Polyphenol-Mediated Autophagy in Cancer: Evidence of In Vitro and In Vivo Studies

 ,
,  , ,
, ,  ,
, 

Abstract
:1. Introduction
2. Polyphenols
3. In Vitro- and In Vivo-Mediated Autophagy by Polyphenols
3.1. Flavonoids
3.1.1. Flavonols
3.1.2. Flavan-3-ols
3.1.3. Flavones
3.1.4. Anthocyanins
3.1.5. Flavanones
3.1.6. Isoflavones
3.2. Non-Flavonoids
3.2.1. Coumarins
3.2.2. Curcuminoids
3.2.3. Phenolic Acids
3.2.4. Lignans
3.2.5. Stilbenes
3.2.6. Xanthones
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- White, E.; Green, D.R.; Letai, A.G. Apoptosis, Necrosis, and Autophagy. In The Molecular Basis of Cancer, 4th ed.; Mendelsohn, J., Howley, P.M., Thompson, C.B., Israel, M.A., Gray, J.W., Eds.; Saunders: Philadelphia, PA, USA, 2014; pp. 209–228.e203. [Google Scholar] [CrossRef]
- Bhattacharyya, S.; Yu, H.; Mim, C.; Matouschek, A. Regulated protein turnover: Snapshots of the proteasome in action. Nat. Rev. Mol. Cell Biol. 2014, 15, 122–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galluzzi, L.; Pietrocola, F.; Levine, B.; Kroemer, G. Metabolic control of autophagy. Cell 2014, 159, 1263–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tooze, S.A.; Abada, A.; Elazar, Z. Endocytosis and autophagy: Exploitation or cooperation? Cold Spring Harb. Perspect. Biol. 2014, 6, a018358. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Baehrecke, E.H.; Ballabio, A.; Boya, P.; Bravo-San Pedro, J.M.; Cecconi, F.; Choi, A.M.; Chu, C.T.; Codogno, P.; Colombo, M.I.; et al. Molecular definitions of autophagy and related processes. EMBO J. 2017, 36, 1811–1836. [Google Scholar] [CrossRef]
- Hosokawa, N.; Hara, T.; Kaizuka, T.; Kishi, C.; Takamura, A.; Miura, Y.; Iemura, S.; Natsume, T.; Takehana, K.; Yamada, N.; et al. Nutrient-dependent mTORC1 association with the ULK1-Atg13-FIP200 complex required for autophagy. Mol. Biol. Cell 2009, 20, 1981–1991. [Google Scholar] [CrossRef] [Green Version]
- Nazio, F.; Strappazzon, F.; Antonioli, M.; Bielli, P.; Cianfanelli, V.; Bordi, M.; Gretzmeier, C.; Dengjel, J.; Piacentini, M.; Fimia, G.M.; et al. mTOR inhibits autophagy by controlling ULK1 ubiquitylation, self-association and function through AMBRA1 and TRAF6. Nat. Cell Biol. 2013, 15, 406–416. [Google Scholar] [CrossRef]
- Mihaylova, M.M.; Shaw, R.J. The AMPK signalling pathway coordinates cell growth, autophagy and metabolism. Nat. Cell Biol. 2011, 13, 1016–1023. [Google Scholar] [CrossRef]
- Russell, R.C.; Tian, Y.; Yuan, H.; Park, H.W.; Chang, Y.Y.; Kim, J.; Kim, H.; Neufeld, T.P.; Dillin, A.; Guan, K.L. ULK1 induces autophagy by phosphorylating Beclin-1 and activating VPS34 lipid kinase. Nat. Cell Biol. 2013, 15, 741–750. [Google Scholar] [CrossRef] [Green Version]
- Mizushima, N.; Yoshimori, T.; Ohsumi, Y. Role of the Apg12 conjugation system in mammalian autophagy. Int. J. Biochem. Cell Biol. 2003, 35, 553–561. [Google Scholar] [CrossRef]
- Geng, J.; Klionsky, D.J. The Atg8 and Atg12 ubiquitin-like conjugation systems in macroautophagy. ‘Protein modifications: Beyond the usual suspects’ review series. EMBO Rep. 2008, 9, 859–864. [Google Scholar] [CrossRef] [Green Version]
- Galluzzi, L.; Pietrocola, F.; Bravo-San Pedro, J.M.; Amaravadi, R.K.; Baehrecke, E.H.; Cecconi, F.; Codogno, P.; Debnath, J.; Gewirtz, D.A.; Karantza, V.; et al. Autophagy in malignant transformation and cancer progression. EMBO J. 2015, 34, 856–880. [Google Scholar] [CrossRef] [PubMed]
- Weidberg, H.; Shvets, E.; Shpilka, T.; Shimron, F.; Shinder, V.; Elazar, Z. LC3 and GATE-16/GABARAP subfamilies are both essential yet act differently in autophagosome biogenesis. EMBO J. 2010, 29, 1792–1802. [Google Scholar] [CrossRef] [PubMed]
- Itakura, E.; Kishi-Itakura, C.; Mizushima, N. The hairpin-type tail-anchored SNARE syntaxin 17 targets to autophagosomes for fusion with endosomes/lysosomes. Cell 2012, 151, 1256–1269. [Google Scholar] [CrossRef] [Green Version]
- Huynh, K.K.; Eskelinen, E.L.; Scott, C.C.; Malevanets, A.; Saftig, P.; Grinstein, S. LAMP proteins are required for fusion of lysosomes with phagosomes. EMBO J. 2007, 26, 313–324. [Google Scholar] [CrossRef]
- Liu, X.M.; Sun, L.L.; Hu, W.; Ding, Y.H.; Dong, M.Q.; Du, L.L. ESCRTs Cooperate with a Selective Autophagy Receptor to Mediate Vacuolar Targeting of Soluble Cargos. Mol. Cell 2015, 59, 1035–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaushik, S.; Cuervo, A.M. The coming of age of chaperone-mediated autophagy. Nat. Rev. Mol. Cell Biol. 2018, 19, 365–381. [Google Scholar] [CrossRef] [PubMed]
- Cuervo, A.M.; Dice, J.F. A receptor for the selective uptake and degradation of proteins by lysosomes. Science 1996, 273, 501–503. [Google Scholar] [CrossRef]
- Bandyopadhyay, U.; Kaushik, S.; Varticovski, L.; Cuervo, A.M. The chaperone-mediated autophagy receptor organizes in dynamic protein complexes at the lysosomal membrane. Mol. Cell Biol. 2008, 28, 5747–5763. [Google Scholar] [CrossRef] [Green Version]
- Bandyopadhyay, U.; Sridhar, S.; Kaushik, S.; Kiffin, R.; Cuervo, A.M. Identification of regulators of chaperone-mediated autophagy. Mol. Cell 2010, 39, 535–547. [Google Scholar] [CrossRef] [Green Version]
- Dice, J.F. Peptide sequences that target cytosolic proteins for lysosomal proteolysis. Trends. Biochem. Sci. 1990, 15, 305–309. [Google Scholar] [CrossRef]
- Kaushik, S.; Massey, A.C.; Cuervo, A.M. Lysosome membrane lipid microdomains: Novel regulators of chaperone-mediated autophagy. EMBO J. 2006, 25, 3921–3933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujiwara, Y.; Furuta, A.; Kikuchi, H.; Aizawa, S.; Hatanaka, Y.; Konya, C.; Uchida, K.; Yoshimura, A.; Tamai, Y.; Wada, K.; et al. Discovery of a novel type of autophagy targeting RNA. Autophagy 2013, 9, 403–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galluzzi, L.; Bravo-San Pedro, J.M.; Levine, B.; Green, D.R.; Kroemer, G. Pharmacological modulation of autophagy: Therapeutic potential and persisting obstacles. Nat. Rev. Drug Discov. 2017, 16, 487–511. [Google Scholar] [CrossRef] [Green Version]
- Rybstein, M.D.; Bravo-San Pedro, J.M.; Kroemer, G.; Galluzzi, L. The autophagic network and cancer. Nat. Cell Biol. 2018, 20, 243–251. [Google Scholar] [CrossRef]
- Liu, Y.; Levine, B. Autosis and autophagic cell death: The dark side of autophagy. Cell Death Differ. 2015, 22, 367–376. [Google Scholar] [CrossRef] [Green Version]
- Nah, J.; Zablocki, D.; Sadoshima, J. Autosis: A New Target to Prevent Cell Death. JACC Basic Transl. Sci. 2020, 5, 857–869. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.H.; Piao, S.; Wang, D.; McAfee, Q.W.; Nathanson, K.L.; Lum, J.J.; Li, L.Z.; Amaravadi, R.K. Measurements of tumor cell autophagy predict invasiveness, resistance to chemotherapy, and survival in melanoma. Clin. Cancer Res. 2011, 17, 3478–3489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Wang, G.; Zhou, Y.; Chen, Y.; Ouyang, L.; Liu, B. Mechanisms of autophagy and relevant small-molecule compounds for targeted cancer therapy. Cell Mol. Life Sci. 2018, 75, 1803–1826. [Google Scholar] [CrossRef]
- Deng, S.; Shanmugam, M.K.; Kumar, A.P.; Yap, C.T.; Sethi, G.; Bishayee, A. Targeting autophagy using natural compounds for cancer prevention and therapy. Cancer 2019, 125, 1228–1246. [Google Scholar] [CrossRef]
- Manach, C.; Scalbert, A.; Morand, C.; Remesy, C.; Jimenez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [Green Version]
- Scalbert, A.; Manach, C.; Morand, C.; Remesy, C.; Jimenez, L. Dietary polyphenols and the prevention of diseases. Crit. Rev. Food Sci. Nutr. 2005, 45, 287–306. [Google Scholar] [CrossRef] [PubMed]
- Di Carlo, G.; Mascolo, N.; Izzo, A.A.; Capasso, F. Flavonoids: Old and new aspects of a class of natural therapeutic drugs. Life Sci. 1999, 65, 337–353. [Google Scholar] [CrossRef]
- Fantini, M.; Benvenuto, M.; Masuelli, L.; Frajese, G.V.; Tresoldi, I.; Modesti, A.; Bei, R. In vitro and in vivo antitumoral effects of combinations of polyphenols, or polyphenols and anticancer drugs: Perspectives on cancer treatment. Int. J. Mol. Sci. 2015, 16, 9236–9282. [Google Scholar] [CrossRef] [Green Version]
- Beecher, G.R. Overview of dietary flavonoids: Nomenclature, occurrence and intake. J. Nutr. 2003, 133, 3248S–3254S. [Google Scholar] [CrossRef] [PubMed]
- Marzocchella, L.; Fantini, M.; Benvenuto, M.; Masuelli, L.; Tresoldi, I.; Modesti, A.; Bei, R. Dietary flavonoids: Molecular mechanisms of action as anti- inflammatory agents. Recent Pat. Inflamm. Allergy Drug Discov. 2011, 5, 200–220. [Google Scholar] [CrossRef] [PubMed]
- Crozier, A.; Jaganath, I.B.; Clifford, M.N. Dietary phenolics: Chemistry, bioavailability and effects on health. Nat. Prod. Rep. 2009, 26, 1001–1043. [Google Scholar] [CrossRef]
- Bei, R.; Masuelli, L.; Turriziani, M.; Li Volti, G.; Malaguarnera, M.; Galvano, F. Impaired expression and function of signaling pathway enzymes by anthocyanins: Role on cancer prevention and progression. Curr. Enzym. Inhib. 2009, 5, 184–197. [Google Scholar] [CrossRef]
- Bei, R.; Palumbo, C.; Masuelli, L.; Turriziani, M.; Frajese, G.V.; Li Volti, G.; Malaguarnera, M.; Galvano, F. Impaired expression and function of cancer-related enzymes by anthocyans: An update. Curr. Enzym. Inhib. 2012, 8, 2–21. [Google Scholar] [CrossRef]
- Tomas-Barberan, F.A.; Clifford, M.N. Flavanones, chalcones and dihydrochalcones—Nature, occurrence and dietary burden. J. Sci. Food Agric. 2000, 80, 1073–1080. [Google Scholar] [CrossRef]
- Cassidy, A.; Hanley, B.; Lamuela-Raventos, M.R. Isoflavones, lignans and stilbenes—Origins, metabolism and potential importance to human health. J. Sci. Food Agric. 2000, 80, 1044–1062. [Google Scholar] [CrossRef]
- Sakai, T.; Kogiso, M. Soy isoflavones and immunity. J. Med. Investig. 2008, 55, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Majnooni, M.B.; Fakhri, S.; Smeriglio, A.; Trombetta, D.; Croley, C.R.; Bhattacharyya, P.; Sobarzo-Sanchez, E.; Farzaei, M.H.; Bishayee, A. Antiangiogenic Effects of Coumarins against Cancer: From Chemistry to Medicine. Molecules 2019, 24, 4278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkata Sairam, K.; Gurupadayya, B.M.; Chandan, R.S.; Nagesha, D.K.; Vishwanathan, B. A Review on Chemical Profile of Coumarins and their Therapeutic Role in the Treatment of Cancer. Curr. Drug Deliv. 2016, 13, 186–201. [Google Scholar] [CrossRef]
- Prasad, S.; Tyagi, A.K.; Aggarwal, B.B. Recent developments in delivery, bioavailability, absorption and metabolism of curcumin: The golden pigment from golden spice. Cancer Res. Treat. 2014, 46, 2–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amalraj, A.; Pius, A.; Gopi, S.; Gopi, S. Biological activities of curcuminoids, other biomolecules from turmeric and their derivatives—A review. J. Tradit. Complement. Med. 2017, 7, 205–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowland, I.; Faughnan, M.; Hoey, L.; Wahala, K.; Williamson, G.; Cassidy, A. Bioavailability of phyto-oestrogens. Br. J. Nutr. 2003, 89 (Suppl. 1), S45–S58. [Google Scholar] [CrossRef] [PubMed]
- Adlercreutz, H. Lignans and human health. Crit. Rev. Clin. Lab. Sci. 2007, 44, 483–525. [Google Scholar] [CrossRef]
- De Silva, S.F.; Alcorn, J. Flaxseed Lignans as Important Dietary Polyphenols for Cancer Prevention and Treatment: Chemistry, Pharmacokinetics, and Molecular Targets. Pharmaceuticals 2019, 12, 68. [Google Scholar] [CrossRef] [Green Version]
- Chong, J.; Poutaraud, A.; Hugueney, P. Metabolism and role of stilbenes in plants. Plant Sci. 2009, 177, 143–155. [Google Scholar] [CrossRef]
- Vervandier-Fasseur, D.; Latruffe, N. The Potential Use of Resveratrol for Cancer Prevention. Molecules 2019, 24, 4506. [Google Scholar] [CrossRef] [Green Version]
- El-Seedi, H.R.; Salem, M.A.; Khattab, O.M.; El-Wahed, A.A.; El-Kersh, D.M.; Khalifa, S.A.M.; Saeed, A.; Abdel-Daim, M.M.; Hajrah, N.H.; Alajlani, M.M.; et al. Dietary Xanthones. In Handbook of Dietary Phytochemicals; Xiao, J., Sarker, S.D., Asakawa, Y., Eds.; Springer Nature Singapore Pte Ltd.: Singapore, 2020. [Google Scholar] [CrossRef]
- Negi, J.S.; Bisht, V.K.; Singh, P.; Rawat, M.S.M.; Joshi, G.P. Naturally Occurring Xanthones: Chemistry and Biology. J. Appl. Chem. 2013, 2013, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Benvenuto, M.; Fantini, M.; Masuelli, L.; De Smaele, E.; Zazzeroni, F.; Tresoldi, I.; Calabrese, G.; Galvano, F.; Modesti, A.; Bei, R. Inhibition of ErbB receptors, Hedgehog and NF-kappaB signaling by polyphenols in cancer. Front. Biosci. (Landmark Ed) 2013, 18, 1290–1310. [Google Scholar] [CrossRef] [PubMed]
- Benvenuto, M.; Mattera, R.; Taffera, G.; Giganti, M.G.; Lido, P.; Masuelli, L.; Modesti, A.; Bei, R. The Potential Protective Effects of Polyphenols in Asbestos-Mediated Inflammation and Carcinogenesis of Mesothelium. Nutrients 2016, 8, 275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Focaccetti, C.; Izzi, V.; Benvenuto, M.; Fazi, S.; Ciuffa, S.; Giganti, M.G.; Potenza, V.; Manzari, V.; Modesti, A.; Bei, R. Polyphenols as Immunomodulatory Compounds in the Tumor Microenvironment: Friends or Foes? Int. J. Mol. Sci. 2019, 20, 1714. [Google Scholar] [CrossRef] [Green Version]
- Mattera, R.; Benvenuto, M.; Giganti, M.G.; Tresoldi, I.; Pluchinotta, F.R.; Bergante, S.; Tettamanti, G.; Masuelli, L.; Manzari, V.; Modesti, A.; et al. Effects of Polyphenols on Oxidative Stress-Mediated Injury in Cardiomyocytes. Nutrients 2017, 9, 523. [Google Scholar] [CrossRef] [Green Version]
- Klappan, A.K.; Hones, S.; Mylonas, I.; Bruning, A. Proteasome inhibition by quercetin triggers macroautophagy and blocks mTOR activity. Histochem. Cell Biol. 2012, 137, 25–36. [Google Scholar] [CrossRef]
- Granato, M.; Rizzello, C.; Romeo, M.A.; Yadav, S.; Santarelli, R.; D’Orazi, G.; Faggioni, A.; Cirone, M. Concomitant reduction of c-Myc expression and PI3K/AKT/mTOR signaling by quercetin induces a strong cytotoxic effect against Burkitt’s lymphoma. Int. J. Biochem. Cell Biol. 2016, 79, 393–400. [Google Scholar] [CrossRef]
- Jia, L.; Huang, S.; Yin, X.; Zan, Y.; Guo, Y.; Han, L. Quercetin suppresses the mobility of breast cancer by suppressing glycolysis through Akt-mTOR pathway mediated autophagy induction. Life Sci. 2018, 208, 123–130. [Google Scholar] [CrossRef]
- Moon, J.H.; Eo, S.K.; Lee, J.H.; Park, S.Y. Quercetin-induced autophagy flux enhances TRAIL-mediated tumor cell death. Oncol. Rep. 2015, 34, 375–381. [Google Scholar] [CrossRef] [Green Version]
- Calgarotto, A.K.; Maso, V.; Junior, G.C.F.; Nowill, A.E.; Filho, P.L.; Vassallo, J.; Saad, S.T.O. Antitumor activities of Quercetin and Green Tea in xenografts of human leukemia HL60 cells. Sci. Rep. 2018, 8, 3459. [Google Scholar] [CrossRef] [Green Version]
- Jakubowicz-Gil, J.; Langner, E.; Badziul, D.; Wertel, I.; Rzeski, W. Quercetin and sorafenib as a novel and effective couple in programmed cell death induction in human gliomas. Neurotox. Res. 2014, 26, 64–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Moon, J.Y.; Ahn, K.S.; Cho, S.K. Quercetin induces mitochondrial mediated apoptosis and protective autophagy in human glioblastoma U373MG cells. Oxidative Med. Cell Longev. 2013, 2013, 596496. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Shen, C.; Li, C.; Liu, Y.; Gao, D.; Shi, C.; Peng, F.; Liu, Z.; Zhao, B.; Zheng, Z.; et al. Inhibition of autophagy induced by quercetin at a late stage enhances cytotoxic effects on glioma cells. Tumor Biol. 2016, 37, 3549–3560. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Liu, R.; Li, J.; Mao, J.; Lei, Y.; Wu, J.; Zeng, J.; Zhang, T.; Wu, H.; Chen, L.; et al. Quercetin induces protective autophagy in gastric cancer cells: Involvement of Akt-mTOR- and hypoxia-induced factor 1alpha-mediated signaling. Autophagy 2011, 7, 966–978. [Google Scholar] [CrossRef] [Green Version]
- Granato, M.; Rizzello, C.; Gilardini Montani, M.S.; Cuomo, L.; Vitillo, M.; Santarelli, R.; Gonnella, R.; D’Orazi, G.; Faggioni, A.; Cirone, M. Quercetin induces apoptosis and autophagy in primary effusion lymphoma cells by inhibiting PI3K/AKT/mTOR and STAT3 signaling pathways. J. Nutr. Biochem. 2017, 41, 124–136. [Google Scholar] [CrossRef]
- Liu, Y.; Gong, W.; Yang, Z.Y.; Zhou, X.S.; Gong, C.; Zhang, T.R.; Wei, X.; Ma, D.; Ye, F.; Gao, Q.L. Quercetin induces protective autophagy and apoptosis through ER stress via the p-STAT3/Bcl-2 axis in ovarian cancer. Apoptosis 2017, 22, 544–557. [Google Scholar] [CrossRef]
- Maso, V.; Calgarotto, A.K.; Franchi, G.C., Jr.; Nowill, A.E.; Filho, P.L.; Vassallo, J.; Saad, S.T. Multitarget effects of quercetin in leukemia. Cancer Prev. Res. 2014, 7, 1240–1250. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.L.; Chow, J.M.; Chang, J.H.; Wen, Y.C.; Lin, Y.W.; Yang, S.F.; Lee, W.J.; Chien, M.H. Quercetin simultaneously induces G0/G1 -phase arrest and caspase-mediated crosstalk between apoptosis and autophagy in human leukemia HL-60 cells. Environ. Toxicol. 2017, 32, 1857–1868. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, W.; Lv, Q.; Zhang, J.; Zhu, D. The critical role of quercetin in autophagy and apoptosis in HeLa cells. Tumor Biol. 2016, 37, 925–929. [Google Scholar] [CrossRef]
- Enayat, S.; Seyma Ceyhan, M.; Taskoparan, B.; Stefek, M.; Banerjee, S. CHNQ, a novel 2-Chloro-1,4-naphthoquinone derivative of quercetin, induces oxidative stress and autophagy both in vitro and in vivo. Arch. Biochem. Biophys. 2016, 596, 84–98. [Google Scholar] [CrossRef]
- Zhao, Y.; Fan, D.; Zheng, Z.P.; Li, E.T.; Chen, F.; Cheng, K.W.; Wang, M. 8-C-(E-phenylethenyl)quercetin from onion/beef soup induces autophagic cell death in colon cancer cells through ERK activation. Mol. Nutr. Food Res. 2017, 61, 1600437. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.X.; Zou, B.Y.; Shi, L.; Du, L.Y.; Zhu, Y.Y.; Jiang, Y.M.; Ma, X.D.; Kang, X.H.; Wang, C.Y.; Zhen, Y.H.; et al. 7-O-geranylquercetin-induced autophagy contributes to apoptosis via ROS generation in human non-small cell lung cancer cells. Life Sci. 2017, 180, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Fu, T.; Wang, L.; Jin, X.N.; Sui, H.J.; Liu, Z.; Jin, Y. Hyperoside induces both autophagy and apoptosis in non-small cell lung cancer cells in vitro. Acta Pharmacol. Sin. 2016, 37, 505–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Ji, M.; Han, Y.; Guo, Y.; Zhu, W.; Gao, F.; Yang, X.; Zhang, C. PGRMC1-dependent autophagy by hyperoside induces apoptosis and sensitizes ovarian cancer cells to cisplatin treatment. Int. J. Oncol. 2017, 50, 835–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, Y.; Hu, K.; Chen, H. Autophagy inhibition enhances isorhamnetininduced mitochondriadependent apoptosis in nonsmall cell lung cancer cells. Mol. Med. Rep. 2015, 12, 5796–5806. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Sun, S.; Li, N.; Ho, A.S.W.; Kiang, K.M.Y.; Zhang, X.; Cheng, Y.S.; Poon, M.W.; Lee, D.; Pu, J.K.S.; et al. Rutin increases the cytotoxicity of temozolomide in glioblastoma via autophagy inhibition. J. Neurooncol. 2017, 132, 393–400. [Google Scholar] [CrossRef]
- Alzaharna, M.; Alqouqa, I.; Cheung, H.Y. Taxifolin synergizes Andrographolide-induced cell death by attenuation of autophagy and augmentation of caspase dependent and independent cell death in HeLa cells. PLoS ONE 2017, 12, e0171325. [Google Scholar] [CrossRef] [Green Version]
- Lou, M.; Zhang, L.N.; Ji, P.G.; Feng, F.Q.; Liu, J.H.; Yang, C.; Li, B.F.; Wang, L. Quercetin nanoparticles induced autophagy and apoptosis through AKT/ERK/Caspase-3 signaling pathway in human neuroglioma cells: In vitro and in vivo. Biomed. Pharmacother. 2016, 84, 1–9. [Google Scholar] [CrossRef]
- Luo, C.L.; Liu, Y.Q.; Wang, P.; Song, C.H.; Wang, K.J.; Dai, L.P.; Zhang, J.Y.; Ye, H. The effect of quercetin nanoparticle on cervical cancer progression by inducing apoptosis, autophagy and anti-proliferation via JAK2 suppression. Biomed. Pharmacother. 2016, 82, 595–605. [Google Scholar] [CrossRef]
- Huang, W.W.; Tsai, S.C.; Peng, S.F.; Lin, M.W.; Chiang, J.H.; Chiu, Y.J.; Fushiya, S.; Tseng, M.T.; Yang, J.S. Kaempferol induces autophagy through AMPK and AKT signaling molecules and causes G2/M arrest via downregulation of CDK1/cyclin B in SK-HEP-1 human hepatic cancer cells. Int. J. Oncol. 2013, 42, 2069–2077. [Google Scholar] [CrossRef]
- Han, X.; Liu, C.F.; Gao, N.; Zhao, J.; Xu, J. Kaempferol suppresses proliferation but increases apoptosis and autophagy by up-regulating microRNA-340 in human lung cancer cells. Biomed. Pharmacother. 2018, 108, 809–816. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Ma, C. Kaempferol suppresses human gastric cancer SNU-216 cell proliferation, promotes cell autophagy, but has no influence on cell apoptosis. Braz. J. Med. Biol. Res. 2019, 52, e7843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.W.; Lee, S.Y.; Kim, M.; Cheon, C.; Ko, S.G. Kaempferol induces autophagic cell death via IRE1-JNK-CHOP pathway and inhibition of G9a in gastric cancer cells. Cell Death Dis. 2018, 9, 875. [Google Scholar] [CrossRef] [PubMed]
- Budisan, L.; Gulei, D.; Jurj, A.; Braicu, C.; Zanoaga, O.; Cojocneanu, R.; Pop, L.; Raduly, L.; Barbat, A.; Moldovan, A.; et al. Inhibitory Effect of CAPE and Kaempferol in Colon Cancer Cell Lines-Possible Implications in New Therapeutic Strategies. Int. J. Mol. Sci. 2019, 20, 1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.L.; Dong, J.L.; Wu, J. Juglanin induces apoptosis and autophagy in human breast cancer progression via ROS/JNK promotion. Biomed. Pharmacother. 2017, 85, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Fudhaili, A.; Oh, S.S.; Lee, K.W.; Madhi, H.; Kim, D.H.; Yoo, J.; Ryu, H.W.; Park, K.H.; Kim, K.D. Cytotoxic effects of kazinol A derived from Broussonetia papyrifera on human bladder cancer cells, T24 and T24R2. Phytomedicine 2016, 23, 1462–1468. [Google Scholar] [CrossRef]
- Tan, M.; Jiang, B.; Wang, H.; Ouyang, W.; Chen, X.; Wang, T.; Dong, D.; Yi, S.; Yi, J.; Huang, Y.; et al. Dihydromyricetin induced lncRNA MALAT1-TFEB-dependent autophagic cell death in cutaneous squamous cell carcinoma. J. Cancer 2019, 10, 4245–4255. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Guo, S.; Fang, T.; Feng, D.; Zhang, X.; Zhang, Q.; Liu, J.; Liu, B.; Li, M.; Zhu, R. Dihydromyricetin induces autophagy in HepG2 cells involved in inhibition of mTOR and regulating its upstream pathways. Food Chem. Toxicol. 2014, 66, 7–13. [Google Scholar] [CrossRef]
- Zhou, D.Z.; Sun, H.Y.; Yue, J.Q.; Peng, Y.; Chen, Y.M.; Zhong, Z.J. Dihydromyricetin induces apoptosis and cytoprotective autophagy through ROS-NF-kappaB signalling in human melanoma cells. Free Radic. Res. 2017, 51, 517–528. [Google Scholar] [CrossRef]
- Fan, T.F.; Wu, T.F.; Bu, L.L.; Ma, S.R.; Li, Y.C.; Mao, L.; Sun, Z.J.; Zhang, W.F. Dihydromyricetin promotes autophagy and apoptosis through ROS-STAT3 signaling in head and neck squamous cell carcinoma. Oncotarget 2016, 7, 59691–59703. [Google Scholar] [CrossRef] [Green Version]
- Chiu, C.T.; Hsuan, S.W.; Lin, H.H.; Hsu, C.C.; Chou, F.P.; Chen, J.H. Hibiscus sabdariffa leaf polyphenolic extract induces human melanoma cell death, apoptosis, and autophagy. J. Food Sci. 2015, 80, H649–H658. [Google Scholar] [CrossRef]
- Irimie, A.I.; Braicu, C.; Zanoaga, O.; Pileczki, V.; Gherman, C.; Berindan-Neagoe, I.; Campian, R.S. Epigallocatechin-3-gallate suppresses cell proliferation and promotes apoptosis and autophagy in oral cancer SSC-4 cells. Onco Targets Ther. 2015, 8, 461–470. [Google Scholar] [CrossRef] [Green Version]
- Wei, R.; Mao, L.; Xu, P.; Zheng, X.; Hackman, R.M.; Mackenzie, G.G.; Wang, Y. Suppressing glucose metabolism with epigallocatechin-3-gallate (EGCG) reduces breast cancer cell growth in preclinical models. Food Funct. 2018, 9, 5682–5696. [Google Scholar] [CrossRef]
- Zhao, L.; Liu, S.; Xu, J.; Li, W.; Duan, G.; Wang, H.; Yang, H.; Yang, Z.; Zhou, R. A new molecular mechanism underlying the EGCG-mediated autophagic modulation of AFP in HepG2 cells. Cell Death Dis. 2017, 8, e3160. [Google Scholar] [CrossRef] [Green Version]
- Enkhbat, T.; Nishi, M.; Yoshikawa, K.; Jun, H.; Tokunaga, T.; Takasu, C.; Kashihara, H.; Ishikawa, D.; Tominaga, M.; Shimada, M. Epigallocatechin-3-gallate Enhances Radiation Sensitivity in Colorectal Cancer Cells Through Nrf2 Activation and Autophagy. Anticancer Res. 2018, 38, 6247–6252. [Google Scholar] [CrossRef]
- Hsieh, C.H.; Lu, C.H.; Kuo, Y.Y.; Chen, W.T.; Chao, C.Y. Studies on the non-invasive anticancer remedy of the triple combination of epigallocatechin gallate, pulsed electric field, and ultrasound. PLoS ONE 2018, 13, e0201920. [Google Scholar] [CrossRef] [Green Version]
- Grube, S.; Ewald, C.; Kogler, C.; Lawson McLean, A.; Kalff, R.; Walter, J. Achievable Central Nervous System Concentrations of the Green Tea Catechin EGCG Induce Stress in Glioblastoma Cells in Vitro. Nutr. Cancer 2018, 70, 1145–1158. [Google Scholar] [CrossRef]
- Satoh, M.; Takemura, Y.; Hamada, H.; Sekido, Y.; Kubota, S. EGCG induces human mesothelioma cell death by inducing reactive oxygen species and autophagy. Cancer Cell Int. 2013, 13, 19. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Yun, J.P.; Yang, Y.N.; Hua, F.; Zhang, X.W.; Lin, H.; Lv, X.X.; Li, K.; Zhang, P.C.; Hu, Z.W. A novel ECG analog 4-(S)-(2,4,6-trimethylthiobenzyl)-epigallocatechin gallate selectively induces apoptosis of B16-F10 melanoma via activation of autophagy and ROS. Sci. Rep. 2017, 7, 42194. [Google Scholar] [CrossRef] [Green Version]
- Izdebska, M.; Klimaszewska-Wisniewska, A.; Halas, M.; Gagat, M.; Grzanka, A. Green tea extract induces protective autophagy in A549 non-small lung cancer cell line. Postepy. Hig. Med. Dosw. 2015, 69, 1478–1484. [Google Scholar]
- Rizzi, F.; Naponelli, V.; Silva, A.; Modernelli, A.; Ramazzina, I.; Bonacini, M.; Tardito, S.; Gatti, R.; Uggeri, J.; Bettuzzi, S. Polyphenon E(R), a standardized green tea extract, induces endoplasmic reticulum stress, leading to death of immortalized PNT1a cells by anoikis and tumorigenic PC3 by necroptosis. Carcinogenesis 2014, 35, 828–839. [Google Scholar] [CrossRef] [Green Version]
- Gu, W.; Lin, Y.; Gou, X.; He, W. Tea Polyphenol inhibits autophagy to sensitize Epirubicin-induced apoptosis in human bladder cancer cells. Neoplasma 2017, 64, 674–680. [Google Scholar] [CrossRef]
- Wang, Q.; He, W.Y.; Zeng, Y.Z.; Hossain, A.; Gou, X. Inhibiting autophagy overcomes docetaxel resistance in castration-resistant prostate cancer cells. Int. Urol. Nephrol. 2018, 50, 675–686. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Ye, H.L.; Zhang, G.; Yao, W.M.; Chen, X.Z.; Zhang, F.C.; Liang, G. Autophagy inhibition contributes to the synergistic interaction between EGCG and doxorubicin to kill the hepatoma Hep3B cells. PLoS ONE 2014, 9, e85771. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Chen, D.; Zhu, K. SOX2OT variant 7 contributes to the synergistic interaction between EGCG and Doxorubicin to kill osteosarcoma via autophagy and stemness inhibition. J. Exp. Clin. Cancer Res. 2018, 37, 37. [Google Scholar] [CrossRef] [Green Version]
- Meng, J.; Chang, C.; Chen, Y.; Bi, F.; Ji, C.; Liu, W. EGCG overcomes gefitinib resistance by inhibiting autophagy and augmenting cell death through targeting ERK phosphorylation in NSCLC. Onco Targets Ther. 2019, 12, 6033–6043. [Google Scholar] [CrossRef] [Green Version]
- Hu, F.; Wei, F.; Wang, Y.; Wu, B.; Fang, Y.; Xiong, B. EGCG synergizes the therapeutic effect of cisplatin and oxaliplatin through autophagic pathway in human colorectal cancer cells. J. Pharmacol. Sci. 2015, 128, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Yuan, C.H.; Horng, C.T.; Lee, C.F.; Chiang, N.N.; Tsai, F.J.; Lu, C.C.; Chiang, J.H.; Hsu, Y.M.; Yang, J.S.; Chen, F.A. Epigallocatechin gallate sensitizes cisplatin-resistant oral cancer CAR cell apoptosis and autophagy through stimulating AKT/STAT3 pathway and suppressing multidrug resistance 1 signaling. Environ. Toxicol. 2017, 32, 845–855. [Google Scholar] [CrossRef]
- Modernelli, A.; Naponelli, V.; Giovanna Troglio, M.; Bonacini, M.; Ramazzina, I.; Bettuzzi, S.; Rizzi, F. EGCG antagonizes Bortezomib cytotoxicity in prostate cancer cells by an autophagic mechanism. Sci. Rep. 2015, 5, 15270. [Google Scholar] [CrossRef]
- Kim, S.W.; Moon, J.H.; Park, S.Y. Activation of autophagic flux by epigallocatechin gallate mitigates TRAIL-induced tumor cell apoptosis via down-regulation of death receptors. Oncotarget 2016, 7, 65660–65668. [Google Scholar] [CrossRef]
- Brunelli, E.; Pinton, G.; Bellini, P.; Minassi, A.; Appendino, G.; Moro, L. Flavonoid-induced autophagy in hormone sensitive breast cancer cells. Fitoterapia 2009, 80, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, M.; Xu, S.; Chen, H.; Chen, X.; Gu, H. Apigenin restores impairment of autophagy and downregulation of unfolded protein response regulatory proteins in keratinocytes exposed to ultraviolet B radiation. J. Photochem. Photobiol. B 2019, 194, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Ruela-de-Sousa, R.R.; Fuhler, G.M.; Blom, N.; Ferreira, C.V.; Aoyama, H.; Peppelenbosch, M.P. Cytotoxicity of apigenin on leukemia cell lines: Implications for prevention and therapy. Cell Death Dis. 2010, 1, e19. [Google Scholar] [CrossRef] [Green Version]
- Masuelli, L.; Benvenuto, M.; Mattera, R.; Di Stefano, E.; Zago, E.; Taffera, G.; Tresoldi, I.; Giganti, M.G.; Frajese, G.V.; Berardi, G.; et al. In Vitro and In Vivo Anti-tumoral Effects of the Flavonoid Apigenin in Malignant Mesothelioma. Front. Pharmacol. 2017, 8, 373. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Xu, H.; Yu, X.; Wang, X.; Zhu, X.; Xu, X. Apigenin inhibits in vitro and in vivo tumorigenesis in cisplatin-resistant colon cancer cells by inducing autophagy, programmed cell death and targeting m-TOR/PI3K/Akt signalling pathway. J. BUON 2019, 24, 488–493. [Google Scholar] [PubMed]
- Zhang, L.; Cheng, X.; Gao, Y.; Zheng, J.; Xu, Q.; Sun, Y.; Guan, H.; Yu, H.; Sun, Z. Apigenin induces autophagic cell death in human papillary thyroid carcinoma BCPAP cells. Food Funct. 2015, 6, 3464–3472. [Google Scholar] [CrossRef]
- Yang, J.; Pi, C.; Wang, G. Inhibition of PI3K/Akt/mTOR pathway by apigenin induces apoptosis and autophagy in hepatocellular carcinoma cells. Biomed. Pharmacother. 2018, 103, 699–707. [Google Scholar] [CrossRef]
- Lee, Y.; Sung, B.; Kang, Y.J.; Kim, D.H.; Jang, J.Y.; Hwang, S.Y.; Kim, M.; Lim, H.S.; Yoon, J.H.; Chung, H.Y.; et al. Apigenin-induced apoptosis is enhanced by inhibition of autophagy formation in HCT116 human colon cancer cells. Int. J. Oncol. 2014, 44, 1599–1606. [Google Scholar] [CrossRef] [Green Version]
- Chow, S.E.; Chen, Y.W.; Liang, C.A.; Huang, Y.K.; Wang, J.S. Wogonin induces cross-regulation between autophagy and apoptosis via a variety of Akt pathway in human nasopharyngeal carcinoma cells. J. Cell Biochem. 2012, 113, 3476–3485. [Google Scholar] [CrossRef]
- Tan, H.; Li, X.; Yang, W.H.; Kang, Y. A flavone, Wogonin from Scutellaria baicalensis inhibits the proliferation of human colorectal cancer cells by inducing of autophagy, apoptosis and G2/M cell cycle arrest via modulating the PI3K/AKT and STAT3 signalling pathways. J. BUON 2019, 24, 1143–1149. [Google Scholar]
- Hong, Z.P.; Wang, L.G.; Wang, H.J.; Ye, W.F.; Wang, X.Z. Wogonin exacerbates the cytotoxic effect of oxaliplatin by inducing nitrosative stress and autophagy in human gastric cancer cells. Phytomedicine 2018, 39, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Jiang, C.; Chen, W.; Zhang, G.; Luo, D.; Cao, Y.; Wu, J.; Ding, Y.; Liu, B. Baicalein induces apoptosis and autophagy via endoplasmic reticulum stress in hepatocellular carcinoma cells. BioMed Res. Int. 2014, 2014, 732516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.F.; Li, T.; Tang, Z.H.; Chang, L.L.; Zhu, H.; Chen, X.P.; Wang, Y.T.; Lu, J.J. Baicalein Triggers Autophagy and Inhibits the Protein Kinase B/Mammalian Target of Rapamycin Pathway in Hepatocellular Carcinoma HepG2 Cells. Phytother. Res. 2015, 29, 674–679. [Google Scholar] [CrossRef]
- Wang, Y.F.; Xu, Y.L.; Tang, Z.H.; Li, T.; Zhang, L.L.; Chen, X.; Lu, J.H.; Leung, C.H.; Ma, D.L.; Qiang, W.A.; et al. Baicalein Induces Beclin 1- and Extracellular Signal-Regulated Kinase-Dependent Autophagy in Ovarian Cancer Cells. Am. J. Chin. Med. 2017, 45, 123–136. [Google Scholar] [CrossRef]
- Li, B.; Lu, M.; Jiang, X.X.; Pan, M.X.; Mao, J.W.; Chen, M. Inhibiting reactive oxygen species-dependent autophagy enhanced baicalein-induced apoptosis in oral squamous cell carcinoma. J. Nat. Med. 2017, 71, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Ding, L.; Zhang, L.; Wang, S.; Wang, Y.; Wang, B.; Li, L. Baicalein Induces Autophagy and Apoptosis through AMPK Pathway in Human Glioma Cells. Am. J. Chin. Med. 2019, 47, 1405–1418. [Google Scholar] [CrossRef]
- Wang, M.; Qiu, S.; Qin, J. Baicalein induced apoptosis and autophagy of undifferentiated thyroid cancer cells by the ERK/PI3K/Akt pathway. Am. J. Transl. Res. 2019, 11, 3341–3352. [Google Scholar]
- Aryal, P.; Kim, K.; Park, P.H.; Ham, S.; Cho, J.; Song, K. Baicalein induces autophagic cell death through AMPK/ULK1 activation and downregulation of mTORC1 complex components in human cancer cells. FEBS J. 2014, 281, 4644–4658. [Google Scholar] [CrossRef]
- Yan, W.; Ma, X.; Zhao, X.; Zhang, S. Baicalein induces apoptosis and autophagy of breast cancer cells via inhibiting PI3K/AKT pathway in vivo and vitro. Drug Des. Dev. Ther. 2018, 12, 3961–3972. [Google Scholar] [CrossRef] [Green Version]
- Wu, R.; Murali, R.; Kabe, Y.; French, S.W.; Chiang, Y.M.; Liu, S.; Sher, L.; Wang, C.C.; Louie, S.; Tsukamoto, H. Baicalein Targets GTPase-Mediated Autophagy to Eliminate Liver Tumor-Initiating Stem Cell-Like Cells Resistant to mTORC1 Inhibition. Hepatology 2018, 68, 1726–1740. [Google Scholar] [CrossRef]
- Chen, Y.K.; Wang, H.C.; Ho, C.T.; Chen, H.Y.; Li, S.; Chan, H.L.; Chung, T.W.; Tan, K.T.; Li, Y.R.; Lin, C.C. 5-demethylnobiletin promotes the formation of polymerized tubulin, leads to G2/M phase arrest and induces autophagy via JNK activation in human lung cancer cells. J. Nutr. Biochem. 2015, 26, 484–504. [Google Scholar] [CrossRef] [PubMed]
- Verschooten, L.; Barrette, K.; Van Kelst, S.; Rubio Romero, N.; Proby, C.; De Vos, R.; Agostinis, P.; Garmyn, M. Autophagy inhibitor chloroquine enhanced the cell death inducing effect of the flavonoid luteolin in metastatic squamous cell carcinoma cells. PLoS ONE 2012, 7, e48264. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Park, H.S.; Lee, J.H.; Chi, G.Y.; Kim, G.Y.; Moon, S.K.; Chang, Y.C.; Hyun, J.W.; Kim, W.J.; Choi, Y.H. Induction of endoplasmic reticulum stress-mediated apoptosis and non-canonical autophagy by luteolin in NCI-H460 lung carcinoma cells. Food Chem. Toxicol. 2013, 56, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Nazim, U.M.; Park, S.Y. Luteolin sensitizes human liver cancer cells to TRAILinduced apoptosis via autophagy and JNKmediated death receptor 5 upregulation. Int. J. Oncol. 2019, 54, 665–672. [Google Scholar] [CrossRef] [Green Version]
- Cao, Z.; Zhang, H.; Cai, X.; Fang, W.; Chai, D.; Wen, Y.; Chen, H.; Chu, F.; Zhang, Y. Luteolin Promotes Cell Apoptosis by Inducing Autophagy in Hepatocellular Carcinoma. Cell Physiol. Biochem. 2017, 43, 1803–1812. [Google Scholar] [CrossRef]
- Chakrabarti, M.; Ray, S.K. Anti-tumor activities of luteolin and silibinin in glioblastoma cells: Overexpression of miR-7-1-3p augmented luteolin and silibinin to inhibit autophagy and induce apoptosis in glioblastoma in vivo. Apoptosis 2016, 21, 312–328. [Google Scholar] [CrossRef]
- Rafatian, G.; Khodagholi, F.; Farimani, M.M.; Abraki, S.B.; Gardaneh, M. Increase of autophagy and attenuation of apoptosis by Salvigenin promote survival of SH-SY5Y cells following treatment with H(2)O(2). Mol. Cell Biochem. 2012, 371, 9–22. [Google Scholar] [CrossRef]
- Zhang, X.; Tang, X.; Liu, H.; Li, L.; Hou, Q.; Gao, J. Autophagy induced by baicalin involves downregulation of CD147 in SMMC-7721 cells in vitro. Oncol. Rep. 2012, 27, 1128–1134. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.; Tsai, S.C.; Tseng, M.T.; Peng, S.F.; Kuo, S.C.; Lin, M.W.; Hsu, Y.M.; Lee, M.R.; Amagaya, S.; Huang, W.W.; et al. AKT serine/threonine protein kinase modulates baicalin-triggered autophagy in human bladder cancer T24 cells. Int. J. Oncol. 2013, 42, 993–1000. [Google Scholar] [CrossRef] [Green Version]
- Lewinska, A.; Adamczyk-Grochala, J.; Kwasniewicz, E.; Deregowska, A.; Wnuk, M. Diosmin-induced senescence, apoptosis and autophagy in breast cancer cells of different p53 status and ERK activity. Toxicol. Lett. 2017, 265, 117–130. [Google Scholar] [CrossRef]
- Silva, D.F.; Vidal, F.C.; Santos, D.; Costa, M.C.; Morgado-Diaz, J.A.; do Desterro Soares Brandao Nascimento, M.; de Moura, R.S. Cytotoxic effects of Euterpe oleracea Mart. in malignant cell lines. BMC Complement. Altern. Med. 2014, 14, 175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sui, Y.; Yao, H.; Li, S.; Jin, L.; Shi, P.; Li, Z.; Wang, G.; Lin, S.; Wu, Y.; Li, Y.; et al. Delicaflavone induces autophagic cell death in lung cancer via Akt/mTOR/p70S6K signaling pathway. J. Mol. Med. 2017, 95, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Shen, S.; Zhao, X.; Gong, X. Luteoloside induces G0/G1 arrest and pro-death autophagy through the ROS-mediated AKT/mTOR/p70S6K signalling pathway in human non-small cell lung cancer cell lines. Biochem. Biophys. Res. Commun. 2017, 494, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Chen, M.; Zhang, R.; Xu, T. Antitumor Effects of Glychionide-A Flavonoid in Human Pancreatic Carcinoma Cells Are Mediated by Activation of Apoptotic and Autophagic Pathways, Cell Cycle Arrest, and Disruption of Mitochondrial Membrane Potential. Med. Sci. Monit. 2019, 25, 962–969. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Wei, S.; Wang, J.; Liu, X. Isoorientin induces apoptosis and autophagy simultaneously by reactive oxygen species (ROS)-related p53, PI3K/Akt, JNK, and p38 signaling pathways in HepG2 cancer cells. J. Agric. Food. Chem. 2014, 62, 5390–5400. [Google Scholar] [CrossRef]
- Lv, S.X.; Qiao, X. Isovitexin (IV) induces apoptosis and autophagy in liver cancer cells through endoplasmic reticulum stress. Biochem. Biophys. Res. Commun. 2018, 496, 1047–1054. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, H.; Cong, Z.; Xu, J.; Zhu, J.; Ji, X.; Ding, K. Wogonoside induces autophagy-related apoptosis in human glioblastoma cells. Oncol. Rep. 2014, 32, 1179–1187. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.P.; Guo, H.; Wang, X.B. Nobiletin (NOB) suppresses autophagic degradation via over-expressing AKT pathway and enhances apoptosis in multidrug-resistant SKOV3/TAX ovarian cancer cells. Biomed. Pharmacother. 2018, 103, 29–37. [Google Scholar] [CrossRef]
- Moon, J.Y.; Cho, S.K. Nobiletin Induces Protective Autophagy Accompanied by ER-Stress Mediated Apoptosis in Human Gastric Cancer SNU-16 Cells. Molecules 2016, 21, 914. [Google Scholar] [CrossRef] [Green Version]
- Toton, E.; Romaniuk, A.; Budzianowski, J.; Hofmann, J.; Rybczynska, M. Zapotin (5,6,2′,6′-tetramethoxyflavone) Modulates the Crosstalk Between Autophagy and Apoptosis Pathways in Cancer Cells with Overexpressed Constitutively Active PKC. Nutr. Cancer 2016, 68, 290–304. [Google Scholar] [CrossRef]
- He, J.D.; Wang, Z.; Li, S.P.; Xu, Y.J.; Yu, Y.; Ding, Y.J.; Yu, W.L.; Zhang, R.X.; Zhang, H.M.; Du, H.Y. Vitexin suppresses autophagy to induce apoptosis in hepatocellular carcinoma via activation of the JNK signaling pathway. Oncotarget 2016, 7, 84520–84532. [Google Scholar] [CrossRef]
- Chen, J.; Zhu, Y.; Zhang, W.; Peng, X.; Zhou, J.; Li, F.; Han, B.; Liu, X.; Ou, Y.; Yu, X. Delphinidin induced protective autophagy via mTOR pathway suppression and AMPK pathway activation in HER-2 positive breast cancer cells. BMC Cancer 2018, 18, 342. [Google Scholar] [CrossRef]
- Farzaei, M.H.; El-Senduny, F.F.; Momtaz, S.; Parvizi, F.; Iranpanah, A.; Tewari, D.; Naseri, R.; Abdolghaffari, A.H.; Rezaei, N. An update on dietary consideration in inflammatory bowel disease: Anthocyanins and more. Expert. Rev. Gastroenterol. Hepatol. 2018, 12, 1007–1024. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Park, Y.J.; Hwang, S.C.; Kim, K.D.; Moon, D.K.; Kim, D.H. Cytotoxic effects of delphinidin in human osteosarcoma cells. Acta Orthop. Traumatol. Turc. 2018, 52, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Tsuyuki, S.; Fukui, S.; Watanabe, A.; Akune, S.; Tanabe, M.; Yoshida, K. Delphinidin induces autolysosome as well as autophagosome formation and delphinidin-induced autophagy exerts a cell protective role. J. Biochem. Mol. Toxicol. 2012, 26, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, S.; Geng, B.; Yi, Z. Pelargonidin induces antitumor effects in human osteosarcoma cells via autophagy induction, loss of mitochondrial membrane potential, G2/M cell cycle arrest and downregulation of PI3K/AKT signalling pathway. J. BUON 2018, 23, 735–740. [Google Scholar] [PubMed]
- Liu, X.; Zhang, D.; Hao, Y.; Liu, Q.; Wu, Y.; Liu, X.; Luo, J.; Zhou, T.; Sun, B.; Luo, X.; et al. Cyanidin Curtails Renal Cell Carcinoma Tumorigenesis. Cell Physiol. Biochem. 2018, 46, 2517–2531. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Buya, M.; Qin, W.; Sun, C.; Cai, H.; Xie, Q.; Xu, B.; Wu, Y. Anthocyanins from Chinese bayberry extract activate transcription factor Nrf2 in beta cells and negatively regulate oxidative stress-induced autophagy. J. Agric. Food Chem. 2013, 61, 8765–8772. [Google Scholar] [CrossRef]
- Cheng, K.C.; Wang, C.J.; Chang, Y.C.; Hung, T.W.; Lai, C.J.; Kuo, C.W.; Huang, H.P. Mulberry fruits extracts induce apoptosis and autophagy of liver cancer cell and prevent hepatocarcinogenesis in vivo. J. Food Drug Anal. 2020, 28, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Long, H.L.; Zhang, F.F.; Wang, H.L.; Yang, W.S.; Hou, H.T.; Yu, J.K.; Liu, B. Mulberry anthocyanins improves thyroid cancer progression mainly by inducing apoptosis and autophagy cell death. Kaohsiung J. Med. Sci. 2018, 34, 255–262. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Zhou, J.; Luo, J.; Wang, Q.; Liu, J.; Zeng, Q.Q. Study on Mulberry Anthocyanins Induced Autophagy and Apoptosis of Human Gastric Cancer SGC-7901 Cell Autophagy. Zhong Yao Cai 2016, 39, 1134–1138. [Google Scholar] [PubMed]
- Choe, Y.J.; Ha, T.J.; Ko, K.W.; Lee, S.Y.; Shin, S.J.; Kim, H.S. Anthocyanins in the black soybean (Glycine max L.) protect U2OS cells from apoptosis by inducing autophagy via the activation of adenosyl monophosphate-dependent protein kinase. Oncol. Rep. 2012, 28, 2049–2056. [Google Scholar] [CrossRef] [PubMed]
- Schiavano, G.F.; De Santi, M.; Brandi, G.; Fanelli, M.; Bucchini, A.; Giamperi, L.; Giomaro, G. Inhibition of Breast Cancer Cell Proliferation and In Vitro Tumorigenesis by a New Red Apple Cultivar. PLoS ONE 2015, 10, e0135840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Symonds, E.L.; Konczak, I.; Fenech, M. The Australian fruit Illawarra plum (Podocarpus elatus Endl., Podocarpaceae) inhibits telomerase, increases histone deacetylase activity and decreases proliferation of colon cancer cells. Br. J. Nutr. 2013, 109, 2117–2125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Way, T.D.; Tsai, S.J.; Wang, C.M.; Jhan, Y.L.; Ho, C.T.; Chou, C.H. Cinnamtannin D1 from Rhododendron formosanum Induces Autophagy via the Inhibition of Akt/mTOR and Activation of ERK1/2 in Non-Small-Cell Lung Carcinoma Cells. J. Agric. Food Chem. 2015, 63, 10407–10417. [Google Scholar] [CrossRef] [PubMed]
- Weh, K.M.; Howell, A.B.; Kresty, L.A. Expression, modulation, and clinical correlates of the autophagy protein Beclin-1 in esophageal adenocarcinoma. Mol. Carcinog. 2016, 55, 1876–1885. [Google Scholar] [CrossRef]
- Saiprasad, G.; Chitra, P.; Manikandan, R.; Sudhandiran, G. Hesperidin induces apoptosis and triggers autophagic markers through inhibition of Aurora-A mediated phosphoinositide-3-kinase/Akt/mammalian target of rapamycin and glycogen synthase kinase-3 beta signalling cascades in experimental colon carcinogenesis. Eur. J. Cancer 2014, 50, 2489–2507. [Google Scholar] [CrossRef]
- Zhang, Y.S.; Wang, F.; Cui, S.X.; Qu, X.J. Natural dietary compound naringin prevents azoxymethane/dextran sodium sulfate-induced chronic colorectal inflammation and carcinogenesis in mice. Cancer Biol. Ther. 2018, 19, 735–744. [Google Scholar] [CrossRef]
- Raha, S.; Yumnam, S.; Hong, G.E.; Lee, H.J.; Saralamma, V.V.; Park, H.S.; Heo, J.D.; Lee, S.J.; Kim, E.H.; Kim, J.A.; et al. Naringin induces autophagy-mediated growth inhibition by downregulating the PI3K/Akt/mTOR cascade via activation of MAPK pathways in AGS cancer cells. Int. J. Oncol. 2015, 47, 1061–1069. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Wang, K.; Wu, Y.; Chen, Y.; Chen, X.; Hu, C.W.; Hu, F. Pinocembrin induces ER stress mediated apoptosis and suppresses autophagy in melanoma cells. Cancer Lett. 2018, 431, 31–42. [Google Scholar] [CrossRef]
- Shin, S.Y.; Hyun, J.; Yu, J.R.; Lim, Y.; Lee, Y.H. 5-Methoxyflavanone induces cell cycle arrest at the G2/M phase, apoptosis and autophagy in HCT116 human colon cancer cells. Toxicol. Appl. Pharmacol. 2011, 254, 288–298. [Google Scholar] [CrossRef]
- Zhao, Y.; Fan, D.; Ru, B.; Cheng, K.W.; Hu, S.; Zhang, J.; Li, E.T.; Wang, M. 6-C-(E-phenylethenyl)naringenin induces cell growth inhibition and cytoprotective autophagy in colon cancer cells. Eur. J. Cancer 2016, 68, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Jiang, X.; Gao, H.Y.; Gao, S.H. Liquiritin induces apoptosis and autophagy in cisplatin (DDP)-resistant gastric cancer cells in vitro and xenograft nude mice in vivo. Int. J. Oncol. 2017, 51, 1383–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prajapati, V.; Kale, R.K.; Singh, R.P. Silibinin combination with arsenic strongly inhibits survival and invasiveness of human prostate carcinoma cells. Nutr. Cancer 2015, 67, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Gossner, G.; Choi, M.; Tan, L.; Fogoros, S.; Griffith, K.A.; Kuenker, M.; Liu, J.R. Genistein-induced apoptosis and autophagocytosis in ovarian cancer cells. Gynecol. Oncol. 2007, 105, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Prietsch, R.F.; Monte, L.G.; da Silva, F.A.; Beira, F.T.; Del Pino, F.A.; Campos, V.F.; Collares, T.; Pinto, L.S.; Spanevello, R.M.; Gamaro, G.D.; et al. Genistein induces apoptosis and autophagy in human breast MCF-7 cells by modulating the expression of proapoptotic factors and oxidative stress enzymes. Mol. Cell Biochem. 2014, 390, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Cook, K.L.; Warri, A.; Cruz, I.M.; Rosim, M.; Riskin, J.; Helferich, W.; Doerge, D.; Clarke, R.; Hilakivi-Clarke, L. Lifetime Genistein Intake Increases the Response of Mammary Tumors to Tamoxifen in Rats. Clin. Cancer. Res. 2017, 23, 814–824. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Jin, F.; Lian, X.; Li, M.; Wang, G.; Lan, B.; He, H.; Liu, G.D.; Wu, Y.; Sun, G.; et al. Genistein promotes ionizing radiation-induced cell death by reducing cytoplasmic Bcl-xL levels in non-small cell lung cancer. Sci. Rep. 2018, 8, 328. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, R.; Kang, Y.; Li, X.; Roife, D.; Zhang, R.; Fleming, J.B. Genistein potentiates the antitumor effect of 5-Fluorouracil by inducing apoptosis and autophagy in human pancreatic cancer cells. Anticancer Res. 2014, 34, 4685–4692. [Google Scholar]
- Nazim, U.M.; Park, S.Y. Genistein enhances TRAIL-induced cancer cell death via inactivation of autophagic flux. Oncol. Rep. 2015, 34, 2692–2698. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, Y.; Yogosawa, S.; Izutani, Y.; Watanabe, H.; Otsuji, E.; Sakai, T. A combination of indol-3-carbinol and genistein synergistically induces apoptosis in human colon cancer HT-29 cells by inhibiting Akt phosphorylation and progression of autophagy. Mol. Cancer 2009, 8, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, D.; Xiao, Z.; Li, H.P. Puerarin leads to K562 cell apoptosis of chronic myelogenous leukemia via induction of autophagy. J. BUON 2017, 22, 1554–1562. [Google Scholar] [PubMed]
- Hu, Y.; Li, X.; Lin, L.; Liang, S.; Yan, J. Puerarin inhibits non-small cell lung cancer cell growth via the induction of apoptosis. Oncol. Rep. 2018, 39, 1731–1738. [Google Scholar] [CrossRef] [PubMed]
- Alvero, A.B.; Montagna, M.K.; Chen, R.; Kim, K.H.; Kyungjin, K.; Visintin, I.; Fu, H.H.; Brown, D.; Mor, G. NV-128, a novel isoflavone derivative, induces caspase-independent cell death through the Akt/mammalian target of rapamycin pathway. Cancer 2009, 115, 3204–3216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Z.; Bao, X.; Gu, J. Furowanin A-induced autophagy alleviates apoptosis and promotes cell cycle arrest via inactivation STAT3/Mcl-1 axis in colorectal cancer. Life Sci. 2019, 218, 47–57. [Google Scholar] [CrossRef]
- Hsieh, M.J.; Chen, M.K.; Chen, C.J.; Hsieh, M.C.; Lo, Y.S.; Chuang, Y.C.; Chiou, H.L.; Yang, S.F. Glabridin induces apoptosis and autophagy through JNK1/2 pathway in human hepatoma cells. Phytomedicine 2016, 23, 359–366. [Google Scholar] [CrossRef]
- Wang, W.B.; Feng, L.X.; Yue, Q.X.; Wu, W.Y.; Guan, S.H.; Jiang, B.H.; Yang, M.; Liu, X.; Guo, D.A. Paraptosis accompanied by autophagy and apoptosis was induced by celastrol, a natural compound with influence on proteasome, ER stress and Hsp90. J. Cell Physiol. 2012, 227, 2196–2206. [Google Scholar] [CrossRef]
- Deng, Y.N.; Shi, J.; Liu, J.; Qu, Q.M. Celastrol protects human neuroblastoma SH-SY5Y cells from rotenone-induced injury through induction of autophagy. Neurochem. Int. 2013, 63, 1–9. [Google Scholar] [CrossRef]
- Miyamoto, M.; Takano, M.; Aoyama, T.; Soyama, H.; Ishibashi, H.; Kato, K.; Iwahashi, H.; Takasaki, K.; Kuwahara, M.; Matuura, H.; et al. Phenoxodiol Increases Cisplatin Sensitivity in Ovarian Clear Cancer Cells Through XIAP Down-regulation and Autophagy Inhibition. Anticancer Res. 2018, 38, 301–306. [Google Scholar] [CrossRef]
- Dong, M.; Ye, T.; Bi, Y.; Wang, Q.; Kuerban, K.; Li, J.; Feng, M.; Wang, K.; Chen, Y.; Ye, L. A novel hybrid of 3-benzyl coumarin seco-B-ring derivative and phenylsulfonylfuroxan induces apoptosis and autophagy in non-small-cell lung cancer. Phytomedicine 2019, 52, 79–88. [Google Scholar] [CrossRef]
- Wang, Q.; Guo, Y.; Jiang, S.; Dong, M.; Kuerban, K.; Li, J.; Feng, M.; Chen, Y.; Ye, L. A hybrid of coumarin and phenylsulfonylfuroxan induces caspase-dependent apoptosis and cytoprotective autophagy in lung adenocarcinoma cells. Phytomedicine 2018, 39, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Kaewpiboon, C.; Surapinit, S.; Malilas, W.; Moon, J.; Phuwapraisirisan, P.; Tip-Pyang, S.; Johnston, R.N.; Koh, S.S.; Assavalapsakul, W.; Chung, Y.H. Feroniellin A-induced autophagy causes apoptosis in multidrug-resistant human A549 lung cancer cells. Int. J. Oncol. 2014, 44, 1233–1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Yang, C.; Zhang, Q.; Wang, C.; Zhou, X.; Zhang, X.; Liu, S. In vitro anticancer effects of esculetin against human leukemia cell lines involves apoptotic cell death, autophagy, G0/G1 cell cycle arrest and modulation of Raf/MEK/ERK signalling pathway. J. BUON 2019, 24, 1686–1691. [Google Scholar]
- Wen, Q.; Luo, K.; Huang, H.; Liao, W.; Yang, H. Xanthoxyletin Inhibits Proliferation of Human Oral Squamous Carcinoma Cells and Induces Apoptosis, Autophagy, and Cell Cycle Arrest by Modulation of the MEK/ERK Signaling Pathway. Med. Sci. Monit. 2019, 25, 8025–8033. [Google Scholar] [CrossRef] [PubMed]
- Sumorek-Wiadro, J.; Zajac, A.; Badziul, D.; Langner, E.; Skalicka-Wozniak, K.; Maciejczyk, A.; Wertel, I.; Rzeski, W.; Jakubowicz-Gil, J. Coumarins modulate the anti-glioma properties of temozolomide. Eur. J. Pharmacol. 2020, 881, 173207. [Google Scholar] [CrossRef]
- Lin, C.H.; Funayama, S.; Peng, S.F.; Kuo, C.L.; Chung, J.G. The ethanol extraction of prepared Psoralea corylifolia induces apoptosis and autophagy and alteres genes expression assayed by cDNA microarray in human prostate cancer PC-3 cells. Environ. Toxicol. 2018, 33, 770–788. [Google Scholar] [CrossRef]
- Suparji, N.S.; Chan, G.; Sapili, H.; Arshad, N.M.; In, L.L.; Awang, K.; Hasima Nagoor, N. Geranylated 4-Phenylcoumarins Exhibit Anticancer Effects against Human Prostate Cancer Cells through Caspase-Independent Mechanism. PLoS ONE 2016, 11, e0151472. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.; Qin, X. Hydroxypyridinone-Coumarin Inhibits the Proliferation of MHCC97 and HepG2 Human Hepatocellular Carcinoma Cells and Down-Regulates the Phosphoinositide-3 Kinase Pathway. Med. Sci. Monit. 2020, 26, e920785. [Google Scholar] [CrossRef]
- Li, J.; Fu, Y.; Hu, X.; Xiong, Y. Psoralidin inhibits the proliferation of human liver cancer cells by triggering cell cycle arrest, apoptosis and autophagy and inhibits tumor growth in vivo. J. BUON 2019, 24, 1950–1955. [Google Scholar]
- Ren, G.; Luo, W.; Sun, W.; Niu, Y.; Ma, D.L.; Leung, C.H.; Wang, Y.; Lu, J.J.; Chen, X. Psoralidin induced reactive oxygen species (ROS)-dependent DNA damage and protective autophagy mediated by NOX4 in breast cancer cells. Phytomedicine 2016, 23, 939–947. [Google Scholar] [CrossRef]
- De Amicis, F.; Aquila, S.; Morelli, C.; Guido, C.; Santoro, M.; Perrotta, I.; Mauro, L.; Giordano, F.; Nigro, A.; Ando, S.; et al. Bergapten drives autophagy through the up-regulation of PTEN expression in breast cancer cells. Mol. Cancer 2015, 14, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Gao, S.; Yang, Y.; Zhao, X.; Fan, Y.; Ma, W.; Yang, D.; Yang, A.; Yu, Y. Antitumor activity of curcumin by modulation of apoptosis and autophagy in human lung cancer A549 cells through inhibiting PI3K/Akt/mTOR pathway. Oncol. Rep. 2018, 39, 1523–1531. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Gao, S.; Yang, Y.; Zhao, X.; Fan, Y.; Ma, W.; Yang, D.; Yang, A.; Yu, Y. Curcumin induced autophagy anticancer effects on human lung adenocarcinoma cell line A549. Oncol. Lett. 2017, 14, 2775–2782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.; Kim, K.S.; Fukushi, A.; Kim, D.H.; Kim, C.H.; Lee, Y.C. Transcriptional Activation of Human GD3 Synthase (hST8Sia I) Gene in Curcumin-Induced Autophagy in A549 Human Lung Carcinoma Cells. Int. J. Mol. Sci. 2018, 19, 1943. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.; Wang, J.; Zhang, S.; Zhang, H.; Xu, Z.; Li, X. Curcumin inhibits the development of non-small cell lung cancer by inhibiting autophagy and apoptosis. Exp. Ther. Med. 2017, 14, 5075–5080. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Qiao, H.; Wu, D.; Lu, H.; Liu, L.; Sang, X.; Li, D.; Zhou, Y. Curcumin potentiates the galbanic acid-induced anti-tumor effect in non-small cell lung cancer cells through inhibiting Akt/mTOR signaling pathway. Life Sci. 2019, 239, 117044. [Google Scholar] [CrossRef]
- Chen, P.; Huang, H.P.; Wang, Y.; Jin, J.; Long, W.G.; Chen, K.; Zhao, X.H.; Chen, C.G.; Li, J. Curcumin overcome primary gefitinib resistance in non-small-cell lung cancer cells through inducing autophagy-related cell death. J. Exp. Clin. Cancer Res. 2019, 38, 254. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Bu, S. Curcumin Induces Autophagy, Apoptosis, and Cell Cycle Arrest in Human Pancreatic Cancer Cells. Evid. Based Complement. Alternat. Med. 2017, 2017, 5787218. [Google Scholar] [CrossRef]
- Fu, H.; Wang, C.; Yang, D.; Wei, Z.; Xu, J.; Hu, Z.; Zhang, Y.; Wang, W.; Yan, R.; Cai, Q. Curcumin regulates proliferation, autophagy, and apoptosis in gastric cancer cells by affecting PI3K and P53 signaling. J. Cell Physiol. 2018, 233, 4634–4642. [Google Scholar] [CrossRef]
- Li, W.; Zhou, Y.; Yang, J.; Li, H.; Zhang, H.; Zheng, P. Curcumin induces apoptotic cell death and protective autophagy in human gastric cancer cells. Oncol. Rep. 2017, 37, 3459–3466. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.D.; Pang, Y.X.; Zhao, X.R.; Li, R.; Jin, C.J.; Xue, J.; Dong, R.Y.; Liu, P.S. Curcumin induces apoptotic cell death and protective autophagy by inhibiting AKT/mTOR/p70S6K pathway in human ovarian cancer cells. Arch. Gynecol. Obstet. 2019, 299, 1627–1639. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Zhao, B.; Xiong, P.; Wang, C.; Zhang, J.; Tian, X.; Huang, Y. Curcumin Induces Autophagy via Inhibition of Yes-Associated Protein (YAP) in Human Colon Cancer Cells. Med. Sci. Monit. 2018, 24, 7035–7042. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Lai, Z.L.; Chen, H.F.; Zhang, M.; Wang, A.; Jia, T.; Sun, W.Q.; Zhu, X.M.; Chen, X.F.; Zhao, Z.; et al. Curcumin synergizes with 5-fluorouracil by impairing AMPK/ULK1-dependent autophagy, AKT activity and enhancing apoptosis in colon cancer cells with tumor growth inhibition in xenograft mice. J. Exp. Clin. Cancer Res. 2017, 36, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, P.; Ke, C.; Guo, X.; Ren, P.; Tong, Y.; Luo, S.; He, Y.; Wei, Z.; Cheng, B.; Li, R.; et al. Both glypican-3/Wnt/beta-catenin signaling pathway and autophagy contributed to the inhibitory effect of curcumin on hepatocellular carcinoma. Dig. Liver Dis. 2019, 51, 120–126. [Google Scholar] [CrossRef]
- Elmansi, A.M.; El-Karef, A.A.; Shishtawy, M.; Eissa, L.A. Hepatoprotective Effect of Curcumin on Hepatocellular Carcinoma through Autophagic and Apoptic Pathways. Ann. Hepatol. 2017, 16, 607–618. [Google Scholar] [CrossRef]
- Boldbaatar, J.; Gunarta, I.K.; Suzuki, R.; Erdenebaatar, P.; Davaakhuu, G.; Hohjoh, H.; Yoshioka, K. Protective role of c-Jun NH2-terminal kinase-associated leucine zipper protein (JLP) in curcumin-induced cancer cell death. Biochem. Biophys. Res. Commun. 2020, 522, 697–703. [Google Scholar] [CrossRef]
- Deng, Q.; Liang, L.; Liu, Q.; Duan, W.; Jiang, Y.; Zhang, L. Autophagy is a major mechanism for the dual effects of curcumin on renal cell carcinoma cells. Eur. J. Pharmacol. 2018, 826, 24–30. [Google Scholar] [CrossRef]
- Masuelli, L.; Benvenuto, M.; Di Stefano, E.; Mattera, R.; Fantini, M.; De Feudis, G.; De Smaele, E.; Tresoldi, I.; Giganti, M.G.; Modesti, A.; et al. Curcumin blocks autophagy and activates apoptosis of malignant mesothelioma cell lines and increases the survival of mice intraperitoneally transplanted with a malignant mesothelioma cell line. Oncotarget 2017, 8, 34405–34422. [Google Scholar] [CrossRef] [Green Version]
- Masuelli, L.; Granato, M.; Benvenuto, M.; Mattera, R.; Bernardini, R.; Mattei, M.; d’Amati, G.; D’Orazi, G.; Faggioni, A.; Bei, R.; et al. Chloroquine supplementation increases the cytotoxic effect of curcumin against Her2/neu overexpressing breast cancer cells in vitro and in vivo in nude mice while counteracts it in immune competent mice. Oncoimmunology 2017, 6, e1356151. [Google Scholar] [CrossRef] [Green Version]
- Masuelli, L.; Di Stefano, E.; Fantini, M.; Mattera, R.; Benvenuto, M.; Marzocchella, L.; Sacchetti, P.; Focaccetti, C.; Bernardini, R.; Tresoldi, I.; et al. Resveratrol potentiates the in vitro and in vivo anti-tumoral effects of curcumin in head and neck carcinomas. Oncotarget 2014, 5, 10745–10762. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Shan, Q.Q.; Gong, P.Y.; Wang, S.C. The autophagy induced by curcumin via MEK/ERK pathway plays an early anti-leukemia role in human Philadelphia chromosome-positive acute lymphoblastic leukemia SUP-B15 cells. J. Cancer Res. Ther. 2018, 14, S125–S131. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Yoon, S.S.; Moon, E.Y. Curcumin-Induced Autophagy Augments Its Antitumor Effect against A172 Human Glioblastoma Cells. Biomol. Ther. 2019, 27, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.U.; Woo, S.M.; Lee, H.S.; Kim, S.H.; Min, K.J.; Kwon, T.K. mTORC1/2 inhibitor and curcumin induce apoptosis through lysosomal membrane permeabilization-mediated autophagy. Oncogene 2018, 37, 5205–5220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, J.L.; Poklepovic, A.; Booth, L. Curcumin interacts with sildenafil to kill GI tumor cells via endoplasmic reticulum stress and reactive oxygen/nitrogen species. Oncotarget 2017, 8, 99451–99469. [Google Scholar] [CrossRef] [Green Version]
- Maiti, P.; Scott, J.; Sengupta, D.; Al-Gharaibeh, A.; Dunbar, G.L. Curcumin and Solid Lipid Curcumin Particles Induce Autophagy, but Inhibit Mitophagy and the PI3K-Akt/mTOR Pathway in Cultured Glioblastoma Cells. Int. J. Mol. Sci. 2019, 20, 399. [Google Scholar] [CrossRef] [Green Version]
- Khorsandi, K.; Hosseinzadeh, R.; Shahidi, F.K. Photodynamic treatment with anionic nanoclays containing curcumin on human triple-negative breast cancer cells: Cellular and biochemical studies. J. Cell Biochem. 2019, 120, 4998–5009. [Google Scholar] [CrossRef]
- Hsiao, Y.T.; Kuo, C.L.; Chueh, F.S.; Liu, K.C.; Bau, D.T.; Chung, J.G. Curcuminoids Induce Reactive Oxygen Species and Autophagy to Enhance Apoptosis in Human Oral Cancer Cells. Am. J. Chin. Med. 2018, 46, 1145–1168. [Google Scholar] [CrossRef]
- Hsiao, Y.T.; Kuo, C.L.; Lin, J.J.; Huang, W.W.; Peng, S.F.; Chueh, F.S.; Bau, D.T.; Chung, J.G. Curcuminoids combined with gefitinib mediated apoptosis and autophagy of human oral cancer SAS cells in vitro and reduced tumor of SAS cell xenograft mice in vivo. Environ. Toxicol. 2018, 33, 821–832. [Google Scholar] [CrossRef]
- Tseng, Y.H.; Chiou, S.S.; Weng, J.P.; Lin, P.C. Curcumin and tetrahydrocurcumin induce cell death in Ara-C-resistant acute myeloid leukemia. Phytother. Res. 2019, 33, 1199–1207. [Google Scholar] [CrossRef] [Green Version]
- Song, G.; Lu, H.; Chen, F.; Wang, Y.; Fan, W.; Shao, W.; Lu, H.; Lin, B. Tetrahydrocurcumininduced autophagy via suppression of PI3K/Akt/mTOR in nonsmall cell lung carcinoma cells. Mol. Med. Rep. 2018, 17, 5964–5969. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Qiang, P.; Yu, J.; Miao, Y.; Chen, Z.; Qu, J.; Zhao, Q.; Chen, Z.; Liu, Y.; Yao, X.; et al. Identification of compound CA-5f as a novel late-stage autophagy inhibitor with potent anti-tumor effect against non-small cell lung cancer. Autophagy 2019, 15, 391–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, G.Z.; Wang, Q.Q.; Wang, P.B.; Wang, Z.C.; Sun, G.C. One novel curcumin derivative ZYX01 induces autophagy of human non-small lung cancer cells A549 through AMPK/ULK1/Beclin-1 signaling pathway. Cell Mol. Biol. 2019, 65, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.Z.; Shi, Y.Y.; Wei, L.L.; Sun, G.C. Autophagy induction and antiproliferative effect of a novel curcumin derivative MOMI-1 on the human lung cancer cells A549. J. Biochem. Mol. Toxicol. 2019, 33, e22280. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.C.; Hsieh, M.T.; Yang, J.S.; Lu, C.C.; Tsai, F.J.; Tsao, J.W.; Chiu, Y.J.; Kuo, S.C.; Lee, K.H. Effect of bis(hydroxymethyl) alkanoate curcuminoid derivative MTH-3 on cell cycle arrest, apoptotic and autophagic pathway in triple-negative breast adenocarcinoma MDA-MB-231 cells: An in vitro study. Int. J. Oncol. 2018, 52, 67–76. [Google Scholar] [CrossRef]
- Wang, L.; Zhu, Z.; Han, L.; Zhao, L.; Weng, J.; Yang, H.; Wu, S.; Chen, K.; Wu, L.; Chen, T. A curcumin derivative, WZ35, suppresses hepatocellular cancer cell growth via downregulating YAP-mediated autophagy. Food Funct. 2019, 10, 3748–3757. [Google Scholar] [CrossRef]
- Duan, J.; Zhan, J.C.; Wang, G.Z.; Zhao, X.C.; Huang, W.D.; Zhou, G.B. The red wine component ellagic acid induces autophagy and exhibits anti-lung cancer activity in vitro and in vivo. J. Cell Mol. Med. 2019, 23, 143–154. [Google Scholar] [CrossRef]
- Elsaid, F.G.; Alshehri, M.A.; Shati, A.A.; Al-Kahtani, M.A.; Alsheri, A.S.; Massoud, E.E.; El-Kott, A.F.; El-Mekkawy, H.I.; Al-Ramlawy, A.M.; Abdraboh, M.E. The anti-tumourigenic effect of ellagic acid in SKOV-3 ovarian cancer cells entails activation of autophagy mediated by inhibiting Akt and activating AMPK. Clin. Exp. Pharmacol. Physiol. 2020, 47, 1611–1621. [Google Scholar] [CrossRef]
- Wang, S.G.; Huang, M.H.; Li, J.H.; Lai, F.I.; Lee, H.M.; Hsu, Y.N. Punicalagin induces apoptotic and autophagic cell death in human U87MG glioma cells. Acta Pharmacol. Sin. 2013, 34, 1411–1419. [Google Scholar] [CrossRef]
- Guaman-Ortiz, L.M.; Romero-Benavides, J.C.; Suarez, A.I.; Torres-Aguilar, S.; Castillo-Veintimilla, P.; Samaniego-Romero, J.; Ortiz-Diaz, K.; Bailon-Moscoso, N. Cytotoxic Property of Grias neuberthii Extract on Human Colon Cancer Cells: A Crucial Role of Autophagy. Evid. Based Complement. Alternat. Med. 2020, 2020, 1565306. [Google Scholar] [CrossRef] [Green Version]
- Patra, S.; Panda, P.K.; Naik, P.P.; Panigrahi, D.P.; Praharaj, P.P.; Bhol, C.S.; Mahapatra, K.K.; Padhi, P.; Jena, M.; Patil, S.; et al. Terminalia bellirica extract induces anticancer activity through modulation of apoptosis and autophagy in oral squamous cell carcinoma. Food Chem. Toxicol. 2020, 136, 111073. [Google Scholar] [CrossRef]
- Gao, L.; Wang, Z.; Lu, D.; Huang, J.; Liu, J.; Hong, L. Paeonol induces cytoprotective autophagy via blocking the Akt/mTOR pathway in ovarian cancer cells. Cell Death Dis. 2019, 10, 609. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; Chai, Y.; Wang, L.; Zhang, J.; Lee, H.J.; Kim, S.H.; Lu, J. Pentagalloylglucose induces autophagy and caspase-independent programmed deaths in human PC-3 and mouse TRAMP-C2 prostate cancer cells. Mol. Cancer Ther. 2009, 8, 2833–2843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Zhang, G.; Tong, Y.; Yuan, J.; Li, Y.; Song, G. Corilagin induces apoptosis, autophagy and ROS generation in gastric cancer cells in vitro. Int. J. Mol. Med. 2019, 43, 967–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.; Wang, Y.; Yin, X.; Liu, X.; Xuan, H. Ethanol extract of propolis and its constituent caffeic acid phenethyl ester inhibit breast cancer cells proliferation in inflammatory microenvironment by inhibiting TLR4 signal pathway and inducing apoptosis and autophagy. BMC Complement. Altern. Med. 2017, 17, 471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, S.H.; Kao, Y.T.; Wu, J.Y.; Huang, S.H.; Huang, S.T.; Lee, C.M.; Cheng, K.T.; Lin, C.M. Inhibition of AMPK-associated autophagy enhances caffeic acid phenethyl ester-induced cell death in C6 glioma cells. Planta Med. 2011, 77, 907–914. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Kuo, Y.H.; Lin, C.C.; Chao, C.Y.; Pai, M.H.; Chiang, E.I.; Tang, F.Y. Decyl caffeic acid inhibits the proliferation of colorectal cancer cells in an autophagy-dependent manner in vitro and in vivo. PLoS ONE 2020, 15, e0232832. [Google Scholar] [CrossRef]
- Endo, S.; Hoshi, M.; Matsunaga, T.; Inoue, T.; Ichihara, K.; Ikari, A. Autophagy inhibition enhances anticancer efficacy of artepillin C, a cinnamic acid derivative in Brazilian green propolis. Biochem. Biophys. Res. Commun. 2018, 497, 437–443. [Google Scholar] [CrossRef]
- Yu, C.H.; Chu, S.C.; Yang, S.F.; Hsieh, Y.S.; Lee, C.Y.; Chen, P.N. Induction of apoptotic but not autophagic cell death by Cinnamomum cassia extracts on human oral cancer cells. J. Cell Physiol. 2019, 234, 5289–5303. [Google Scholar] [CrossRef]
- Gao, J.; Yu, H.; Guo, W.; Kong, Y.; Gu, L.; Li, Q.; Yang, S.; Zhang, Y.; Wang, Y. The anticancer effects of ferulic acid is associated with induction of cell cycle arrest and autophagy in cervical cancer cells. Cancer Cell Int. 2018, 18, 102. [Google Scholar] [CrossRef] [Green Version]
- Pellerito, C.; Emanuele, S.; Ferrante, F.; Celesia, A.; Giuliano, M.; Fiore, T. Tributyltin(IV) ferulate, a novel synthetic ferulic acid derivative, induces autophagic cell death in colon cancer cells: From chemical synthesis to biochemical effects. J. Inorg. Biochem. 2020, 205, 110999. [Google Scholar] [CrossRef]
- Shailasree, S.; Venkataramana, M.; Niranjana, S.R.; Prakash, H.S. Cytotoxic effect of p-Coumaric acid on neuroblastoma, N2a cell via generation of reactive oxygen species leading to dysfunction of mitochondria inducing apoptosis and autophagy. Mol. Neurobiol. 2015, 51, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Reis, F.S.; Lima, R.T.; Morales, P.; Ferreira, I.C.; Vasconcelos, M.H. Methanolic Extract of Ganoderma lucidum Induces Autophagy of AGS Human Gastric Tumor Cells. Molecules 2015, 20, 17872–17882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaushik, G.; Venugopal, A.; Ramamoorthy, P.; Standing, D.; Subramaniam, D.; Umar, S.; Jensen, R.A.; Anant, S.; Mammen, J.M. Honokiol inhibits melanoma stem cells by targeting notch signaling. Mol. Carcinog. 2015, 54, 1710–1721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaushik, G.; Ramalingam, S.; Subramaniam, D.; Rangarajan, P.; Protti, P.; Rammamoorthy, P.; Anant, S.; Mammen, J.M. Honokiol induces cytotoxic and cytostatic effects in malignant melanoma cancer cells. Am. J. Surg. 2012, 204, 868–873. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Dong, H.; Li, M.; Wu, Y.; Liu, Y.; Zhao, Y.; Chen, X.; Ma, M. Honokiol induces autophagy and apoptosis of osteosarcoma through PI3K/Akt/mTOR signaling pathway. Mol. Med. Rep. 2018, 17, 2719–2723. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.H.; Chen, S.H.; Chang, Y.S.; Liu, Y.W.; Wu, J.Y.; Lim, Y.P.; Yu, H.I.; Lee, Y.R. Honokiol, a potential therapeutic agent, induces cell cycle arrest and program cell death in vitro and in vivo in human thyroid cancer cells. Pharmacol. Res. 2017, 115, 288–298. [Google Scholar] [CrossRef]
- Lin, M.C.; Lee, Y.W.; Tseng, Y.Y.; Lin, Y.W.; Chen, J.T.; Liu, S.H.; Chen, R.M. Honokiol Induces Autophagic Apoptosis in Neuroblastoma Cells through a P53-Dependent Pathway. Am. J. Chin. Med. 2019, 47, 895–912. [Google Scholar] [CrossRef]
- Chio, C.C.; Chen, K.Y.; Chang, C.K.; Chuang, J.Y.; Liu, C.C.; Liu, S.H.; Chen, R.M. Improved effects of honokiol on temozolomide-induced autophagy and apoptosis of drug-sensitive and -tolerant glioma cells. BMC Cancer 2018, 18, 379. [Google Scholar] [CrossRef] [Green Version]
- Hahm, E.R.; Sakao, K.; Singh, S.V. Honokiol activates reactive oxygen species-mediated cytoprotective autophagy in human prostate cancer cells. Prostate 2014, 74, 1209–1221. [Google Scholar] [CrossRef] [Green Version]
- Huang, K.J.; Kuo, C.H.; Chen, S.H.; Lin, C.Y.; Lee, Y.R. Honokiol inhibits in vitro and in vivo growth of oral squamous cell carcinoma through induction of apoptosis, cell cycle arrest and autophagy. J. Cell Mol. Med. 2018, 22, 1894–1908. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.C.; Hueng, D.Y.; Huang, H.Y.; Chen, J.Y.; Chen, Y. Magnolol and honokiol exert a synergistic anti-tumor effect through autophagy and apoptosis in human glioblastomas. Oncotarget 2016, 7, 29116–29130. [Google Scholar] [CrossRef]
- Zhou, J.; Hu, H.; Long, J.; Wan, F.; Li, L.; Zhang, S.; Shi, Y.E.; Chen, Y. Vitexin 6, a novel lignan, induces autophagy and apoptosis by activating the Jun N-terminal kinase pathway. Anticancer Drugs 2013, 24, 928–936. [Google Scholar] [CrossRef] [PubMed]
- Maheswari, U.; Ghosh, K.; Sadras, S.R. Licarin A induces cell death by activation of autophagy and apoptosis in non-small cell lung cancer cells. Apoptosis 2018, 23, 210–225. [Google Scholar] [CrossRef] [PubMed]
- Moura, A.F.; Lima, K.S.B.; Sousa, T.S.; Marinho-Filho, J.D.B.; Pessoa, C.; Silveira, E.R.; Pessoa, O.D.L.; Costa-Lotufo, L.V.; Moraes, M.O.; Araujo, A.J. In vitro antitumor effect of a lignan isolated from Combretum fruticosum, trachelogenin, in HCT-116 human colon cancer cells. Toxicol. In Vitro 2018, 47, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Yin, S.; Zhou, S.; Shao, Y.; Sun, J.; Pang, X.; Han, L.; Zhang, Y.; Gao, X.; Jin, C.; et al. Magnolin promotes autophagy and cell cycle arrest via blocking LIF/Stat3/Mcl-1 axis in human colorectal cancers. Cell Death Dis. 2018, 9, 702. [Google Scholar] [CrossRef] [Green Version]
- Won, S.J.; Yen, C.H.; Liu, H.S.; Wu, S.Y.; Lan, S.H.; Jiang-Shieh, Y.F.; Lin, C.N.; Su, C.L. Justicidin A-induced autophagy flux enhances apoptosis of human colorectal cancer cells via class III PI3K and Atg5 pathway. J. Cell Physiol. 2015, 230, 930–946. [Google Scholar] [CrossRef]
- Ning, Y.; Fu, Y.L.; Zhang, Q.H.; Zhang, C.; Chen, Y. Inhibition of in vitro and in vivo ovarian cancer cell growth by pinoresinol occurs by way of inducing autophagy, inhibition of cell invasion, loss of mitochondrial membrane potential and inhibition Ras/MEK/ERK signalling pathway. J. BUON 2019, 24, 709–714. [Google Scholar]
- Dou, H.; Yang, S.; Hu, Y.; Xu, D.; Liu, L.; Li, X. Sesamin induces ER stress-mediated apoptosis and activates autophagy in cervical cancer cells. Life Sci. 2018, 200, 87–93. [Google Scholar] [CrossRef]
- Tanabe, H.; Kuribayashi, K.; Tsuji, N.; Tanaka, M.; Kobayashi, D.; Watanabe, N. Sesamin induces autophagy in colon cancer cells by reducing tyrosine phosphorylation of EphA1 and EphB2. Int. J. Oncol. 2011, 39, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Li, H.B.; Yi, X.; Gao, J.M.; Ying, X.X.; Guan, H.Q.; Li, J.C. Magnolol-induced H460 cells death via autophagy but not apoptosis. Arch. Pharmacal Res. 2007, 30, 1566–1574. [Google Scholar] [CrossRef]
- Shen, J.; Ma, H.; Zhang, T.; Liu, H.; Yu, L.; Li, G.; Li, H.; Hu, M. Magnolol Inhibits the Growth of Non-Small Cell Lung Cancer via Inhibiting Microtubule Polymerization. Cell Physiol. Biochem. 2017, 42, 1789–1801. [Google Scholar] [CrossRef] [PubMed]
- Rasul, A.; Yu, B.; Khan, M.; Zhang, K.; Iqbal, F.; Ma, T.; Yang, H. Magnolol, a natural compound, induces apoptosis of SGC-7901 human gastric adenocarcinoma cells via the mitochondrial and PI3K/Akt signaling pathways. Int. J. Oncol. 2012, 40, 1153–1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maxwell, T.; Lee, K.S.; Kim, S.; Nam, K.S. Arctigenin inhibits the activation of the mTOR pathway, resulting in autophagic cell death and decreased ER expression in ER-positive human breast cancer cells. Int. J. Oncol. 2018, 52, 1339–1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Lina, L.; Xu, L.; Yang, Z.; Qian, Z.; Zhou, J.; Suoni, L. Arctigenin enhances the sensitivity of cisplatin resistant colorectal cancer cell by activating autophagy. Biochem. Biophys. Res. Commun. 2019, 520, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Okubo, S.; Ohta, T.; Shoyama, Y.; Uto, T. Arctigenin suppresses cell proliferation via autophagy inhibition in hepatocellular carcinoma cells. J. Nat. Med. 2020, 74, 525–532. [Google Scholar] [CrossRef]
- Kwon, J.; Lee, Y.; Jeong, J.H.; Ryu, J.H.; Kim, K.I. Inhibition of autophagy sensitizes lignan-induced endoplasmic reticulum stress-mediated cell death. Biochem. Biophys. Res. Commun. 2020, 526, 300–305. [Google Scholar] [CrossRef]
- Fan, Y.; Chiu, J.F.; Liu, J.; Deng, Y.; Xu, C.; Zhang, J.; Li, G. Resveratrol induces autophagy-dependent apoptosis in HL-60 cells. BMC Cancer 2018, 18, 581. [Google Scholar] [CrossRef] [Green Version]
- Puissant, A.; Robert, G.; Fenouille, N.; Luciano, F.; Cassuto, J.P.; Raynaud, S.; Auberger, P. Resveratrol promotes autophagic cell death in chronic myelogenous leukemia cells via JNK-mediated p62/SQSTM1 expression and AMPK activation. Cancer Res. 2010, 70, 1042–1052. [Google Scholar] [CrossRef] [Green Version]
- Miki, H.; Uehara, N.; Kimura, A.; Sasaki, T.; Yuri, T.; Yoshizawa, K.; Tsubura, A. Resveratrol induces apoptosis via ROS-triggered autophagy in human colon cancer cells. Int. J. Oncol. 2012, 40, 1020–1028. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Fang, Q.; Ji, S.; Han, Z.; Cheng, W.; Zhang, H. Resveratrol-mediated apoptosis in renal cell carcinoma via the p53/AMPactivated protein kinase/mammalian target of rapamycin autophagy signaling pathway. Mol. Med. Rep. 2018, 17, 502–508. [Google Scholar] [CrossRef]
- Zhang, B.; Yin, X.; Sui, S. Resveratrol inhibited the progression of human hepatocellular carcinoma by inducing autophagy via regulating p53 and the phosphoinositide 3kinase/protein kinase B pathway. Oncol. Rep. 2018, 40, 2758–2765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Zepeda, S.P.; Garcia-Villa, E.; Diaz-Chavez, J.; Hernandez-Pando, R.; Gariglio, P. Resveratrol induces cell death in cervical cancer cells through apoptosis and autophagy. Eur. J. Cancer Prev. 2013, 22, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.H.; Lee, C.Y.; Lu, C.C.; Tsai, F.J.; Hsu, Y.M.; Tsao, J.W.; Juan, Y.N.; Chiu, H.Y.; Yang, J.S.; Wang, C.C. Resveratrol-induced autophagy and apoptosis in cisplatin-resistant human oral cancer CAR cells: A key role of AMPK and Akt/mTOR signaling. Int. J. Oncol. 2017, 50, 873–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, M.; Suzuki, S.O.; Himeno, M. Resveratrol-induced autophagy in human U373 glioma cells. Oncol. Lett. 2010, 1, 489–493. [Google Scholar] [CrossRef] [PubMed]
- Filippi-Chiela, E.C.; Villodre, E.S.; Zamin, L.L.; Lenz, G. Autophagy interplay with apoptosis and cell cycle regulation in the growth inhibiting effect of resveratrol in glioma cells. PLoS ONE 2011, 6, e20849. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Qin, Z.; Liang, Z. The prosurvival role of autophagy in Resveratrol-induced cytotoxicity in human U251 glioma cells. BMC Cancer 2009, 9, 215. [Google Scholar] [CrossRef] [Green Version]
- Lang, F.; Qin, Z.; Li, F.; Zhang, H.; Fang, Z.; Hao, E. Apoptotic Cell Death Induced by Resveratrol Is Partially Mediated by the Autophagy Pathway in Human Ovarian Cancer Cells. PLoS ONE 2015, 10, e0129196. [Google Scholar] [CrossRef]
- Wang, H.; Peng, Y.; Wang, J.; Gu, A.; Li, Q.; Mao, D.; Guo, L. Effect of autophagy on the resveratrol-induced apoptosis of ovarian cancer SKOV3 cells. J. Cell Biochem. 2018, 120, 7788–7793. [Google Scholar] [CrossRef]
- Zhong, L.X.; Zhang, Y.; Wu, M.L.; Liu, Y.N.; Zhang, P.; Chen, X.Y.; Kong, Q.Y.; Liu, J.; Li, H. Resveratrol and STAT inhibitor enhance autophagy in ovarian cancer cells. Cell Death Discov. 2016, 2, 15071. [Google Scholar] [CrossRef]
- Tan, L.; Wang, W.; He, G.; Kuick, R.D.; Gossner, G.; Kueck, A.S.; Wahl, H.; Opipari, A.W.; Liu, J.R. Resveratrol inhibits ovarian tumor growth in an in vivo mouse model. Cancer 2016, 122, 722–729. [Google Scholar] [CrossRef]
- Fukuda, T.; Oda, K.; Wada-Hiraike, O.; Sone, K.; Inaba, K.; Ikeda, Y.; Makii, C.; Miyasaka, A.; Kashiyama, T.; Tanikawa, M.; et al. Autophagy inhibition augments resveratrol-induced apoptosis in Ishikawa endometrial cancer cells. Oncol. Lett. 2016, 12, 2560–2566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Ma, K.; Qi, T.; Wei, X.; Zhang, Q.; Li, G.; Chiu, J.F. P62 regulates resveratrol-mediated Fas/Cav-1 complex formation and transition from autophagy to apoptosis. Oncotarget 2015, 6, 789–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Li, J.; Cao, N.; Li, Z.; Han, J.; Li, L. Resveratrol, an activator of SIRT1, induces protective autophagy in non-small-cell lung cancer via inhibiting Akt/mTOR and activating p38-MAPK. Onco Targets Ther. 2018, 11, 7777–7786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; He, W.; Gao, X.; Li, B.; Mei, C.; Xu, R.; Chen, H. Resveratrol overcomes gefitinib resistance by increasing the intracellular gefitinib concentration and triggering apoptosis, autophagy and senescence in PC9/G NSCLC cells. Sci. Rep. 2015, 5, 17730. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.; Li, X.; Xu, R.; Ye, L.; Kong, H.; Zeng, X.; Wang, H.; Xie, W. The synergistic effect of resveratrol in combination with cisplatin on apoptosis via modulating autophagy in A549 cells. Acta Biochim. Biophys. Sin. 2016, 48, 528–535. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.J.; Lee, G.J.; Yi, S.S.; Heo, S.H.; Park, C.R.; Nam, H.S.; Cho, M.K.; Lee, S.H. Cisplatin and resveratrol induce apoptosis and autophagy following oxidative stress in malignant mesothelioma cells. Food Chem. Toxicol. 2016, 97, 96–107. [Google Scholar] [CrossRef]
- Tang, Q.; Li, G.; Wei, X.; Zhang, J.; Chiu, J.F.; Hasenmayer, D.; Zhang, D.; Zhang, H. Resveratrol-induced apoptosis is enhanced by inhibition of autophagy in esophageal squamous cell carcinoma. Cancer Lett. 2013, 336, 325–337. [Google Scholar] [CrossRef]
- Wang, M.; Yu, T.; Zhu, C.; Sun, H.; Qiu, Y.; Zhu, X.; Li, J. Resveratrol triggers protective autophagy through the ceramide/Akt/mTOR pathway in melanoma B16 cells. Nutr. Cancer 2014, 66, 435–440. [Google Scholar] [CrossRef]
- Scarlatti, F.; Maffei, R.; Beau, I.; Codogno, P.; Ghidoni, R. Role of non-canonical Beclin 1-independent autophagy in cell death induced by resveratrol in human breast cancer cells. Cell Death Differ. 2008, 15, 1318–1329. [Google Scholar] [CrossRef]
- Alayev, A.; Berger, S.M.; Kramer, M.Y.; Schwartz, N.S.; Holz, M.K. The combination of rapamycin and resveratrol blocks autophagy and induces apoptosis in breast cancer cells. J. Cell Biochem. 2015, 116, 450–457. [Google Scholar] [CrossRef] [Green Version]
- Rai, G.; Mishra, S.; Suman, S.; Shukla, Y. Resveratrol improves the anticancer effects of doxorubicin in vitro and in vivo models: A mechanistic insight. Phytomedicine 2016, 23, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Rai, G.; Suman, S.; Mishra, S.; Shukla, Y. Evaluation of growth inhibitory response of Resveratrol and Salinomycin combinations against triple negative breast cancer cells. Biomed. Pharmacother. 2017, 89, 1142–1151. [Google Scholar] [CrossRef]
- Back, J.H.; Zhu, Y.; Calabro, A.; Queenan, C.; Kim, A.S.; Arbesman, J.; Kim, A.L. Resveratrol-mediated downregulation of Rictor attenuates autophagic process and suppresses UV-induced skin carcinogenesis. Photochem. Photobiol. 2012, 88, 1165–1172. [Google Scholar] [CrossRef] [PubMed]
- Siedlecka-Kroplewska, K.; Jozwik, A.; Boguslawski, W.; Wozniak, M.; Zauszkiewicz-Pawlak, A.; Spodnik, J.H.; Rychlowski, M.; Kmiec, Z. Pterostilbene induces accumulation of autophagic vacuoles followed by cell death in HL60 human leukemia cells. J. Physiol. Pharmacol. 2013, 64, 545–556. [Google Scholar] [PubMed]
- Wang, Y.; Ding, L.; Wang, X.; Zhang, J.; Han, W.; Feng, L.; Sun, J.; Jin, H.; Wang, X.J. Pterostilbene simultaneously induces apoptosis, cell cycle arrest and cyto-protective autophagy in breast cancer cells. Am. J. Transl. Res. 2012, 4, 44–51. [Google Scholar] [PubMed]
- Chang, H.P.; Lu, C.C.; Chiang, J.H.; Tsai, F.J.; Juan, Y.N.; Tsao, J.W.; Chiu, H.Y.; Yang, J.S. Pterostilbene modulates the suppression of multidrug resistance protein 1 and triggers autophagic and apoptotic mechanisms in cisplatin-resistant human oral cancer CAR cells via AKT signaling. Int. J. Oncol. 2018, 52, 1504–1514. [Google Scholar] [CrossRef]
- Chao, A.C.; Hsu, Y.L.; Liu, C.K.; Kuo, P.L. alpha-Mangostin, a dietary xanthone, induces autophagic cell death by activating the AMP-activated protein kinase pathway in glioblastoma cells. J. Agric. Food Chem. 2011, 59, 2086–2096. [Google Scholar] [CrossRef]
- Wang, F.; Ma, H.; Liu, Z.; Huang, W.; Xu, X.; Zhang, X. alpha-Mangostin inhibits DMBA/TPA-induced skin cancer through inhibiting inflammation and promoting autophagy and apoptosis by regulating PI3K/Akt/mTOR signaling pathway in mice. Biomed. Pharmacother. 2017, 92, 672–680. [Google Scholar] [CrossRef]
- Chen, J.J.; Long, Z.J.; Xu, D.F.; Xiao, R.Z.; Liu, L.L.; Xu, Z.F.; Qiu, S.X.; Lin, D.J.; Liu, Q. Inhibition of autophagy augments the anticancer activity of alpha-mangostin in chronic myeloid leukemia cells. Leuk Lymphoma 2014, 55, 628–638. [Google Scholar] [CrossRef]
- Chen, J.; Zhou, M.; Zhang, Q.; Xu, J.; Ouyang, J. Gambogic acid induces death of K562 cells through autophagy and apoptosis mechanisms. Leuk Lymphoma 2015, 56, 2953–2958. [Google Scholar] [CrossRef]
- Ye, L.; Zhou, J.; Zhao, W.; Jiao, P.; Ren, G.; Wang, S. Gambogic acid-induced autophagy in nonsmall cell lung cancer NCI-H441 cells through a reactive oxygen species pathway. J. Cancer Res. Ther. 2018, 14, S942–S947. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhou, Y.; Cheng, X.; Fan, Y.; He, S.; Li, S.; Ye, H.; Xie, C.; Wu, W.; Li, C.; et al. Isogambogenic acid induces apoptosis-independent autophagic cell death in human non-small-cell lung carcinoma cells. Sci. Rep. 2015, 5, 7697. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Lei, Y.; Yuan, P.; Li, L.; Luo, C.; Gao, R.; Tian, J.; Feng, Z.; Nice, E.C.; Sun, J. ROS-mediated autophagy induced by dysregulation of lipid metabolism plays a protective role in colorectal cancer cells treated with gambogic acid. PLoS ONE 2014, 9, e96418. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.J.; Han, Q.B.; Wen, Z.S.; Ma, L.; Gao, J.; Zhou, G.B. Gambogenic acid induces G1 arrest via GSK3beta-dependent cyclin D1 degradation and triggers autophagy in lung cancer cells. Cancer Lett. 2012, 322, 185–194. [Google Scholar] [CrossRef]
- Mei, W.; Dong, C.; Hui, C.; Bin, L.; Fenggen, Y.; Jingjing, S.; Cheng, P.; Meiling, S.; Yawen, H.; Xiaoshan, W.; et al. Gambogenic acid kills lung cancer cells through aberrant autophagy. PLoS ONE 2014, 9, e83604. [Google Scholar] [CrossRef]
- Kim, M.O.; Lee, H.S.; Chin, Y.W.; Moon, D.O.; Ahn, J.S. Gartanin induces autophagy through JNK activation which extenuates caspase-dependent apoptosis. Oncol. Rep. 2015, 34, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Antalek, M.; Nguyen, L.; Li, X.; Tian, X.; Le, A.; Zi, X. The effect of gartanin, a naturally occurring xanthone in mangosteen juice, on the mTOR pathway, autophagy, apoptosis, and the growth of human urinary bladder cancer cell lines. Nutr. Cancer 2013, 65 (Suppl. 1), 68–77. [Google Scholar] [CrossRef] [Green Version]
- Luo, M.; Liu, Q.; He, M.; Yu, Z.; Pi, R.; Li, M.; Yang, X.; Wang, S.; Liu, A. Gartanin induces cell cycle arrest and autophagy and suppresses migration involving PI3K/Akt/mTOR and MAPK signalling pathway in human glioma cells. J. Cell Mol. Med. 2017, 21, 46–57. [Google Scholar] [CrossRef]
- Pham, V.; Rendon, R.; Le, V.X.; Tippin, M.; Fu, D.J.; Le, T.H.; Miller, M.; Agredano, E.; Cedano, J.; Zi, X. Gartanin is a novel NEDDylation inhibitor for induction of Skp2 degradation, FBXW2 expression, and autophagy. Mol. Carcinog. 2020, 59, 193–201. [Google Scholar] [CrossRef]
- Kaewpiboon, C.; Boonnak, N.; Kaowinn, S.; Chung, Y.H. Formoxanthone C, isolated from Cratoxylum formosum ssp. pruniflorum, reverses anticancer drug resistance by inducing both apoptosis and autophagy in human A549 lung cancer cells. Bioorg. Med. Chem. Lett. 2018, 28, 820–825. [Google Scholar] [CrossRef]
- Yu, L.; Chen, M.; Zhang, R.; Jin, Z. Inhibition of cancer cell growth in gemcitabine-resistant pancreatic carcinoma by mangiferin phytochemical involves induction of autophagy, endogenous ROS production, cell cycle disruption, mitochondrial mediated apoptosis and suppression of cancer cell migration and invasion. J. BUON 2019, 24, 1581–1586. [Google Scholar] [PubMed]
- Tovilovic-Kovacevic, G.; Krstic-Milosevic, D.; Vinterhalter, B.; Toljic, M.; Perovic, V.; Trajkovic, V.; Harhaji-Trajkovic, L.; Zogovic, N. Xanthone-rich extract from Gentiana dinarica transformed roots and its active component norswertianin induce autophagy and ROS-dependent differentiation of human glioblastoma cell line. Phytomedicine 2018, 47, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; Zhang, H.; Xu, D.; Lao, Y.; Fu, W.; Tan, H.; Cao, P.; Yang, L.; Xu, H. Xanthones from the Leaves of Garcinia cowa Induce Cell Cycle Arrest, Apoptosis, and Autophagy in Cancer Cells. Molecules 2015, 20, 11387–11399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, S.B.; Kang, H.M.; Park, D.B.; Park, B.S.; Kim, I.R. Cudraxanthone D Regulates Epithelial-Mesenchymal Transition by Autophagy Inhibition in Oral Squamous Cell Carcinoma Cell Lines. Evid. Based Complement. Alternat. Med. 2019, 2019, 5213028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyes-Farias, M.; Carrasco-Pozo, C. The Anti-Cancer Effect of Quercetin: Molecular Implications in Cancer Metabolism. Int. J. Mol. Sci. 2019, 20, 3177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, M.A.; Khathayer, F.; Ray, S.K. Quercetin and Sodium Butyrate Synergistically Increase Apoptosis in Rat C6 and Human T98G Glioblastoma Cells through Inhibition of Autophagy. Neurochem. Res. 2019, 44, 1715–1725. [Google Scholar] [CrossRef]
- Tomas-Hernandez, S.; Blanco, J.; Rojas, C.; Roca-Martinez, J.; Ojeda-Montes, M.J.; Beltran-Debon, R.; Garcia-Vallve, S.; Pujadas, G.; Arola, L.; Mulero, M. Resveratrol Potently Counteracts Quercetin Starvation-Induced Autophagy and Sensitizes HepG2 Cancer Cells to Apoptosis. Mol. Nutr. Food Res. 2018, 62, 11700610. [Google Scholar] [CrossRef]
- Boukes, G.J.; van de Venter, M. The apoptotic and autophagic properties of two natural occurring prodrugs, hyperoside and hypoxoside, against pancreatic cancer cell lines. Biomed. Pharmacother. 2016, 83, 617–626. [Google Scholar] [CrossRef]
- Chen, J.; Zhou, J.; Li, F.; Zhu, Y.; Zhang, W.; Yu, X. Delphinidin induces autophagy in HER-2+ breast cancer cells via inhibition of AKT/mTOR pathway. Zhong Nan Da Xue Xue Bao Yi Xue Ban 2017, 42, 264–270. [Google Scholar] [CrossRef]
- Wu, S.; Hu, Y.; Bai, W.; Zhao, J.; Huang, C.; Wen, C.; Deng, L.; Lu, D. Cyanidin-3-O-glucoside inhibits UVA-induced human dermal fibroblast injury by upregulating autophagy. Photodermatol. Photoimmunol. Photomed. 2019, 35, 360–368. [Google Scholar] [CrossRef]
- Noda, Y.; Kaneyuki, T.; Mori, A.; Packer, L. Antioxidant activities of pomegranate fruit extract and its anthocyanidins: Delphinidin, cyanidin, and pelargonidin. J. Agric. Food Chem. 2002, 50, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.A.; Ahn, K.S.; Song, Y.W.; Moon, J.Y.; Cho, M.; Lim, Y.; Cho, S.K. Mechanism of 2′,3′-dimethoxyflavanone-induced apoptosis in breast cancer stem cells: Role of ubiquitination of caspase-8 and LC3. Arch. Biochem. Biophys. 2014, 562, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Pons, D.G.; Nadal-Serrano, M.; Torrens-Mas, M.; Oliver, J.; Roca, P. The Phytoestrogen Genistein Affects Breast Cancer Cells Treatment Depending on the ERalpha/ERbeta Ratio. J. Cell Biochem. 2016, 117, 218–229. [Google Scholar] [CrossRef]
- Ahmad, B.; Khan, S.; Liu, Y.; Xue, M.; Nabi, G.; Kumar, S.; Alshwmi, M.; Qarluq, A.W. Molecular Mechanisms of Anticancer Activities of Puerarin. Cancer Manag. Res. 2020, 12, 79–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, H.H.; Huang, C.Y.; Chou, C.W.; Makondi, P.T.; Huang, M.T.; Wei, P.L.; Chang, Y.J. Heat shock protein 27 influences the anti-cancer effect of curcumin in colon cancer cells through ROS production and autophagy activation. Life Sci. 2018, 209, 43–51. [Google Scholar] [CrossRef]
- Chung, Y.C.; Lu, L.C.; Tsai, M.H.; Chen, Y.J.; Chen, Y.Y.; Yao, S.P.; Hsu, C.P. The inhibitory effect of ellagic Acid on cell growth of ovarian carcinoma cells. Evid. Based Complement. Alternat. Med. 2013, 2013, 306705. [Google Scholar] [CrossRef]
- Elshaer, M.; Chen, Y.; Wang, X.J.; Tang, X. Resveratrol: An overview of its anti-cancer mechanisms. Life Sci. 2018, 207, 340–349. [Google Scholar] [CrossRef]
- Zhao, Y.; Tang, H.; Zeng, X.; Ye, D.; Liu, J. Resveratrol inhibits proliferation, migration and invasion via Akt and ERK1/2 signaling pathways in renal cell carcinoma cells. Biomed. Pharmacother. 2018, 98, 36–44. [Google Scholar] [CrossRef]
- Ferraresi, A.; Phadngam, S.; Morani, F.; Galetto, A.; Alabiso, O.; Chiorino, G.; Isidoro, C. Resveratrol inhibits IL-6-induced ovarian cancer cell migration through epigenetic up-regulation of autophagy. Mol. Carcinog. 2017, 56, 1164–1181. [Google Scholar] [CrossRef]
- Madeo, F.; Carmona-Gutierrez, D.; Hofer, S.J.; Kroemer, G. Caloric Restriction Mimetics against Age-Associated Disease: Targets, Mechanisms, and Therapeutic Potential. Cell Metab. 2019, 29, 592–610. [Google Scholar] [CrossRef] [Green Version]
- Kumar, B.; Iqbal, M.A.; Singh, R.K.; Bamezai, R.N. Resveratrol inhibits TIGAR to promote ROS induced apoptosis and autophagy. Biochimie 2015, 118, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, V.; Srivastava, P.; Yadav, N.; Amadori, M.; Schneider, A.; Seshadri, A.; Pitarresi, J.; Scott, R.; Zhang, H.; Koochekpour, S.; et al. Resveratrol depletes mitochondrial DNA and inhibition of autophagy enhances resveratrol-induced caspase activation. Mitochondrion 2013, 13, 493–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, D.; Jeong, H.; Lee, M.N.; Koh, A.; Kwon, O.; Yang, Y.R.; Noh, J.; Suh, P.G.; Park, H.; Ryu, S.H. Resveratrol induces autophagy by directly inhibiting mTOR through ATP competition. Sci. Rep. 2016, 6, 21772. [Google Scholar] [CrossRef] [Green Version]
- Modak, M.; Dixit, P.; Londhe, J.; Ghaskadbi, S.; Devasagayam, T.P. Indian herbs and herbal drugs used for the treatment of diabetes. J. Clin. Biochem. Nutr. 2007, 40, 163–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, H.Y.; Ho, C.T.; Chen, Y.K. Biological actions and molecular effects of resveratrol, pterostilbene, and 3′-hydroxypterostilbene. J. Food. Drug Anal. 2017, 25, 134–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, K.; Liu, X.; Chen, L.; Lv, J.M.; Qu, F.J.; Pan, X.W.; Li, L.; Cui, X.G.; Gao, Y.; Xu, D.F. alpha-Viniferin activates autophagic apoptosis and cell death by reducing glucocorticoid receptor expression in castration-resistant prostate cancer cells. Med. Oncol. 2018, 35, 105. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Sun, X.; Yang, Y.; Yang, X.; Zhu, H.; Dai, S.; Chen, X.; Zhang, H.; Guo, Q.; Song, Y.; et al. Gambogic acid enhances the radiosensitivity of human esophageal cancer cells by inducing reactive oxygen species via targeting Akt/mTOR pathway. Tumor Biol. 2016, 37, 1853–1862. [Google Scholar] [CrossRef]
- Kimmelman, A.C. The dynamic nature of autophagy in cancer. Genes Dev. 2011, 25, 1999–2010. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Chen, X.; Wang, X.; Zhao, Z.; Hu, W.; Zeng, S.; Wei, J.; Yang, X.; Qian, L.; Zhou, S.; et al. The effects and the mechanisms of autophagy on the cancer-associated fibroblasts in cancer. J. Exp. Clin. Cancer Res. 2019, 38, 171. [Google Scholar] [CrossRef] [Green Version]
- Thuwajit, C.; Ferraresi, A.; Titone, R.; Thuwajit, P.; Isidoro, C. The metabolic cross-talk between epithelial cancer cells and stromal fibroblasts in ovarian cancer progression: Autophagy plays a role. Med. Res. Rev. 2018, 38, 1235–1254. [Google Scholar] [CrossRef]
- Thongchot, S.; Ferraresi, A.; Vidoni, C.; Loilome, W.; Yongvanit, P.; Namwat, N.; Isidoro, C. Erratum to “Resveratrol interrupts the pro-invasive communication between Cancer associated Fibroblasts and Cholangiocarcinoma cells” [Cancer Letters 430C (2018) 160-171]. Cancer Lett. 2018, 434, 206–207. [Google Scholar] [CrossRef]
- Huang, D.; Cui, L.; Ahmed, S.; Zainab, F.; Wu, Q.; Wang, X.; Yuan, Z. An overview of epigenetic agents and natural nutrition products targeting DNA methyltransferase, histone deacetylases and microRNAs. Food Chem. Toxicol. 2019, 123, 574–594. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, M.F.; Saleem, A.; Rasul, A.; Faran Ashraf Baig, M.M.; Bin-Jumah, M.; Abdel Daim, M.M. Anticancer natural medicines: An overview of cell signaling and other targets of anticancer phytochemicals. Eur. J. Pharmacol. 2020, 888, 173488. [Google Scholar] [CrossRef] [PubMed]
- Vidoni, C.; Ferraresi, A.; Secomandi, E.; Vallino, L.; Dhanasekaran, D.N.; Isidoro, C. Epigenetic targeting of autophagy for cancer prevention and treatment by natural compounds. Semin. Cancer Biol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Patra, S.; Mishra, S.R.; Behera, B.P.; Mahapatra, K.K.; Panigrahi, D.P.; Bhol, C.S.; Praharaj, P.P.; Sethi, G.; Patra, S.K.; Bhutia, S.K. Autophagy-modulating phytochemicals in cancer therapeutics: Current evidences and future perspectives. Semin. Cancer Biol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Davatgaran-Taghipour, Y.; Masoomzadeh, S.; Farzaei, M.H.; Bahramsoltani, R.; Karimi-Soureh, Z.; Rahimi, R.; Abdollahi, M. Polyphenol nanoformulations for cancer therapy: Experimental evidence and clinical perspective. Int. J. Nanomed. 2017, 12, 2689–2702. [Google Scholar] [CrossRef] [Green Version]
- Yadav, N.; Parveen, S.; Banerjee, M. Potential of nano-phytochemicals in cervical cancer therapy. Clin. Chim. Acta. 2020, 505, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.N.; Almoyad, M.; Huq, F. Polyphenols in Colorectal Cancer: Current State of Knowledge including Clinical Trials and Molecular Mechanism of Action. BioMed Res. Int. 2018, 2018, 4154185. [Google Scholar] [CrossRef] [Green Version]
- Avila-Galvez, M.A.; Gimenez-Bastida, J.A.; Espin, J.C.; Gonzalez-Sarrias, A. Dietary Phenolics against Breast Cancer. A Critical Evidence-Based Review and Future Perspectives. Int. J. Mol. Sci. 2020, 21, 5718. [Google Scholar] [CrossRef]
- Bisol, A.; de Campos, P.S.; Lamers, M.L. Flavonoids as anticancer therapies: A systematic review of clinical trials. Phytother. Res. 2020, 34, 568–582. [Google Scholar] [CrossRef] [PubMed]
- Kanai, M.; Imaizumi, A.; Otsuka, Y.; Sasaki, H.; Hashiguchi, M.; Tsujiko, K.; Matsumoto, S.; Ishiguro, H.; Chiba, T. Dose-escalation and pharmacokinetic study of nanoparticle curcumin, a potential anticancer agent with improved bioavailability, in healthy human volunteers. Cancer Chemother. Pharmacol. 2012, 69, 65–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livingstone, T.L.; Beasy, G.; Mills, R.D.; Plumb, J.; Needs, P.W.; Mithen, R.; Traka, M.H. Plant Bioactives and the Prevention of Prostate Cancer: Evidence from Human Studies. Nutrients 2019, 11, 2245. [Google Scholar] [CrossRef] [Green Version]
- Miller, K.; Feucht, W.; Schmid, M. Bioactive Compounds of Strawberry and Blueberry and Their Potential Health Effects Based on Human Intervention Studies: A Brief Overview. Nutrients 2019, 11, 1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Chen, W.; Tu, G.; Chen, X.; Lu, Y.; Wu, L.; Zheng, D. Enhanced Chemotherapeutic Efficacy of PLGA-Encapsulated Epigallocatechin Gallate (EGCG) Against Human Lung Cancer. Int. J. Nanomed. 2020, 15, 4417–4429. [Google Scholar] [CrossRef]




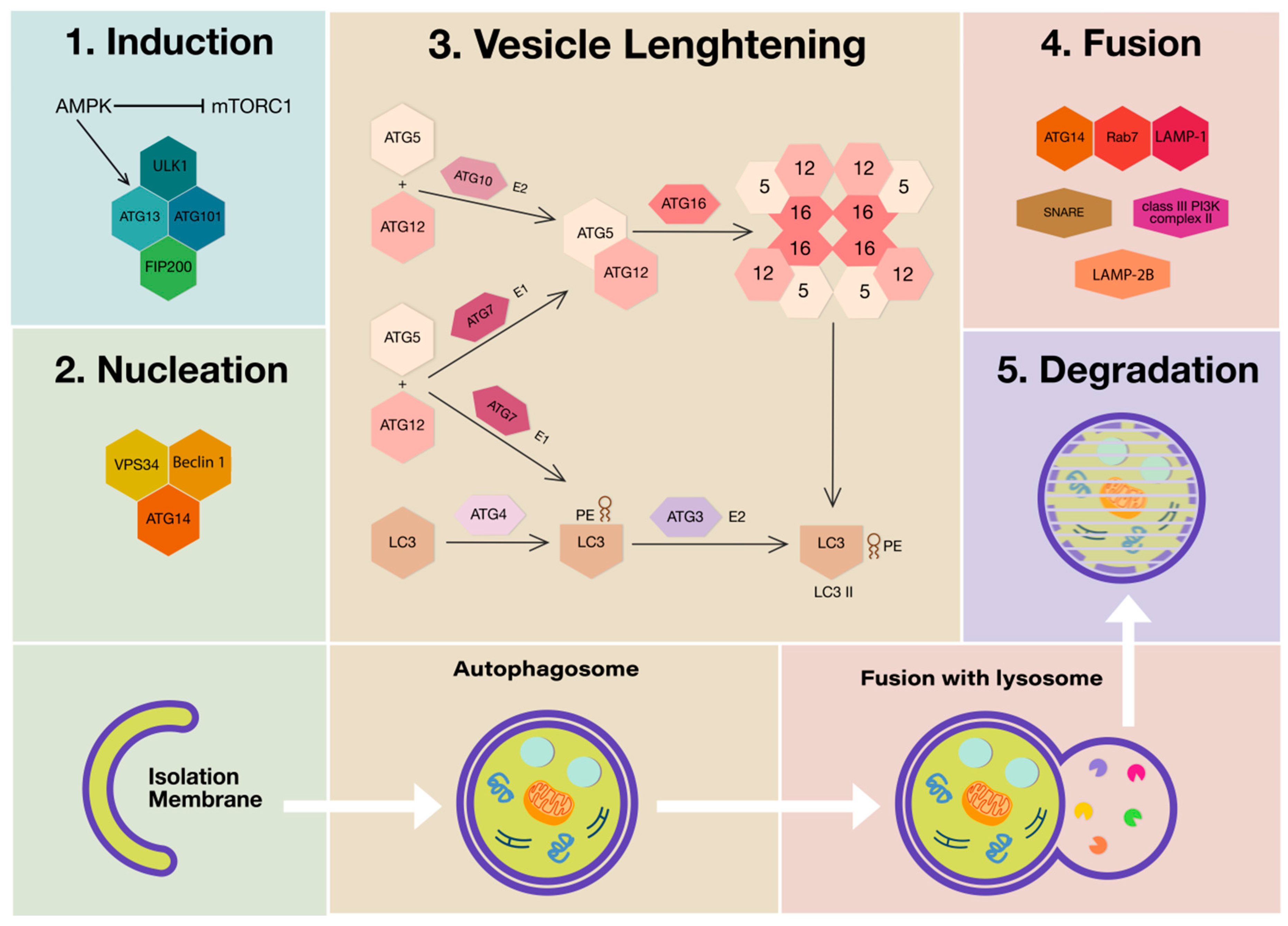{kind=link}
{kind=link}
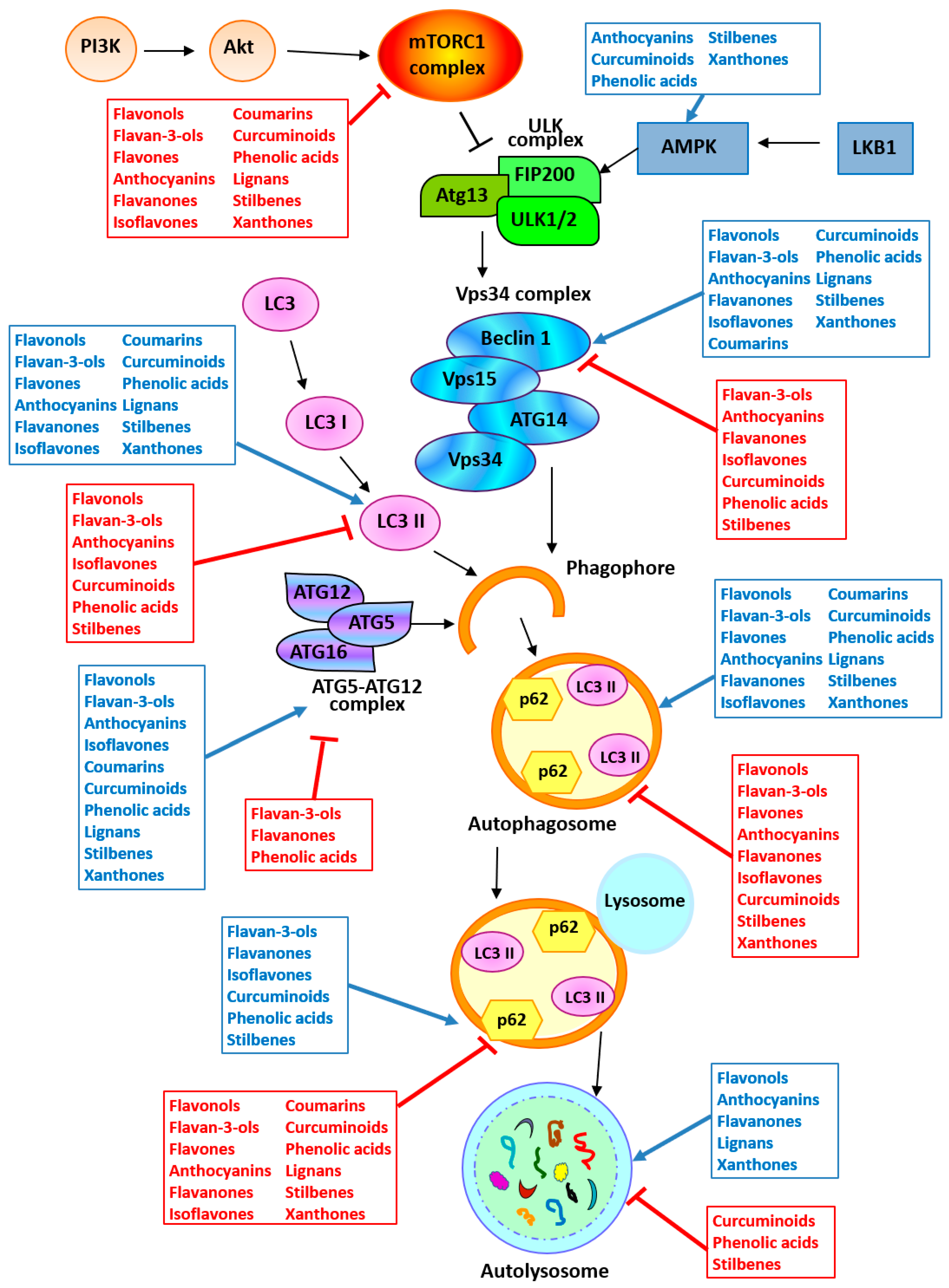{kind=link}
| Polyphenol | In Vitro Model | In Vivo Model | Effects on Autophagy | Reference |
|---|---|---|---|---|
| Flavonoids | ||||
| Flavonols | ||||
| Quercetin | MCF-7 breast cancer cells and HeLa cervical cancer cells (90 µM) | Induction of autophagy ↑ Autophagosomes and autolysosomes ↓ mTOR pathway | [58] | |
| Burkitt lymphoma cells (100 µM) | Induction of autophagy ↑ LC3 II expression ↓ PI3K/Akt/mTOR pathway | [59] | ||
| MCF-7 and MDA-MB-231 breast cancer cells (30 µM) | BALB/c nude mice s.c. injected with MCF-7 cells (50 mg/kg, i.p., twice daily) | Induction of autophagy ↑ LC3 II/I ratio ↑ LC3-puncta structures ↓ Akt/mTOR pathway | [60] | |
| A549 NSCLC cells (20–80 µM + TRAIL) | Induction of autophagy ↑ LC3 II expression ↑ Autophagosomes ↓ Akt/mTOR pathway ↑ TRAIL-induced cell death | [61] | ||
| NOD/SCID mice s.c. injected with HL-60 AML cells (Quercetin 120 mg/kg, i.p., every 4 days; green tea: 100 mg/kg, p.o., daily) | Induction of autophagy ↑ Beclin 1, ATG7, ATG5-ATG12 expression ↑ LC3 II positive cells | [62] | ||
| MOGGCCM anaplastic astrocytoma and T98G glioblastoma multiforme (30–50 µM + Sorafenib) | Induction of autophagy ↑ AVOs ↑ Beclin 1 and LC3 II expression | [63] | ||
| U373MG malignant glioma cells (25–100 µM) | Induction of cytoprotective autophagy ↑ AVOs ↑ LC3 I to LC3 II conversion | [64] | ||
| U87 and U251 glioma cells (25–100 µM) | Sprague Dawley rats intracranially injected with rat glioma C6 cells (100 mg/kg, i.v., daily) | Induction of cytoprotective autophagy ↑ Autophagosomes ↑ LC3 II expression | [65] | |
| AGS and MKN28 gastric cancer cells (AGS: 10–40 µM; MKN28: 40–160 µM) | BALB/c mice s.c. injected with MKN28 cells (50 mg/kg, i.p., daily) | Induction of cytoprotective autophagy ↑ Autophagosomes ↑ AVOs ↑ LC3 I to LC3 II conversion ↑ Beclin 1, ATG7, ATG5-ATG12 expression ↓ Akt/mTOR pathway | [66] | |
| BC3 and BCBL1 primary effusion lymphoma cells (50 µM) | Induction of cytoprotective autophagy ↑ LC3-puncta structures ↑ LC3 II expression ↓ PI3K/Akt/mTOR pathway | [67] | ||
| CAOV-3 and primary P#1 ovarian cancer cells (40–80 µM) | NOD/SCID mice i.p. injected with CAOV-3 cells (80 mg/kg, i.p., twice a week) | Induction of cytoprotective autophagy ↑ Autophagosomes ↑ LC3-puncta structures ↑ LC3 I to LC3 II conversion ↑ Beclin 1 and ATG5 expression | [68] | |
| P39 leukemia cells (50 µM) | Induction of cytoprotective autophagy | [69] | ||
| HL-60 AML cells (100 µM) | Induction of cytoprotective autophagy | [70] | ||
| HeLa cervical cancer cells (50 µM) | Induction of cytoprotective autophagy | [71] | ||
| CHNQ | HCT-116 and HT-29 colon cancer cells (HCT-116: 25 µM) (HT-29: 40 µM) | Induction of autophagy ↑ LC3 I to LC3 II conversion ↑ LC3-puncta structures ↑ AVOs ↓ Akt phosphorylation | [72] | |
| 8-CEPQ | SW620 and HCT-116 colon cancer cells (15 µM) | Induction of autophagy ↑ AVOs ↑ LC3-puncta structures ↑ LC3 II, Beclin 1 and ATG7 expression ↓ p62 expression ↑ ERK pathway | [73] | |
| GQ | A549 and NCI-H1975 NSCLC cells (25–35 µM) | Induction of autophagy ↑ Autophagosomes ↑ LC3 II and Beclin 1 expression ↓ p62 expression | [74] | |
| Hyperoside | A549 NSCLC cells (0.5–2 mM) | Induction of autophagy ↑ LC3 II expression ↑ Autophagosomes ↓ p62 expression ↓ Akt/mTOR/p70S6K pathway ↑ ERK pathway | [75] | |
| SKOV-3 and HO-8910 ovarian cancer cells (50–100 µM) | Induction of autophagy ↑ MDC-labelled vacuoles ↑ LC3 II expression | [76] | ||
| Isorhamnetin | A549 NSCLC cells (2–8 µM) | BALB/c nu/nu mice s.c. injected with A549 cells (0.5 mg/kg, i.p., daily) | Induction of cytoprotective autophagy ↑ LC3 II and Beclin 1 expression ↑ LC3-puncta structures ↑ MDC-labelled vacuoles | [77] |
| Rutin | D54MG glioblastoma multiforme cells (50–200 µM + TMZ) | BALB/c athymic mice s.c. injected with U87 glioma cells or intracranially injected with U87 cells (20 mg/kg, i.p., daily) | Inhibition TMZ-induced autophagy ↓ LC3 II expression ↓ JNK activity ↑ Drug-induced cytotoxicity | [78] |
| Taxifolin | HeLa cervical cancer cells (100 µM + Andrographolide) | Inhibition andrographolide-induced autophagy ↓ LC3-puncta structures ↓ AVOs ↑ Drug-induced cytotoxicity | [79] | |
| Quercetin nanoparticles (NPs) | U87 glioma cells (30–40 µg/mL) | BALB/c nu/nu nude mice s.c. injected with U87 cells (40 and 80 mg/kg, i.p., daily) | Induction of autophagy ↑ LC3 I to LC3 II conversion ↑ Beclin 1 expression ↓ p62 expression ↓ Akt/mTOR pathway | [80] |
| Caski cervical cancer cells (10–20 µg/mL) | BALB/c nu/nu nude mice s.c. injected with Caski cells treated with NPs | Induction of autophagy ↑ Autophagosomes ↓ Akt/mTOR pathway | [81] | |
| Kaempferol | SK-Hep-1 HCC cells (50–100 µM) | Induction of autophagy ↑ Autophagosomes ↑ AVOs ↑ LC3-puncta structures ↑ LC3 II, ATG5, ATG7, ATG12 and Beclin 1 expression ↓ Akt/mTOR pathway | [82] | |
| A549 NSCLC cells (20–50 µM) | Induction of autophagy ↑ LC3 II/I ratio ↑ ATG7 and Beclin 1 expression ↓ p62 expression ↑ LC3-puncta structures ↓ PI3K/Akt pathway | [83] | ||
| SNU-216 gastric cancer cells (50 µM) | Induction of autophagy ↑ LC3 II/I ratio ↑ ATG7 and Beclin 1 expression ↓ p62 expression | [84] | ||
| AGS and SNU-638 gastric cancer cells (50 µM) | Induction of autophagy ↑ LC3 I to LC3 II conversion ↑ LC3 II, Beclin 1 and ATG5 expression ↓ p62 expression ↑ LC3-puncta structures | [85] | ||
| Kaempferol or CAPE | RKO and HCT-116 colon cancer cells (RKO: CAPE 36.87 µM, kaempferol 17.42 µM) (HCT-116: CAPE 3.326 µM, kaempferol 9.427 µM) | Induction of autophagy ↑ MDC-labelled vacuoles | [86] | |
| Juglanin | MCF-7 and SK-BR-3 breast cancer cells (2.5–10 µM) | BALB/c-nude mice s.c. injected with MCF-7 cells (5 and 10 mg/kg, i.p., daily) | Induction of autophagy ↑ Autophagosomes ↑ LC3 II expression | [87] |
| Kazinol A | T24 and cisplatin-resistant T24R2 bladder cancer cells (20 µM) | Induction of autophagy ↑ LC3-puncta structures ↑ LC3 I to LC3 II conversion ↓ mTOR phosphorylation | [88] | |
| Dihydromyricetin | A431 epidermoid carcinoma cells (25–100 µM) | Induction of autophagy ↑ Beclin 1 and LC3 II expression | [89] | |
| HepG2 HCC cells (10–50 µM) | Induction of autophagy ↑ Beclin 1 and LC3 II expression | [90] | ||
| SK-MEL-28 human melanoma cells (100 μM) | Induction of cytoprotective autophagy ↑ Beclin 1 and LC3 II expression ↓ p62 expression Activation NF-κB pathway | [91] | ||
| CAL-27 OSCC cells (50 μM) | Induction of cytoprotective autophagy ↑ Beclin 1 and LC3 II expression ↓ p62 expression | [92] | ||
| Flavan-3-ols | ||||
| HLP extract and ECG | A375 melanoma cells (HLP: 100–250 μg/mL; ECG: 100 µM) | Induction of autophagy ↑ AVOs ↑ LC3 II, Beclin 1, ATG9, ATG16, ATG5-ATG12 expression ↓ Akt/mTOR pathway | [93] | |
| EGCG | SSC-4 OSCC cells (20 µM) | Induction of autophagy ↑ MDC-labelled vacuoles | [94] | |
| 4T1 breast cancer cells (10, 20, 40 µM) | BALB/c mice s.c. injected with 4T1 cells (5, 10, 20 mg/kg, i.p., daily) | Induction of autophagy ↑ LC3 II/I ratio, Beclin 1, ATG5 expression ↑ LC3-puncta structures ↑ Autophagosomes | [95] | |
| HepG2 HCC cells (25–50 µM) | Induction of autophagy ↑ MDC-labelled vacuoles ↑ LC3-puncta structures ↑ LC3 II expression Degradation of AFP aggregates | [96] | ||
| HCT-116 colorectal cancer cells (12.5 µM + 2 Gy radiation) | Induction of autophagy ↑ LC3 II mRNA expression ↑ Sensitivity to radiation | [97] | ||
| HepG2 HCC cells PANC-1 pancreatic cancer cells (20 µM + PEF + US) | Induction of autophagy ↑ MDC-labelled vacuoles ↑ LC3 II expression ↓ Akt phosphorylation | [98] | ||
| GBM15, GBM16 primary glioblastoma cells (500 µM) | Induction of autophagy ↑ MDC-labelled vacuoles ↑ LC3 II expression | [99] | ||
| EHMES-10, ACC-meso and Y-meso malignant mesothelioma cells (50–250 µM) | Induction of cytoprotective autophagy ↑ LC3 II expression | [100] | ||
| JP8 | B16-F10 melanoma cells (20 µM) | C57BL/6 mice s.c. injected with B16-F10 cells (25 and 50 mg/kg, i.p., daily) | Induction of autophagy ↑ LC3 I to LC3 II conversion ↑ LC3-puncta structures ↓ p62 expression | [101] |
| Green tea extract | A549 NSCLC cells (25, 50, 150 µM) | Induction of cytoprotective autophagy ↑ Autophagosomes and autolysosomes ↑ AVOs ↑ LC3-puncta structures | [102] | |
| Polyphenon E® | PNT1a prostate cancer cells (35 µg/mL) | Induction of cytoprotective autophagy ↑ LC3-puncta structures ↑ LC3 I to LC3 II conversion | [103] | |
| Tea polyphenols + anticancer drugs | T24 cells and BIU87 bladder cancer cells (100 µM + Epirubicin) | Inhibition epirubicin-induced autophagy ↓ LC3 II induction ↓ Autophagosomes and autolysosomes ↑ Sensitivity to epirubicin | [104] | |
| PC-3 and DU145 castration-resistant prostate cancer cells (20 µM + Docetaxel) | Inhibition docetaxel-induced autophagy ↓ LC3 II expression ↑ mTOR activation ↑ Sensitivity to docetaxel | [105] | ||
| EGCG + anticancer drugs | Hep3B hepatoma cells (10–40 µg/mL + DOX) | Nude mice s.c. injected with Hep3B cells (50 mg/kg, intragastric, daily + DOX) | Inhibition DOX-induced autophagy ↓ Autophagosomes ↓ ATG5, Beclin 1 mRNA and protein ↑ Sensitivity to DOX | [106] |
| SaoS2 and U2OS cells osteosarcoma cells (20 µg/mL + DOX) | Inhibition DOX-induced autophagy ↓ LC3-puncta structures ↓ ATG5, Beclin 1 mRNA ↓ LC3 II/I ratio ↑ p62 expression ↑ Sensitivity to DOX | [107] | ||
| A549 NSCLC cells (34 µM + Gefitinib) | Inhibition gefitinib-induced autophagy ↓ LC3-puncta structures ↓ LC3 II/I ratio, ATG5 expression ↑ p62 expression ↑ Sensitivity to gefitinib | [108] | ||
| DLD-1 and HT-29 colon cancer cells (100 µM + Cisplatin/Oxaliplatin) | ↑ Drugs-induced autophagy ↑ LC3 I to LC3 II conversion ↑ LC3 II expression ↑ Autophagosomes ↑ AVOs | [109] | ||
| Cisplatin-resistant CAR oral cancer cells (50 µM) | Induction of autophagy ↑ AVOs ↑ Autophagosomes ↑ LC3-puncta structures ↑ ATG5, ATG7, ATG12, Beclin 1 and LC3 II expression ↓ Akt/STAT3 pathway | [110] | ||
| PC-3 prostate cancer cells (5, 50 µM + Bortezomib) | Induction of autophagy ↑ LC3-puncta structures ↑ LC3 I to LC3 II conversion Antagonized drug-induced cytotoxic effect | [111] | ||
| HCT-116 colorectal cancer cells (5–20 µM + TRAIL) | Induction of autophagy ↓ p62 expression and accumulation Antagonized drug-induced cytotoxic effect | [112] | ||
| Flavones | ||||
| Isocannflavin B | Estrogens sensitive ER+ T47-D and insensitive ER− MDA-MB-231 cells (25 µM) | Induction of autophagy in ER+ cells | [113] | |
| Apigenin | Primary epidermal keratinocytes and COLO16 cutaneous squamous cell carcinoma cells (20–80 µM + UVB) | Restoring of autophagy ↑ LC3 II | [114] | |
| TF1 erythroid subtype leukemia cells (100 µM) | Induction of autophagy ↑ ATG5, ATG12 expression | [115] | ||
| MM-F1, MM-B1 and H-Meso-1 malignant mesothelioma cells (50 μM) | No effects on autophagy | [116] | ||
| HT-29 colon cancer cells (15–60 µM) | Induction of autophagy ↑ Autophagosomes ↑ Beclin 1 and LC3 II expression ↓ p62 expression | [117] | ||
| BCPAP papillary thyroid carcinoma cells (12.5–50 µM) | Induction of autophagy ↑ Beclin 1 and LC3 II expression ↓ p62 expression | [118] | ||
| HepG2 HCC cells (10–40 µM) | Induction of cytoprotective autophagy ↑ LC3 II, ATG5 and Beclin 1 expression ↓ PI3K/Akt/mTOR pathway | [119] | ||
| HCT-116 colon cancer cells (6.25–50 µM) | Induction of cytoprotective autophagy ↑ Beclin 1 and LC3 II expression ↓ p62 expression | [120] | ||
| Wogonin | NPC-TW076 and NPC-TW039 nasopharyngeal carcinoma cells (50 µM) | Induction of cytoprotective autophagy ↑ LC3 II expression ↑ Autophagosomes and autolysosomes ↓ Akt/cRaf/ERK pathway | [121] | |
| SW48 colorectal cancer cells (4–16 µM) | Induction of autophagy ↑ Beclin 1 and LC3 II | [122] | ||
| BGC-823 gastric cancer cells (10, 50 and 200 µM + Oxaliplatin) | Induction of autophagy ↑ LC3 II | [123] | ||
| Baicalein | SMMC-7721 and Bel-7402 HCC cells (100 and 200 µM) | Induction of cytoprotective autophagy ↑ LC3 II | [124] | |
| HepG2 HCC cells (12.5–50 µM) | Induction of cytoprotective autophagy ↓ Akt/mTOR pathway | [125] | ||
| HEY and A2780 ovarian cancer cells (12.5–50 µM) | Induction of cytoprotective autophagy ↑ LC3 II expression and AVOs | [126] | ||
| CAL-27 OSCC cells (25–100 µM) | Induction of cytoprotective autophagy ↑ LC3 II, Beclin 1 and p62 | [127] | ||
| U251MG glioma cells (10–80 µM) | Induction of autophagy ↑ LC3 II expression ↑ AMPK pathway | [128] | ||
| Follicular undifferentiated thyroid cancer cells (10–80 µM) | Induction of autophagy ↑ p62, Beclin 1, ATG5, ATG12 expression | [129] | ||
| PC-3, MDA-MB-231 and DU145 cancer cells (5 µg/mL) | Induction of autophagy ↑ Autophagosomes ↑ AMPKα and ULK1 ↓ mTOR and Raptor | [130] | ||
| MCF-7 and MDA-MB-231 breast cancer cells (10, 20, 40 µM) | BALB/c-nude mice s.c. injected with MCF-7 or MDA-MB-231 cells (100 mg/kg, p.o., daily) | Induction of autophagy ↑ LC3 II and Beclin 1 expression ↓ PI3K/Akt pathway | [131] | |
| Stem cell-like cells (TICs) isolated from mouse and human liver tumors (30 µM) | Inhibition of autophagy ↓ Autophagosomes formation ↓ GTP binding of SAR1B GTPase | [132] | ||
| Polymethoxyflavone 5-demethylnobiletin (5-DMN) | CL1-5 and A549 NSCLC cells (12.5 μM) | Induction of cytoprotective autophagy ↑ Beclin 1 expression | [133] | |
| Luteolin | MET4 cells derived from a primary cutaneous invasive squamous cell carcinoma (50 µM) | Induction of cytoprotective autophagy ↑ Autophagosomes ↓ p62 expression | [134] | |
| NCI-H460 NSCLC cells (200 µM) | Induction of autophagy ↑ LC3B II expression | [135] | ||
| Huh7 HCC cells (20 µM) | Induction of autophagy ↑ LC3 II expression ↓ p62 expression | [136] | ||
| SMMC-7721 HCC cells (25–100 µM) | Induction of autophagy ↑ Autophagosomes ↑ LC3B II and Beclin 1 expression | [137] | ||
| Luteolin and Silibinin | U87MG and T98G glioma cells (Luteolin: 20 µM; Silibinin: 50 µM) | Inhibition of autophagy ↓ Beclin 1, LC3B I and II expression | [138] | |
| Salvigenin | SH-SY5Y neuroblastoma cells (25–50 µM) | Induction of cytoprotective autophagy ↑ LC3 II/I ratio ↑ ATG7 and ATG12 expression | [139] | |
| Baicalin | SMMC-7721 HCC cells (40–160 µM) | Induction of autophagy ↑ Beclin 1 expression | [140] | |
| Human bladder cancer T24 cells (100–200 µM) | Induction of autophagy ↓ p-Akt (Ser473) protein level and Akt kinase activity ↑ ATG complex, LC-3 and Beclin 1 expression | [141] | ||
| Diosmin | MCF-7, MDA-MB-231 and SK-BR-3 breast cancer cells (5–20 μM) | Induction of cytostatic and cytotoxic autophagy ↑ Oxidative stress and DNA damage | [142] | |
| Seed extract from Euterpe oleracea Mart. | MCF-7 breast cancer cells (10, 20 and 40 μg/mL) | Induction of autophagy ↑ LC3B II expression | [143] | |
| Delicaflavone | A549 and PC-9 NSCLC cells (40 μg/mL) | Induction of autophagy ↑ Autophagosomes ↑ LC3 II/I ratio ↓ p-Akt, p-mTOR and p-p70S6K | [144] | |
| Luteoloside | A549 and H292 NSCLC cells (60 µM) | Induction of autophagy ↓ Akt/mTOR/p70S6K signaling pathway ↑ Beclin 1 and LC3 II expression ↓ p62 expression | [145] | |
| Glychionide-A | PANC-1 pancreatic cancer cells (7–28 µM) | Induction of autophagy ↑ Beclin 1 and LC3 II expression ↓ p62 expression | [146] | |
| Isoorientin | HepG2 HCC cells (20–80 μM) | Induction of autophagy ↑ Beclin 1 and LC3 II expression | [147] | |
| Isovitexin | HepG2 and SK-Hep-1 HCC cells (12.5–50 μg/mL) | Induction of autophagy ↑ Beclin 1, LC3B II, ATG3 and ATG5 expression | [148] | |
| Wogonoside | U251MG and U87MG glioma cells (250–300 µM) | Induction of autophagy ↑ AVOs ↑ LC3 II expression ↓ p62 expression ↑ p38 MAPK ↓ PI3K/Akt/mTOR/p70S6K pathways | [149] | |
| Nobiletin | CAOV-3 and ES-2 ovarian cancer cells (40 μM) | Induction of autophagy ↑ p62 expression | [150] | |
| SNU-16 gastric cancer cells (12.5–50 μM) | Induction of autophagy ↑ LC3 II/I ratio ↓ p62 expression | [151] | ||
| Zapotin | HeLaPKCeA/E cancer cells with overexpressed constitutively active protein kinase C epsilon (30 μM) | Inhibition of autophagy ↓ Autophagosomes formation ↓ LC3 expression | [152] | |
| Vitexin | SK-Hep-1 and Hepa1-6 HCC cells (100 µM) | Inhibition of cytoprotective autophagy ↓ LC3 II expression | [153] | |
| Anthocyanins | ||||
| Delphinidin | MDA-MB-453 and BT474 HER-2+ breast cancer cells (MDA-MB-453: 80 µM) (BT474: 140 µM) | Induction of autophagy ↑ Autophagic vacuoles ↑ LC3 II expression ↑ ATG5-ATG12 expression ↓ p-Akt, p-mTOR, p70S6K, eIF4E ↑ LKB1, AMPK, ULK1, FOXO3a | [154] | |
| SMMC7721, HCCLM3 and MHCC97L HCC cells (80–150 µM) | Induction of autophagy ↑ Autophagic vacuoles ↑ LC3 II expression | [155] | ||
| U2OS osteosarcoma cells (10–200 µM) | Induction of autophagy ↑ Autophagosomes ↑ LC3 II expression ↓ p62 expression | [156] | ||
| HeLa cervical cancer cells (100 µM) | ATG5-deficient mouse embryonic fibroblasts (100 µM) | Induction of autophagy ↑ Autophagosomes and autolysosomes ↑ Colocalization LC3 II/LAMP-1
↓ Autophagy | [157] | |
| Pelargonidin | U2OS osteosarcoma cells (15–30 µM) | Induction of autophagy ↑ Beclin 1 and LC3 II expression ↓ LC3 I expression ↓ p-PI3K, p-Akt ↑ ROS | [158] | |
| Cyanidin | 786-O and ACHN RCC (25–100 µM) | ↓ ATG4, p62 expression ↓ LC3 II expression ↑ EGR1, SEPW1 expression | [159] | |
| C3G | INS-1 rat pancreatic β cells under oxidative stress condition (H2O2 treatment) (0.5–1 µM) | Reduction in H2O2-induced autophagy ↓ LC3 II expression ↓ Autophagic vacuoles ↓ MDC-labelled vacuoles ↑ HO-1, Nrf2 | [160] | |
| Polyphenols from Mulberry water extract (MPE) | p53+ HepG2 and p53− Hep3B HCC cells (0.25–1 mg/mL) | DEN-induced liver cancer in Wistar rat fed with normal diet + 1–2% MPE | ↑ Apoptosis of p53+ HepG2 ↑ Autophagy of p53− Hep3B, ↑ AMPK ↓ PI3K/Akt/mTOR signaling Protection from liver damage and HCC formation | [161] |
| SW1736 and HTh-7 thyroid cancer cells (10 µg/mL) | Induction of autophagy ↑ LC3 II/LC3 I ratio ↑ Autophagic vesicles ↑ LC3-puncta structures ↓ Akt/mTOR signaling | [162] | ||
| SGC-7901 gastric cancer cells | Induction of autophagy ↑ LC3 II/I ratio ↑ Beclin 1 expression | [163] | ||
| Anthocyanins, extracted from black soybean | U2OS osteosarcoma cells (100–300 µg/mL) | Induction of autophagy ↑ LC3 I to LC3 II conversion ↑ LC3-puncta structures ↑ p-ERK1/2, p-p38 MAPK, p-JNK, p-Akt ↓ p-mTOR ↑ AMPK | [164] | |
| Anthocyanins from Pelingo apple | MCF-7 and MDA-MB-231 breast cancer cells (2.5% v/v of Pelingo juice) | Induction of autophagy ↑ LC3 II/I ratio ↑ Autophagic vacuoles ↑ p21 expression ↓ ERK1/2 | [165] | |
| Illawarra plum extract | HT-29 colonic cancer cells (100 µg/mL) | Induction of autophagy ↑ Alteration of morphology ↑ Autophagic vacuoles ↑ SIRT1 expression ↑ Nuclear buds, micronuclei, and nucleoplasmic bridges | [166] | |
| Cinnamtannin D1 | A549 and H460 NSCLC cells (125–175 μM) | Induction of autophagy ↑ LC3-puncta structures ↑ LC3 II expression ↑ ATG5 expression ↓ Akt/mTOR ↑ ERK1/2 | [167] | |
| Proanthocyanidin-rich cranberry extract | JHAD1 and OE19 esophageal adenocarcinoma cells (75 μg/mL) | Induction of autophagy ↓ Beclin 1 expression ↑ LC3 II expression ↑ Autophagic vacuoles | [168] | |
| Flavanones | ||||
| Hesperidin | Male Swiss albino mice i.p. injected with AOM (25 mg/kg, p.o., daily) | Induction of autophagy ↑ Beclin 1 expression ↑ LC3 II expression ↓ PI3K/Akt/GSK-3β and mTOR pathways | [169] | |
| Naringin | Male C57BL/6 mice i.p. injected with AOM and DSS (50 and 100 mg/kg, p.o., daily) | Inhibition ER-stress mediated autophagy Inhibition formation of autophagosomes | [170] | |
| AGS gastric cancer cells (2 mM) | Induction of autophagy Formation of cytoplasmic vacuoles and autophagosomes ↑ Beclin 1 and LC3 II expression ↓ PI3K/Akt/mTOR pathway ↑ MAPKs | [171] | ||
| Pinocembrin | B16-F10 and A375 melanoma cells (50–150 µM) | C57BL/6 mice s.c. implanted into the oxter with B16-F10 cells (50 mg/kg or 75 mg/kg, i.v., daily) | Inhibition of autophagy ↑ LC3 I expression ↓ ATG5 and ATG5-ATG12 expression ↓ Beclin 1 expression ↑ p62 expression ↓ AVOs ↑ PI3K/Akt/mTOR pathway | [172] |
| 5-Methoxyflavanone | HCT-116 colon cancer cells (40 µM) | Induction of cytoprotective autophagy ↑ Autolysosomes ↑ LC3-puncta structures ↑ LC3 II expression ↑ LC3 I to LC3 II conversion | [173] | |
| 6-CEPN | SW620 and HCT-116 colon cancer cells (10 µM) | Induction of cytoprotective autophagy ↑ AVOs ↑ MDC-labelled vacuoles ↑ LC3-puncta structures ↑ LC3 II expression | [174] | |
| Liquiritin | SGC-7901/DDP cisplatin (DDP)-resistant gastric cancer cells (80 µM + DDP) | BALB/c-nu mice s.c. injected with SGC-7901/DDP cells (15 mg/kg, i.p., daily + DDP) | Induction of autophagy ↑ Beclin 1 expression ↑ LC3 II expression ↓ p62 expression | [175] |
| Silibinin | DU145 prostate cancer cells (100 µM + Arsenic) | Induction of autophagy ↑ Autophagic vacuoles ↑ Beclin 1 expression | [176] | |
| Isoflavones | ||||
| Genistein | A2780 ovarian cancer cells (50–100 µM) | Induction of autophagy ↑ LC3-puncta structures ↓ Akt phosphorylation | [177] | |
| MCF-7 breast cancer cells (100 µM) | Induction of autophagy ↑ LC3-puncta structures | [178] | ||
| DMBA-induced mammary tumors in Sprague–Dawley rats fed with AIN93G diet supplemented with 500 ppm genistein | Inhibition of autophagy ↓ GRP78, IRE1α, ATF4 and Beclin 1 genes ↑ Sensitivity to tamoxifen | [179] | ||
| A549 NSCLC cells (60 µM) | Induction of autophagy ↓ Bcl-xL levels ↑ LC3 II expression ↓ p62 expression Dissociation of Bcl-xL/Beclin 1 proteins | [180] | ||
| MIA PaCa-2 pancreatic cancer cells (100 µM) | Nude mice s.c. injected with MIA PaCa-2 cells (1.3 mg, i.p., every 4 days + 5-FU) | Induction of autophagy ↓ Bcl-2 expression ↑ Beclin 1 expression ↑ AVOs ↑ 5-FU anticancer effects | [181] | |
| TRAIL-resistant A549 NSCLC cells (10–40 µM) | Inhibition of autophagy ↑ LC3 II expression ↑ p62 expression ↑ TRAIL-induced cell death | [182] | ||
| I3C | HT-29 colon cancer cells (40 µM + I3C 300 µM) | Induction of autophagy ↑ LC3 II expression ↓ Akt/mTOR pathway ↓ Maturation of autophagosomes | [183] | |
| Puerarin | K562 CML cells (100 µM) | Induction of autophagy ↑ Autophagosomes ↑ LC3 II/I ratio ↑ ATG5 expression | [184] | |
| NCI-H441 NSCLC cells (20 µM) | Induction of autophagy ↑ MDC-labelled vacuoles ↑ ATG5 expression ↓ LC3 I expression ↓ Akt/mTOR pathway | [185] | ||
| NV-128 | EOC and R182 paclitaxel- and carboplatin-resistant ovarian cancer cells (0.1–10 µM) | Induction of autophagy ↑ LC3 II expression | [186] | |
| Furowanin A | HT-29 and SW480 colon cancer cells (2 and 5 µM) | Induction of autophagy ↑ AVOs ↑ LC3 II and Beclin 1 expression ↓ p62 expression ↑ Autophagosomes | [187] | |
| Glabridin | Huh7 hepatoma cells (1–100 µM) | Induction of cytoprotective autophagy ↑ AVOs ↑ LC3 II and Beclin 1 expression | [188] | |
| Celastrol | HeLa cervical cancer cells, A549 NSCLC cells, PC-3 prostate cancer cells (1.2 µM) | Induction of cytoprotective autophagy ↑ Autophagosomes ↑ LC3 II expression | [189] | |
| SH-SY5Y neuroblastoma cells (500 nM) | Induction of autophagy ↑ LC3 II/I ratio ↑ Autophagosomes | [190] | ||
| Phenoxodiol | KK ovarian clear cell carcinoma cells (0.5–2 µg/mL) | Inhibition of autophagy ↓ ATG7, ATG12, Beclin 1 expression ↑ Sensitivity to cisplatin | [191] | |
| Non-Flavonoids | ||||
| Coumarins | ||||
| Hybrid of 3-benzyl coumarin seco-B-ring derivative and phenylsulfonylfuroxan | A549 NSCLC cells (50 nM) | Induction of autophagy ↑ Autophagosomes ↑ LC3 II expression ↑ LC3 I to LC3 II conversion ↑ Autophagic flux | [192] | |
| Hybrid compound of coumarin and phenylsulfonylfuroxan | A549 and H1299 NSCLC cells (200 nM) | Induction of cytoprotective autophagy ↑ Autophagosomes ↑ LC3 I to LC3 II conversion | [193] | |
| Feroniellin A | A549RT-eto NSCLC cells (0.05–1 mM) | Induction of autophagy ↑ LC3 I to LC3 II conversion ↑ LC3-puncta structures ↑ Beclin 1 and ATG5 expression ↓ mTOR pathway | [194] | |
| Esculetin | HL-60 AML cells (20 µM) | Induction of autophagy ↑ Autophagosomes ↑ LC3 II and Beclin 1 expression ↓ ATG3 and p62 expression | [195] | |
| Xanthoxyletin | SCC-1 OSCC cells (5–20 µM) | Induction of autophagy ↑ Autophagosomes ↑ Beclin 1 and LC3 II expression ↓ p62 expression | [196] | |
| Osthole | T98G glioblastoma multiforme cells and MOGGCCM anaplastic astrocytoma cells (150–250 µM) | Induction of autophagy ↑ Autophagic cells ↑ Beclin 1 expression | [197] | |
| Psoralen and isopsoralen | PC-3 prostate cancer cells (250 µg/mL) | Induction of autophagy ↑ AVOs ↑ LC3 II expression ↓ p62 expression | [198] | |
| Geranylated 4-phenylcoumarin | PC-3 and DU145 prostate cancer cells (9 µM) | Induction of autophagy ↑ LC3 I to LC3 II conversion ↑ LC3-puncta structures ↑ Autophagosomes ↓ p62 expression | [199] | |
| Hydroxypyridinone-coumarin | MHCC-97 and HepG2 HCC cells (2 µM) | Induction of autophagy ↑ ATG5, ATG3, Beclin 1 and LC3 II expression ↓ p62 expression ↑ ERK1/2 ↓ Akt | [200] | |
| Psoralidin | HepG2 HCC cells (9–26 µM) | Induction of autophagy ↑ Autophagosomes ↑ Beclin 1 and LC3 II expression | [201] | |
| MCF-7 breast cancer cells (2.5–10 µM) | Induction of cytoprotective autophagy ↑ Beclin 1, LC3 II and p-ULK1 (Ser317) expression ↓ p62 expression ↓ Akt/mTOR pathway ↑ MDC-labelled vacuoles | [202] | ||
| Bergapten | MCF-7 and ZR-75 breast cancer cells (20–50 µM) | Induction of autophagy ↑ Beclin 1, PI3KII, UVRAG, AMBRA ↑ LC3 I to LC3 II conversion ↑ Autophagosomes ↑ pTEN and p38 MAPK/NF-Y pathway ↓ Akt/mTOR pathway | [203] | |
| Curcuminoids | ||||
| Curcumin | A549 NSCLC cells (5–40 µM) | Induction of autophagy ↑ Autophagic vesicles ↑ MDC-labelled vacuoles ↑ Double membrane-enclosed structures ↑ LC3 II and Beclin 1 expression ↑ LC3 II/I ratio ↓ p62 expression ↓ Akt, mTOR expression | [204,205] | |
| A549 NSCLC cells (40 µM) | Induction of autophagy ↑ LC3-puncta structures ↑ AMPK pathway ↑ hST8Sia I, GD | [206] | ||
| H1299 and A549 NSCLC cells (10 µM) | Induction of autophagy ↑ Beclin 1 expression ↑ LC3 II/I ratio ↑ Autophagosomes ↓ p-mTOR, p-S6, p-PI3K, p-Akt | [207] | ||
| A549 NSCLC cells (40 µM + Galbanic acid) | Induction of autophagy ↑ LC3-puncta structures ↑ LC3 II expression ↑ Beclin 1 expression ↓ p-Akt, p-mTOR, p-p70S6K | [208] | ||
| H157 and H1299 NSCLC cells (10 µM + Gefitinib) | BALB/c athymic nude mice s.c. injected with H157 or H1299 cells (1 g/kg, p.o., daily + Gefitinib) | Induction of autophagy ↑ LC3-puncta structures ↑ AVOs ↓ SQSTM1
↑ LC3, Beclin 1 ↓ EGR, survivin, Sp1, HDAC1 | [209] | |
| PANC1 and BxPC3 pancreatic cancer cells (10–80 µg/mL) | Induction of autophagy ↑ LC3 II expression ↑ Autophagosomes ↑ LC3-puncta structures | [210] | ||
| SGC-7901 and BGC-823 gastric cancer cells (10–40 µM) | Induction of autophagy ↑ Beclin 1, ATG5, ATG3 expression ↑ LC3 I to LC3 II conversion ↓ p-mTOR, PI3K, p-Akt | [211] | ||
| SGC-7901, BGC-823 and MKN-28 gastric cancer cells (5–20 µM) | Induction of autophagy ↑ AVOs ↑ Beclin 1, ATG7, ATG5-ATG12 expression ↑ LC3 I to LC3 II conversion ↓ p-Akt, p-mTOR, p-p70S6K | [212] | ||
| SKOV-3 and A2780 ovarian cancer cells (SKOV-3: 10–40 µM) (A2780: 7.5–30 µM) | Induction of autophagy ↑ AVOs ↑ LC3 I/II ratio ↑ LC3-puncta structures ↑ Beclin 1, ATG3 expression ↓ p-Akt, p-mTOR, p-p70S6K, p-4EBP1 | [213] | ||
| SW620 and HCT-116 colon cancer cells (10–30 µM) | Induction of autophagy ↑ LC3 expression ↓ p62 expression ↓ YAP expression | [214] | ||
| HCT-116 and HT-29 colon cancer cells (10–30 µM + 5-FU) | BALB/c nu/nu mice s.c. injected with HCT-116 cells (40 mg/kg, i.p., daily + 5-FU) | Induction of autophagy ↓ LC3 II/II ratio, Beclin 1 expression ↑ p62 expression ↓ p-Akt, p-mTOR, p-AMPK, p-ULK1 | [215] | |
| HepG2 HCC cells (5–20 µM) | BALB/c nude mice s.c. injected with HepG2 cells (200 mg/kg, i.p., daily) | Induction of autophagy ↑ Beclin 1, LC3 expression ↓ Glypican-3 (GPC3) ↓ Wnt/β-catenin pathway | [216] | |
| Sprague Dawley rats (100–200 mg/kg, p.o., daily + TAA) | Induction of autophagy ↑ LC3 II expression ↓ ALT, AST, albumin ↑ Survival | [217] | ||
| HeLa cervical cancer cells, HCT-116 colon carcinoma cells, HepG2 HCC cells (40 µM + JLP silencing) | Induction of cytoprotective autophagy ↑ LC3 II expression ↓ Autophagosomes-lysosome fusion and degradation with JLP silencing | [218] | ||
| 786-O and ACHN RCC cells (5–80 µM) | ↑ LC3 II expression
↑ p-AMPK ↑ GRP78, CHOP expression ↓ ROS production
↓ p-AMPK ↓ GRP78, CHOP expression ↑ ROS production | [219] | ||
| MM-B1, H-Meso-1, MM-F1 and murine #40a malignant mesothelioma cells (25 µM) | Induction of autophagy, but autophagic flux blocked ↑ p62/SQSMT1, LC3 I | [220] | ||
| TUBO murine Her2/neu+ breast cancer cells (25 μM + CQ) | Immunocompetent or immunocompromised BALB/c mice s.c. injected with TUBO cells (100 mg/kg, p.o., 3 times a week + CQ) | ↓ p62 expression ↑ LC3 II expression
↑ Tumor growth ↑ Autophagosomes in tumor tissues ↓ p62 expression ↓ Foxp3+ T regulatory cells ↑ CD8+ T cells
↓ Tumor growth | [221] | |
| CAL-27, SCC-15 and FaDu HNSCC cancer cells (25 μM + Resveratrol) | Induction of autophagy ↑ LC3 I to LC3 II conversion ↑ LC3 II expression ↑ Autophagosomes | [222] | ||
| SUP-B15 Ph+ acute lymphoblastic leukemia cells (30 µM) | Induction of autophagy ↑ RAF/MEK/ERK pathway ↑ Autophagy | [223] | ||
| A172 glioblastoma cells (10 µM) | Induction of autophagy ↑ LC3 II expression ↑ LC3-puncta structures ↑ ATG5, ATG12, Beclin 1 expression | [224] | ||
| Caki, ACHN and A498 renal carcinoma cells; U87MG glioma cells; MDA-MB-231 breast carcinoma cells (20 µM + PP242) | Induction of autophagy ↓ mTORC2/Akt pathway ↓ Rictor, Akt ↑ Cytosolic Ca2+, MMP, cytosolic pH ↑ Autophagy, Lysophagy, galectin-3 Colocalization of LAMP-1 and p62 ↑ LC3-puncta structures ↑ LC3 II expression ↑ p62 expression | [225] | ||
| HCT-116, HT-29, HepG2 and Huh7 gastrointestinal cancer cells (2 µM + Sildenafil) | Induction of autophagy ↓ mTORC1 and mTORC2 activity ↑ Beclin 1 ↑ Autophagosome and autolysosome | [226] | ||
| CUR or solid lipid CUR particles (SLCP) | U87MG; mouse, GL261; rat, F98 glioblastoma cells; C6-glioma rat glial tumor cells; N2a cells mouse neuroblastoma cells (25 µM) | Induction of autophagy ↑ ATG5, ATG7, Beclin 1 ↑ LC3A/B-II/LC3A/B-I ratio ↓ Mitophagy markers PINK-1, NIP3L/NIX, BNIP3, HIH-1α ↓ Akt/mTOR pathway ↓ Akt, p-Akt, mTOR and p-mTOR ↓ LAMP-2a ↑ Autophagic vacuoles and membrane blebbing | [227] | |
| CUR-LDH | MDA-MB-231 breast cancer cells (25–100 µg/mL + Photodynamic therapy) | ↑ Autophagosomes | [228] | |
| Curcumin DMC BDMC | SAS oral cancer cell line (CUR: 30 µM) (DMC, BDMC: 15 µM) | Induction of autophagy ↑ Autophagic vacuoles ↑ MDC-labelled vacuoles ↓ p-mTOR, ↑ AMPKα1, Vps34, ULK1, ATG16L1, ATG5 ↑ LC3, Beclin 1 | [229] | |
| Curcumin DMC BDMC TetrahydroCUR | SAS oral cancer cell line + Gefitinib (CUR: 20 µM) (DMC, BDMC: 5 µM) | BALB/c athymic nude mice s.c. injected with SAS cells (30 mg/kg, i.p., every two days + gefitinib) | Induction of autophagy ↑ Cleaved-caspase-3 ↓ MMP ↑ Autophagic vacuoles ↑ ATG5, p62/SQSTM1, ULK1, Vps34 ↑ LC3, Beclin 1
↑ Beclin 1 | [230] |
| Chemotherapy-resistant HL60 human leukemia cells | Induction of autophagy ↑ LC3 II expression ↑ p62 expression | [231] | ||
| TetrahydroCUR | A549 NSCLC cells (10–130 µM) | Induction of autophagy ↑ AVOs ↑ Beclin 1 expression ↓ mTOR, p-mTOR, p-Akt ↓ p62 expression ↑ LC3 II/I ratio ↑ PI3K | [232] | |
| CA-5f | A549, H1299 and H157 NSCLC cells; HUVEC umbilical vein endothelial cells; HepG2 HCC cells; HeLa cervical cancer cells; HEK293 embryonic kidney 293 cells (1–40 µM) | BALB/c nude mice s.c. injected with A549 cells (40 mg/kg, i.v., every two days) | Inhibition of autophagy ↑ SQSTM1 expression ↓ Autophagophores degradation Modulation of cytoskeleton protein, membrane trafficking, vesicles mediated transport
↑ LC3 II expression ↑ SQSTM1 | [233] |
| ZYX01 | A549 NSCLC cells | Induction of autophagy ↑ AMPK/ULK1/Beclin 1 pathway ↑ LC3 II/I ratio ↑ Beclin 1 ↓ p62 expression | [234] | |
| MOMI-1 | A549 NSCLC cells; MCF-7 breast cancer cells; HepG2 HCC cells (20 µM) | Induction of autophagy ↑ Autophagic vacuoles ↑ MDC-labelled vacuoles ↑ LC3-puncta structures ↓ p62 expression ↑ Beclin 1 ↑ LC3 I to LC3 II conversion | [235] | |
| MTH-3 | MDA-MB-231 breast cancer cells (10 µM) | Induction of autophagy ↑ LC3 expression ↑ p62 expression ↑ ATG5, ATG7, ATG12, Beclin 1 | [236] | |
| WZ35 | HCCLM3 HCC cells (20 µg/mL) | Induction of autophagy ↑ Autophagic vacuoles Deregulated YAP signaling ↓ LC3 I/II ratio ↓ ATG7, Beclin 1 expression ↑ p62 expression | [237] | |
| Phenolic Acids | ||||
| Ellagic acid | HOP62 and H1975 lung cancer cells (10–50 µM) | BALB/c nude mice s.c. injected with HOP62 cells (40 mg/kg, i.p., every 2 days) | Induction of autophagy ↑ LC3-positive autophagosomes ↑ LC3 II and ATG5 expression ↓ p62 expression ↑ Activation AMPK ↓ mTORC1 and Akt | [238] |
| SKOV-3 ovarian cancer cells (36.6 µM) | Induction of autophagy ↑ LC3 II/I ratio ↑ Beclin 1 and ATG5 expression ↓ p62 expression ↑ Activation AMPK ↓ mTORC1 and Akt | [239] | ||
| Punicalagin | U87MG glioma cells (1–30 µg/mL) | Induction of autophagy ↑ LC3 II expression ↓ LC3 I expression ↑ LC3-puncta structures ↑ Activation AMPK and p27 | [240] | |
| Grias Nuberthii extract | RKO and SW613-B3 colon cancer cells (20, 30, 50 µg/mL) | Induction of autophagy ↑ Beclin 1 and LC3 II expression ↓ p62 expression | [241] | |
| Gallic acid | Cal33 OSCC cells (0.1, 0.5, 1 mg/mL) | Induction of cytoprotective autophagy, but autophagic flux blocked ↑ LC3 I to LC3 II conversion ↑ Beclin 1 and ATG5-ATG12 expression ↑ p62 expression ↓ Autolysosome formation | [242] | |
| Paeonol | A2780 and SKOV-3 ovarian cancer cells (0.6–1.2 mM) | BALB/c nude mice (nu/nu) s.c injected with A2780 cells (40 mg/kg, i.p., every 2 days) | Induction of cytoprotective autophagy ↑ LC3 I to LC3 II conversion ↑ Autophagosomes ↓ p62 expression ↓ Akt/mTOR pathway | [243] |
| PGG | DU145, PC-3, TRAMP-C2 prostate cancer cells (25–75 µM) | Induction of cytoprotective autophagy ↑ Autophagosomes ↑ LC3 II expression ↓ S6K and 4EBP1 ↑ Akt activation | [244] | |
| Corilagin | SGC-7901 and BGC-823 gastric cancer cells (10–30 µM) | Induction of cytoprotective autophagy ↑ LC3 II expression ↑ Autophagosomes | [245] | |
| CAPE + EECP | MDA-MB-231 breast cancer cells (CAPE: 25 µg/mL + EECP: 25–100 µg/mL) | Induction of autophagy ↑ LC3 II expression ↓ p62 expression | [246] | |
| CAPE | C6 glioma cells (10 µM) | Induction of cytoprotective autophagy ↑ LC3 II/I ratio ↑ Autophagosomes ↑ AMPK activation | [247] | |
| Decyl caffeic acid | HCT-116 colorectal cancer cells (40 µM) | Induction of cytoprotective autophagy ↑ ATG3, ATG16, Beclin 1 and LC3 I/II expression | [248] | |
| Artepillin C | CWR22Rv1 prostate cancer (50–100 µM) | Induction of cytoprotective autophagy ↑ LC3 II expression | [249] | |
| Cinnamomum cassia extracts | SASVO3 oral cancer cells (50–100 µg/mL) | Induction of cytoprotective autophagy ↑ AVOs ↑ LC3 I, LC3 II, ATG14, rubicon and p62 expression ↓ PI3K/Akt/mTOR pathway | [250] | |
| Ferulic acid | HeLa and Caski cervical cancer cells (2.0–4.0 mM) | Inhibition of autophagy ↓ LC3 II, Beclin 1, ATG5-ATG12 expression | [251] | |
| Tributyltin (IV) ferulate | HCT-116, HT-29, Caco-2 colon cancer cells (400 nM) | Induction of autophagy ↑ MDC-labelled vacuoles ↑ LC3 II and p62 expression | [252] | |
| p-coumaric acid | N2a neuroblastoma cells (150–200 µM) | Induction of autophagy ↑ Autophagosomes ↑ LC3 II expression | [253] | |
| Ganoderma lucidum methanolic extract | AGS gastric cancer cells (66.6 and 133.2 µM) | Induction of autophagy ↑ Autophagosomes ↑ LC3 II expression ↓ p62 expression | [254] | |
| Lignans | ||||
| Honokiol | B16-F10, SKMEL-28 melanoma cancer cells (30–40 µM) | Induction of autophagy Formation of autophagosomes ↑ LC3 II expression and cytoplasmic accumulation ↓ Akt/mTOR pathway and Notch signaling | [255,256] | |
| MG-63 osteosarcoma cells (10–20 µg/mL) | Induction of autophagy ↑ LC3 II expression ↓ Akt/mTOR pathway | [257] | ||
| ARO, WRO, SW579 thyroid cancer cells (20–60 µM) | BALB/cAnN.Cg-Foxn1nu/CrlNarl nude mice s.c. injected with ARO cells (5 or 15 mg/kg, p.o., every 3 days) | Induction of autophagy ↑ LC3 II expression ↓ p62 expression | [258] | |
| Neuro-2a and NB41A3 neuroblastoma cells (50 µM) | Induction of autophagy ↑ AVOs ↑ LC3 II/I ratio | [259] | ||
| Drug sensitive (U87MG, murine GL261) and resistant (U87-MR-R9) glioma cells (40 µM + TMZ) | Induction of autophagy ↑ Autophagic cells percentage | [260] | ||
| PC-3, LNCaP, murine Myc-CaP prostate cancer cells (40 µM) | Induction of cytoprotective autophagy ↑ LC3 II expression Formation of autophagic vacuoles ↑ LC3-puncta structures | [261] | ||
| OC2 and OCSL OSCC cells (20–40 µM) | BALB/cAnN.Cg-Foxn1nu/CrlNarl nude mice s.c. injected with SAS cells (5 and 15 mg/kg, p.o., twice a week) | Induction of autophagy ↑ LC3 II expression | [262] | |
| Honokiol + Magnolol | U87MG and LN229 glioma cells (40 µM, each) | Induction of cytoprotective autophagy ↑ LC3 II expression | [263] | |
| Vitexin 6 | T-47D breast cancer cells and RKO colon cancer cells (5–20 µM) | Induction of autophagy Formation of autophagosomes ↑ LC3 II conversion ↑ LC3 II and Beclin 1 expression | [264] | |
| Licarin A | A549 and NCI-H23 NSCLC cells (10–25 µM) | Induction of autophagy ↑ AVOs ↑ Beclin 1, LC3 II mRNA levels ↓ p62 levels | [265] | |
| Trachelogenin | HCT-116 colon cancer cells (5–10 µM) | Induction of autophagy ↑ AVOs ↑ Beclin 1 ↑ LC3 I to LC3 II conversion | [266] | |
| Magnolin | HCT-116 and SW480 colon cancer cells (10–40 µM) | BALB/c athymic nude mice s.c. injected with HCT-116 cells (20 mg/kg, i.p., daily) | Induction of autophagy ↑ LC3 II expression ↓ p62 expression Accumulation of double membrane vesicles ↑ LC3-puncta structures | [267] |
| Justicidin A | HT-29 colon cancer cells (0.5–1.5 µM) | NOD-SCID mice s.c. injected with HT-29 cells (6.2 mg, p.o., daily) | Induction of autophagy ↑ LC3 I to LC3 II conversion ↑ LC3 II expression ↑ AVOs ↑ LC3-puncta structures ↓ p62 expression ↓ p-mTOR, p-p70S6K expression ↑ Beclin 1, ATG5-ATG12 expression | [268] |
| Pinoresinol | SKOV-3 ovarian cancer cells (10–40 µM) | Mice s.c. injected with SKOV-3 cells (40 mg/kg, i.p., thrice a week) | Induction of autophagy Formation of autophagic vesicles ↑ LC3 II expression ↑ Beclin 1 expression ↓ p62 expression | [269] |
| Sesamin | HeLa cervical cancer cells (50 µM) | Induction of autophagy ↑ Autophagosomes ↑ LC3 II expression ↑ Beclin 1 expression | [270] | |
| HT-29 and LS180 colon cancer cells (50 µM) | Induction of autophagy Formation of double membrane vacuoles ↑ LC3-puncta structures ↑ LC3 I to LC3 II conversion ↑ MDC-labelled vacuoles | [271] | ||
| Magnolol | H460, A549 and NCI-H1299 NSCLC cells (A549: 80 µM) (A549 and NCI-H1299: 10–20 µM) | Induction of autophagy Formation of autophagosomes ↑ MDC-labeled vacuoles ↑ ATG5, ATG12 expression ↑ LC3 II/I ratio ↓ p62 expression ↓ Akt/mTOR pathway | [272,273] | |
| SGC-7901 human gastric adenocarcinoma cells (40–80 µM) | Induction of autophagy ↑ AVOs | [274] | ||
| Arctigenin | MCF-7 breast cancer cells (1–200 µM) | Induction of autophagy ↑ LC3 II expression ↑ LC3 II/I ratio ↓ mTOR pathway | [275] | |
| R-SW480, R-SW620 cisplatin resistant colorectal cancer (100 µM + Cisplatin) | Induction of autophagy ↑ LC3 II expression ↑ p65 expression ↓ LC3 I expression | [276] | ||
| HepG2 HCC cells (1.25–10 µM) | Inhibition of autophagy ↑ LC3 II expression ↑ Beclin 1 phosphorylation ↑ p62 levels | [277] | ||
| DFS | DU145 prostate cancer cells and SW480 colon cancer cells (10 µM) | Induction of cytoprotective autophagy ↑ LC3 II levels ↑ Autophagosomes and autolysosomes | [278] | |
| Stilbenes | ||||
| Resveratrol | HL-60 AML cells (12.5–100 μM) | Induction of autophagy ↑ LC3 II, ATG5 and Beclin 1 expression ↑ LKB1/AMPK activation ↓ PI3K/Akt/mTOR pathway | [279] | |
| K562 CML cells (50 μM) | Induction of autophagy ↑ AMPK activation ↓ mTOR pathway ↑ LC3 II and ATG3 expression ↑ p62 expression | [280] | ||
| HT-29 and COLO201 colon cancer cells (HT-29: 150 μM) (COLO201: 75 μM) | Induction of autophagy ↑ LC3 II expression ↑ Autophagic vacuoles ↑ LC3-puncta structures | [281] | ||
| HK-2 and Ketr-3 renal carcinoma cells (12.5–100 μM) | Induction of autophagy ↑AMPK activation ↓ mTOR phosphorylation ↑ LC3, ATG5, ATG7 expression | [282] | ||
| MHCC-97 HCC cells (20–100 μM) | Induction of autophagy ↑ LC3 II/I ratio ↑ Beclin 1 expression ↓ p62 expression ↑ LC3-puncta structures ↓ p-Akt/Akt ratio | [283] | ||
| C33A, CaLo, and HeLa cervical cancer cells (30–50 μM) | Induction of autophagy ↑ Lysosomal permeability ↑ Lysosomal swelling and degranulation ↑ Vacuoles and autophagosomes | [284] | ||
| CAR cisplatin-resistant oral carcinoma cells (50 μM) | Induction of autophagy ↑ AVOs ↑ MDC-labeled vacuoles ↑ LC3-puncta structures ↑ AMPK activation ↓ Akt activation ↑ ATG5, ATG7, ATG12, ATG14, ATG16L1, Beclin 1 and LC3 II expression | [285] | ||
| U373 glioma cells (100 μM) | Induction of autophagy ↑ LC3-labeled vesicles | [286] | ||
| U87 glioma cells (30 μM) | Induction of cytoprotective autophagy ↑ Autophagosomes ↑ ATG5, Beclin 1 and LC3 II expression ↓ Akt and p70S6K activation | [287] | ||
| U251 glioma cells (150 μM) | Induction of cytoprotective autophagy ↑ MDC-labeled vacuoles ↑ LC3-puncta structures ↑ LC3 II and Beclin 1 expression | [288] | ||
| OVCAR-3 and CAOV-3 ovarian cancer cells (30μM) | Induction of autophagy ↑ LC3 II and ATG5 expression | [289] | ||
| SKOV-3 ovarian cancer cells (25 μM) | Induction of cytoprotective autophagy ↑ LC3 II and Beclin 1 expression ↑ MDC-labeled vacuoles | [290] | ||
| OVCAR-3 and CAOV-3 ovarian cancer cells (120 μM) | Induction of autophagy ↑ LC3 II and Beclin 1 expression ↑ LC3-labeled vesicles ↑ Autophagosomes ↓ STAT3 activation | [291] | ||
| Female nu/nu mice injected i.p. with GFP-labeled A2780 ovarian carcinoma cells (160 mg/kg, i.p., daily) | Induction of autophagy ↑ Autophagosomes | [292] | ||
| Ishikawa endometrial carcinoma cells (20 μM) | Induction of cytoprotective autophagy ↑ LC3 II expression ↑ Autophagosomes ↑ p-ERK and p-AMPKα | [293] | ||
| A549 NSCLC cells (50 μM) | Induction of autophagy ↑ p62 degradation ↑ LC3 II expression ↑ MDC-labeled vacuoles | [294] | ||
| A549 and H1299 NSCLC cells (200 μM) | Induction of cytoprotective autophagy ↑ Beclin 1 expression ↑ LC3 II/I ratio ↓ p62 expression ↑ SIRT 1 expression ↓ Akt/mTOR pathway ↑ p38 MAPK activation | [295] | ||
| PC9 NSCLC cells (40 μΜ + Gefitinib) | Induction of cytoprotective autophagy ↑ LC3B II expression ↑ MDC-labelled vacuoles | [296] | ||
| A549 NSCLC cells (2.5 μM + Cisplatin) | Induction of autophagy ↑ Autophagosomes ↑ LC3 II expression ↓ p62 expression ↓ p-Akt ↑ LC3-puncta structures ↑ Autophagosomes | [297] | ||
| MSTO-211H and H-2452 malignant mesothelioma cells (30 μM + Cisplatin) | Induction of cytoprotective autophagy ↑ LC3 A and Beclin 1 expression | [298] | ||
| EC109 and EC9706 squamous esophageal carcinoma cells (10–150 μM) | Induction of cytoprotective autophagy ↑ MDC-labelled vacuoles ↑ AVOs ↑ LC3 II, ATG5 and Beclin 1 expression ↑ Autophagosomes | [299] | ||
| B16 melanoma cells (25–100 μM) | Induction of cytoprotective autophagy ↑ LC3 II and Beclin 1 expression ↓ Akt/mTOR pathway | [300] | ||
| MCF-7 breast cancer cells (64 μM) | Induction of autophagy ↑ LC3-puncta structure ↑ LC3 II expression ATG7, Beclin 1 and Vps34 expression unchanged ↓ Akt/mTOR pathway | [301] | ||
| MCF-7 and MDA-MB-231 breast cancer cells (100 μM + Rapamycin) | Inhibition of rapamycin-induced autophagy ↓ LC3 II expression ↑ p62 expression ↓ Akt activation | [302] | ||
| MCF-7 and MDA-MB-231 breast cancer cells (MCF-7: 84.6 μM + DOX) (MDA-MB-231: 108 μM + DOX) | Inhibition of DOX-induced autophagy ↓ LC3B expression ↓ Beclin 1 expression ↓ Akt activation | [303] | ||
| MDA-MB-231 breast cancer cells (72 μM + Salinomycin) | Inhibition of salinomycin-induced autophagy ↓ LC3 expression ↓ Beclin 1 expression | [304] | ||
| A431 epidermoid carcinoma cells (50 μM) | No autolysosome formation ↑ LAMP-2 expression ↑ LC3 II expression ↓ Rictor expression | [305] | ||
| Pterostilbene | HL-60 AML cells | Induction of autophagy ↑ LC3 II expression ↑ LC3-puncta structures Accumulation autophagic vacuoles Reduction in autophagic degradation | [306] | |
| Bcap-37 and MCF-7 breast cancer cells (50 μM) | Induction of cytoprotective autophagy ↑ LC3B II expression ↑ Autophagosomes | [307] | ||
| CAR cisplatin-resistant human oral cancer cells (50 and 75 μM) | Induction of autophagy ↑ AVOs ↑ MDC-labelled vacuoles ↑ ATG5, ATG7, ATG12, Beclin 1 and LC3 II expression ↓ p-Akt | [308] | ||
| Xanthones | ||||
| α-Mangostin | GBM8401 and DBTRG-05MG glioblastoma cells (2.5–10 µM) | BALB/cA-ν (ν/ν) nude mice s.c. injected with GBM8401 cells (2 mg/kg, i.p., daily) | Induction of autophagy ↑ AVOs ↑ MDC-labelled vacuoles ↑ Autophagosomes ↑ LC3-puncta structures ↑ AMPK pathway ↓ mTORC1 activity | [309] |
| ICR mice treated topically with DMBA/TPA (5 and 20 mg/kg, i.p., daily) | Induction of autophagy ↑ LC3, LC3 II, Beclin 1 expression ↓ LC3 I and p62 expression ↓ PI3K/Akt/mTOR pathway | [310] | ||
| K562, KBM5 and KBM5-T135I CML cells (5–20 µM) | Induction of cytoprotective autophagy ↑ LC3 II expression ↑ Autophagic vacuoles | [311] | ||
| Gambogic acid | K562 CML cells (0.5–2 µM) | Induction of autophagy ↑ Autophagic vacuoles ↑ LC3 II and Beclin 1 expression ↓ p62 expression | [312] | |
| NCI-H441 NSCLC cells | Induction of autophagy ↑ Beclin 1 expression ↑ LC3 I to LC3 II conversion ↑ Autophagosomes | [313] | ||
| TE13 esophageal cancer cells (0.25–1 µM) | Induction of autophagy, but autophagic flux blocked ↑ LC3 II expression ↑ Autophagosomes ↓ Akt/mTOR pathway | [314] | ||
| HCT-116 and SW260 colon cancer cells (0.25–1.5 µM) | BALB/c mice s.c. injected with C26 cells (8 mg/kg, i.p., daily) | Induction of cytoprotective autophagy ↑ Autophagic vacuoles ↑ AVOs ↑ LC3 I to LC3 II conversion ↑ Beclin 1, ATG7, ATG5-ATG12 expression ↓ p62 expression ↓ Akt/mTOR pathway | [315] | |
| Gambogenic acid | H1975, H460 lung cancer cells (1.25–25 µM) | Induction of autophagy ↑ Autophagosomes ↑ LC3-puncta structures ↑ LC3 II expression ↑ GSK3β activation ↓ Akt/mTOR pathway | [316] | |
| Isogambogenic acid | A549 NSCLC cells and HeLa cervical cancer cells (1.5–12 µM) | BALB/cA nude mice s.c. injected with A549 cells (16 mg/kg, i.v., twice a week) | Induction of autophagy, but autophagic flux blocked ↑ Autophagic vacuoles ↑ LC3 II and Beclin 1 expression ↓ p70S6K phosphorylation Inhibition degradation p62 Inhibition acidification of vacuoles | [317] |
| Gartanin | Hep3B, HepG2, Huh7 HCC cells (10–40 µM) | Induction of cytoprotective autophagy ↑ AVOs ↑ LC3 I to LC3 II conversion ↑ LC3-puncta structures ↑ Autophagosomes and autolysosomes ↓ p62 expression | [318] | |
| T24 and RT4 urinary bladder cancer cells (10–25 µM) | Induction of autophagy ↑ LC3-puncta structures ↑ Autophagosomes ↑ LC3 I to LC3 II conversion ↓ p70S6K and 4E-BP1 | [319] | ||
| T98G glioma cells (10 µM) | Induction of autophagy ↑ LC3-puncta structures ↑ LC3 II and Beclin 1 expression ↓ p62 expression ↓ PI3K/Akt/mTOR pathway | [320] | ||
| 22Rv1 and PC-3 prostate cancer cells (6–24 µM) | Induction of autophagy ↑ LC3-puncta structures ↑ LC3 II expression | [321] | ||
| Formoxanthone C | MDR A549RT-eto NSCLC cells (20 µg/mL) | Induction of autophagy ↑ Autophagic vacuoles ↑ LC3-puncta structures ↑ LC3 I to LC3 II conversion ↑ Beclin 1 expression ↓ p-mTOR levels ↑ Sensitivity to etoposide | [322] | |
| Mangiferin | Gemcitabine-resistant Mia-PaCa2 pancreatic carcinoma cells (5–20 µM) | Induction of autophagy ↑ LC3 II and Beclin 1 expression | [323] | |
| Gentiana dinarica extract and norswertianin | U251 glioblastoma cells (Extract: 50 µg/mL; noswertianin 40 µM) | Induction of autophagy ↑ AVOs ↑ LC3 I to LC3 II conversion ↓ p62 expression ↓ Akt/mTOR pathway | [324] | |
| Xanthone V1 | HeLa cervical cancer cells (10–20 µM) | Induction of autophagy ↑ LC3-puncta structures ↑ LC3 I to LC3 II conversion ↓ p62 expression | [325] | |
| Cudraxanthone D | Ca9-22 and SCC25 OSCC cells (50 µM) | Inhibition of autophagy ↓ Autophagic vacuoles | [326] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benvenuto, M.; Albonici, L.; Focaccetti, C.; Ciuffa, S.; Fazi, S.; Cifaldi, L.; Miele, M.T.; De Maio, F.; Tresoldi, I.; Manzari, V.; et al. Polyphenol-Mediated Autophagy in Cancer: Evidence of In Vitro and In Vivo Studies. Int. J. Mol. Sci. 2020, 21, 6635. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186635
Benvenuto M, Albonici L, Focaccetti C, Ciuffa S, Fazi S, Cifaldi L, Miele MT, De Maio F, Tresoldi I, Manzari V, et al. Polyphenol-Mediated Autophagy in Cancer: Evidence of In Vitro and In Vivo Studies. International Journal of Molecular Sciences. 2020; 21(18):6635. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186635
Chicago/Turabian StyleBenvenuto, Monica, Loredana Albonici, Chiara Focaccetti, Sara Ciuffa, Sara Fazi, Loredana Cifaldi, Martino Tony Miele, Fernando De Maio, Ilaria Tresoldi, Vittorio Manzari, and et al. 2020. "Polyphenol-Mediated Autophagy in Cancer: Evidence of In Vitro and In Vivo Studies" International Journal of Molecular Sciences 21, no. 18: 6635. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186635