The Impact of Genetic Polymorphisms in Organic Cation Transporters on Renal Drug Disposition

 , , and
, , and
Abstract
:1. Introduction
2. Role of the Organic Cation Transporters
2.1. Role of OCT1
2.2. Role of OCT2
2.3. Role of OCT3
3. Drug-Related Genetic Polymorphisms in the Organic Cation Transporter Genes
3.1. Genetic Polymorphisms in the OCT1 Gene (SLC22A1)
3.2. Genetic Polymorphism in the OCT2 Gene (SLC22A2)
3.3. Genetic Polymorphisms in the OCT3 Gene (SLC22A3)
4. Impact of Pharmacogenetic Variants in OCTs in Precision Medicine
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| SLC | Solute carrier |
| OAT | Organic anion transporters |
| OCT | Organic cation transporter |
| SNP | Single nucleotide polymorphism |
| MATE | Multidrug and toxic compound extrusion |
| CTR | Copper transporter |
| ABC | ATP-binding cassette |
| MRP | Multidrug resistance protein |
| OC | Organic cation |
| CNS | Central nervous system |
| MAF | Minor allele frequency |
| HEK293 | Human embryonic kidney 293 cells |
| T2DM | Type 2 diabetes mellitus |
| eGFR | Estimated glomerular filtration rate |
| MPP+ | 1-methyl-4-phenylpyridinium |
| TBA | Tetrabutylammonium |
| HOMA-IR | Homeostatic model assessment for insulin resistance |
| 3′UTR | Three prime untranslated region |
| MTT | 3-[4,5-dimethylthiazol-2-yl]-2,5 diphenyl tetrazolium bromide |
| KIM-1 | Kidney injury molecule-1 |
| B2M | β2-microglobulin |
| TFF-3 | Trefoil factor-3 |
| NSCLC | Non-small cell lung cancer |
| CKD | Chronic kidney disease |
| GWAS | Genome-wide association studies |
| CRISPR-Cas9 | Clustered Regularly Interspaced Short Palindromic Repeats and Cas genes |
References
- Ivanyuk, A.; Livio, F.; Biollaz, J.; Buclin, T. Renal Drug Transporters and Drug Interactions. Clin. Pharmacokinet. 2017, 56, 825–892. [Google Scholar] [CrossRef] [PubMed]
- Ciarimboli, G. Organic Cation Transporters. Xenobiotica 2008, 38, 936–971. [Google Scholar] [CrossRef] [PubMed]
- Koepsell, H.; Endou, H. The SLC22 drug transporter family. Pflug. Arch. Eur. J. Physiol. 2004, 447, 666–676. [Google Scholar] [CrossRef]
- Koepsell, H.; Lips, K.; Volk, C. Polyspecific organic cation transporters: Structure, function, physiological roles, and biopharmaceutical implications. Pharm. Res. 2007, 24, 1227–1251. [Google Scholar] [CrossRef] [PubMed]
- Koepsell, H.; Schmitt, B.M.; Gorboulev, V. Organic cation transporters. Rev. Physiol. Biochem. Pharmacol. 2003, 150, 36–90. [Google Scholar] [CrossRef] [PubMed]
- Van Montfoort, J.; Hagenbuch, B.; Groothuis, G.; Koepsell, H.; Meier, P.; Meijer, D. Drug Uptake Systems in Liver and Kidney. Curr. Drug Metab. 2005, 4, 185–211. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Gorset, W.; Dresser, M.J.; Giacomini, K.M. The Interaction of n-Tetraalkylammonium Compounds with a Human Organic Cation Transporter, hOCT1. J. Pharmacol. Exp. Ther. 1999, 288, 1192–1198. [Google Scholar]
- Wagner, D.J.; Hu, T.; Wang, J. Polyspecific organic cation transporters and their impact on drug intracellular levels and pharmacodynamics. Pharmacol. Res. 2016, 111, 237–246. [Google Scholar] [CrossRef] [Green Version]
- Koepsell, H. Polyspecific organic cation transporters: Their functions and interactions with drugs. Trends Pharmacol. Sci. 2004, 25, 375–381. [Google Scholar] [CrossRef]
- Moraes, F.; Góes, A. A decade of human genome project conclusion: Scientific diffusion about our genome knowledge. Biochem. Mol. Biol. Educ. 2016, 44, 215–223. [Google Scholar] [CrossRef] [Green Version]
- Roden, D.M.; McLeod, H.L.; Relling, M.V.; Williams, M.S.; Mensah, G.A.; Peterson, J.F.; Van Driest, S.L. Pharmacogenomics. Lancet 2019, 394, 521–532. [Google Scholar] [CrossRef]
- Yee, S.W.; Brackman, D.J.; Ennis, E.A.; Sugiyama, Y.; Kamdem, L.K.; Blanchard, R.; Galetin, A.; Zhang, L.; Giacomini, K.M. Influence of Transporter Polymorphisms on Drug Disposition and Response: A Perspective From the International Transporter Consortium. Clin. Pharmacol. Ther. 2018, 104, 803–817. [Google Scholar] [CrossRef] [PubMed]
- Pelis, R.M.; Zhang, X.; Dangprapai, Y.; Wright, S.H. Cysteine accessibility in the hydrophilic cleft of human organic cation transporter 2. J. Biol. Chem. 2006, 281, 35272–35280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorboulev, V.; Ulzheimer, J.C.; Akhoundova, A.; Ulzheimer-Teuber, I.; Karbach, U.; Quester, S.; Baumann, C.; Lang, F.; Koepsell, H. Cloning and characterization of two human polyspecific organic cation transporters. DNA Cell Biol. 1997, 16, 871–881. [Google Scholar] [CrossRef] [PubMed]
- Nies, A.T.; Koepsell, H.; Winter, S.; Burk, O.; Klein, K.; Kerb, R.; Zanger, U.M.; Keppler, D.; Schwab, M.; Schaeffeler, E. Expression of organic cation transporters OCT1 (SLC22A1) and OCT3 (SLC22A3) is affected by genetic factors and cholestasis in human liver. Hepatology 2009, 50, 1227–1240. [Google Scholar] [CrossRef]
- Nishimura, M.; Naito, S. Tissue-specific mRNA Expression Profiles of Human ATP-binding Cassette and Solute Carrier Transporter Superfamilies. Drug Metab. Pharmacokinet. 2005, 20, 452–477. [Google Scholar] [CrossRef]
- Wessler, I.; Roth, E.; Deutsch, C.; Brockerhoff, P.; Bittinger, F.; Kirkpatrick, C.J.; Kilbinger, H. Release of non-neuronal acetylcholine from the isolated human placenta is mediated by organic cation transporters. Br. J. Pharmacol. 2001, 134, 951–956. [Google Scholar] [CrossRef] [Green Version]
- Lips, K.S.; Volk, C.; Schmitt, B.M.; Pfeil, U.; Arndt, P.; Miska, D.; Ermert, L.; Kummer, W.; Koepsell, H. Polyspecific cation transporters mediate luminal release of acetylcholine from bronchial epithelium. Am. J. Respir. Cell Mol. Biol. 2005, 33, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Nies, A.T.; Koepsell, H.; Damme, K.; Schwab, M. Organic Cation Transporters (OCTs, MATEs), In Vitro and In Vivo Evidence for the Importance in Drug Therapy. In Handbook of Experimental Pharmacology; Springer: Berlin, Germany, 2011; Volume 201, pp. 105–167. ISBN 9783642145407. [Google Scholar]
- Tzvetkov, M.V.; Vormfelde, S.J.H.V.; Balen, D.; Meineke, I.; Schmidt, T.; Sehrt, D.; Sabolić, I.; Koepsell, H.; Brockmöller, J. The effects of genetic polymorphisms in the organic cation transporters OCT1, OCT2, and OCT3 on the renal clearance of metformin. Clin. Pharmacol. Ther. 2009, 86, 299–306. [Google Scholar] [CrossRef]
- Koepsell, H. The SLC22 family with transporters of organic cations, anions and zwitterions. Mol. Asp. Med. 2013, 34, 413–435. [Google Scholar] [CrossRef]
- Motohashi, H.; Inui, K.I. Organic cation transporter OCTs (SLC22) and MATEs (SLC47) in the human kidney. AAPS J. 2013, 15, 581–588. [Google Scholar] [CrossRef] [Green Version]
- Sugawara-Yokoo, M.; Urakami, Y.; Koyama, H.; Fujikura, K.; Masuda, S.; Saito, H.; Naruse, T.; Inui, K.I.; Takata, K. Differential localization of organic cation transporters rOCT1 and rOCT2 in the basolateral membrane of rat kidney proximal tubules. Histochem. Cell Biol. 2000, 114, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Prasad, B. Kidney cortical transporter expression across species using quantitative proteomics. Drug Metab. Dispos. 2019, 47, 802–808. [Google Scholar] [CrossRef]
- Andreev, E.; Brosseau, N.; Carmona, E.; Mes-Masson, A.M.; Ramotar, D. The human organic cation transporter OCT1 mediates high affinity uptake of the anticancer drug daunorubicin. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Koepsell, H. Role of organic cation transporters in drug-drug interaction. Expert Opin. Drug Metab. Toxicol. 2015, 11, 1619–1633. [Google Scholar] [CrossRef]
- Hendrickx, R.; Johansson, J.G.; Lohmann, C.; Jenvert, R.M.; Blomgren, A.; Börjesson, L.; Gustavsson, L. Identification of novel substrates and structure-activity relationship of cellular uptake mediated by human organic cation transporters 1 and 2. J. Med. Chem. 2013, 56, 7232–7242. [Google Scholar] [CrossRef]
- Tojo, A.; Sekine, T.; Nakajima, N.; Hosoyamada, M.; Kanai, Y.; Kimura, K.; Endou, H. Immunohistochemical localization of multispecific renal organic anion transporter 1 in rat kidney. J. Am. Soc. Nephrol. 1999, 10, 464–471. [Google Scholar]
- Ciarimboli, G.; Lancaster, C.S.; Schlatter, E.; Franke, R.M.; Sprowl, J.A.; Pavenstädt, H.; Massmann, V.; Guckel, D.; Mathijssen, R.H.J.; Yang, W.; et al. Proximal tubular secretion of creatinine by organic cation transporter OCT2 in cancer patients. Clin. Cancer Res. 2012, 18, 1101–1108. [Google Scholar] [CrossRef] [Green Version]
- Busch, A.E.; Karbach, U.; Miska, D.; Gorboulev, V.; Akhoundova, A.; Volk, C.; Arndt, P.; Ulzheimer, J.C.; Sonders, M.S.; Baumann, C.; et al. Human neurons express the polyspecific cation transporter hOCT2, which translocates monoamine neurotransmitters, amantadine, and memantine. Mol. Pharmacol. 1998, 54, 342–352. [Google Scholar] [CrossRef] [Green Version]
- Dresser, M.J.; Xiao, G.; Leabman, M.K.; Gray, A.T.; Giacomini, K.M. Interactions of n-tetraalkylammonium compounds and biguanides with a human renal organic cation transporter (hOCT2). Pharm. Res. 2002, 19, 1244–1247. [Google Scholar] [CrossRef]
- Kimura, N.; Masuda, S.; Tanihara, Y.; Ueo, H.; Okuda, M.; Katsura, T.; Inui, K.I. Metformin is a superior substrate for renal organic cation transporter OCT2 rather than hepatic OCT1. Drug Metab. Pharmacokinet. 2005, 20, 379–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urakami, Y.; Akazawa, M.; Saito, H.; Okuda, M.; Inui, K.I. cDNA cloning, functional characterization, and tissue distribution of an alternatively spliced variant of organic cation transporter hOCT2 predominantly expressed in the human kidney. J. Am. Soc. Nephrol. 2002, 13, 1703–1710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sata, R.; Ohtani, H.; Tsujimoto, M.; Murakami, H.; Koyabu, N.; Nakamura, T.; Uchiumi, T.; Kuwano, M.; Nagata, H.; Tsukimori, K.; et al. Functional analysis of organic cation transporter 3 expressed in human placenta. J. Pharmacol. Exp. Ther. 2005, 315, 888–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kekuda, R.; Prasad, P.D.; Wu, X.; Wang, H.; Fei, Y.-J.; Leibach, F.H.; Ganapathy, V. Cloning and Functional Characterization of a Potential-sensitive, Polyspecific Organic Cation Transporter (OCT3) Most Abundantly Expressed in Placenta. J. Biol. Chem. 1998, 273, 15971–15979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasser, P.J.; Lowry, C.A. Organic cation transporter 3: A cellular mechanism underlying rapid, non-genomic glucocorticoid regulation of monoaminergic neurotransmission, physiology, and behavior. Horm. Behav. 2018, 104, 173–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Wei, H.; Ganapathy, M.E.; Wang, H.; Kekuda, R.; Conway, S.J.; Leibach, F.H.; Ganapathy, V. Structure, function, and regional distribution of the organic cation transporter OCT3 in the kidney. Am. J. Physiol.-Ren. Physiol. 2000, 279. [Google Scholar] [CrossRef]
- Dresser, M.J.; Leabman, M.K.; Giacomini, K.M. Transporters involved in the elimination of drugs in the kidney: Organic anion transporters and organic cation transporters. J. Pharm. Sci. 2001, 90. [Google Scholar] [CrossRef]
- Christensen, M.M.H.; Brasch-Andersen, C.; Green, H.; Nielsen, F.; Damkier, P.; Beck-Nielsen, H.; Brosen, K. The pharmacogenetics of metformin and its impact on plasma metformin steady-state levels and glycosylated hemoglobin A1c. Pharmacogenet. Genom. 2011, 21, 837–850. [Google Scholar] [CrossRef]
- Tzvetkov, M.V.; Dos Santos Pereira, J.N.; Meineke, I.; Saadatmand, A.R.; Stingl, J.C.; Brockmöller, J. Morphine is a substrate of the organic cation transporter OCT1 and polymorphisms in OCT1 gene affect morphine pharmacokinetics after codeine administration. Biochem. Pharmacol. 2013, 86, 666–678. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, A.J.T.; Stage, T.B.; Glintborg, D.; Andersen, M.; Christensen, M.M.H. The Pharmacogenetics of Metformin in Women with Polycystic Ovary Syndrome: A Randomized Trial. Basic Clin. Pharmacol. Toxicol. 2018, 122, 239–244. [Google Scholar] [CrossRef]
- Santoro, A.B.; Botton, M.R.; Struchiner, C.J.; Suarez-Kurtz, G. Influence of pharmacogenetic polymorphisms and demographic variables on metformin pharmacokinetics in an admixed Brazilian cohort. Br. J. Clin. Pharmacol. 2018, 84, 987–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarasova, L.; Kalnina, I.; Geldnere, K.; Bumbure, A.; Ritenberga, R.; Nikitina-Zake, L.; Fridmanis, D.; Vaivade, I.; Pirags, V.; Klovins, J. Association of genetic variation in the organic cation transporters OCT1, OCT2 and multidrug and toxin extrusion 1 transporter protein genes with the gastrointestinal side effects and lower BMI in metformin-treated type 2 diabetes patients. Pharmacogenet. Genom. 2012, 22, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Cheong, H.S.; Kim, H.D.; Na, H.S.; Kim, J.O.; Kim, L.H.; Kim, S.H.; Bae, J.S.; Chung, M.W.; Shin, H.D. Screening of genetic variations of SLC15A2, SLC22A1, SLC22A2 and SLC22A6 genes. J. Hum. Genet. 2011, 56, 666–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkatasubramanian, R.; Fukuda, T.; Niu, J.; Mizuno, T.; Chidambaran, V.; Vinks, A.A.; Sadhasivam, S. ABCC3 and OCT1 genotypes influence pharmacokinetics of morphine in children. Pharmacogenomics 2014, 15, 1297–1309. [Google Scholar] [CrossRef] [Green Version]
- Balyan, R.; Zhang, X.; Chidambaran, V.; Martin, L.J.; Mizuno, T.; Fukuda, T.; Vinks, A.A.; Sadhasivam, S. OCT1 genetic variants are associated with postoperative morphine-related adverse effects in children. Pharmacogenomics 2017, 18, 621–629. [Google Scholar] [CrossRef] [Green Version]
- Kroetz, D.L.; Yee, S.W.; Giacomini, K.M. The Pharmacogenomics of Membrane Transporters Project: Research at the Interface of Genomics and Transporter Pharmacology. Clin. Pharmacol. Ther. 2010, 87, 109–116. [Google Scholar] [CrossRef]
- Chen, L.; Takizawa, M.; Chen, E.; Schlessinger, A.; Segenthelar, J.; Choi, J.H.; Sali, A.; Kubo, M.; Nakamura, S.; Iwamoto, Y.; et al. Genetic Polymorphisms in Organic Cation Transporter 1 (OCT1) in Chinese and Japanese Populations Exhibit Altered Function. J. Pharmacol. Exp. Ther. 2010, 335, 42–50. [Google Scholar] [CrossRef]
- Singh, O.; Chan, J.Y.; Lin, K.; Heng, C.C.T.; Chowbay, B. SLC22A1-ABCB1 Haplotype Profiles Predict Imatinib Pharmacokinetics in Asian Patients with Chronic Myeloid Leukemia. PLoS ONE 2012, 7, e51771. [Google Scholar] [CrossRef] [Green Version]
- Altall, R.M.; Qusti, S.Y.; Filimban, N.; Alhozali, A.M.; Alotaibi, N.A.; Dallol, A.; Chaudhary, A.G.; Bakhashab, S. SLC22A1 And ATM Genes Polymorphisms Are Associated With The Risk Of Type 2 Diabetes Mellitus In Western Saudi Arabia: A Case-Control Study. Appl. Clin. Genet. 2019, 12, 213–219. [Google Scholar] [CrossRef] [Green Version]
- Jablonski, K.A.; McAteer, J.B.; de Bakker, P.I.W.; Franks, P.W.; Pollin, T.I.; Hanson, R.L.; Saxena, R.; Fowler, S.; Shuldiner, A.R.; Knowler, W.C.; et al. Common Variants in 40 Genes Assessed for Diabetes Incidence and Response to Metformin and Lifestyle Intervention in the Diabetes Prevention Program. Diabetes 2010, 59, 2672–2681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, H.-B.; Zhuang, W.; Wu, T.; Xin, S.; Lin, C.-Z.; Ruan, H.-L.; Zhu, X.; Huang, M.; Li, J.-L.; Hou, X.-Y.; et al. Imatinib-induced ophthalmological side-effects in GIST patients are associated with the variations of EGFR, SLC22A1, SLC22A5 and ABCB1. Pharm. J. 2018, 18, 460–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arimany-Nardi, C.; Koepsell, H.; Pastor-Anglada, M. Role of SLC22A1 polymorphic variants in drug disposition, therapeutic responses, and drug-drug interactions. Pharmacogenom. J. 2015, 15, 473–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, C.; Pearce, B.; Du Plessis, M.; Hoosain, N.; Benjeddou, M. Genetic polymorphisms and haplotypes of the organic cation transporter 1 gene (SLC22A1) in the Xhosa population of South Africa. Genet. Mol. Biol. 2014, 37, 350–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naja, K.; El Shamieh, S.; Fakhoury, R. rs622342A>C in SLC22A1 is associated with metformin pharmacokinetics and glycemic response. Drug Metab. Pharmacokinet. 2020, 35, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Sajib, A.A.; Islam, T.; Paul, N.; Yeasmin, S. Interaction of rs316019 variants of SLC22A2 with metformin and other drugs- an in silico analysis. J. Genet. Eng. Biotechnol. 2018, 16, 769–775. [Google Scholar] [CrossRef]
- Wang, Z.J.; Yin, O.Q.P.; Tomlinson, B.; Chow, M.S.S. OCT2 polymorphisms and in-vivo renal functional consequence: Studies with metformin and cimetidine. Pharmacogenet. Genom. 2008, 18, 637–645. [Google Scholar] [CrossRef]
- Song, I.; Shin, H.; Shim, E.; Jung, I.; Kim, W.; Shon, J.; Shin, J. Genetic Variants of the Organic Cation Transporter 2 Influence the Disposition of Metformin. Clin. Pharmacol. Ther. 2008, 84, 559–562. [Google Scholar] [CrossRef] [PubMed]
- Frenzel, D.; Köppen, C.; Bolle Bauer, O.; Karst, U.; Schröter, R.; Tzvetkov, M.V.; Ciarimboli, G. Effects of Single Nucleotide Polymorphism Ala270Ser (rs316019) on the Function and Regulation of hOCT2. Biomolecules 2019, 9, 578. [Google Scholar] [CrossRef] [Green Version]
- Filipski, K.K.; Mathijssen, R.H.; Mikkelsen, T.S.; Schinkel, A.H.; Sparreboom, A. Contribution of organic cation transporter 2 (OCT2) to cisplatin-induced nephrotoxicity. Clin. Pharmacol. Ther. 2009, 86, 396–402. [Google Scholar] [CrossRef]
- Zazuli, Z.; Otten, L.S.; Drögemöller, B.I.; Medeiros, M.; Monzon, J.G.; Wright, G.E.; Kollmannsberger, C.K.; Bedard, P.L.; Chen, Z.; Gelmon, K.A.; et al. Outcome Definition Influences the Relationship between Genetic Polymorphisms of ERCC1, ERCC2, SLC22A2 and Cisplatin Nephrotoxicity in Adult Testicular Cancer Patients. Genes (Basel) 2019, 10, 364. [Google Scholar] [CrossRef] [Green Version]
- Kashi, Z.; Masoumi, P.; Mahrooz, A.; Hashemi-Soteh, M.B.; Bahar, A.; Alizadeh, A. The variant organic cation transporter 2 (OCT2)–T201M contribute to changes in insulin resistance in patients with type 2 diabetes treated with metformin. Diabetes Res. Clin. Pract. 2015, 108, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.-K.; Song, I.-S. Genetic variants of organic cation transporter 1 (OCT1) and OCT2 significantly reduce lamivudine uptake. Biopharm. Drug Dispos. 2012, 33, 170–178. [Google Scholar] [CrossRef]
- Leabman, M.K.; Huang, C.C.; Kawamoto, M.; Johns, S.J.; Stryke, D.; Ferrin, T.E.; DeYoung, J.; Taylor, T.; Clark, A.G.; Herskowitz, I.; et al. Polymorphisms in a human kidney xenobiotic transporter, OCT2, exhibit altered function. Pharmacogenetics 2002, 12, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Moeez, S.; Khalid, Z.; Jalil, F.; Irfan, M.; Ismail, M.; Arif, M.A.; Niazi, R.; Khalid, S. Effects of SLC22A2 (rs201919874) and SLC47A2 (rs138244461) genetic variants on Metformin Pharmacokinetics in Pakistani T2DM patients. J. Pak. Med. Assoc. 2019, 69, 155–163. [Google Scholar] [PubMed]
- Chang, C.; Hu, Y.; Hogan, S.L.; Mercke, N.; Gomez, M.; O’Bryant, C.; Bowles, D.W.; George, B.; Wen, X.; Aleksunes, L.M.; et al. Pharmacogenomic variants may influence the urinary excretion of novel kidney injury biomarkers in patients receiving cisplatin. Int. J. Mol. Sci. 2017, 18, 1333. [Google Scholar] [CrossRef] [Green Version]
- Sallinen, R.; Kaunisto, M.A.; Forsblom, C.; Thomas, M.; Fagerudd, J.; Pettersson-Fernholm, K.; Groop, P.H.; Wessman, M. Association of the SLC22A1, SLC22A2, and SLC22A3 genes encoding organic cation transporters with diabetic nephropathy and hypertension. Ann. Med. 2010, 42, 296–304. [Google Scholar] [CrossRef]
- Choi, C.-I.; Bae, J.-W.; Keum, S.-K.; Lee, Y.-J.; Lee, H.-I.; Jang, C.-G.; Lee, S.-Y. Effects of OCT2 c.602C > T genetic variant on the pharmacokinetics of lamivudine. Xenobiotica 2013, 43, 636–640. [Google Scholar] [CrossRef]
- Hakooz, N.; Jarrar, Y.B.; Zihlif, M.; Imraish, A.; Hamed, S.; Arafat, T. Effects of the genetic variants of organic cation transporters 1 and 3 on the pharmacokinetics of metformin in Jordanians. Drug Metab. Pers. Ther. 2017, 32, 157–162. [Google Scholar] [CrossRef]
- Mahrooz, A.; Alizadeh, A.; Hashemi-Soteh, M.B.; Ghaffari-Cherati, M.; Hosseyni-Talei, S.R. The Polymorphic Variants rs3088442 and rs2292334 in the Organic Cation Transporter 3 (OCT3) Gene and Susceptibility Against Type 2 Diabetes: Role of their Interaction. Arch. Med. Res. 2017, 48, 162–168. [Google Scholar] [CrossRef]
- Moeez, S.; Riaz, S.; Masood, N.; Kanwal, N.; Arif, M.A.; Niazi, R.; Khalid, S. Evaluation of the rs3088442 G>A SLC22A3 Gene Polymorphism and the Role of microRNA 147 in Groups of Adult Pakistani Populations With Type 2 Diabetes in Response to Metformin. Can. J. Diabetes 2019, 43, 128–135.e3. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Pawlikowski, B.; Schlessinger, A.; More, S.S.; Stryke, D.; Johns, S.J.; Portman, M.A.; Chen, E.; Ferrin, T.E.; Sali, A.; et al. Role of organic cation transporter 3 (SLC22A3) and its missense variants in the pharmacologic action of metformin. Pharmacogenet. Genom. 2010, 20, 687–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakata, T.; Anzai, N.; Kimura, T.; Miura, D.; Fukutomi, T.; Takeda, M.; Sakurai, H.; Endou, H. Functional Analysis of Human Organic Cation Transporter OCT3 (SLC22A3) Polymorphisms. J. Pharmacol. Sci. 2010, 113, 263–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozano, E.; Briz, O.; Macias, R.; Serrano, M.; Marin, J.; Herraez, E. Genetic Heterogeneity of SLC22 Family of Transporters in Drug Disposition. J. Pers. Med. 2018, 8, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Hong, C.; Chen, E.C.; Yee, S.W.; Xu, L.; Almof, E.U.; Wen, C.; Fujii, K.; Johns, S.J.; Stryke, D.; et al. Genetic and epigenetic regulation of the organic cation transporter 3, SLC22A3. Pharmacogenom. J. 2013, 13, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Cacabelos, R.; Torrellas, C. Epigenetics of Aging and Alzheimer’s Disease: Implications for Pharmacogenomics and Drug Response. Int. J. Mol. Sci. 2015, 16, 30483–30543. [Google Scholar] [CrossRef] [Green Version]
- Chen, E.C.; Liang, X.; Yee, S.W.; Geier, E.G.; Stocker, S.L.; Chen, L.; Giacomini, K.M. Targeted disruption of organic cation transporter 3 attenuates the pharmacologic response to metformin. Mol. Pharmacol. 2015, 88, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Zaharenko, L.; Kalnina, I.; Geldnere, K.; Konrade, I.; Grinberga, S.; Židzik, J.; Javorský, M.; Lejnieks, A.; Nikitina-Zake, L.; Fridmanis, D.; et al. Single nucleotide polymorphisms in the intergenic region between metformin transporter OCT2 and OCT3 coding genes are associated with short-Term response to metformin monotherapy in type 2 diabetes mellitus patients. Eur. J. Endocrinol. 2016, 175, 531–540. [Google Scholar] [CrossRef]
- Kang, H.J.; Song, I.S.; Ho, J.S.; Kim, W.Y.; Lee, C.H.; Shim, J.C.; Zhou, H.H.; Sang, S.L.; Shin, J.G. Identification and functional characterization of genetic variants of human organic cation transporters in a Korean population. Drug Metab. Dispos. 2007, 35, 667–675. [Google Scholar] [CrossRef] [Green Version]
- Graham, G.G.; Punt, J.; Arora, M.; Day, R.O.; Doogue, M.P.; Duong, J.K.; Furlong, T.J.; Greenfield, J.R.; Greenup, L.C.; Kirkpatrick, C.M.; et al. Clinical pharmacokinetics of metformin. Clin. Pharmacokinet. 2011, 50, 81–98. [Google Scholar] [CrossRef]
- Scheen, A.J. Clinical pharmacokinetics of metformin. Clin. Pharmacokinet. 1996, 30, 359–371. [Google Scholar] [CrossRef]
- Zhou, K.; Yee, S.W.; Seiser, E.L.; Van Leeuwen, N.; Tavendale, R.; Bennett, A.J.; Groves, C.J.; Coleman, R.L.; Van Der Heijden, A.A.; Beulens, J.W.; et al. Variation in the glucose transporter gene SLC2A2 is associated with glycemic response to metformin. Nat. Genet. 2016, 48, 1055–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotroff, D.M.; Yee, S.W.; Zhou, K.; Marvel, S.W.; Shah, H.S.; Jack, J.R.; Havener, T.M.; Hedderson, M.M.; Kubo, M.; Herman, M.A.; et al. Genetic variants in CPA6 and PRPF31 are associated with variation in response to metformin in individuals with type 2 diabetes. Diabetes 2018, 67, 1428–1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, Y.; Sheardown, S.A.; Brown, C.; Owen, R.P.; Zhang, S.; Castro, R.A.; Ianculescu, A.G.; Yue, L.; Lo, J.C.; Burchard, E.G.; et al. Effect of genetic variation in the organic cation transporter 1 (OCT1) on metformin action. J. Clin. Investig. 2007, 117, 1422–1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, M.M.H.; Højlund, K.; Hother-Nielsen, O.; Stage, T.B.; Damkier, P.; Beck-Nielsen, H.; Brøsen, K. Steady-state pharmacokinetics of metformin is independent of the OCT1 genotype in healthy volunteers. Eur. J. Clin. Pharmacol. 2015, 71, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Christensen, M.M.H.; Højlund, K.; Hother-Nielsen, O.; Stage, T.B.; Damkier, P.; Beck-Nielsen, H.; Brøsen, K. Endogenous glucose production increases in response to metformin treatment in the glycogen-depleted state in humans: A randomised trial. Diabetologia 2015, 58, 2494–2502. [Google Scholar] [CrossRef] [Green Version]
- Dujic, T.; Zhou, K.; Donnelly, L.A.; Tavendale, R.; Palmer, C.N.A.; Pearson, E.R. Association of organic cation transporter 1 with intolerance to metformin in type 2 diabetes: A GoDARTS study. Diabetes 2015, 64, 1786–17931. [Google Scholar] [CrossRef] [Green Version]
- Becker, M.L.; Visser, L.E.; van Schaik, R.H.N.; Hofman, A.; Uitterlinden, A.G.; Stricker, B.H.C. Genetic variation in the organic cation transporter 1 is associated with metformin response in patients with diabetes mellitus. Pharmacogenom. J. 2009, 9, 242–247. [Google Scholar] [CrossRef]
- Dujic, T.; Zhou, K.; Yee, S.W.; van Leeuwen, N.; de Keyser, C.E.; Javorský, M.; Goswami, S.; Zaharenko, L.; Hougaard Christensen, M.M.; Out, M.; et al. Variants in Pharmacokinetic Transporters and Glycemic Response to Metformin: A Metgen Meta-Analysis. Clin. Pharmacol. Ther. 2017, 101, 763–772. [Google Scholar] [CrossRef] [Green Version]
- Goswami, S.; Yee, S.W.; Xu, F.; Sridhar, S.B.; Mosley, J.D.; Takahashi, A.; Kubo, M.; Maeda, S.; Davis, R.L.; Roden, D.M.; et al. A Longitudinal HbA1c Model Elucidates Genes Linked to Disease Progression on Metformin. Clin. Pharmacol. Ther. 2016, 100, 537–547. [Google Scholar] [CrossRef]
- Chen, Y.; Li, S.; Brown, C.; Cheatham, S.; Castro, R.A.; Leabman, M.K.; Urban, T.J.; Chen, L.; Yee, S.W.; Choi, J.H.; et al. Effect of genetic variation in the organic cation transporter 2 on the renal elimination of metformin. Pharmacogenet. Genom. 2009, 19, 497–504. [Google Scholar] [CrossRef] [Green Version]
- Yanagisawa, R.; Kubota, N.; Hidaka, E.; Sakashita, K.; Tanaka, M.; Nakazawa, Y.; Nakamura, T. Cisplatin-induced nephrotoxicity in patients with advanced neuroblastoma. Pediatr. Blood Cancer 2018, 65, 1–2. [Google Scholar] [CrossRef]
- Iwata, K.; Aizawa, K.; Kamitsu, S.; Jingami, S.; Fukunaga, E.; Yoshida, M.; Yoshimura, M.; Hamada, A.; Saito, H. Effects of genetic variants in SLC22A2 organic cation transporter 2 and SLC47A1 multidrug and toxin extrusion 1 transporter on cisplatin-induced adverse events. Clin. Exp. Nephrol. 2012, 16, 843–851. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhou, W. Ameliorative effects of SLC22A2 gene polymorphism 808 G/T and cimetidine on cisplatin-induced nephrotoxicity in Chinese cancer patients. Food Chem. Toxicol. 2012, 50, 2289–2293. [Google Scholar] [CrossRef] [PubMed]
- Qian, C.-Y.; Zheng, Y.; Wang, Y.; Chen, J.; Liu, J.-Y.; Zhou, H.-H.; Yin, J.-Y.; Liu, Z.-Q. Associations of genetic polymorphisms of the transporters organic cation transporter 2 (OCT2), multidrug and toxin extrusion 1 (MATE1), and ATP-binding cassette subfamily C member 2 (ABCC2) with platinum-based chemotherapy response and toxicity in non-sma. Chin. J. Cancer 2016, 35, 85. [Google Scholar] [CrossRef] [Green Version]
- Ciarimboli, G.; Deuster, D.; Knief, A.; Sperling, M.; Holtkamp, M.; Edemir, B.; Pavenstädt, H.; Lanvers-Kaminsky, C.; Zehnhoff-Dinnesen, A.A.; Schinkel, A.H.; et al. Organic cation transporter 2 mediates cisplatin-induced oto- and nephrotoxicity and is a target for protective interventions. Am. J. Pathol. 2010, 176, 1169–1180. [Google Scholar] [CrossRef]
- Zazuli, Z.; Vijverberg, S.; Slob, E.; Liu, G.; Carleton, B.; Veltman, J.; Baas, P.; Masereeuw, R.; Maitland-Van Der Zee, A.H. Genetic variations and cisplatin nephrotoxicity: A systematic review. Front. Pharmacol. 2018, 9, 1111. [Google Scholar] [CrossRef]
- Hinai, Y.; Motoyama, S.; Niioka, T.; Miura, M. Absence of effect of SLC22A2 genotype on cisplatin-induced nephrotoxicity in oesophageal cancer patients receiving cisplatin and 5-fluorouracil: Report of results discordant with those of earlier studies. J. Clin. Pharm. Ther. 2013, 38, 498–503. [Google Scholar] [CrossRef]
- Williams, L.K.; Padhukasahasram, B.; Ahmedani, B.K.; Peterson, E.L.; Wells, K.E.; Burchard, E.G.; Lanfear, D.E. Differing effects of metformin on glycemic control by race-ethnicity. J. Clin. Endocrinol. Metab. 2014, 99, 3160–3168. [Google Scholar] [CrossRef] [Green Version]
- Tanner, C.; Wang, G.; Liu, N.; Andrikopoulos, S.; Zajac, J.D.; Ekinci, E.I. Metformin: Time to review its role and safety in chronic kidney disease. Med. J. Aust. 2019, 211, 37–42. [Google Scholar] [CrossRef]
- Rocha, A.; Almeida, M.; Santos, J.; Carvalho, A. Metformin in patients with chronic kidney disease: Strengths and weaknesses. J. Nephrol. 2013, 26, 55–60. [Google Scholar] [CrossRef]
- Mato, E.P.M.; Guewo-Fokeng, M.; Essop, M.F.; Owira, P.M.O. Genetic polymorphisms of organic cation transporter 1 (OCT1) and responses to metformin therapy in individuals with type 2 diabetes. Medicine 2018, 97. [Google Scholar] [CrossRef]
- Pearson, E.R. Diabetes: Is There a Future for Pharmacogenomics Guided Treatment? Clin. Pharmacol. Ther. 2019, 106, 329–337. [Google Scholar] [CrossRef] [Green Version]
- Manohar, S.; Leung, N. Cisplatin nephrotoxicity: A review of the literature. J. Nephrol. 2018, 31, 15–25. [Google Scholar] [CrossRef]
- Trendowski, M.R.; El-Charif, O.; Ratain, M.J.; Monahan, P.; Mu, Z.; Wheeler, H.E.; Dinh, P.C.; Feldman, D.R.; Ardeshir-Rouhani-Fard, S.; Hamilton, R.J.; et al. Clinical and genome-wide analysis of serum platinum levels after cisplatin-based chemotherapy. Clin. Cancer Res. 2019, 25, 5913–5924. [Google Scholar] [CrossRef]
- Thiesen, S.; Yin, P.; Jorgensen, A.L.; Zhang, J.E.; Manzo, V.; McEvoy, L.; Barton, C.; Picton, S.; Bailey, S.; Brock, P.; et al. TPMT, COMT and ACYP2 genetic variants in paediatric cancer patients with cisplatin-induced ototoxicity. Pharmacogenet. Genom. 2017, 27, 213–222. [Google Scholar] [CrossRef]
- Teft, W.A.; Winquist, E.; Nichols, A.C.; Kuruvilla, S.; Richter, S.; Parker, C.; Francis, P.; Trinnear, M.; Lukovic, J.; Bukhari, N.; et al. Predictors of cisplatin-induced ototoxicity and survival in chemoradiation treated head and neck cancer patients. Oral Oncol. 2019, 89, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Ross, C.J.D.; Katzov-Eckert, H.; Dubé, M.P.; Brooks, B.; Rassekh, S.R.; Barhdadi, A.; Feroz-Zada, Y.; Visscher, H.; Brown, A.M.K.; Rieder, M.J.; et al. Genetic variants in TPMT and COMT are associated with hearing loss in children receiving cisplatin chemotherapy. Nat. Genet. 2009, 41, 1345–1349. [Google Scholar] [CrossRef]
- Yang, J.J.; Lim, J.Y.S.; Huang, J.; Bass, J.; Wu, J.; Wang, C.; Fang, J.; Stewart, E.; Harstead EH, E.S.; Robinson, G.W.; et al. The role of inherited TPMT and COMT genetic variation in cisplatin-induced ototoxicity in children with cancer. Clin. Pharmacol. Ther. 2013, 94, 252–259. [Google Scholar] [CrossRef] [Green Version]
- Hagleitner, M.M.; Coenen, M.J.H.; Patino-Garcia, A.; De Bont, E.S.J.M.; Gonzalez-Neira, A.; Vos, H.I.; Van Leeuwen, F.N.; Gelderblom, H.; Hoogerbrugge, P.M.; Guchelaar, H.J.; et al. Influence of genetic variants in TPMT and COMT associated with cisplatin induced hearing loss in patients with cancer: Two new cohorts and a meta-analysis reveal significant heterogeneity between cohorts. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [Green Version]
- Drögemöller, B.I.; Monzon, J.G.; Bhavsar, A.P.; Borrie, A.E.; Brooks, B.; Wright, G.E.B.; Liu, G.; Renouf, D.J.; Kollmannsberger, C.K.; Bedard, P.L.; et al. Association between slc16a5 genetic variation and cisplatin-induced ototoxic effects in adult patients with testicular cancer. JAMA Oncol. 2017, 3, 1558–1562. [Google Scholar] [CrossRef] [Green Version]
- Lui, G.; Bouazza, N.; Denoyelle, F.; Moine, M.; Brugières, L.; Chastagner, P.; Corradini, N.; Entz-Werle, N.; Vérité, C.; Landmanparker, J.; et al. Association between genetic polymorphisms and platinuminduced ototoxicity in children. Oncotarget 2018, 9, 30883–30893. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Robinson, G.W.; Huang, J.; Lim, J.Y.S.; Zhang, H.; Bass, J.K.; Broniscer, A.; Chintagumpala, M.; Bartels, U.; Gururangan, S.; et al. Common variants in ACYP2 influence susceptibility to cisplatin-induced hearing loss. Nat. Genet. 2015, 47, 263–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheeler, H.E.; Gamazon, E.R.; Frisina, R.D.; Perez-Cervantes, C.; El Charif, O.; Mapes, B.; Fossa, S.D.; Feldman, D.R.; Hamilton, R.J.; Vaughn, D.J.; et al. Variants in WFS1 and other mendelian deafness genes are associated with cisplatin-associated ototoxicity. Clin. Cancer Res. 2017, 23, 3325–3333. [Google Scholar] [CrossRef] [Green Version]
- Drögemöller, B.I.; Wright, G.E.B.; Lo, C.; Le, T.; Brooks, B.; Bhavsar, A.P.; Rassekh, S.R.; Ross, C.J.D.; Carleton, B.C. Pharmacogenomics of Cisplatin-Induced Ototoxicity: Successes, Shortcomings, and Future Avenues of Research. Clin. Pharmacol. Ther. 2019, 106, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Griffin, B.R.; Faubel, S.; Edelstein, C.L. Biomarkers of drug-induced kidney toxicity. Ther. Drug Monit. 2019, 41, 213–226. [Google Scholar] [CrossRef]



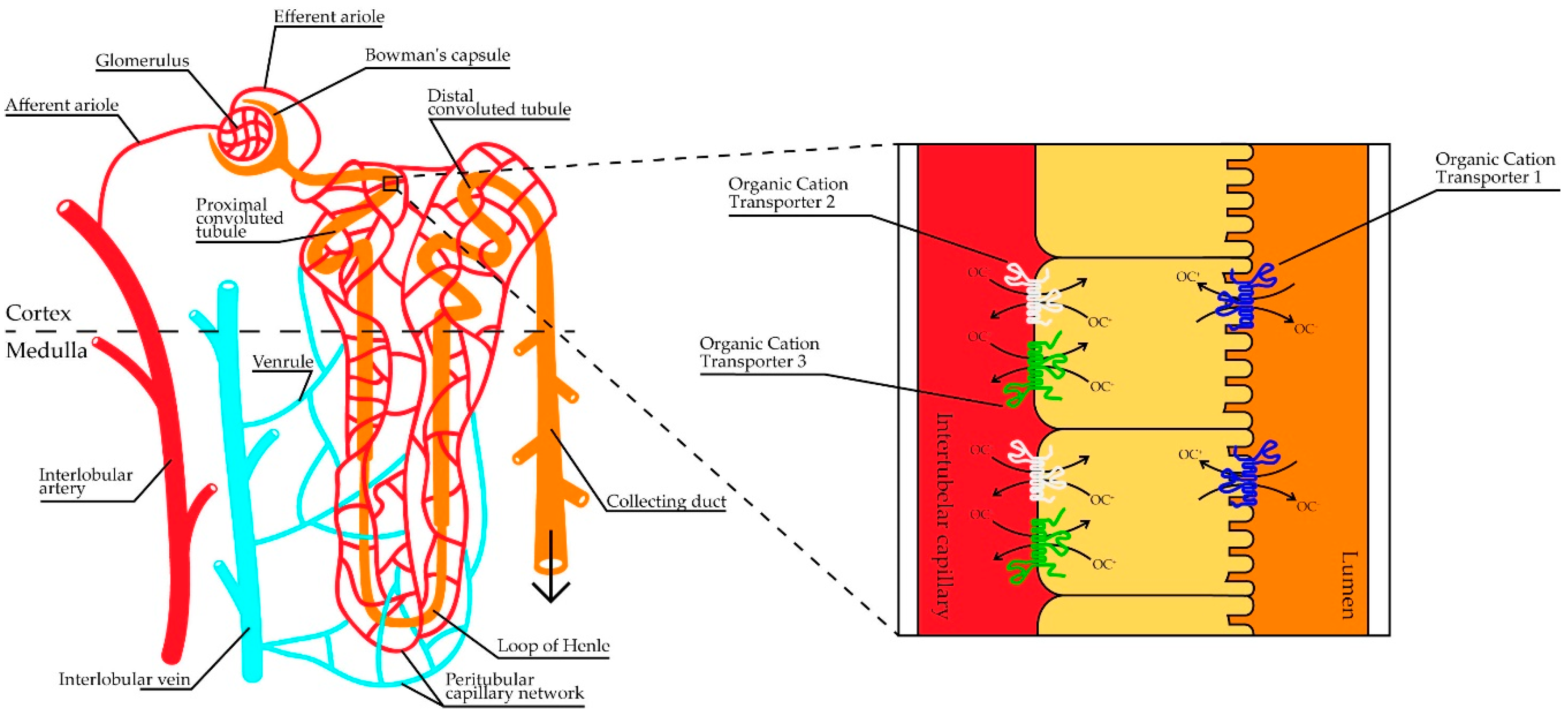{kind=link}
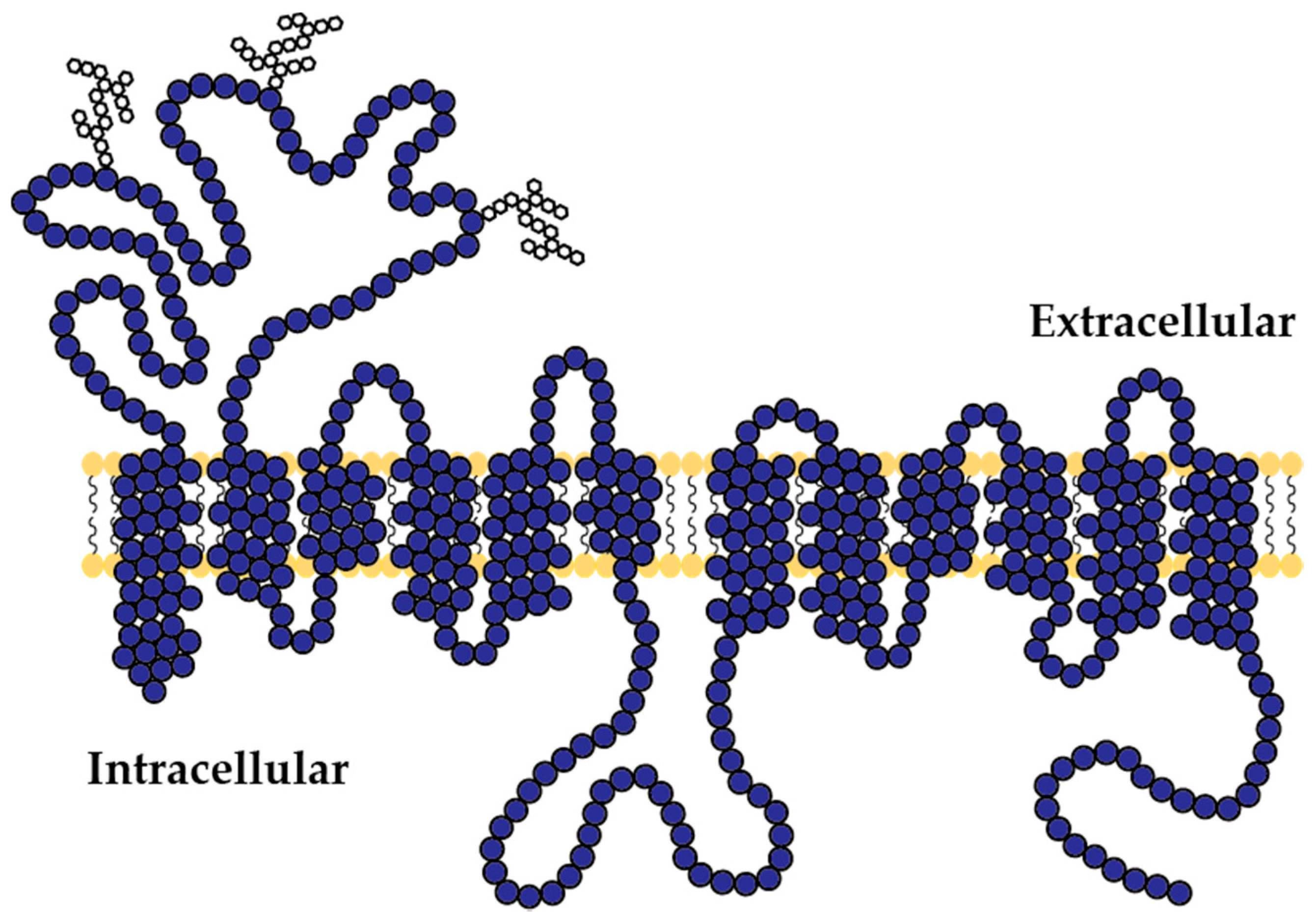{kind=link}
| Type of OCT | Endogenous Substrates | Drug Substrates | References | ||
|---|---|---|---|---|---|
| OCT1 | monoamine neurotransmitters ** norepinephrine ** serotonin ** histamine ** dopamine ** | Acyclovir albuterol * amiloride * amisulpride ** atenolol * atropine * barberine ** cimetidine * clidinium daunorubicin debrisoquine diltiazem evafirenz fenoterol * | furamidine ganciclovir ipratropium * irinotecan lamivudine ** lamotrigine metformin ** metoclopramide * oxaliplatin ** paclitacel pentamidine picoplatin * procainamide * | Ranitidine * salbutamol selegiline sumatriptan * terazosin * terbutaline * tiotropium * triamterene * trimethoprim * tropisetron varenicline * zalcitabine | [8,21,25,26] |
| OCT2 | creatinine monoamine neurotransmitters ** norepinephrine ** serotonin ** histamine ** dopamine ** | Albuterol * amantadine amiloride * amiodarone amisulpride ** atenolol * atropine * barberine ** cimetidine * cisplatin famotidine fenoterol * | ganciclovir ifosfamide ipratropium * lamivudine ** memantine metformin ** metoclopramide * oxaliplatin ** picoplatin * procainamide * | Ranitidine * Sumatriptan * Terazosin * Terbutaline * Tiotropium * Triamterene * Trimethoprim * Varenicline * zalcitabine zidovudine | [1,21,27] |
| OCT3 | monoamine neurotransmitters ** dopamine ** norepinephrine ** serotonin ** histamine ** | Amisulpride ** berberine ** cisplatin etilefrine | Lamuvidine ** lidocaine metformin ** | pramipexole oxaliplatin ** quinidine | [8,21] |
| Gene | Polymorphism | AA * | Allele Change | Annotation (DNA Strand **) | Alleles | MAF per Ethnicity # | References | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Minor | Major | EU | Af | As | EA | SA | C | LA | Other | ||||||
| SLC22A1 (OCT1) | rs12208357 | R61C | 181C>T | Coding (plus) | T | C | 0.0723 | 0.0281 | 0 | 0 | 0.04 | 0.037 | 0.0216 | 0.0701 | [12,20,39,40,41,42,43] |
| rs55918055 | C88R | 262T>C | Coding (plus) | C | T | 0.00303 | 0.006 | 0 | 0 | 0 | 0 | 0 | 0 | [12,44,45] | |
| rs34130495 | G401S | 1201G>A | Coding (plus) | A | G | 0.0261 | 0.005 | 0 | 0 | 0 | 0.02 | 0.0127 | 0.0197 | [39,45,46] | |
| rs72552763 | M420del | 1260-1262delGAT | Coding (plus) | del | GAT | 0.1148 | 0.053 | 0 | 0 | 0 | 0 | 0 | 0.0805 | [12,20,39,40,41,43,46] | |
| rs34059508 | G465R | 1393G>A | Coding (plus) | A | G | 0.02274 | 0.008 | 0 | 0 | 0 | 0 | 0 | 0.0194 | [12,20,39,40,43,45,47] | |
| rs628031 | M408V | 1222A>G | Coding (plus) | A | G | 0.402712 | 0.2663 | 0.3 | 0.308 | 0.3666 | N/D | 0.2058 | 0.3701 | [20,40,43,48,49] | |
| rs683369 | F160L | 480G>C | Coding (plus) | G | C | 0.1977 | 0.123 | 0.22 | 0.36 | 0.2 | N/D *** | 0 | 0.1712 | [48,50,51,52] | |
| N/A **** | R206C | 616C>T | Coding (plus) | T | C | N/D | N/D | 0.008 1 | N/D | N/D | N/D | N/D | N/D | [48] | |
| N/A | Q97K | 289C>A | Coding (plus) | A | C | N/D | N/D | 0.017 1 | N/D | N/D | N/D | N/D | N/D | [48] | |
| rs200684404 | P117L | 350C>T | Coding (plus) | T | C | 0.00008 | 0.001 | 0.012 | 0.018 | 0 | N/D | 0 | 0.0004 | [48] | |
| rs34447885 | S14F | 41C>T | Coding (plus) | T | C | 0.00033 | 0.012 | 0 | 0 | 0 | N/D | 0 | 0.0011 | [20,40,53] | |
| rs36103319 | G220V | 659G>T | Coding (plus) | T | G | 0.00013 | 0 | 0 | 0 | N/D | N/D | N/D | 0 | [44] | |
| rs34104736 | S189L | 566C>T | Coding (plus) | T | C | 0.00152 | 0 | 0 | 0 | N/D | N/D | N/D | 0.0014 | [47,53,54] | |
| rs2282143 | P341L | 1022C>T | Coding (plus) | T | C | 0.014788 | 0.0621 | 0.094 | 0.086 | 0.08 | 0.069 | 0.0384 | 0.0181 | [47,49,53] | |
| rs622342 | N/A | C>A | Intron 9 | C | A | 0.363843 | 0.1871 | 0.192 | 0.155 | 0.2764 | N/D | 0.3689 | 0.3048 | [53,55] | |
| SLC22A2 (OCT2) | rs316019 | A270S | 808A>C | Coding (minus) | A | C | 0.103508 | 0.1541 | 0.11 | 0.11 | 0.1156 | N//D | 0.0533 | 0.10206 | [20,43,56,57,58,59,60,61,62,63] |
| rs8177516 | R400S | 1198G>T | Coding (minus) | T | G | 0.000205 | 0.0121 | 0 | 0 | 0.0002 | 0.026 | N/D | 0.0014 | [44,47] | |
| rs8177517 | K432Q | 1294A>G | Coding (minus) | G | A | 0.001096 | 0.0243 | 0 | 0 | 0.0022 | 0.003 | N/D | 0.0022 | [44,47] | |
| rs8177507 | M165I | 495G>A | Coding (minus) | A | G | 0.000009 | 0.0062 | 0 | 0 | 0 | 0 | N/D | 0.0002 | [44,64] | |
| rs201919874 | T199I | 596C>T | Coding (minus) | T | C | 0 2 | 02 | 02 | 0.0007 2 | 0 2 | N/D | N/D | N/D | [63,65] | |
| rs596881 | N/A | T>C | 3′UTR (plus) | T | C | 0.101541 | 0.2983 | 0.104 | 0.102 | 0.1294 | 0.136 | 0.0707 | 0.1224 | [66,67] | |
| rs145450955 | T201M | 602C>T | Coding (minus) | T | C | 0.00001 | 0 | 0.045 | 0 | N/D | N/D | N/D | 0.0001 | [57,58,62,68] | |
| SLC22A3 (OCT3) | rs2292334 | A411 | 1233G>A | Coding (plus) | A | G | 0.364435 | 0.1458 | 0.448 | 0.403 | 0.2702 | 0.331 | 0.5086 | 0.3458 | [69,70,71] |
| rs8187715 | T44M | 131C>T | Coding (plus) | T | T | 0.006 4 | 0.006 3 | 0.012 3 | N/D | N/D | N/D | N/D | 0.0009 | [72] | |
| rs8187717 | A116S | 346G>T | Coding (plus) | T | G | 0 | 0.0017 4 | 0 | 0 | 0 | 0 | 0 | 0 | [72] | |
| rs8187725 | T400I | 1199C>T | Coding (plus) | T | C | 0.00005 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | [72,73,74] | |
| rs12212246 | A439V | 1316C>T | Coding (plus) | T | C | 0.00001 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | [73,74] | |
| rs9365165 | G475S | 1423G>A | Coding (plus) | A | G | 0.00013 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | [73,74] | |
| rs8187722 | L346 | 1038A>G | Coding (plus) | G | A | 0.001233 | 0.0338 | 0 | 0 | 0.0002 | 0 | 0 | 0.0079 | [69,72] | |
| N/A | V423F | 1267G>T | Coding (plus) | T | G | 0 4 | 0 4 | N/D | 0.068 4 | N/D | N/D | N/D | N/D | [72] | |
| rs3088442 | N/A | 564G>A | 3′UTR (plus) | A | G | 0.36755 | 0.08969 | 0.35 | 0.35 | 0.08 | 0.5 | 0.92 | 0.335 | [70,71] | |
| rs555754 | N/A | −29G>A | 5′UTR (plus) | A | G | 0.46766 | 0.5441 | 0.25 | 0.21 | 0.33 | 0.515 | 0.3764 | 0.477 | [75,76] | |
| rs60515630 | N/A | −81G>delAG | Upstream | G | del | 0.0029 | 0.1 | N/D | N/D | N/D | N/D | N/D | N/D | [75,76] | |
| rs376563 | N/A | 976-6046T>C | Intron 5 (plus) | T | C | 0.47480 | 0.2523 | 0.35 | 0.386 | 0.47 | 0.444 | 0.3652 | 0.4119 | [67] | |
| rs2076828 | N/A | 698C>G | 3′UTR (plus) | G | C | 0.4249 | 0.46 | 0.8 | 0.5377 | 0.8 | N/D | N/D | 0.4 | [77] | |
| rs2481030 | N/A | A>G | Intergenic | G | A | 0.3404 | 0.177 | 0.17 | 0.17 | 0.81 | N/D | 0.2671 | 0.306 | [78] | |
| Gene | SNP | Ref | Drugs and Chemicals | Effect |
|---|---|---|---|---|
| SLC22A1 (OCT1) | rs12208357 | [20,39,41,42,43,87] | Metformin, morphine | Reduced uptake activity; decrease in steady-state concentration of metformin; associated with metformin intolerance |
| rs55918055 | [44,45] | Metformin | Reduced uptake activity | |
| rs34130495 | [39,45,46,87] | Metformin, * MPP+ | Reduced uptake activity; decrease in steady-state concentration of metformin; associated with metformin intolerance | |
| rs72552763 | [20,39,40,41,42,43,46,87] | Metformin, morphine, MPP+ | Reduced uptake activity and decreased morphine clearance; decrease in steady-state concentration of metformin; associated with metformin intolerance | |
| rs34059508 | [20,39,43,45,47,87] | Metformin, MPP+ | Reduced uptake activity; decrease in steady-state concentration of metformin; associated with metformin intolerance | |
| rs628031 | [20,43,48,49] | Metformin, imatinib | Reduced imatinib clearance | |
| rs683369 | [48,50,51,52] | Metformin, imatinib | Reduced function and reduced imatinib clearance | |
| R206C | [48] | Metformin | Reduced uptake activity, reduced function | |
| Q97K | [48] | Metformin | Reduced uptake activity | |
| rs200684404 | [48] | Metformin | Reduced uptake activity | |
| rs34447885 | [20,53] | Metformin | Reduced uptake activity | |
| rs36103319 | [44] | Metformin | Reduced uptake activity | |
| rs34104736 | [47,53,54] | Metformin | Reduced uptake activity | |
| rs2282143 | [47,49,53] | MPP+ | Reduced uptake activity | |
| rs622342 | [39,53,55,88] | Metformin | Reduced uptake activity; decrease in steady-state concentration of metformin; smaller HbA1c lowering effect | |
| SLC22A2 (OCT2) | rs316019 | [20,43,56,57,58,59,60,61,62,63,66,90,91,92,93,94,95,96] | Metformin, cisplatin, creatinine, MPP+, lamivudine | Reduced uptake activity, lower renal clearance of metformin, higher renal clearance of metformin, lower HbA1c levels in metformin users, lower nephrotoxicity, higher nephrotoxicity, lower hematotoxicity, lower hepatotoxicity |
| rs8177516 | [44,47] | Metformin, MPP+, ** TBA | Reduced uptake activity | |
| rs8177517 | [44,47] | Metformin, MPP+, TBA | Reduced uptake activity | |
| rs8177507 | [44,64] | Metformin | Reduced uptake activity | |
| rs201919874 | [63,65] | Metformin, MPP+, lamivudine | Damaged protein, reduced activity | |
| rs596881 | [66,67] | N/A | Renoprotective effect and maintenance of eGFR, hypertension | |
| rs145450955 | [57,58,62,68] | Metformin, MPP+, lamivudine, insulin | Reduced activity, changed insulin resistance | |
| SLCC22A3 (OCT3) | rs2292334 | [69,70,71] | Metformin | Reduced activity |
| rs8187715 | [72] | Metformin, * MPP+, *** catecholamines | Enhanced uptake activity | |
| rs8187717 | [72] | Catecholamines, metformin, MPP+, histamine | Reduced uptake activity | |
| rs8187725 | [72,73] | Catecholamines, metformin, MPP+, histamine | Reduced uptake activity | |
| rs12212246 | [73] | Catecholamines, metformin, MPP+, histamine | Reduced uptake activity | |
| rs9365165 | [73] | Histamine | Reduced histamine uptake | |
| rs8187722 | [69,72] | Metformin | Reduced uptake activity | |
| V423F | [72] | Catecholamines | Reduced uptake activity | |
| rs3088442 | [70,71] | Metformin | Genetic risk marker for T2DM #, A allele has protective effect | |
| rs555754 | [75] | N/A | Higher transcription rate, higher expression | |
| rs60515630 | [75] | N/A | Higher transcription rate, higher expression | |
| rs376563 | [67] | N/A | Effect on diabetic nephropathy and hypertension | |
| rs2076828 | [77] | Metformin | Reduced response to metformin | |
| rs2481030 | [78] | Metformin | Metformin inefficiency |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zazuli, Z.; Duin, N.J.C.B.; Jansen, K.; Vijverberg, S.J.H.; Maitland-van der Zee, A.H.; Masereeuw, R. The Impact of Genetic Polymorphisms in Organic Cation Transporters on Renal Drug Disposition. Int. J. Mol. Sci. 2020, 21, 6627. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186627
Zazuli Z, Duin NJCB, Jansen K, Vijverberg SJH, Maitland-van der Zee AH, Masereeuw R. The Impact of Genetic Polymorphisms in Organic Cation Transporters on Renal Drug Disposition. International Journal of Molecular Sciences. 2020; 21(18):6627. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186627
Chicago/Turabian StyleZazuli, Zulfan, Naut J. C. B. Duin, Katja Jansen, Susanne J. H. Vijverberg, Anke H. Maitland-van der Zee, and Rosalinde Masereeuw. 2020. "The Impact of Genetic Polymorphisms in Organic Cation Transporters on Renal Drug Disposition" International Journal of Molecular Sciences 21, no. 18: 6627. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186627