 246759 | NM_145672 | Cxcl9 | C-X-C motif chemokine ligand 9 | OM | 2.03 |
| Olfaction | |||||
| 405186 | NM_001000886 | Olr1710 | olfactory receptor 1710 | Olfactory transduction, perception of smell | −8.68 |
| 296694 | NM_001000397 | Olr434 | olfactory receptor 434 | OM, Olfactory transduction, perception of smell | −7.07 |
| 295743 | NM_001000301 | Olr472 | olfactory receptor 472 | OM, Olfactory transduction, perception of smell | −4.23 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Shehawi, A.M.; El-Shazly, S.; Ahmed, M.; Alkafafy, M.; Sayed, S.; Farouk, S.; Alotaibi, S.S.; Elseehy, M.M. Transcriptome Analysis of Testis from HFD-Induced Obese Rats (Rattus norvigicus) Indicated Predisposition for Male Infertility. Int. J. Mol. Sci. 2020, 21, 6493. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186493
El-Shehawi AM, El-Shazly S, Ahmed M, Alkafafy M, Sayed S, Farouk S, Alotaibi SS, Elseehy MM. Transcriptome Analysis of Testis from HFD-Induced Obese Rats (Rattus norvigicus) Indicated Predisposition for Male Infertility. International Journal of Molecular Sciences. 2020; 21(18):6493. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186493
Chicago/Turabian StyleEl-Shehawi, Ahmed M., Samir El-Shazly, Mohamed Ahmed, Mohamed Alkafafy, Samy Sayed, Samy Farouk, Saqer S. Alotaibi, and Mona M. Elseehy. 2020. "Transcriptome Analysis of Testis from HFD-Induced Obese Rats (Rattus norvigicus) Indicated Predisposition for Male Infertility" International Journal of Molecular Sciences 21, no. 18: 6493. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186493