MALDI-TOF Mass Spectrometry Revealed Significant Lipid Variations in Follicular Fluid and Somatic Follicular Cells but Not in Enclosed Oocytes between the Large Dominant and Small Subordinate Follicles in Bovine Ovary

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
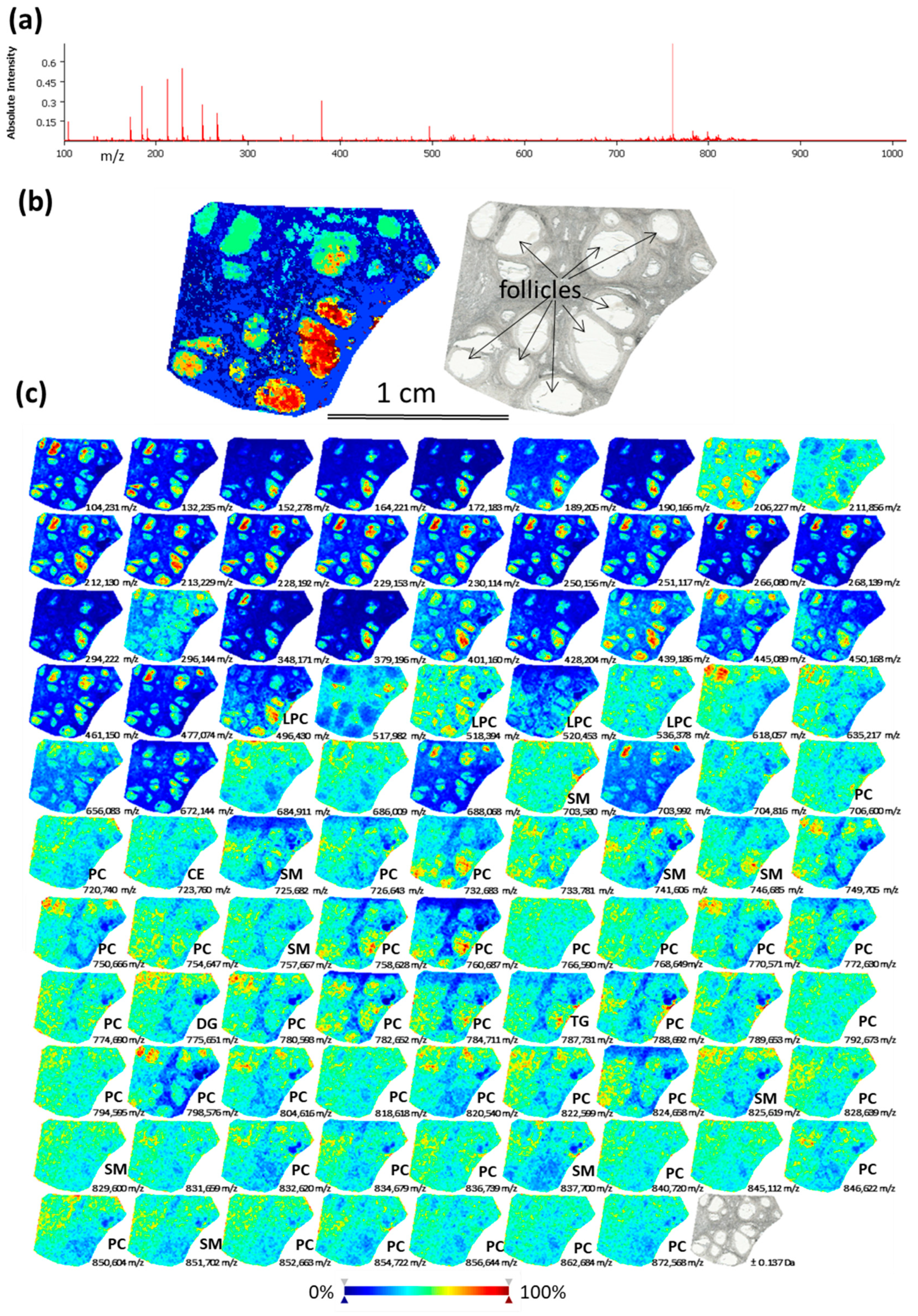
2.1. Analysis of Lipids by Mass Spectrometry Imaging (MSI) on Bovine Ovarian Sections
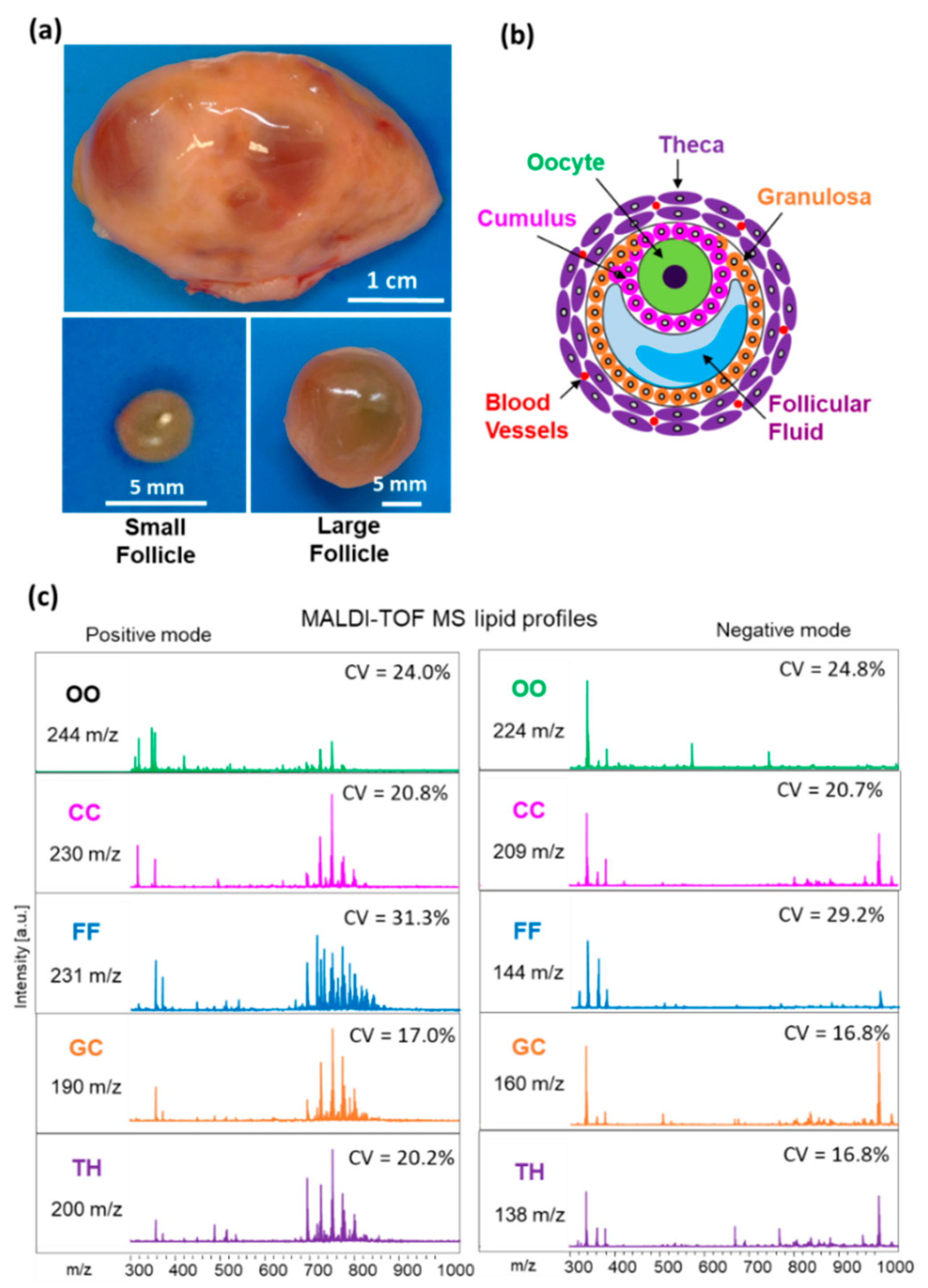
2.2. Comparison of Lipid Profiles in Follicular Cells and Fluids of Small and Large Follicles
2.3. Gene Expression Analysis
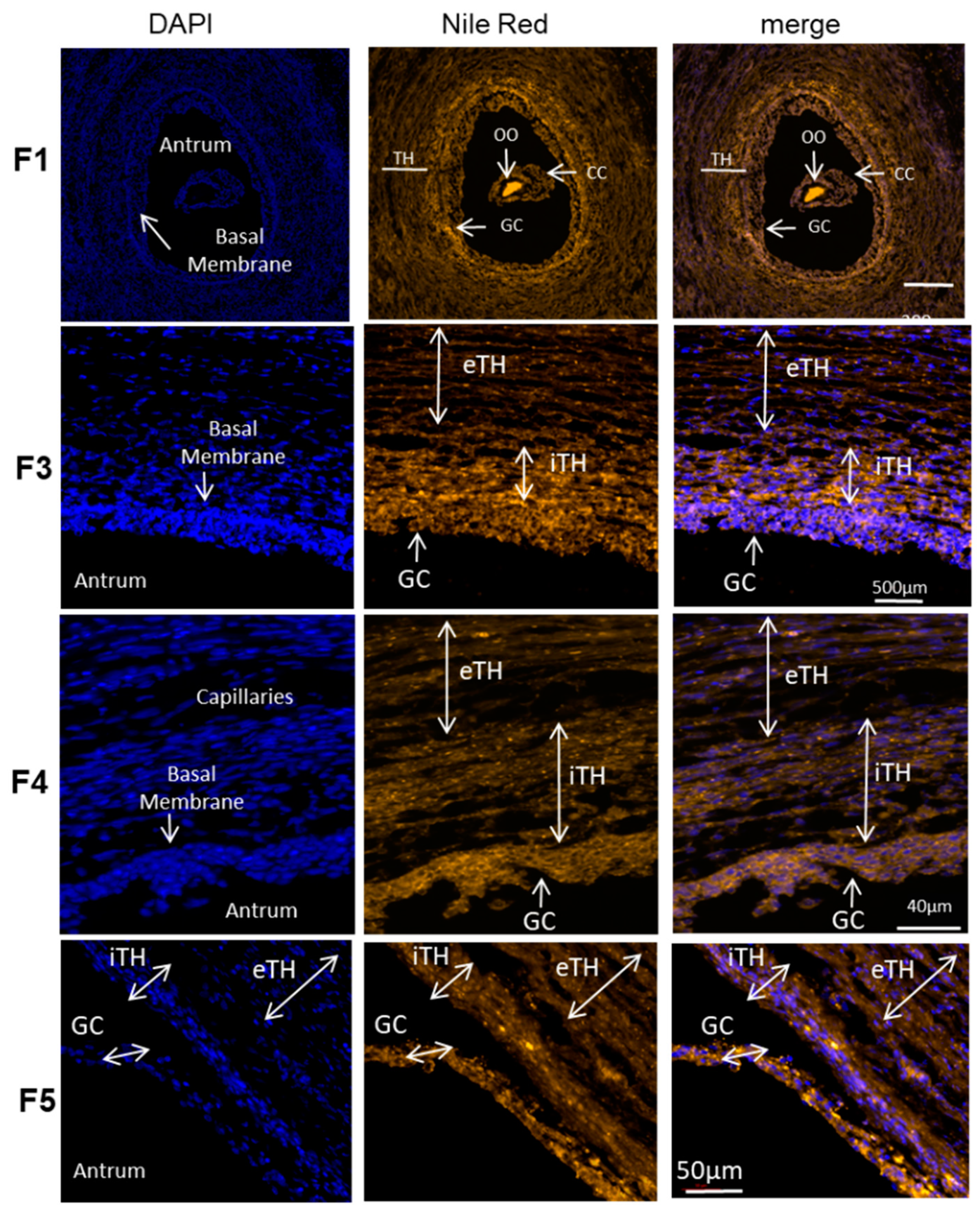
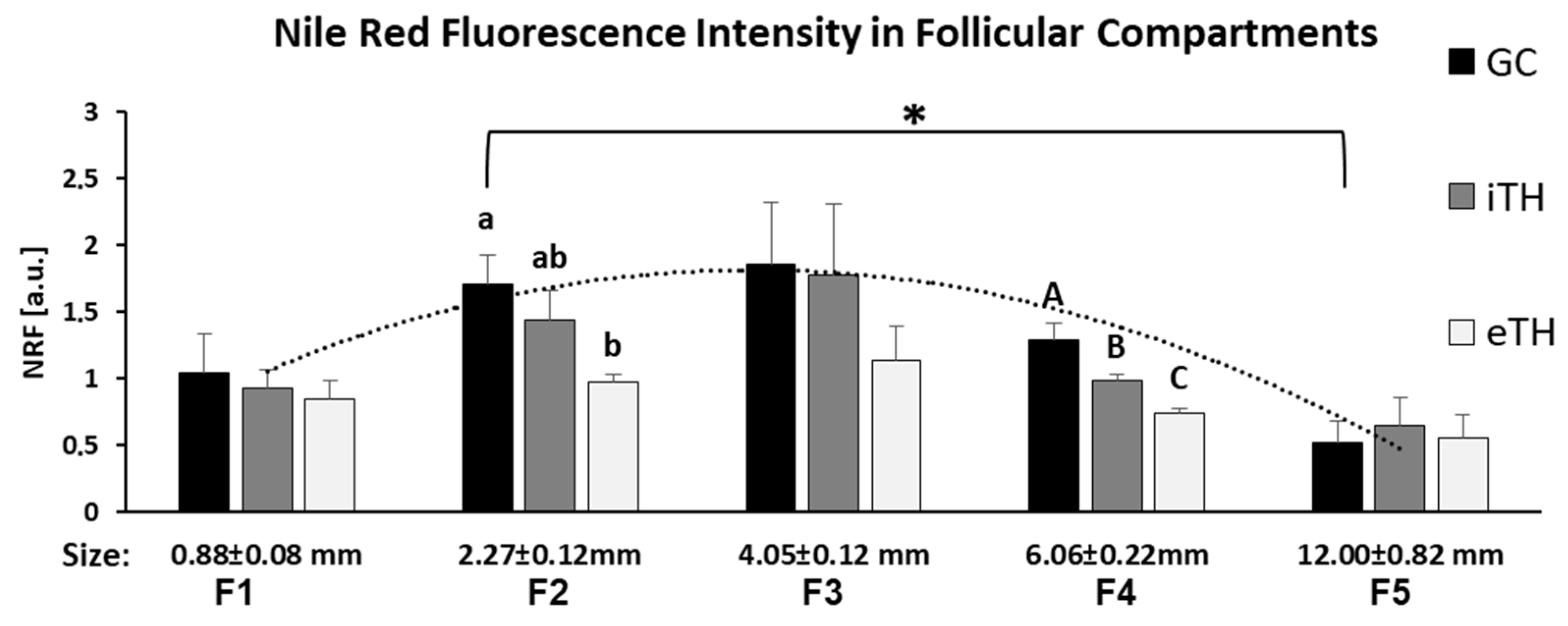
2.4. Quantification of Neutral Lipids in Ovarian Follicles of Different Size
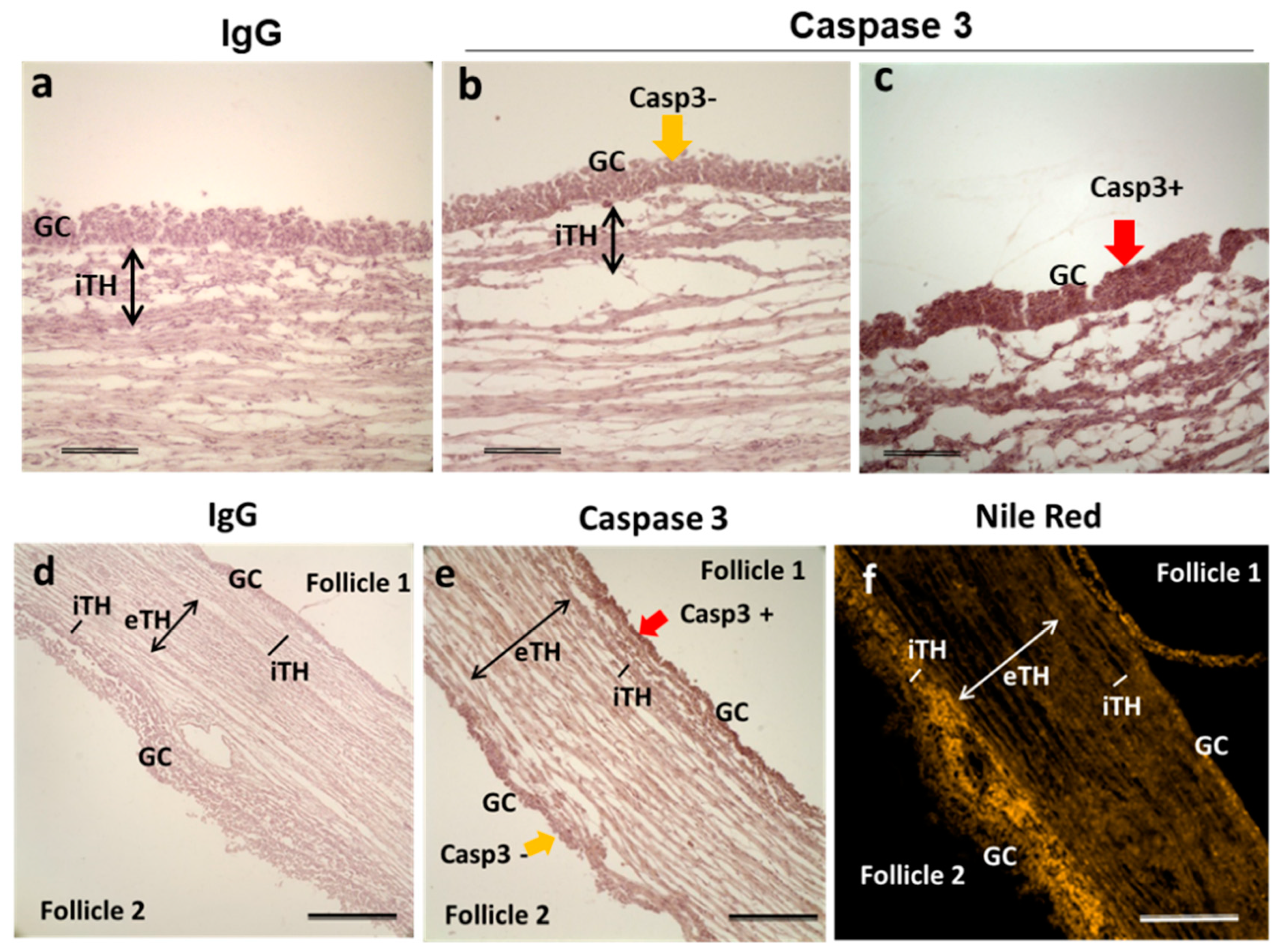
2.5. Caspase-3 and Lipid Accumulation in Bovine Follicular Cells
3. Discussion
3.1. MSI Mapping of Lipids Within Ovarian Section Revealed Difference Between the Antral Follicles
3.2. Lipid Fingerprints Changes along Follicle Growth Are Specific to Follicular Compartment
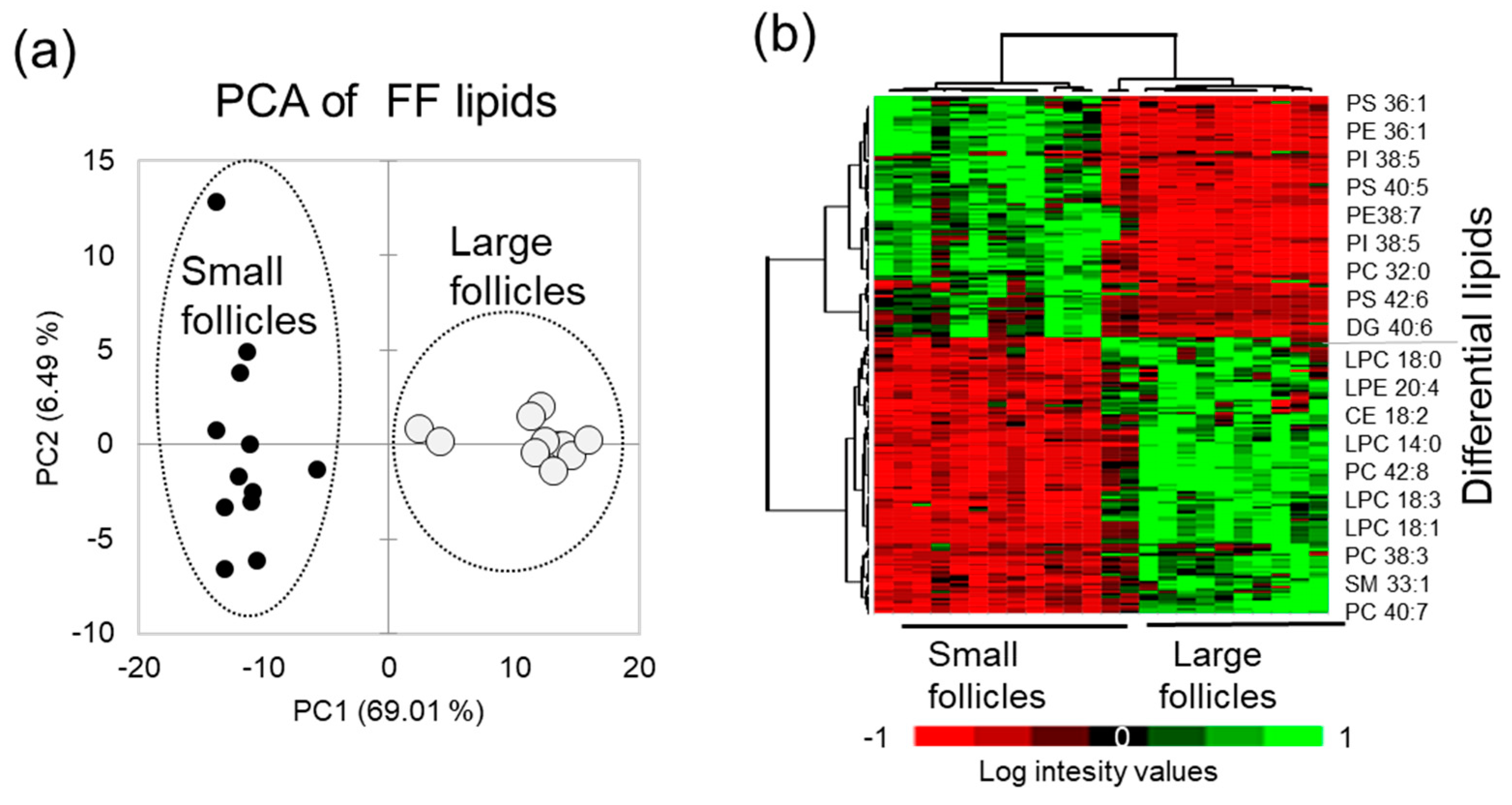
3.3. Follicular Fluid Lipid Composition is the Most Different Compartment between the Large and Small Follicles
3.4. Lipid Changes in Granulosa Cells During Follicle Growth Impact Follicular Fluid Composition
3.5. Minor Lipid Changes in the Oocyte Contrasted to Lipid Modulations in Surrounding Cumulus Cells During Follicle Growth
3.6. Lipid Changes in Theca Cells
3.7. Lipid Accumulation in Follicular Cells Associated with Apoptotic State of the Follicles
4. Materials and Methods
4.1. Ethics
4.2. Chemicals
4.3. Biological Materials
4.4. Mass Spectrometry Imaging by MALDI-TOF MS
4.5. MALDI-TOF MS Profiling of Lipids in Follicular Cells and Fluid
4.6. Lipid Characterization
4.7. Gene Expression Analysis
4.7.1. RNA Extraction and Reverse Transcription
4.7.2. Real Time PCR Analysis
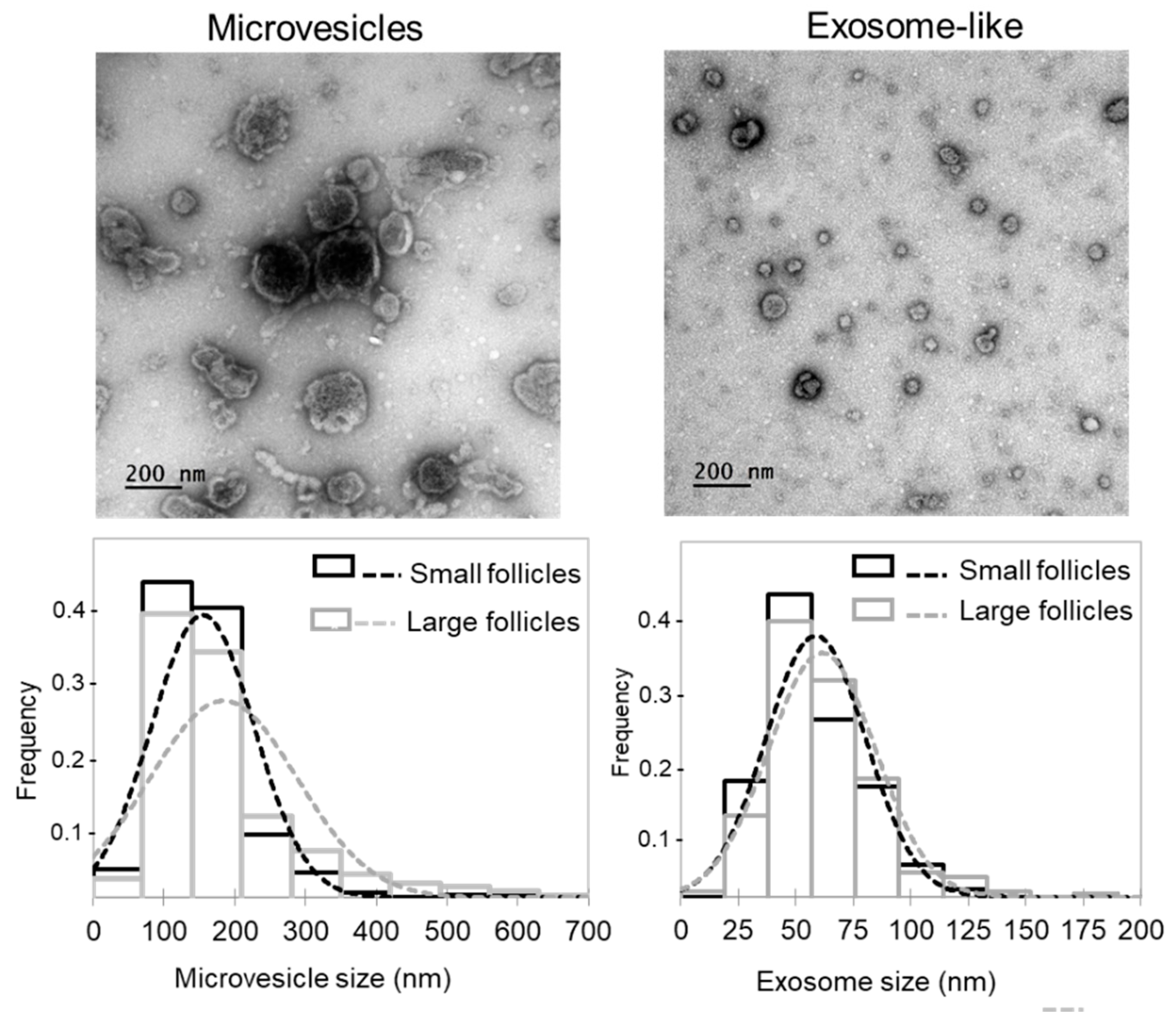
4.8. Isolation of Extracellular Vesicles from Follicular Fluid
4.9. Transmission Electron Microscopy (TEM)
4.10. Nile Red Fluorescence (NRF)
4.11. Immunohistochemistry on Ovary Sections
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ACACA | Acetyl-CoA Carboxylase Alpha |
| ACADVL | Acyl-CoA Dehydrogenase Very Long Chain |
| ACOT9 | Acyl-CoA Thioesterase 9 |
| APOA1 | Apolipoprotein A1 |
| ATT | 6-Aza-2-thiothymine matrix |
| Casp3 | Caspase-3 |
| cAMP | Cyclic adenosine monophosphate |
| CC | Cumulus cells |
| CD36 | CD36 Molecule (Thrombospondin Receptor, Fatty Acid Translocase) |
| Cer | Ceramide |
| CE | Cholesteryl esters |
| CHCA | α-Cyano-4-hydroxycinnamic acid matrix |
| CPT2 | Carnitine Palmitoyltranferase 2 |
| CYP11A1 | Cytochrome P450 Family 11 Subfamily A Member 1 |
| DAPI | 4′,6-Diamidine-2′-phenylindole dihydrochloride |
| DG | Diacylglycerol |
| DHAP | 2,5-dihydroxyacetophenone matrix |
| DHB | 2,5-Dihydroxybenzoic acid matrix |
| EV | Extracellular vesicle |
| FA | Fatty acyl |
| FABP3 | Fatty acid binding protein 3 |
| FABP5 | Fatty acid binding protein 5 |
| FAO | Fatty acid oxidation |
| FF | Follicular fluid |
| ffEVs | Extracellular vesicles isolated from FF |
| GC | Granulosa cells |
| GLUT1 | Glucose transporter 1 |
| GPX4 | Glutathione peroxidase 4 |
| HABA | 2-(4′-Hydroxybenzeneazo) benzoic acid matrix |
| HADHA | Hydroxyacyl-CoA Dehydrogenase Trifunctional Multienzyme Complex Subunit α |
| HDL | High Density Lipoprotein |
| HSD3B1 | Hydroxy-Delta-5-Steroid Dehydrogenase, 3 Beta- and Steroid Delta-Isomerase 1 |
| LF | Large Follicle |
| LPC | Lyso-phosphatidylcholines |
| LPE | Lyso-phosphatidylethanolamines |
| LPL | Lipoprotein Lipase |
| m/z | Ion Mass to Ion Charge number ratio |
| MALDI-TOF | Matrix Assisted Laser Desorption Ionization -Time of Flight |
| MBT | 2-Mercaptobenzothiazole matrix |
| MS | Mass Spectrometry |
| MSI | Mass Spectrometry Imaging |
| MV | Microvesicle |
| NRF | Nile Red Fluorescence |
| OO | Oocyte |
| COC | Cumulus-oocyte complex |
| PC | Phosphatidylcholines |
| PE | Phosphatidylethanolamines |
| PI | Phosphatidylinositols |
| PI3K | Phosphatidylinositol-3-Kinase |
| PLIN2 | Perilipin 2 |
| PON1 | Paraxoxonase 1 |
| PS | Phosphatidylserines |
| ROI | Region of Interest |
| SCARB1 | Scavenger Receptor Class B Member 1 |
| SCARB2 | Scavenger Receptor Class B Member 2 |
| SCD | Stearoyl-CoA desaturase |
| SF | Small Follicles |
| SM | Sphingomyelins |
| SOD | Superoxide dismutase |
| TG | Triacylglycerol |
| TH | Theca cells |
| eTH | External Theca |
| iTH | Internal Theca |
| TRIB2 | Tribbles homolog 2 |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Description | Primer’s Sequence (5′-3′) | Efficiency% (E) | |
|---|---|---|---|---|---|
| ACACA | NM_174224 | Acetyl-CoA Carboxylase Alpha | Fw Rev | TGCTTCCCATTTGCCATC CTGCCATCCTCACGACCT | 103.2% (2.03) |
| ACADVL | MN_174494 | Acyl-CoA Dehydrogenase, Very Long Chain | Fw Rev | CATCGTCCACCAGGAACTGAG GAACCACCACCATGGCATAGA | 103.129% (2.03) |
| ACOT9 | NM_001034560.1 | Acyl-CoA Thioesterase 9 | Fw Rev | GTGAGGTGGCCTCTCTTCAG GGGCTTCAACAAGAGCAGTC | 114.5% (2.14) |
| APOA1 | NM_174242.3 | Apolipoprotein A1 | Fw Rev | TTTGGGAAAACAGCTCAACC CAGGTCCTTGTGCATCTCCT | 84.1% (1.84) |
| CD36 | NM_001044622.1 | CD36-molecule (Thrombospondin Receptor) | Fw Rev | TTGGCCTATGAACCGTTTACT CGTTCTGAAGTTGCCAAGCA | 119.6% (2.20) |
| CPT2 | NC_037330.1 | Carnitine Palmitoyltransferase 2 | Fw Rev | AACCGCAAGTCTGGAGAAGA TGACCGTAACCACCATGAGA | 105.1% (2.05) |
| CYP11A1 | NM_176644.1 | Cytochrome P450, Family 11, Subfamily A, Polypeptide 1 | Fw Rev | CGGAAAGTTTGTAGGGGACA ACGTTGAGCAGAGGGACACT | 100.5% (2.00) |
| FABP3 | NM_174313 | Fatty acid binding protein 3 | Fw Rev | ATCGTGACGCTGGATGGCGG GCCGAGTCCAGGAGTAGCCCA | 99.8% (1.99) |
| FABP5 | NM_001129805.1 | Fatty acid binding protein 5 | Fw Rev | ACGTGTCGAAAGAAGGAGGT ACATCCCAAAGAGCATATCGTA | 100.7% (2.00) |
| GAPDH | NM_001034034 | Glyceraldehyde 3 phosphate dehydrogenase | Fw Rev | TTCAACGGCACAGTCAAGG ACATACTCAGCACCAGCATCAC | 100.1% (2.00) |
| GPX4 | NM_174770 | Glutathione Peroxidase 4 | Fw Rev | CGATACGCCGAGTGTGGTTTAC ACAGCCGTTCTTGTCAATGAGG | 101% (2.01) |
| HADHA | NM_174335 | Hydroxyacyl-CoA Dehydrogenase Trifunctional Multienzyme Complex Subunit Alpha | Fw Rev | CTGTACGGGGCACAGAAAAT TGCCAGAGTTCGTCTTCCTT | 102.2% (2.02) |
| HSD3B1 | NM_174343 | Hydroxy-Delta-5-Steroid Dehydrogenase, 3 Beta- and Steroid Delta-Isomerase 1 | Fw Rev | TCCACACCAGCACCATAGAA AAGGTGCCACCATTTTTCAG | 102.2% (2.02) |
| LPL | NM_001075120 | Lipoprotein lipase | Fw Rev | GGGTTTTGAGCAAGGGTACA GCCACAATGACCTTTCCAGT | 111.1% (2.11) |
| PLIN2 | NM_173980 | Perilipin 2 | Fw Rev | ACAACACACCCCTCAACTGG CTGCCTGCCTACTTCAGACC | 95.0% (1.95) |
| RPL19 | BC102223 | Ribosomal protein L19 | Fw Rev | AATCGCCAATGCCAACTC CCCTTTCGCTTACCTATACC | 101.0% (2.01) |
| RPS9 | BC148016 | Ribosomial protein S9 | Fw Rev | GGAGACCCTTCGAGAAGTCC GGGCATTACCTTCGAACAGA | 95.8% (1.96) |
| SCARB1 | NM_174597.2 | Scavenger Receptor Class B, member 1 or CD36 Antigen-Like | Fw Rev | ATGCCCAGTATGTGCTCCTT AAGAAGCGGGGTGTAGGG | 100.1% (2.00) |
| SCARB2 | NM_001102153.1 | Scavenger Receptor Class B, member 2 | Fw Rev | TCCAGGCTTCTCCCACTAGA TTCTGGGCAGCATTCTCTTT | 109.6% (2.10) |
| SLC2A1 (GLUT1) | NM_174602 | Solute Carrier Family 2 Member 1 (facilitative glucose transporter) | Fw Rev | CTGATCCTGGGTCGCTTCAT ACGTACATGGGCACAAAACCA | 100% (2.00) |
| TRIB2 | NM_178317.3 | Tribbles homolog 2 (Drosophila) | Fw Rev | GTGGCATGTAGTGCAGACC ACAGGACAAAGCACCAGAG | 100.2% (2.00) |
References
- Dalbies-Tran, R.; Cadoret, V.; Desmarchais, A.; Elis, S.; Maillard, V.; Monget, P.; Monniaux, D.; Reynaud, K.; Saint-Dizier, M.; Uzbekova, S.V. A Comparative Analysis of Oocyte Development in Mammals. Cells 2020, 9, 1002. [Google Scholar] [CrossRef] [Green Version]
- Young, J.M.; McNeilly, A.S. Theca: The forgotten cell of the ovarian follicle. Reproduction 2010, 140, 489–504. [Google Scholar] [CrossRef] [Green Version]
- Irving-Rodgers, H.F.; Harland, M.L.; Sullivan, T.; Rodgers, R.J. Studies of granulosa cell maturation in dominant and subordinate bovine follicles: Novel extracellular matrix focimatrix is co-ordinately regulated with cholesterol side-chain cleavage CYP11A1. Reproduction 2009, 137, 825–834. [Google Scholar] [CrossRef]
- Fernández, E.C.; Picton, H.M.; Dumollard, R. Metabolism throughout follicle and oocyte development in mammals. Int. J. Dev. Biol. 2012, 56, 799–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunning, K.R.; Russell, D.L.; Robker, R.L. Lipids and oocyte developmental competence: The role of fatty acids and β-oxidation. Reproduction 2014, 148, R15–R27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paczkowski, M.; Silva, E.; Schoolcraft, W.B.; Krisher, R.L. Comparative Importance of Fatty Acid Beta-Oxidation to Nuclear Maturation, Gene Expression, and Glucose Metabolism in Mouse, Bovine, and Porcine Cumulus Oocyte Complexes1. Biol. Reprod. 2013, 88, 111. [Google Scholar] [CrossRef] [PubMed]
- Sturmey, R.; Reis, A.; Leese, H.; McEvoy, T.; Sturmey, R.G. Role of Fatty Acids in Energy Provision During Oocyte Maturation and Early Embryo Development. Reprod. Domest. Anim. 2009, 44, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, H.F.; Goodman, J.M. The life cycle of lipid droplets. Curr. Opin. Cell Biol. 2015, 33, 119–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahli, W.; Michalik, L. PPARs at the crossroads of lipid signaling and inflammation. Trends Endocrinol. Metab. 2012, 23, 351–363. [Google Scholar] [CrossRef] [PubMed]
- Lucki, N.C.; Sewer, M.B. Multiple Roles for Sphingolipids in Steroid Hormone Biosynthesis. Subcell. Biochem. 2008, 49, 387–412. [Google Scholar] [CrossRef] [Green Version]
- Hillier, S.G.; Whitelaw, P.F.; Smyth, D. Follicular oestrogen synthesis: The ‘two-cell, two-gonadotrophin’ model revisited. Mol. Cell. Endocrinol. 1994, 100, 51–54. [Google Scholar] [CrossRef]
- Bao, B.; Garverick, H.A. Expression of steroidogenic enzyme and gonadotropin receptor genes in bovine follicles during ovarian follicular waves: A review. J. Anim. Sci. 1998, 76, 1903–1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertevello, P.S.; Teixeira-Gomes, A.-P.; Seyer, A.; Carvalho, A.V.; Labas, V.; Blache, M.-C.; Banliat, C.; Cordeiro, L.A.V.; Duranthon, V.; Papillier, P.; et al. Lipid Identification and Transcriptional Analysis of Controlling Enzymes in Bovine Ovarian Follicle. Int. J. Mol. Sci. 2018, 19, 3261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donadeu, F.X.; Fahiminiya, S.; Esteves, C.L.; Nadaf, J.; Miedzinska, K.; McNeilly, A.S.; Waddington, D.; Gérard, N. Transcriptome Profiling of Granulosa and Theca Cells During Dominant Follicle Development in the Horse1. Biol. Reprod. 2014, 91, 111. [Google Scholar] [CrossRef]
- Hatzirodos, N.; Hummitzsch, K.; Irving-Rodgers, H.F.; Rodgers, R.J. Transcriptome Comparisons Identify New Cell Markers for Theca Interna and Granulosa Cells from Small and Large Antral Ovarian Follicles. PLoS ONE 2015, 10, e0119800. [Google Scholar] [CrossRef] [Green Version]
- Skinner, M.K.; Schmidt, M.; Savenkova, M.I.; Sadler-Riggleman, I.; Nilsson, E.E. Regulation of granulosa and theca cell transcriptomes during ovarian antral follicle development. Mol. Reprod. Dev. 2008, 75, 1457–1472. [Google Scholar] [CrossRef] [Green Version]
- Girard, A.; Dufort, I.; Douville, G.; Sirard, M.-A. Global gene expression in granulosa cells of growing, plateau and atretic dominant follicles in cattle. Reprod. Biol. Endocrinol. 2015, 13, 17. [Google Scholar] [CrossRef] [Green Version]
- Assidi, M.; Dieleman, S.J.; Sirard, M.-A. Cumulus cell gene expression following the LH surge in bovine preovulatory follicles: Potential early markers of oocyte competence. Reproduction 2010, 140, 835–852. [Google Scholar] [CrossRef] [Green Version]
- Labrecque, R.; Fournier, E.; Sirard, M.-A. Transcriptome analysis of bovine oocytes from distinct follicle sizes: Insights from correlation network analysis. Mol. Reprod. Dev. 2016, 83, 558–569. [Google Scholar] [CrossRef]
- Aardema, H.; Vos, P.L.A.M.; Gadella, B.M. Cumulus cells protect the oocyte against saturated free fatty acids. Anim. Reprod. 2018, 15, 737–750. [Google Scholar] [CrossRef]
- Sanchez-Lazo, L.; Brisard, D.; Elis, S.; Maillard, V.; Uzbekov, R.; Labas, V.; Desmarchais, A.; Papillier, P.; Monget, P.; Uzbekova, S. Fatty Acid Synthesis and Oxidation in Cumulus Cells Support Oocyte Maturation in Bovine. Mol. Endocrinol. 2014, 28, 1502–1521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutton-McDowall, M.; Feil, D.; Robker, R.L.; Thompson, J.G.; Dunning, K.R. Utilization of endogenous fatty acid stores for energy production in bovine preimplantation embryos. Theriogenology 2012, 77, 1632–1641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blondin, P. Oocyte and follicular morphology as determining characteristics for developmental competence in bovine oocytes. Mol. Reprod. Dev. 1995, 41, 54–62. [Google Scholar] [CrossRef]
- Lonergan, P.; Monaghan, P.; Rizos, D.; Boland, M.P.; Gordon, I. Effect of follicle size on bovine oocyte quality and developmental competence following maturation, fertilization, and culture in vitro. Mol. Reprod. Dev. 1994, 37, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Jeong, W.; Cho, S.; Lee, H.; Deb, G.; Lee, Y.; Kwon, T.; Kong, I. Effect of cytoplasmic lipid content on in vitro developmental efficiency of bovine IVP embryos. Theriogenology 2009, 72, 584–589. [Google Scholar] [CrossRef]
- Nagano, M. Acquisition of developmental competence and in vitro growth culture of bovine oocytes. J. Reprod. Dev. 2019, 65, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Oseikria, M.; Elis, S.; Maillard, V.; Corbin, E.; Uzbekova, S. N-3 polyunsaturated fatty acid DHA during IVM affected oocyte developmental competence in cattle. Theriogenology 2016, 85, 1625–1634. [Google Scholar] [CrossRef]
- Elis, S.; Oseikria, M.; Carvalho, A.V.; Bertevello, P.S.; Corbin, E.; Teixeira-Gomes, A.-P.; Lecardonnel, J.; Archilla, C.; Duranthon, V.; Labas, V.; et al. Docosahexaenoic acid mechanisms of action on the bovine oocyte-cumulus complex. J. Ovarian Res. 2017, 10, 74. [Google Scholar] [CrossRef]
- Freret, S.; Oseikria, M.; Le Bourhis, D.; Desmarchais, A.; Briant, E.; Desnoes, O.; Dupont, M.; Le Berre, L.; Ghazouani, O.; Bertevello, P.S.; et al. Effects of a n-3 PUFA enriched diet on embryo production in dairy cows. Reproduction 2019, 158, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Leroy, J.; Vanholder, T.; Delanghe, J.; Opsomer, G.; Van Soom, A.; Bols, P.; De Kruif, A. Metabolite and ionic composition of follicular fluid from different-sized follicles and their relationship to serum concentrations in dairy cows. Anim. Reprod. Sci. 2004, 80, 201–211. [Google Scholar] [CrossRef]
- Hatzirodos, N.; Rodgers, R.J.; Hummitzsch, K.; Harland, M.L.; Morris, S.E.; Rodgers, R.J. Transcriptome profiling of granulosa cells of bovine ovarian follicles during growth from small to large antral sizes. BMC Genom. 2014, 15, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatzirodos, N.; Hummitzsch, K.; Irving-Rodgers, H.F.; Rodgers, R.J. Transcriptome Profiling of the Theca Interna in Transition from Small to Large Antral Ovarian Follicles. PLoS ONE 2014, 9, e97489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Théry, C.; Witwer, K.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenwick, M.A.; Hurst, P.R. Immunohistochemical localization of active caspase-3 in the mouse ovary: Growth and atresia of small follicles. Reproduction 2002, 124, 659–665. [Google Scholar] [CrossRef]
- Glamočlija, V.; Vilović, K.; Saraga-Babić, M.; Baranović, A.; Sapunar, D. Apoptosis and active caspase-3 expression in human granulosa cells. Fertil. Steril. 2005, 83, 426–431. [Google Scholar] [CrossRef]
- Uzbekova, S.; Elis, S.; Teixeira-Gomes, A.-P.; Desmarchais, A.; Maillard, V.; Labas, V. MALDI Mass Spectrometry Imaging of Lipids and Gene Expression Reveals Differences in Fatty Acid Metabolism between Follicular Compartments in Porcine Ovaries. Biology 2015, 4, 216–236. [Google Scholar] [CrossRef] [Green Version]
- Cordeiro, F.B.; Jarmusch, A.K.; León, M.; Ferreira, C.R.; Pirro, V.; Eberlin, L.S.; Hallett, J.; Miglino, M.A.; Cooks, R.G. Mammalian ovarian lipid distributions by desorption electrospray ionization-mass spectrometry (DESI-MS) imaging. Anal. Bioanal. Chem. 2020, 412, 1251–1262. [Google Scholar] [CrossRef]
- Campbell, D.I.; Ferreira, C.R.; Eberlin, L.S.; Cooks, R.G. Improved spatial resolution in the imaging of biological tissue using desorption electrospray ionization. Anal. Bioanal. Chem. 2012, 404, 389–398. [Google Scholar] [CrossRef]
- Kruip, T.; Dieleman, S. Steroid hormone concentrations in the fluid of bovine follicles relative to size, quality and stage of the oestrus cycle. Theriogenology 1985, 24, 395–408. [Google Scholar] [CrossRef]
- Brantmeier, S.; Grummer, R.; Ax, R. Concentrations of High Density Lipoproteins Vary Among Follicular Sizes in the Bovine. J. Dairy Sci. 1987, 70, 2145–2149. [Google Scholar] [CrossRef]
- Hung, W.-T.; Hong, X.; Christenson, L.K.; McGinnis, L.K. Extracellular Vesicles from Bovine Follicular Fluid Support Cumulus Expansion1. Biol. Reprod. 2015, 93, 117. [Google Scholar] [CrossRef] [PubMed]
- Skotland, T.; Sandvig, K.; Llorente, A. Lipids in exosomes: Current knowledge and the way forward. Prog. Lipid Res. 2017, 66, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Navakanitworakul, R.; Hung, W.-T.; Gunewardena, S.; Davis, J.S.; Chotigeat, W.; Christenson, L.K. Characterization and Small RNA Content of Extracellular Vesicles in Follicular Fluid of Developing Bovine Antral Follicles. Sci. Rep. 2016, 6, 25486. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Kuang, Y.; Zhou, L.; Yu, B.; Chen, Q.; Fu, Y.; Yan, Z.; Guo, H.; Lyu, Q.; Xie, J.; et al. Lipidomic Components Alterations of Human Follicular Fluid Reveal the Relevance of Improving Clinical Outcomes in Women Using Progestin-Primed Ovarian Stimulation Compared to Short-Term Protocol. Med. Sci. Monit. 2018, 24, 3357–3365. [Google Scholar] [CrossRef]
- Lapidot, T.; Graff, J.M. Form(ul)ation of adipocytes by lipids. Adipocyte 2017, 6, 176–186. [Google Scholar] [CrossRef] [Green Version]
- Prokazova, N.V.; Zvezdina, N.D.; Korotaeva, A.A. Effect of lysophosphatidylcholine on transmembrane signal transduction. Biochem. Mosc. 1998, 63, 31–37. [Google Scholar]
- Lauber, K.; Bohn, E.; Kröber, S.M.; Xiao, Y.-J.; Blumenthal, S.G.; Lindemann, R.K.; Marini, P.; Wiedig, C.; Zobywalski, A.; Baksh, S.; et al. Apoptotic Cells Induce Migration of Phagocytes via Caspase-3-Mediated Release of a Lipid Attraction Signal. Cell 2003, 113, 717–730. [Google Scholar] [CrossRef] [Green Version]
- McNatty, K.; Heath, D.A.; Henderson, K.M.; Lun, S.; Hurst, P.R.; Ellis, L.M.; Montgomery, G.W.; Morrison, L.; Thurley, D.C. Some aspects of thecal and granulosa cell function during follicular development in the bovine ovary. Reproduction 1984, 72, 39–53. [Google Scholar] [CrossRef] [Green Version]
- Serna, J.; García-Seisdedos, D.; Alcázar, A.; Lasunción, M.A.; Busto, R.; Pastor, Ó. Quantitative lipidomic analysis of plasma and plasma lipoproteins using MALDI-TOF mass spectrometry. Chem. Phys. Lipids 2015, 189, 7–18. [Google Scholar] [CrossRef]
- Drzazga, A.; Sowińska, A.; Koziołkiewicz, M. Lysophosphatidylcholine and lysophosphatidylinosiol—Novel promissing signaling molecules and their possible therapeutic activity. Acta Pol. Pharm. 2015, 71, 887–899. [Google Scholar]
- Jaspard, B.; Fournier, N.; Vieitez, G.; Atger, V.; Barbaras, R.; Vieu, C.; Manent, J.; Chap, H.; Perret, B.; Collet, X. Structural and Functional Comparison of HDL From Homologous Human Plasma and Follicular Fluid. Arter. Thromb. Vasc. Biol. 1997, 17, 1605–1613. [Google Scholar] [CrossRef] [PubMed]
- Dadarwal, D.; Adams, G.P.; Maddox-Hyttel, P.; Brogliatti, G.M.; Caldwell, S.; Singh, J. Organelle reorganization in bovine oocytes during dominant follicle growth and regression. Reprod. Biol. Endocrinol. 2015, 13, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imai, H.; Matsuoka, M.; Kumagai, T.; Sakamoto, T.; Koumura, T. Lipid Peroxidation-Dependent Cell Death Regulated by GPx4 and Ferroptosis; Springer: Berlin/Heidelberg, Germany, 2016; Volume 403, pp. 143–170. [Google Scholar]
- Roy, R.; Bélanger, A. Formation of lipoidal steroids in follicular fluid. J. Steroid Biochem. 1989, 33, 257–262. [Google Scholar] [CrossRef]
- Tokumura, A.; Miyake, M.; Nishioka, Y.; Yamano, S.; Aono, T.; Fukuzawa, K. Production of lysophosphatidic acids by lysophospholipase D in human follicular fluids of In vitro fertilization patients. Biol. Reprod. 1999, 61, 195–199. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Bloom, M.S.; Fujimoto, V.Y.; Browne, R.W. Associations between PON1 enzyme activities in human ovarian follicular fluid and serum specimens. PLoS ONE 2017, 12, e0172193. [Google Scholar] [CrossRef]
- Cigliano, L.; Spagnuolo, M.S.; Dale, B.; Balestrieri, M.; Abrescia, P. Estradiol esterification in the human preovulatory follicle. Steroids 2001, 66, 889–896. [Google Scholar] [CrossRef]
- Bausenwein, J.; Serke, H.; Eberle, K.; Hirrlinger, J.; Jogschies, P.; Hmeidan, F.A.; Blumenauer, V.; Spanel-Borowski, K.; Abu Hmeidan, F. Elevated levels of oxidized low-density lipoprotein and of catalase activity in follicular fluid of obese women. Mol. Hum. Reprod. 2009, 16, 117–124. [Google Scholar] [CrossRef] [Green Version]
- Combelles, C.M.H.; Holick, E.A.; Paolella, L.J.; Walker, D.C.; Wu, Q. Profiling of superoxide dismutase isoenzymes in compartments of the developing bovine antral follicles. Reproduction 2010, 139, 871–881. [Google Scholar] [CrossRef] [Green Version]
- Schneider, A.; Absalon-Medina, V.; Esposito, G.; Corrêa, M.; Butler, W.R. Paraoxonase (PON) 1, 2 and 3 Expression in Granulosa Cells and PON1 Activity in Follicular Fluid of Dairy Cows. Reprod. Domest. Anim. 2013, 48, 989–994. [Google Scholar] [CrossRef]
- Monniaux, D. Driving folliculogenesis by the oocyte-somatic cell dialog: Lessons from genetic models. Theriogenology 2016, 86, 41–53. [Google Scholar] [CrossRef]
- Hao, Q.; Zhu, Z.; Xu, D.; Liu, W.; Lyu, L.; Li, P. Proteomic characterization of bovine granulosa cells in dominant and subordinate follicles. Hereditas 2019, 156, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Falkenburger, B.; Jensen, J.B.; Dickson, E.J.; Suh, B.-C.; Hille, B. Phosphoinositides: Lipid regulators of membrane proteins. J. Physiol. 2010, 588, 3179–3185. [Google Scholar] [CrossRef] [PubMed]
- Sobinoff, A.; Sutherland, J.; McLaughlin, E.A. Intracellular signalling during female gametogenesis. Mol. Hum. Reprod. 2012, 19, 265–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatzirodos, N.; Hummitzsch, K.; Rodgers, R.J.; Harland, M.L.; Morris, S.E.; Rodgers, R.J. Transcriptome profiling of granulosa cells from bovine ovarian follicles during atresia. BMC Genom. 2014, 15, 40. [Google Scholar] [CrossRef] [Green Version]
- Annes, K.; Müller, D.B.; Vilela, J.A.P.; Valente, R.S.; Caetano, D.P.; Cibin, F.; Milazzotto, M.P.; Mesquita, F.S.; Belaz, K.R.A.; Eberlin, M.N.; et al. Influence of follicle size on bovine oocyte lipid composition, follicular metabolic and stress markers, embryo development and blastocyst lipid content. Reprod. Fertil. Dev. 2019, 31, 462. [Google Scholar] [CrossRef]
- Assey, R.J.; Hyttel, P.; Greve, T.; Purwantara, B. Oocyte morphology in dominant and subordinate follicles. Mol. Reprod. Dev. 1994, 37, 335–344. [Google Scholar] [CrossRef]
- Milhas, D.; Clarke, C.J.; Hannun, Y.A. Sphingomyelin metabolism at the plasma membrane: Implications for bioactive sphingolipids. FEBS Lett. 2009, 584, 1887–1894. [Google Scholar] [CrossRef] [Green Version]
- Mathias, S.; Peña, L.A.; Kolesnick, R. Signal transduction of stress via ceramide. Biochem. J. 1998, 335, 465–480. [Google Scholar] [CrossRef]
- Lolicato, F.; Brouwers, J.F.; Van De Lest, C.H.; Wubbolts, R.; Aardema, H.; Priore, P.; Roelen, B.A.; Helms, J.B.; Gadella, B.M.; De Lest, C.H.A.V. The Cumulus Cell Layer Protects the Bovine Maturing Oocyte Against Fatty Acid-Induced Lipotoxicity1. Biol. Reprod. 2015, 92, 16. [Google Scholar] [CrossRef]
- Aardema, H.; Van Tol, H.T.A.; Wubbolts, R.W.; Brouwers, J.F.H.M.; Gadella, B.M.; Roelen, B.A.J. Stearoyl-CoA desaturase activity in bovine cumulus cells protects the oocyte against saturated fatty acid stress. Biol. Reprod. 2017, 96, 982–992. [Google Scholar] [CrossRef] [Green Version]
- Humblot, P.; Holm, P.; Lonergan, P.; Wrenzycki, C.; Lequarré, A.-S.; Joly, C.G.; Herrmann, D.; Lopes, A.; Rizos, D.; Niemann, H.; et al. Effect of stage of follicular growth during superovulation on developmental competence of bovine oocytes. Theriogenology 2005, 63, 1149–1166. [Google Scholar] [CrossRef] [PubMed]
- Lequarré, A.; Vigneron, C.; Ribaucour, F.; Holm, P.; Donnay, I.; Dalbiès-Tran, R.; Callesen, H.; Mermillod, P. Influence of antral follicle size on oocyte characteristics and embryo development in the bovine. Theriogenology 2005, 63, 841–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jewgenow, K.; Heerdegen, B.; Müller, K. In vitro development of individually matured bovine oocytes in relation to follicular wall atresia. Theriogenology 1999, 51, 745–756. [Google Scholar] [CrossRef]
- Zeuner, A.; Müller, K.; Reguszynski, K.; Jewgenow, K. Apoptosis within bovine follicular cells and its effect on oocyte development during in vitro maturation. Theriogenology 2003, 59, 1421–1433. [Google Scholar] [CrossRef]
- Salhab, M.; Tosca, L.; Cabau, C.; Papillier, P.; Perreau, C.; Dupont, J.; Mermillod, P.; Uzbekova, S. Kinetics of gene expression and signaling in bovine cumulus cells throughout IVM in different mediums in relation to oocyte developmental competence, cumulus apoptosis and progesterone secretion. Theriogenology 2011, 75, 90–104. [Google Scholar] [CrossRef] [PubMed]
- Brisard, D.; Chesnel, F.; Elis, S.; Desmarchais, A.; Sánchez-Lazo, L.; Chasles, M.; Maillard, V.; Uzbekova, S. Tribbles expression in cumulus cells is related to oocyte maturation and fatty acid metabolism. J. Ovarian Res. 2014, 7, 44. [Google Scholar] [CrossRef] [Green Version]
- Demarquoy, J.; Le Borgne, F. Crosstalk between mitochondria and peroxisomes. World J. Biol. Chem. 2015, 6, 301–309. [Google Scholar] [CrossRef]
- Grevengoed, T.J.; Klett, E.L.; Coleman, R.A. Acyl-CoA metabolism and partitioning. Annu. Rev. Nutr. 2014, 34, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Tillander, V.; Nordström, E.A.; Reilly, J.; Strozyk, M.; Van Veldhoven, P.P.; Hunt, M.C.; Alexson, S.E.H. Acyl-CoA thioesterase 9 (ACOT9) in mouse may provide a novel link between fatty acid and amino acid metabolism in mitochondria. Cell. Mol. Life Sci. 2013, 71, 933–948. [Google Scholar] [CrossRef]
- Robenek, H.; Hofnagel, O.; Buers, I.; Robenek, M.J.; Troyer, D.; Severs, N.J. Adipophilin-enriched domains in the ER membrane are sites of lipid droplet biogenesis. J. Cell Sci. 2006, 119, 4215–4224. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, F.; Inoue, N.; Manabe, N.; Ohkura, S. Follicular growth and atresia in mammalian ovaries: Regulation by survival and death of granulosa cells. J. Reprod. Dev. 2012, 58, 44–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valdez, K.E.; Cuneo, S.P.; Turzillo, A.M. Regulation of apoptosis in the atresia of dominant bovine follicles of the first follicular wave following ovulation. Reproduction 2005, 130, 71–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Y.; Jin, Y.; Wu, X.; Ren, Z. PSMD1 and PSMD2 regulate HepG2 cell proliferation and apoptosis via modulating cellular lipid droplet metabolism. BMC Mol. Biol. 2019, 20, 1–20. [Google Scholar] [CrossRef]
- Dautel, S.E.; Kyle, J.E.; Clair, G.C.; Sontag, R.L.; Weitz, K.K.; Shukla, A.K.; Nguyen, S.N.; Kim, Y.-M.; Zink, E.M.; Luders, T.; et al. Lipidomics reveals dramatic lipid compositional changes in the maturing postnatal lung. Sci. Rep. 2017, 7, 40555. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.-S.; Qi, W.-T.; Guo, W.; Wang, C.-L.; Hu, Z.-B.; Li, A.-K. Genistein and daidzein induce apoptosis of colon cancer cells by inhibiting the accumulation of lipid droplets. Food Nutr. Res. 2018, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]








| Follicular Compartment | Number of Detected Peaks | Differentially Abundant Peaks, n (%) | LF vs. SF (Relative Abundance of the Peaks) | |
|---|---|---|---|---|
| Up-Regulated (n) | Down-Regulated (n) | |||
| OO | 468 | 8 (1.7%) | 2 | 6 |
| CC | 439 | 43 (9.8%) | 41 | 2 |
| FF | 375 | 208 (55.5%) | 111 | 97 |
| GC | 350 | 52 (14.9%) | 35 | 17 |
| TH | 338 | 18 (5.3%) | 15 | 3 |
| m/z | p-Value | Ratio LF vs. SF | Lipid Ion (Carbons: Unsaturation [Add]) |
|---|---|---|---|
| m/z 546.293+ | 5.84 × 107 | 6.14 | LPC 18:0 [Na]+ |
| m/z 524.310+ | 6.21 × 107 | 5.21 | LPE 20:4 [Na]+ or LPC 18:0 [H]+ |
| m/z 671.543+ | 7.21 × 105 | 3.99 | CE 18:2 [Na]+ |
| m/z 506.197+ | 1.33 × 106 | 3.90 | LPC 14:0 [K]+ |
| m/z 858.613+ | 1.02 × 104 | 3.49 | PC 42:8 [H]+ or PC 40:5 [Na]+ |
| m/z 860.595+ | 7.80 × 105 | 3.31 | PC 42:7 [H]+ or PC 40:4 [Na]+ |
| m/z 810.601+ | 2.75 × 108 | 2.82 | PC 38:4 [H]+ |
| m/z 808.592+ | 2.73 × 105 | 2.74 | PC 38:5 [H]+ |
| m/z 518.223+ | 2.88 × 104 | 2.56 | LPC 18:3 [H]+ |
| m/z 544.283+ | 4.55 × 106 | 2.46 | LPC 18:1 [Na]+ |
| m/z 788.619+ | 6.14 × 106 | 2.43 | PC 36:1 [H]+ |
| m/z 832.584+ | 1.47 × 106 | 2.39 | PC 40:7 [H]+ or PC 38:4 [Na]+ |
| m/z 812.625+ | 2.32 × 107 | 2.37 | PC 38:3 [H]+ |
| m/z 711.516+ | 1.80 × 104 | 2.36 | SM 33:1 [Na]+ |
| m/z 669.576+ | 5.15 × 105 | 2.34 | CE 18:3 [Na]+ |
| m/z 790.629+ | 2.34 × 105 | 2.07 | PC O-38:7 or PC P-38:6 [H]+ |
| m/z 716.424+ | 1.87 × 104 | 2.01 | PC 28:0 [K]+ |
| m/z 872.530+ | 1.44 × 102 | 0.49 | PC 40:6 [K]+ |
| m/z 605.487+ | 3.98 × 105 | 0.48 | DG 32:1 [K]+ |
| m/z 683.450+ | 1.44 × 103 | 0.48 | DG 38:4 [K]+ |
| m/z 502.218+ | 3.15 × 103 | 0.46 | LPC P-16:0 [Na]+ |
| m/z 826.575+ | 2.98 × 102 | 0.46 | PC 36:1 [K]+ |
| m/z 748.537+ | 6.20 × 104 | 0.43 | PC 33:0 [H]+ |
| m/z 834.607− | 9.57 × 107 | 0.43 | PS 40:6 [H]− |
| m/z 846.501+ | 2.10 × 102 | 0.41 | PC 38:5 [K]+ |
| m/z 870.506+ | 7.08 × 1011 | 0.41 | PC 40:7 [K]+ |
| m/z 794.668− | 1.21 × 109 | 0.38 | PE 40:4 [H]− |
| m/z 800.540+ | 7.12 × 1010 | 0.37 | PC 34:0 [K]+ |
| m/z 706.527+ | 1.20 × 107 | 0.33 | PC 30:0 [H]+ |
| m/z 770.517+ | 1.12 × 1012 | 0.32 | PC 32:1 [K]+ |
| m/z 762.576+ | 3.23 × 1010 | 0.31 | PC 34:0 [H]+ |
| m/z 741.510+ | 5.38 × 1010 | 0.30 | SM 34:1 [K]+ |
| m/z 769.515+ | 8.65 × 109 | 0.30 | SM 36:1 [K]+ |
| m/z 887.662− | 1.23 × 107 | 0.30 | PI 38:3 [H]− |
| m/z 707.525+ | 2.32 × 107 | 0.29 | DG 40:6 [K]+ |
| m/z 716.652− | 3.02 × 106 | 0.27 | PE 34:1 [H]− |
| m/z 750.654− | 6.63 × 106 | 0.27 | PE O-38:5 or PE P-38:4 [H]− |
| m/z 798.517+ | 1.51 × 1011 | 0.23 | PC 34:1 [K]+ |
| m/z 846.658− | 2.45 × 1011 | 0.23 | PS P-42:6 [H]− |
| m/z 772.501+ | 4.55 × 1013 | 0.21 | PC 32:0 [K]+ |
| m/z 734.551+ | 3.02 × 1012 | 0.16 | PC 32:0 [H]+ |
| m/z 774.645− | 1.45 × 1010 | 0.14 | PE O-40:7 or PE P-40:6 [H]− |
| m/z 885.652− | 9.09 × 1010 | 0.11 | PI 38:4 [H]− |
| m/z 742.641− | 1.97 × 108 | 0.10 | PE 36:2 [H]− |
| m/z 760.606− | 2.60 × 105 | 0.10 | PE 38:7 [H]− |
| m/z 836.624− | 4.67 × 108 | 0.10 | PS 40:5 [H]− |
| m/z 883.624− | 2.32 × 108 | 0.09 | PI 38:5 [H]− |
| m/z 744.664− | 8.79 × 1012 | 0.07 | PE 36:1 [H]− |
| m/z 788.660− | 1.92 × 109 | 0.02 | PS 36:1 [H]− |
| m/z | p-Value | Ratio LF vs. SF | Lipid Ion (Carbons: Unsaturation [Add]) |
|---|---|---|---|
| m/z 702.431+ | 3.66 × 102 | 0.43 | PC O-31:2 or PC P-31:1 [H]+ |
| m/z 728.456+ | 3.41 × 102 | 0.43 | PC 29:1 [K]+ |
| m/z 713.467+ | 2.28 × 102 | 0.39 | SM 32:1 [K]+ |
| m/z | p-Value | Ratio LF vs. SF | Lipid Ion (Carbons: Unsaturation [Add]) |
|---|---|---|---|
| m/z 714.352+ | 2.93 × 105 | 4.65 | PC 29:0 [Na]+ |
| m/z 734.595+ | 7.08 × 103 | 2.90 | PC 32:0 [H]+ |
| m/z 813.701+ | 8.74 × 105 | 2.78 | TG 47:1 [Na]+ |
| m/z 808.598+ | 4.51 × 104 | 2.57 | PC 38:5 [H]+ |
| m/z 828.758+ | 2.28 × 103 | 2.57 | PC 39:2 [H]+ |
| m/z 800.647+ | 7.75 × 103 | 2.49 | PC 34:0 [K]+ or PC 37:2 [H]+ |
| m/z 810.634+ | 2.96 × 104 | 2.45 | PC 38:4 [H]+ |
| m/z 824.744+ | 3.46 × 105 | 2.45 | PC O-40:4 or PC P-40:3 [H]+ |
| m/z 834.642+ | 2.54 × 105 | 2.35 | PC 40:6 [H]+ or PC 38:3 [Na]+ |
| m/z 826.754+ | 7.79 × 103 | 2.34 | PC 39:3[H]+ |
| m/z 786.626+ | 1.97 × 103 | 2.33 | PC 36:2 [H]+ or PC 33:0 [K]+ |
| m/z 836.677+ | 3.26 × 104 | 2.13 | PC 40:5 [H]+ or PC 38:2 [Na]+ |
| m/z 725.533+ | 9.83 × 103 | 2.06 | SM 34:1 [Na]+ |
| m/z 748.593+ | 6.23 × 105 | 2.06 | PC 33:0 [H]+ |
| m/z 788.634+ | 1.28 × 103 | 2.06 | PC 36:1 [H]+ |
| m/z 802.659+ | 3.46 × 102 | 2.02 | PC 37:1 [H]+ |
| m/z 744.522+ | 1.57 × 103 | 2.01 | PC O-31:0 [K]+ |
| m/z 721.406+ | 2.38 × 102 | 2.00 | SM 34:3 [Na]+ |
| m/z | p-Value | Ratio LF vs. SF | Lipid Ion (Carbons: Unsaturation [Add]) |
|---|---|---|---|
| m/z 861.431− | 5.83 × 103 | 2.81 | PI 36:2 [H]− |
| m/z 844.546+ | 1.03 × 104 | 0.50 | PC 38:6 [K]+ |
| m/z 560.274+ | 3.70 × 103 | 0.49 | LPC 18:1 [K]+ |
| m/z 863.704+ | 2.54 × 102 | 0.47 | SM 44:2 [Na]+ |
| m/z 848.559+ | 1.01 × 104 | 0.46 | PC 38:4 [K]+ |
| m/z 722.463− | 8.16 × 104 | 0.45 | PE O-36:5 or PE P-36:4 [H]− |
| m/z 582.303+ | 1.22 × 102 | 0.40 | LPC 20:4 [K]+ |
| m/z 534.302+ | 3.84 × 103 | 0.38 | LPC 16:0 [K]+ |
| m/z 428.379+ | 8.21 × 104 | 0.37 | C18:0 Carnitine [H]+ |
| m/z | p-Value | Ratio LF vs. SF | Lipid Ion (Carbons: Unsaturation [Add]) |
|---|---|---|---|
| m/z 667.572+ | 2.20 | 2.2 | DG 38:4 [Na]+ |
| m/z 683.481+ | 2.14 | 2.14 | DG 38:4 [K]+ |
| m/z 760.544− | 2.08 | 2.08 | PE 38:7 [H]− |
| m/z 714.550− | 2.02 | 2.02 | PE 34:2 [H]− |
| Theca Cells | Granulosa Cells | |||
|---|---|---|---|---|
| Gene | Small Follicles | Large Follicles | Small Follicles | Large Follicles |
| ACACA | 1579.6 ± 373.7 | 1044.4 ± 246.6 | 986.4 ± 309.1 | 906.6 ± 325.8 |
| ACADVL | 1463.2 ± 674.1 | 4280.2 ± 3191.1 | 1692.7 ± 816.9 | 4716.8 ± 1635.5 |
| ACOT9 | 11.8 ± 2.9 | 8.1 ± 3.5 | 0.88 ± 0.2 | 1.04 ± 0.2 |
| APOA1 | 0.42 ± 0.2 | 0.31 ± 0.1 | 0.22 ± 0.04 | 0.42 ± 0.2 |
| CD36 | 253.3 ± 69.5 | 168.9 ± 89.6 | 2302.1 ± 979.9 | 1820.9 ± 778.8 |
| CPT2 | 12774.0 ± 4910.7 | 10332.6 ± 5231.8 | 8687.4 ± 2408.4 | 8464.22 ± 3618.4 |
| CYP11 | 1740.53 ± 1205.2 | 5463.51 ± 2995.6 | 1583.9 ± 547.3 | 9277.1 ± 3107.9 |
| FABP3 | 3346.1 ± 1456.9 | 1573.1 ± 298.9 | 859.8 ± 241.7 | 1629.3 ± 540.7 |
| FABP5 | 6860.6 ± 2342.1 | 8872.8 ± 4077.2 | 547.9 ± 272.5 | 1646.8 ± 1043.2 |
| GLUT1 | 965.2 ± 255.6 | 3198.5 ± 2068.3 | 13126.3 ± 5844.3 | 44136.2 ± 23953.6 |
| GPX4 | 2927.1 ± 1100.8 | 12189.2 ± 7173.6 | 11672.5 ± 3132.3 | 34370.6 ± 8799.6 |
| HADHA | 852.1 ± 236.7 | 883.7 ± 156.9 | 885.1 ± 137.7 | 1229.3 ± 130.3 |
| HSD3B1 | 1060.6 ± 331.9 | 566.9 ± 189.5 | 885.3 ± 291.9 | 1387.5 ± 330.5 |
| LPL | 164762.8 ± 57444.5 | 149005.3 ± 77915.5 | 973.7 ± 388.5 | 1055.4 ± 424.3 |
| PLIN2 | 18.5 ± 6.5 | 12.4 ± 6.5 | 5.9 ± 2.6 | 2.7 ± 0.6 |
| SCARB1 | 10.9 ± 4.3 | 15.7 ± 4.6 | 16.3 ± 8.1 | 18.6 ± 5.0 |
| SCARB2 | 2.7 ± 2.0 | 3.1 ± 1.4 | 7.7 ± 3.2 | 11.5 ± 4.7 |
| TRIB2 | 5.76 ± 1.4 | 6.26 ± 3.3 | 1.92 ± 0.9 | 3.96 ± 1.9 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertevello, P.S.; Teixeira-Gomes, A.-P.; Labas, V.; Cordeiro, L.; Blache, M.-C.; Papillier, P.; Singina, G.; Uzbekov, R.; Maillard, V.; Uzbekova, S. MALDI-TOF Mass Spectrometry Revealed Significant Lipid Variations in Follicular Fluid and Somatic Follicular Cells but Not in Enclosed Oocytes between the Large Dominant and Small Subordinate Follicles in Bovine Ovary. Int. J. Mol. Sci. 2020, 21, 6661. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186661
Bertevello PS, Teixeira-Gomes A-P, Labas V, Cordeiro L, Blache M-C, Papillier P, Singina G, Uzbekov R, Maillard V, Uzbekova S. MALDI-TOF Mass Spectrometry Revealed Significant Lipid Variations in Follicular Fluid and Somatic Follicular Cells but Not in Enclosed Oocytes between the Large Dominant and Small Subordinate Follicles in Bovine Ovary. International Journal of Molecular Sciences. 2020; 21(18):6661. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186661
Chicago/Turabian StyleBertevello, Priscila Silvana, Ana-Paula Teixeira-Gomes, Valerie Labas, Luiz Cordeiro, Marie-Claire Blache, Pascal Papillier, Galina Singina, Rustem Uzbekov, Virginie Maillard, and Svetlana Uzbekova. 2020. "MALDI-TOF Mass Spectrometry Revealed Significant Lipid Variations in Follicular Fluid and Somatic Follicular Cells but Not in Enclosed Oocytes between the Large Dominant and Small Subordinate Follicles in Bovine Ovary" International Journal of Molecular Sciences 21, no. 18: 6661. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186661