In Vitro Innovation of Tendon Tissue Engineering Strategies

 ,
,  ,
,  , ,
, ,  , , , , and
, , , , and
Abstract
:1. Introduction
- Identify and/or compare the tenogenic plasticity of different stem/progenitor cell sources,
- Define and drive cell mechanism and environmental conditions leading tenogenesis,
- Control step-wise signaling molecules and pathways,
- Direct stem cell pre-commitment before transplantation (reducing tumorigenic risks with embryonic stem cells (ESCs), unwilling differentiation path of mesenchymal stem cells (MSCs) or to increase tissue integration),
- Study the tenogenic properties of stem cells,
- Test teno-inductive properties of new scaffolds,
- Validate biomechanical teno-inductive stimuli.
1.1. Tendon Structure
1.1.1. Paratenon, Epitenon, and Endotenon
1.1.2. Tendon Properties
1.1.3. Tendon Components
- (1)
- Collagen
- (2)
- Elastin
- (3)
- Proteoglycans
- (4)
- Glycoprotein
1.1.4. Tendon Vasculature
1.1.5. Tendon Innervation
1.2. Tendon Function
1.3. Tendon Biology
2. In Vitro Tenogenesis Techniques
2.1. Stem Cells
2.2. Hypoxia
2.3. Physical Stimuli
2.4. Biomaterials
2.4.1. Materials
- (1)
- Natural Polymers
- (2)
- Synthetic Polymers
2.4.2. Scaffold Fabrication
2.4.3. Tendon Biomimetic Scaffold Structure and Mechanical Properties
2.4.4. Teno-Inductive Potential of Electrospun Produced Materials
2.5. Growth Factors
2.5.1. A Lesson from the Role of Growth Factors In Vitro
2.5.2. TGF β
2.5.3. BMPs
2.5.4. CTGF
2.5.5. FGFs
2.5.6. IGF-1, VEGF, and PDGF
2.5.7. Comparative Studies with Growth Factors
2.6. Co-Culture
3. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Docheva, D.; Müller, S.A.; Majewski, M.; Evans, C.H. Biologics for tendon repair. Adv. Drug Deliv. Rev. 2014, 84, 222–239. [Google Scholar] [CrossRef] [Green Version]
- Maffulli, N. Overuse tendon conditions: Time to change a confusing terminology. Arthrosc. J. Arthrosc. Relat. Surg. 1998, 14, 840–843. [Google Scholar] [CrossRef]
- Sharma, P.; Maffulli, N. Tendon Injury and Tendinopathy: Healing and Repair. J. Bone Jt. Surg. Am. Vol. 2005, 87, 187–202. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, M. Structure and metabolism of tendons. Scand. J. Med. Sci. Sports 1997, 7, 55–61. [Google Scholar] [CrossRef]
- Russo, V.; Mauro, A.; Martelli, A.; Di Giacinto, O.; Di Marcantonio, L.; Nardinocchi, D.; Berardinelli, P.; Barboni, B. Cellular and molecular maturation in fetal and adult ovine calcaneal tendons. J. Anat. 2014, 226, 126–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, G.-C.; Li, Y.-J.; Chen, M.-H.; Lu, P.-P.; Rui, Y.-F. Tendon stem/progenitor cell ageing: Modulation and rejuvenation. World J. Stem Cells 2019, 11, 677–692. [Google Scholar] [CrossRef] [PubMed]
- Matough, F.A.; Budin, S.B.; Hamid, Z.A.; Alwahaibi, N.; Mohamed, J. The role of oxidative stress and antioxidants in diabetic complications. Sultan Qaboos Univ. Med. J. 2012, 12, 5–18. [Google Scholar] [CrossRef]
- Maeda, E.; Shelton, J.C.; Bader, D.L.; Lee, D.A. Differential regulation of gene expression in isolated tendon fascicles exposed to cyclic tensile strain in vitro. J. Appl. Physiol. 2009, 106, 506–512. [Google Scholar] [CrossRef] [Green Version]
- Maeda, E.; Sugimoto, M.; Ohashi, T. Cytoskeletal tension modulates MMP-1 gene expression from tenocytes on micropillar substrates. J. Biomech. 2013, 46, 991–997. [Google Scholar] [CrossRef]
- Favata, M.; Beredjiklian, P.K.; Zgonis, M.H.; Beason, D.P.; Crombleholme, T.M.; Jawad, A.F.; Soslowsky, L.J. Regenerative properties of fetal sheep tendon are not adversely affected by transplantation into an adult environment. J. Orthop. Res. 2006, 24, 2124–2132. [Google Scholar] [CrossRef]
- Benjamin, M.; Qin, S.; Ralphs, J.R. Fibrocartilage associated with human tendons and their pulleys. J. Anat. 1995, 187, 625–633. [Google Scholar] [PubMed]
- Kannus, P. Structure of the tendon connective tissue. Scand. J. Med. Sci. Sports 2000, 10, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Thorpe, C.T.; Birch, H.L.; Clegg, P.; Screen, H.R.C. The role of the non-collagenous matrix in tendon function. Int. J. Exp. Pathol. 2013, 94, 248–259. [Google Scholar] [CrossRef] [PubMed]
- Thorpe, C.T.; Udeze, C.P.; Birch, H.L.; Clegg, P.; Screen, H.R.C. Specialization of tendon mechanical properties results from interfascicular differences. J. R. Soc. Interface 2012, 9, 3108–3117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silver, F.H.; Freeman, J.W.; Seehra, G.P. Collagen self-assembly and the development of tendon mechanical properties. J. Biomech. 2003, 36, 1529–1553. [Google Scholar] [CrossRef]
- Chuen, F.S.; Chuk, C.Y.; Ping, W.Y.; Nar, W.W.; Kim, H.L.; Ming, C.K. Immunohistochemical Characterization of Cells in Adult Human Patellar Tendons. J. Histochem. Cytochem. 2004, 52, 1151–1157. [Google Scholar] [CrossRef]
- McNeilly, C.M.; Banes, A.J.; Benjamin, M.; Ralphs, J.R. Tendon cells in vivo form a three dimensional network of cell processes linked by gap junctions. J. Anat. 1996, 189, 593–600. [Google Scholar] [PubMed]
- Willecke, K.; Eiberger, J.; Degen, J.; Eckardt, D.; Romualdi, A.; Güldenagel, M.; Deutsch, U.; Söhl, G. Structural and Functional Diversity of Connexin Genes in the Mouse and Human Genome. Biol. Chem. 2002, 383, 725–737. [Google Scholar] [CrossRef]
- Tanji, K.; Shimizu, T.; Satou, T.; Hashimoto, S.; Bonilla, E. Gap Junctions between Fibroblasts in Rat Myotendon. Arch. Histol. Cytol. 1995, 58, 97–102. [Google Scholar] [CrossRef] [Green Version]
- Giordano, L.; Della Porta, G.; Peretti, G.M.; Maffulli, N. Therapeutic potential of microRNA in tendon injuries. Br. Med. Bull. 2020, 133, 79–94. [Google Scholar] [CrossRef]
- Bi, Y.; Ehirchiou, D.; Kilts, T.M.; Inkson, C.A.; Embree, M.C.; Sonoyama, W.; Li, L.; Leet, I.A.; Seo, B.-M.; Zhang, L.; et al. Identification of tendon stem/progenitor cells and the role of the extracellular matrix in their niche. Nat. Med. 2007, 13, 1219–1227. [Google Scholar] [CrossRef] [PubMed]
- Józsa, L.G.; Kannus, P. Human Tendons: Anatomy, Physiology, and Pathology; Human Kinetics: Champaign, IL, USA, 1997; ISBN 978-0-87322-484-0. [Google Scholar]
- Salingcarnboriboon, R. Establishment of tendon-derived cell lines exhibiting pluripotent mesenchymal stem cell-like property. Exp. Cell Res. 2003, 287, 289–300. [Google Scholar] [CrossRef]
- Lui, P.P.Y. Identity of tendon stem cells—How much do we know? J. Cell. Mol. Med. 2012, 17, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Mienaltowski, M.J.; Adams, S.M.; Birk, D.E. Regional Differences in Stem Cell/Progenitor Cell Populations from the Mouse Achilles Tendon. Tissue Eng. Part A 2013, 19, 199–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruzzini, L.; Abbruzzese, F.; Rainer, A.; Longo, U.G.; Trombetta, M.; Maffulli, N.; Denaro, V. Characterization of age-related changes of tendon stem cells from adult human tendons. Knee Surg. Sports Traumatol. Arthrosc. 2013, 22, 2856–2866. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Pan, T.; Liu, Y.; Wang, J.H.-C. Mouse treadmill running enhances tendons by expanding the pool of tendon stem cells (TSCs) and TSC-related cellular production of collagen. J. Orthop. Res. 2010, 28, 1178–1183. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, J.H.-C. Mechanobiological response of tendon stem cells: Implications of tendon homeostasis and pathogenesis of tendinopathy. J. Orthop. Res. 2009, 28, 639–643. [Google Scholar] [CrossRef] [PubMed]
- Kirkendall, D.T.; Garrett, W.E. Function and biomechanics of tendons. Scand. J. Med. Sci. Sports 1997, 7, 62–66. [Google Scholar] [CrossRef]
- Curwin, S.L. Biomechanics of tendon and the effects of immobilization. Foot Ankle Clin. 1997, 2, 371–389. [Google Scholar]
- Tuderman, L.; Kivirikko, K.I.; Prockop, D.J. Partial purification and characterization of a neutral protease which cleaves the N-terminal propeptides from procollagen. Biochemistry 1978, 17, 2948–2954. [Google Scholar] [CrossRef]
- Kadler, K.E.; Hojima, Y.; Prockop, D.J. Collagen fibrils in vitro grow from pointed tips in the C- to N-terminal direction. Biochem. J. 1990, 268, 339–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Södersten, F.; Hultenby, K.; Heinegård, D.; Johnston, C.; Ekman, S. Immunolocalization of Collagens (I and III) and Cartilage Oligomeric Matrix Protein in the Normal and Injured Equine Superficial Digital Flexor Tendon. Connect. Tissue Res. 2012, 54, 62–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riley, G.P. The pathogenesis of tendinopathy. A molecular perspective. Rheumatology 2003, 43, 131–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banos, C.C.; Thomas, A.H.; Kuo, C.K. Collagen fibrillogenesis in tendon development: Current models and regulation of fibril assembly. Birth Defects Res. Part C Embryo Today Rev. 2008, 84, 228–244. [Google Scholar] [CrossRef] [PubMed]
- Grant, T.M.; Thompson, M.S.; Urban, J.; Yu, J. Elastic fibres are broadly distributed in tendon and highly localized around tenocytes. J. Anat. 2013, 222, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Flamia, R.; Zhdan, P.A.; Martino, M.; Castle, J.E.; Tamburro, A.M. AFM Study of the Elastin-like Biopolymer Poly(ValGlyGlyValGly). Biomacromolecules 2004, 5, 1511–1518. [Google Scholar] [CrossRef]
- Godinho, M.S.C.; Thorpe, C.T.; Greenwald, S.E.; Screen, H.R.C. Elastin is Localised to the Interfascicular Matrix of Energy Storing Tendons and Becomes Increasingly Disorganised with Ageing. Sci. Rep. 2017, 7, 9713. [Google Scholar] [CrossRef]
- Zhang, G.; Chen, S.; Goldoni, S.; Calder, B.W.; Simpson, H.C.; Owens, R.T.; McQuillan, D.J.; Young, M.F.; Iozzo, R.V.; Birk, D.E. Genetic Evidence for the Coordinated Regulation of Collagen Fibrillogenesis in the Cornea by Decorin and Biglycan. J. Biol. Chem. 2009, 284, 8888–8897. [Google Scholar] [CrossRef] [Green Version]
- Rigozzi, S.; Muller, R.; Stemmer, A.; Snedecker, J. Tendon glycosaminoglycan proteoglycan sidechains promote collagen fibril sliding—AFM observations at the nanoscale. J. Biomech. 2013, 46, 813–818. [Google Scholar] [CrossRef]
- Yoon, J.H.; Halper, J. Tendon proteoglycans: Biochemistry and function. J. Musculoskelet. Neuronal. Interact. 2005, 5, 22–34. [Google Scholar]
- Svensson, L.; Aszódi, A.; Heinegård, D.; Hunziker, E.B.; Reinholt, F.P.; Fässler, R.; Oldberg, A. Cartilage Oligomeric Matrix Protein-Deficient Mice Have Normal Skeletal Development. Mol. Cell. Biol. 2002, 22, 4366–4371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Järvinen, T.A.H.; Józsa, L.; Kannus, P.; Järvinen, T.; Hurme, T.; Kvist, M.; Pelto-Huikko, M.; Kalimo, H.; Järvinen, M. Mechanical loading regulates the expression of tenascin-C in the myotendinous junction and tendon but does not induce de novo synthesis in the skeletal muscle. J. Cell Sci. 2003, 116, 857–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kastelic, J.; Galeski, A.; Baer, E. The Multicomposite Structure of Tendon. Connect. Tissue Res. 1978, 6, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Peacock, E.E. A Study of the Circulation in Normal Tendons and Healing Grafts*. Ann. Surg. 1959, 149, 415–428. [Google Scholar] [CrossRef]
- Ahmed, I.M.; Lagopoulos, M.; McConnell, P.; Soames, R.W.; Sefton, G.K. Blood supply of the achilles tendon. J. Orthop. Res. 1998, 16, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Fenwick, S.A.; Hazleman, B.L.; Riley, G.P. The vasculature and its role in the damaged and healing tendon. Arthritis Res. 2002, 4, 252–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bidder, M.; Towler, D.A.; Gelberman, R.H.; Boyer, M.I. Expression of mRNA for vascular endothelial growth factor at the repair site of healing canine flexor tendon. J. Orthop. Res. 2000, 18, 247–252. [Google Scholar] [CrossRef]
- Kuroda, R.; Kurosaka, M.; Yoshiya, S.; Mizuno, K. Localization of growth factors in the reconstructed anterior cruciate ligament: Immunohistological study in dogs. Knee Surg. Sports Traumatol. Arthrosc. 2000, 8, 120–126. [Google Scholar] [CrossRef]
- Barboni, B.; Russo, V.; Gatta, V.; Bernabò, N.; Berardinelli, P.; Mauro, A.; Martelli, A.; Valbonetti, L.; Muttini, A.; Di Giacinto, O.; et al. Therapeutic potential of hAECs for early Achilles tendon defect repair through regeneration. J. Tissue Eng. Regen. Med. 2017, 12, e1594–e1608. [Google Scholar] [CrossRef]
- Sharma, P.; Maffulli, N. Biology of tendon injury: Healing, modeling and remodeling. J. Musculoskelet. Neuronal. Interact. 2006, 6, 181–190. [Google Scholar]
- Barboni, B.; Russo, V.; Curini, V.; Mauro, A.; Martelli, A.; Muttini, A.; Bernabò, N.; Valbonetti, L.; Marchisio, M.; Di Giacinto, O.; et al. Achilles Tendon Regeneration can be Improved by Amniotic Epithelial Cell Allotransplantation. Cell Transplant. 2012, 21, 2377–2395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ju, Y.-J.; Tohyama, H.; Kondo, E.; Yoshikawa, T.; Muneta, T.; Shinomiya, K.; Yasuda, K. Effects of Local Administration of Vascular Endothelial Growth Factor on Properties of the in Situ Frozen-Thawed Anterior Cruciate Ligament in Rabbits. Am. J. Sports Med. 2006, 34, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, T.; Tohyama, H.; Katsura, T.; Kondo, E.; Kotani, Y.; Matsumoto, H.; Toyama, Y.; Yasuda, K. Effects of Local Administration of Vascular Endothelial Growth Factor on Mechanical Characteristics of the Semitendinosus Tendon Graft after Anterior Cruciate Ligament Reconstruction in Sheep. Am. J. Sports Med. 2006, 34, 1918–1925. [Google Scholar] [CrossRef]
- Wei, X.; Mao, Z.; Hou, Y.; Lin, L.; Xue, T.; Chen, L.; Wang, H.; Yu, C. Local administration of TGFβ-1/VEGF165 gene-transduced bone mesenchymal stem cells for Achilles allograft replacement of the anterior cruciate ligament in rabbits. Biochem. Biophys. Res. Commun. 2011, 406, 204–210. [Google Scholar] [CrossRef]
- Liang, M.; Cornell, H.R.; Baboldashti, N.Z.; Thompson, M.S.; Carr, A.J.; Hulley, P.A. Regulation of Hypoxia-Induced Cell Death in Human Tenocytes. Adv. Orthop. 2012, 2012, 1–12. [Google Scholar] [CrossRef]
- Ackermann, P.W. Tendinopathies in Sports: From Basic Research to the Field. In Sports Injuries; Doral, M.N., Karlsson, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 1–15. ISBN 978-3-642-36801-1. [Google Scholar]
- Ackermann, P.W.; Salo, P.; Hart, D.A. Tendon Innervation. Adv. Exp. Med. Biol. 2016, 920, 35–51. [Google Scholar] [CrossRef]
- Greve, K.; Labruto, F.; Edman, G.; Bring, D.; Nilsson, G.; Ackermann, P.W.; Domeij-Arverud, E. Metabolic activity in early tendon repair can be enhanced by intermittent pneumatic compression. Scand. J. Med. Sci. Sports 2012, 22, e55–e63. [Google Scholar] [CrossRef]
- Nilsson, J.; Von Euler, A.M.; Dalsgaard, C.-J. Stimulation of connective tissue cell growth by substance P and substance K. Nature 1985, 315, 61–63. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.S.; Lee, J.; Lee, E.; Kwon, Y.S.; Lee, E.; Ahn, W.; Jiang, M.H.; Kim, J.C.; Son, Y. A new role of substance P as an injury-inducible messenger for mobilization of CD29+ stromal-like cells. Nat. Med. 2009, 15, 425–435. [Google Scholar] [CrossRef]
- Haegerstrand, A.; Dalsgaard, C.J.; Jonzon, B.; Larsson, O.; Nilsson, J. Calcitonin gene-related peptide stimulates proliferation of human endothelial cells. Proc. Natl. Acad. Sci. USA 1990, 87, 3299–3303. [Google Scholar] [CrossRef] [Green Version]
- Ziche, M.; Morbidelli, L.; Pacini, M.; Geppetti, P.; Alessandri, G.; Maggi, C.A. Substance P stimulates neovascularization in vivo and proliferation of cultured endothelial cells. Microvasc. Res. 1990, 40, 264–278. [Google Scholar] [CrossRef]
- Andersson, G.; Backman, L.J.; Scott, A.; Lorentzon, R.; Forsgren, S.; Danielson, P. Substance P accelerates hypercellularity and angiogenesis in tendon tissue and enhances paratendinitis in response to Achilles tendon overuse in a tendinopathy model. Br. J. Sports Med. 2011, 45, 1017–1022. [Google Scholar] [CrossRef] [Green Version]
- Ackermann, P.W. Neuronal regulation of tendon homoeostasis. Int. J. Exp. Pathol. 2013, 94, 271–286. [Google Scholar] [CrossRef]
- Aicale, R.; Tarantino, D.; Maffulli, N. Basic Science of Tendons. In Bio-Orthopaedics; Gobbi, A., Espregueira-Mendes, J., Lane, J.G., Karahan, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 249–273. ISBN 978-3-662-54180-7. [Google Scholar]
- Oxlund, H. Relationships between the Biomechanical Properties, Composition and Molecular Structure of Connective Tissues. Connect. Tissue Res. 1986, 15, 65–72. [Google Scholar] [CrossRef]
- Fyfe, I.; Stanish, W.D. The Use of Eccentric Training and Stretching in the Treatment and Prevention of Tendon Injuries. Clin. Sports Med. 1992, 11, 601–624. [Google Scholar] [CrossRef]
- Diamant, J.; Keller, A.; Baer, E.; Litt, M.; Arridge, R.G.C. Collagen; ultrastructure and its relation to mechanical properties as a function of ageing. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1972, 180, 293–315. [Google Scholar] [CrossRef]
- Kastelic, J.; Baer, E. Deformation in tendon collagen. Symp. Soc. Exp. Biol. 1980, 34, 397–435. [Google Scholar]
- Sasaki, N.; Shukunami, N.; Matsushima, N.; Izumi, Y. Time-resolved X-ray diffraction from tendon collagen during creep using synchrotron radiation. J. Biomech. 1999, 32, 285–292. [Google Scholar] [CrossRef]
- Oakes, B.W.; Singleton, C.; Haut, R.C. Correlation of collagen fibril morphology and tensile modulus in the repairing and normal rabbit patella tendon. In Proceedings of the 44th Annual Meeting of the Orthopaedic Res Soc, Chicago, IL, USA, 16–19 March 1998. [Google Scholar]
- Komi, P.V.; Fukashiro, S.; Järvinen, M. Biomechanical Loading of Achilles Tendon During Normal Locomotion. Clin. Sports Med. 1992, 11, 521–531. [Google Scholar] [CrossRef]
- Alexander, R.M. Energy-saving mechanisms in walking and running. J. Exp. Biol. 1991, 160, 55–69. [Google Scholar]
- Benjamin, M.; Ralphs, J.R. Tendons and ligaments—An overview. Histol. Histopathol. 1997, 12, 1135–1144. [Google Scholar]
- Kubo, K.; Kawakami, Y.; Kanehisa, H.; Fukunaga, T. Measurement of viscoelastic properties of tendon structures in vivo. Scand. J. Med. Sci. Sports 2002, 12, 3–8. [Google Scholar] [CrossRef]
- Gerard, M.P.; Hodgson, D.R.; Rose, R.J.; Walsh, W. Effects of Recombinant Equine Growth Hormone on In Vitro Biomechanical Properties of the Superficial Digital Flexor Tendon of Standardbred Yearlings in Training. Vet. Surg. 2005, 34, 253–259. [Google Scholar] [CrossRef]
- Ker, R. Mechanics of tendon, from an engineering perspective. Int. J. Fatigue 2007, 29, 1001–1009. [Google Scholar] [CrossRef]
- Eyre, D.R.; Paz, M.A.; Gallop, P.M. Cross-Linking in Collagen and Elastin. Annu. Rev. Biochem. 1984, 53, 717–748. [Google Scholar] [CrossRef]
- Bailey, A.J.; Paul, R.; Knott, L. Mechanisms of maturation and ageing of collagen. Mech. Ageing Dev. 1998, 106, 1–56. [Google Scholar] [CrossRef]
- Avery, N.C.; Bailey, A.J. Enzymic and non-enzymic cross-linking mechanisms in relation to turnover of collagen: Relevance to aging and exercise. Scand. J. Med. Sci. Sports 2005, 15, 231–240. [Google Scholar] [CrossRef]
- Birch, H.L.; Wilson, A.M.; Goodship, A.E. Physical activity: Does long-term, high-intensity exercise in horses result in tendon degeneration? J. Appl. Physiol. 2008, 105, 1927–1933. [Google Scholar] [CrossRef]
- Bank, R.A.; Tekoppele, J.M.; Oostingh, G.; Hazleman, B.L.; Riley, G.P. Lysylhydroxylation and non-reducible crosslinking of human supraspinatus tendon collagen: Changes with age and in chronic rotator cuff tendinitis. Ann. Rheum. Dis. 1999, 58, 35–41. [Google Scholar] [CrossRef] [Green Version]
- Thorpe, C.T.; Clegg, P.; Birch, H.L. A review of tendon injury: Why is the equine superficial digital flexor tendon most at risk? Equine Vet. J. 2010, 42, 174–180. [Google Scholar] [CrossRef]
- Birch, H.L. Tendon matrix composition and turnover in relation to functional requirements. Int. J. Exp. Pathol. 2007, 88, 241–248. [Google Scholar] [CrossRef]
- Maffulli, N.; Almekinders, L. (Eds.) The Achilles Tendon; Springer: London, UK, 2007; ISBN 978-1-84628-628-5. [Google Scholar]
- Batson, E.L.; Paramour, R.J.; Smith, T.J.; Birch, H.L.; Patterson-Kane, J.C.; Goodship, A.E. Are the material properties and matrix composition of equine flexor and extensor tendons determined by their functions? Equine Vet. J. 2003, 35, 314–318. [Google Scholar] [CrossRef]
- Dowling, B.A.; Dart, A.J.; Hodgson, D.R.; Rose, R.J.; Walsh, W. The effect of recombinant equine growth hormone on the biomechanical properties of healing superficial digital flexor tendons in horses. Vet. Surg. 2002, 31, 320–324. [Google Scholar] [CrossRef]
- Kubo, K.; Kanehisa, H.; Takeshita, D.; Kawakami, Y.; Fukashiro, S.; Fukunaga, T. In vivo dynamics of human medial gastrocnemius muscle-tendon complex during stretch-shortening cycle exercise. Acta Physiol. Scand. 2000, 170, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, Y.; Muraoka, T.; Ito, S.; Kanehisa, H.; Fukunaga, T. In vivomuscle fibre behaviour during counter-movement exercise in humans reveals a significant role for tendon elasticity. J. Physiol. 2002, 540, 635–646. [Google Scholar] [CrossRef]
- Thorpe, C.T.; Udeze, C.P.; Birch, H.L.; Clegg, P.D.; Screen, H.R. Capacity for sliding between tendon fascicles decreases with ageing in injury prone equine tendons: A possible mechanism for age-related tendinopathy? Eur. Cell Mater. 2013, 25, 48–60. [Google Scholar] [CrossRef]
- Hooley, C.; McCrum, N.; Cohen, R. The viscoelastic deformation of tendon. J. Biomech. 1980, 13, 521–528. [Google Scholar] [CrossRef]
- Zhang, C.; Zhu, J.; Zhou, Y.; Thampatty, B.P.; Wang, J.H.-C. Tendon Stem/Progenitor Cells and Their Interactions with Extracellular Matrix and Mechanical Loading. Stem Cells Int. 2019, 2019, 3674647. [Google Scholar] [CrossRef] [Green Version]
- Lavagnino, M.; Wall, M.E.; Little, D.; Banes, A.J.; Guilak, F.; Arnoczky, S.P. Tendon mechanobiology: Current knowledge and future research opportunities. J. Orthop. Res. 2015, 33, 813–822. [Google Scholar] [CrossRef] [Green Version]
- Galloway, M.T.; Lalley, A.L.; Shearn, J.T. The Role of Mechanical Loading in Tendon Development, Maintenance, Injury, and Repair. J. Bone Jt. Surg. Am. Vol. 2013, 95, 1620–1628. [Google Scholar] [CrossRef] [PubMed]
- Herod, T.W.; Veres, S.P. Development of overuse tendinopathy: A new descriptive model for the initiation of tendon damage during cyclic loading. J. Orthop. Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Riemersa, D.; Schamhardt, H. The cryo-jaw, a clamp designed for in vitro rheology studies of horse digital flexor tendons. J. Biomech. 1982, 15, 619–620. [Google Scholar] [CrossRef]
- David, L.; Grood, E.S.; Noyes, F.R.; Zernicke, R.E. Biomechanics of Ligaments and Tendons. Exerc. Sport Sci. Rev. 1978, 6, 125–182. [Google Scholar] [CrossRef]
- Cohen, R.E.; Hooley, C.J.; McCrum, N.G. Mechanism of the Viscoelastic Deformation of Collagenous Tissue. Nature 1974, 247, 59–61. [Google Scholar] [CrossRef]
- Sensini, A.; Cristofolini, L. Biofabrication of Electrospun Scaffolds for the Regeneration of Tendons and Ligaments. Materials 2018, 11, 1963. [Google Scholar] [CrossRef] [Green Version]
- Seynnes, O.R.; Bojsen-Møller, J.; Albracht, K.; Arndt, A.; Cronin, N.J.; Finni, T.; Magnusson, S.P. Ultrasound-based testing of tendon mechanical properties: A critical evaluation. J. Appl. Physiol. 2015, 118, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Chaudhry, S.; Morrissey, D.; Woledge, R.C.; Bader, D.L.; Screen, H.R.C. Eccentric and concentric loading of the triceps surae: An in vivo study of dynamic muscle and tendon biomechanical parameters. J. Appl. Biomech. 2014, 31, 69–78. [Google Scholar] [CrossRef]
- Arruda, E.M.; Calve, S.; Dennis, R.G.; Mundy, K.; Baar, K. Regional variation of tibialis anterior tendon mechanics is lost following denervation. J. Appl. Physiol. 2006, 101, 1113–1117. [Google Scholar] [CrossRef] [PubMed]
- Lavagnino, M.; Bedi, A.; Walsh, C.P.; Enselman, E.R.S.; Sheibani-Rad, S.; Arnoczky, S.P. Tendon Contraction After Cyclic Elongation Is an Age-Dependent Phenomenon. Am. J. Sports Med. 2014, 42, 1471–1477. [Google Scholar] [CrossRef]
- Brent, A.E.; Schweitzer, R.; Tabin, C.J. A Somitic Compartment of Tendon Progenitors. Cell 2003, 113, 235–248. [Google Scholar] [CrossRef] [Green Version]
- Brent, A. Developmental regulation of somite derivatives: Muscle, cartilage and tendon. Curr. Opin. Genet. Dev. 2002, 12, 548–557. [Google Scholar] [CrossRef]
- Cserjesi, P.; Brown, D.; Ligon, K.L.; Lyons, G.E.; Copeland, N.G.; Gilbert, D.J.; Jenkins, N.A.; Olson, E.N. Scleraxis: A basic helix-loop-helix protein that prefigures skeletal formation during mouse embryogenesis. Development 1995, 121, 1099–1110. [Google Scholar] [PubMed]
- Schweitzer, R.; Chyung, J.H.; Murtaugh, L.C.; Brent, A.E.; Rosen, V.; Olson, E.N.; Lassar, A.; Tabin, C.J. Analysis of the tendon cell fate using Scleraxis, a specific marker for tendons and ligaments. Development 2001, 128, 3855–3866. [Google Scholar] [PubMed]
- Caceres, M.D.; Pfeifer, C.G.; Docheva, D. Understanding Tendons: Lessons from Transgenic Mouse Models. Stem Cells Dev. 2018, 27, 1161–1174. [Google Scholar] [CrossRef] [PubMed]
- Brent, A.E.; Braun, T.; Tabin, C.J. Genetic analysis of interactions between the somitic muscle, cartilage and tendon cell lineages during mouse development. Development 2005, 132, 515–528. [Google Scholar] [CrossRef] [Green Version]
- Tozer, S.; Duprez, D. Tendon and ligament: Development, repair and disease. Birth Defects Res. Part C Embryo Today Rev. 2005, 75, 226–236. [Google Scholar] [CrossRef]
- Chevallier, A.; Kieny, M.; Mauger, A. Limb-somite relationship: Origin of the limb musculature. J. Embryol. Exp. Morphol. 1977, 41, 245–258. [Google Scholar]
- Kuo, C.K.; Petersen, B.C.; Tuan, R.S. Spatiotemporal protein distribution of TGF-βs, their receptors, and extracellular matrix molecules during embryonic tendon development. Dev. Dyn. 2008, 237, 1477–1489. [Google Scholar] [CrossRef] [Green Version]
- Pryce, B.A.; Watson, S.S.; Murchison, N.D.; Staverosky, J.A.; Dünker, N.; Schweitzer, R. Recruitment and maintenance of tendon progenitors by TGFbeta signaling are essential for tendon formation. Development 2009, 136, 1351–1361. [Google Scholar] [CrossRef] [Green Version]
- Havis, E.; Bonnin, M.-A.; Olivera-Martinez, I.; Nazaret, N.; Ruggiu, M.; Weibel, J.; Durand, C.; Guerquin, M.J.; Bonod-Bidaud, C.; Ruggiero, F.; et al. Transcriptomic analysis of mouse limb tendon cells during development. Development 2014, 141, 3683–3696. [Google Scholar] [CrossRef] [Green Version]
- Dünker, N.; Krieglstein, K. Tgfß2 -/- Tgfß3 -/- double knockout mice display severe midline fusion defects and early embryonic lethality. Brain Struct. Funct. 2002, 206, 73–83. [Google Scholar] [CrossRef]
- Liu, H.; Zhu, S.; Zhang, C.; Lu, P.; Hu, J.; Yin, Z.; Ma, Y.; Chen, X.; Ouyang, H. Crucial transcription factors in tendon development and differentiation: Their potential for tendon regeneration. Cell Tissue Res. 2014, 356, 287–298. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, C.; Zhu, S.; Lu, P.; Zhu, T.; Gong, X.; Zhang, Z.; Hu, J.; Yin, Z.; Heng, B.C.; et al. Mohawk Promotes the Tenogenesis of Mesenchymal Stem Cells Through Activation of the TGFβ Signaling Pathway. Stem Cells 2015, 33, 443–455. [Google Scholar] [CrossRef]
- Berthet, E.; Chen, C.; Butcher, K.; Schneider, R.A.; Alliston, T.; Amirtharajah, M. Smad3 binds scleraxis and mohawk and regulates tendon matrix organization. J. Orthop. Res. 2013, 31, 1475–1483. [Google Scholar] [CrossRef] [Green Version]
- Ito, Y.; Toriuchi, N.; Yoshitaka, T.; Ueno-Kudoh, H.; Sato, T.; Yokoyama, S.; Nishida, K.; Akimoto, T.; Takahashi, M.; Miyaki, S.; et al. The Mohawk homeobox gene is a critical regulator of tendon differentiation. Proc. Natl. Acad. Sci. USA 2010, 107, 10538–10542. [Google Scholar] [CrossRef] [Green Version]
- Kimura, W.; Machii, M.; Xue, X.; Sultana, N.; Hikosaka, K.; Sharkar, M.T.; Uezato, T.; Matsuda, M.; Koseki, H.; Miura, N. Irxl1 mutant mice show reduced tendon differentiation and no patterning defects in musculoskeletal system development. Genes 2010, 49, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Watson, S.S.; Lan, Y.; Keene, D.R.; Ovitt, C.E.; Liu, H.; Schweitzer, R.; Jiang, R. The Atypical Homeodomain Transcription Factor Mohawk Controls Tendon Morphogenesis. Mol. Cell. Biol. 2010, 30, 4797–4807. [Google Scholar] [CrossRef] [Green Version]
- Docheva, D.; Hunziker, E.B.; Fässler, R.; Brandau, O. Tenomodulin Is Necessary for Tenocyte Proliferation and Tendon Maturation. Mol. Cell. Biol. 2005, 25, 699–705. [Google Scholar] [CrossRef] [Green Version]
- Alberton, P.; Dex, S.; Popov, C.; Shukunami, C.; Schieker, M.; Docheva, D. Loss of tenomodulin results in reduced self-renewal and augmented senescence of tendon stem/progenitor cells. Stem Cells Dev. 2014, 24, 597–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dex, S.; Alberton, P.; Willkomm, L.; Söllradl, T.; Bago, S.; Milz, S.; Shakibaei, M.; Ignatius, A.; Bloch, W.; Clausen-Schaumann, H.; et al. Tenomodulin is Required for Tendon Endurance Running and Collagen I Fibril Adaptation to Mechanical Load. EBioMedicine 2017, 20, 240–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandau, O.; Meindl, A.; Fässler, R.; Aszodi, A. A novel gene, tendin, is strongly expressed in tendons and ligaments and shows high homology with chondromodulin-I. Dev. Dyn. 2001, 221, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Nourissat, G.; Berenbaum, F.; Duprez, D. Tendon injury: From biology to tendon repair. Nat. Rev. Rheumatol. 2015, 11, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Melichian, D.S.; De La Garza, M.; Gruner, K.; Bhattacharyya, S.; Barr, L.; Nair, A.; Shahrara, S.; Sporn, P.H.S.; Mustoe, T.A.; et al. Essential Roles for Early Growth Response Transcription Factor Egr-1 in Tissue Fibrosis and Wound Healing. Am. J. Pathol. 2009, 175, 1041–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerquin, M.J.; Charvet, B.; Nourissat, G.; Havis, E.; Ronsin, O.; Bonnin, M.-A.; Ruggiu, M.; Olivera-Martinez, I.; Robert, N.; Lu, Y.; et al. Transcription factor EGR1 directs tendon differentiation and promotes tendon repair. J. Clin. Investig. 2013, 123, 3564–3576. [Google Scholar] [CrossRef] [Green Version]
- Lejard, V.; Blais, F.; Guerquin, M.-J.; Bonnet, A.; Bonnin, M.-A.; Havis, E.; Malbouyres, M.; Bidaud, C.B.; Maro, G.; Gilardi-Hebenstreit, P.; et al. EGR1 and EGR2 Involvement in Vertebrate Tendon Differentiation. J. Biol. Chem. 2010, 286, 5855–5867. [Google Scholar] [CrossRef] [Green Version]
- Cong, X.X.; Rao, X.S.; Lin, J.X.; Liu, X.C.; Zhang, G.A.; Gao, X.K.; He, M.Y.; Shen, W.L.; Fan, W.; Pioletti, D.; et al. Activation of AKT-mTOR Signaling Directs Tenogenesis of Mesenchymal Stem Cells. Stem Cells 2018, 36, 527–539. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, A.; Schilling, T.F. Tendon development and musculoskeletal assembly: Emerging roles for the extracellular matrix. Development 2015, 142, 4191–4204. [Google Scholar] [CrossRef] [Green Version]
- Murchison, N.D.; Price, B.A.; Conner, D.A.; Keene, D.R.; Olson, E.N.; Tabin, C.J.; Schweitzer, R. Regulation of tendon differentiation by scleraxis distinguishes force-transmitting tendons from muscle-anchoring tendons. Development 2007, 134, 2697–2708. [Google Scholar] [CrossRef] [Green Version]
- Espira, L.; Lamoureux, L.; Jones, S.C.; Gerard, R.D.; Dixon, I.M.; Czubryt, M.P. The basic helix–loop–helix transcription factor scleraxis regulates fibroblast collagen synthesis. J. Mol. Cell. Cardiol. 2009, 47, 188–195. [Google Scholar] [CrossRef]
- Léjard, V.; Brideau, G.; Blais, F.; Salingcarnboriboon, R.; Wagner, G.; Roehrl, M.H.; Noda, M.; Duprez, D.; Houillier, P.; Rossert, J. Scleraxis and NFATc regulate the expression of the pro-alpha1(I) collagen gene in tendon fibroblasts. J. Biol. Chem. 2007, 282, 17665–17675. [Google Scholar] [CrossRef] [Green Version]
- Shukunami, C.; Takimoto, A.; Oro, M.; Hiraki, Y. Scleraxis positively regulates the expression of tenomodulin, a differentiation marker of tenocytes. Dev. Biol. 2006, 298, 234–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scadden, D.T. The stem-cell niche as an entity of action. Nature 2006, 441, 1075–1079. [Google Scholar] [CrossRef] [PubMed]
- Vinhas, A.; Rodrigues, M.T.; Gomes, M.E. Exploring Stem Cells and Inflammation in Tendon Repair and Regeneration. In Cell Biology and Translational Medicine, Volume 2; Turksen, K., Ed.; Springer International Publishing: Cham, Switzerland, 2018; Volume 1089, pp. 37–46. ISBN 978-3-030-04169-4. [Google Scholar]
- Bedi, A.; Kovacevi, D.; Hettrich, C.; Gulotta, L.V.; Ehteshami, J.R.; Warren, R.F.; Rodeo, S.A. The effect of matrix metalloproteinase inhibition on tendon-to-bone healing in a rotator cuff repair model. J. Shoulder Elb. Surg. 2010, 19, 384–391. [Google Scholar] [CrossRef]
- Gotoh, M.; Mitsui, Y.; Shibata, H.; Yamada, T.; Shirachi, I.; Nakama, K.; Okawa, T.; Higuchi, F.; Nagata, K. Increased matrix metalloprotease-3 gene expression in ruptured rotator cuff tendons is associated with postoperative tendon retear. Knee Surg. Sports Traumatol. Arthrosc. 2012, 21, 1807–1812. [Google Scholar] [CrossRef]
- Jones, G.C.; Corps, A.N.; Pennington, C.J.; Clark, I.M.; Edwards, D.R.; Bradley, M.M.; Hazleman, B.L.; Riley, G.P. Expression profiling of metalloproteinases and tissue inhibitors of metalloproteinases in normal and degenerate human achilles tendon. Arthritis Rheum. 2006, 54, 832–842. [Google Scholar] [CrossRef] [Green Version]
- Ueda, Y.; Inui, A.; Mifune, Y.; Takase, F.; Kataoka, T.; Kurosawa, T.; Yamaura, K.; Kokubu, T.; Kuroda, R. Molecular changes to tendons after collagenase-induced acute tendon injury in a senescence-accelerated mouse model. BMC Musculoskelet. Disord. 2019, 20, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, D.; Alberton, P.; Caceres, M.D.; Volkmer, E.; Schieker, M.; Docheva, D. Tenomodulin is essential for prevention of adipocyte accumulation and fibrovascular scar formation during early tendon healing. Cell Death Dis. 2017, 8, e3116. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Akinbiyi, T.; Xu, L.; Ramcharan, M.; Leong, D.J.; Ros, S.J.; Colvin, A.C.; Schaffler, M.B.; Majeska, R.J.; Flatow, E.L.; et al. Tendon-derived stem/progenitor cell aging: Defective self-renewal and altered fate. Aging Cell 2010, 9, 911–915. [Google Scholar] [CrossRef] [Green Version]
- Mary, T.J. Proteomics of tendinopathy. Front. Biosci. 2009, 14, 1505. [Google Scholar] [CrossRef] [Green Version]
- Sejersen, M.H.J.; Frost, P.; Hansen, T.; Deutch, S.R.; Svendsen, S.W. Proteomics Perspectives in Rotator Cuff Research: A Systematic Review of Gene Expression and Protein Composition in Human Tendinopathy. PLoS ONE 2015, 10, e0119974. [Google Scholar] [CrossRef]
- Maffulli, N.; Ewen, S.W.B.; Waterston, S.W.; Reaper, J.; Barrass, V. Tenocytes from Ruptured and Tendinopathic Achilles Tendons Produce Greater Quantities of Type III Collagen than Tenocytes from Normal Achilles Tendons: An in Vitro Model of Human Tendon Healing. Am. J. Sports Med. 2000, 28, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Mendias, C.L.; Gumucio, J.P.; Bakhurin, K.I.; Lynch, E.B.; Brooks, S.V. Physiological loading of tendons induces scleraxis expression in epitenon fibroblasts. J. Orthop. Res. 2011, 30, 606–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nichols, A.E.C.; Settlage, R.E.; Werre, S.R.; Dahlgren, L.A. Novel roles for scleraxis in regulating adult tenocyte function. BMC Cell Biol. 2018, 19, 14. [Google Scholar] [CrossRef] [PubMed]
- Dyment, N.A.; Liu, C.-F.; Kazemi, N.; Aschbacher-Smith, L.E.; Kenter, K.; Breidenbach, A.P.; Shearn, J.T.; Wylie, C.; Rowe, D.W.; Butler, D.L. The Paratenon Contributes to Scleraxis-Expressing Cells during Patellar Tendon Healing. PLoS ONE 2013, 8, e59944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shukunami, C.; Oshima, Y.; Hiraki, Y. Molecular Cloning of tenomodulin, a Novel Chondromodulin-I Related Gene. Biochem. Biophys. Res. Commun. 2001, 280, 1323–1327. [Google Scholar] [CrossRef]
- Kimura, N.; Shukunami, C.; Hakuno, D.; Yoshioka, M.; Miura, S.; Docheva, D.; Kimura, T.; Okada, Y.; Matsumura, G.; Shinoka, T.; et al. Local Tenomodulin Absence, Angiogenesis, and Matrix Metalloproteinase Activation Are Associated With the Rupture of the Chordae Tendineae Cordis. Circulation 2008, 118, 1737–1747. [Google Scholar] [CrossRef]
- Dex, S.; Lin, D.; Shukunami, C.; Docheva, D. Tenogenic modulating insider factor: Systematic assessment on the functions of tenomodulin gene. Gene 2016, 587, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Jelinsky, S.A.; Archambault, J.; Li, L.; Seeherman, H. Tendon-selective genes identified from rat and human musculoskeletal tissues. J. Orthop. Res. 2009, 28, 289–297. [Google Scholar] [CrossRef]
- Frolova, E.G.; Drazba, J.; Krukovets, I.; Kostenko, V.; Blech, L.; Harry, C.; Vasanji, A.; Drumm, C.; Sul, P.; Jenniskens, G.J.; et al. Control of organization and function of muscle and tendon by thrombospondin-4. Matrix Biol. 2014, 37, 35–48. [Google Scholar] [CrossRef]
- Subramanian, A.; Schilling, T.F. Thrombospondin-4 controls matrix assembly during development and repair of myotendinous junctions. eLife 2014, 3. [Google Scholar] [CrossRef]
- Mackie, E.J.; Ramsey, S. Expression of tenascin in joint-associated tissues during development and postnatal growth. J. Anat. 1996, 188, 157–165. [Google Scholar] [PubMed]
- Riley, G.P.; Harrall, R.L.; Cawston, T.E.; Hazleman, B.L.; Mackie, E.J. Tenascin-C and human tendon degeneration. Am. J. Pathol. 1996, 149, 933–943. [Google Scholar] [PubMed]
- Järvinen, T.A.; Jozsa, L.; Kannus, P.; Kvist, M.; Hurme, T.; Isola, J.; Kalimo, H.; Järvinen, M. Mechanical loading regulates tenascin-C expression in the osteotendinous junction. J. Cell Sci. 1999, 112, 3157–3166. [Google Scholar] [PubMed]
- Mehr, D.; Pardubsky, P.D.; Martin, J.A.; Buckwalter, J.A. Tenascin-C in tendon regions subjected to compression. J. Orthop. Res. 2000, 18, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Qi, F.; Deng, Z.; Ma, Y.; Wang, S.; Liu, C.; Lyu, F.; Wang, T.; Zheng, Q. From the perspective of embryonic tendon development: Various cells applied to tendon tissue engineering. Ann. Transl. Med. 2020, 8, 131. [Google Scholar] [CrossRef] [PubMed]
- Guillot, P.V.; Cui, W.; Fisk, N.M.; Polak, J. Stem cell differentiation and expansion for clinical applications of tissue engineering. J. Cell. Mol. Med. 2007, 11, 935–944. [Google Scholar] [CrossRef] [Green Version]
- Crook, J.M.; Peura, T.T.; Kravets, L.; Bosman, A.G.; Buzzard, J.J.; Horne, R.; Hentze, H.; Dunn, N.R.; Zweigerdt, R.; Chua, F.; et al. The generation of six clinical-grade human embryonic stem cell lines. Cell Stem Cell 2007, 1, 490–494. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Song, X.-H.; Yin, Z.; Zou, X.-H.; Wang, L.-L.; Hu, H.; Cao, T.; Zheng, M.; Ouyang, H.W. Stepwise Differentiation of Human Embryonic Stem Cells Promotes Tendon Regeneration by Secreting Fetal Tendon Matrix and Differentiation Factors. Stem Cells 2009, 27, 1276–1287. [Google Scholar] [CrossRef]
- Chen, X.; Yin, Z.; Chen, J.; Shen, W.-L.; Liu, H.-H.; Tang, Q.-M.; Fang, Z.; Lu, L.-R.; Ji, J.; Ouyang, H.-W. Force and scleraxis synergistically promote the commitment of human ES cells derived MSCs to tenocytes. Sci. Rep. 2012, 2, 977. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Yin, Z.; Shen, W.-L.; Chen, X.; Heng, B.C.; Zou, X.H.; Ouyang, H.W. Efficacy of hESC-MSCs in knitted silk-collagen scaffold for tendon tissue engineering and their roles. Biomaterials 2010, 31, 9438–9451. [Google Scholar] [CrossRef]
- Dale, T.P.; Mazher, S.; Webb, W.R.; Zhou, J.; Maffulli, N.; Chen, G.-Q.; El Haj, A.J.; Forsyth, N.R. Tenogenic Differentiation of Human Embryonic Stem Cells. Tissue Eng. Part A 2018, 24, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bavin, E.P.; Smith, O.; Baird, A.E.G.; Smith, L.C.; Guest, D. Equine Induced Pluripotent Stem Cells have a Reduced Tendon Differentiation Capacity Compared to Embryonic Stem Cells. Front. Vet. Sci. 2015, 2. [Google Scholar] [CrossRef] [Green Version]
- Komura, S.; Satake, T.; Goto, A.; Aoki, H.; Shibata, H.; Ito, K.; Hirakawa, A.; Yamada, Y.; Akiyama, H. Induced pluripotent stem cell-derived tenocyte-like cells promote the regeneration of injured tendons in mice. Sci. Rep. 2020, 10, 3992. [Google Scholar] [CrossRef] [Green Version]
- Parolini, O.; Soncini, M.; Evangelista, M.; Schmidt, D. Amniotic membrane and amniotic fluid-derived cells: Potential tools for regenerative medicine? Regen. Med. 2009, 4, 275–291. [Google Scholar] [CrossRef] [Green Version]
- Miki, T. Amnion-derived stem cells: In quest of clinical applications. Stem Cell Res. Ther. 2011, 2, 25. [Google Scholar] [CrossRef] [Green Version]
- Muttini, A.; Barboni, B.; Valbonetti, L.; Russo, V.; Maffulli, N. Amniotic Epithelial Stem Cells. Sports Med. Arthrosc. Rev. 2018, 26, 70–74. [Google Scholar] [CrossRef]
- Miki, T. Stem cell characteristics and the therapeutic potential of amniotic epithelial cells. Am. J. Reprod. Immunol. 2018, 80, e13003. [Google Scholar] [CrossRef]
- Hao, Y.; Ma, D.H.-K.; Hwang, D.G.; Kim, W.-S.; Zhang, F. Identification of antiangiogenic and antiinflammatory proteins in human amniotic membrane. Cornea 2000, 19, 348–352. [Google Scholar] [CrossRef]
- Kim, J.S.; Na, B.K.; Jeong, J.M.; Song, C.Y.; Kim, J.C. Amniotic Membrane Patching Promotes Healing and Inhibits Proteinase Activity on Wound Healing Following Acute Corneal Alkali Burn. Exp. Eye Res. 2000, 70, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Solomon, A.; Rosenblatt, M.; Monroy, D.; Ji, Z.; Pflugfelder, S.C.; Tseng, S.C.G. Suppression of interleukin 1α and interleukin 1β in human limbal epithelial cells cultured on the amniotic membrane stromal matrix. Br. J. Ophthalmol. 2001, 85, 444–449. [Google Scholar] [CrossRef] [Green Version]
- Tseng, S.C.; Li, D.Q.; Ma, X. Suppression of transforming growth factor-beta isoforms, TGF-beta receptor type II, and myofibroblast differentiation in cultured human corneal and limbal fibroblasts by amniotic membrane matrix. J. Cell. Physiol. 1999, 179, 325–335. [Google Scholar] [CrossRef]
- Inge, E.; Talmi, Y.P.; Sigler, L.; Finkelstein, Y.; Zohar, Y. Antibacterial properties of human amniotic membranes. Placenta 1991, 12, 285–288. [Google Scholar] [CrossRef]
- Hori, J.; Wang, M.; Kamiya, K.; Takahashi, H.; Sakuragawa, N. Immunological Characteristics of Amniotic Epithelium. Cornea 2006, 25, S53–S58. [Google Scholar] [CrossRef]
- Toda, A.; Okabe, M.; Yoshida, T.; Nikaido, T. The potential of amniotic membrane/amnion-derived cells for regeneration of various tissues. J. Pharmacol. Sci. 2007, 105, 215–228. [Google Scholar] [CrossRef] [Green Version]
- Parolini, O. From fetal development and beyond: A continued role for placenta in sustaining life? Placenta 2011, 32, S283–S284. [Google Scholar] [CrossRef]
- Barboni, B.; Curini, V.; Russo, V.; Mauro, A.; Di Giacinto, O.; Marchisio, M.; Alfonsi, M.; Mattioli, M. Indirect Co-Culture with Tendons or Tenocytes Can Program Amniotic Epithelial Cells towards Stepwise Tenogenic Differentiation. PLoS ONE 2012, 7, e30974. [Google Scholar] [CrossRef] [Green Version]
- Barboni, B.; Russo, V.; Berardinelli, P.; Mauro, A.; Valbonetti, L.; Sanyal, H.; Canciello, A.; Greco, L.; Muttini, A.; Gatta, V.; et al. Placental Stem Cells from Domestic Animals. Cell Transplant. 2018, 27, 93–116. [Google Scholar] [CrossRef]
- Lange-Consiglio, A.; Rossi, D.; Tassan, S.; Perego, R.; Cremonesi, F.; Parolini, O. Conditioned Medium from Horse Amniotic Membrane-Derived Multipotent Progenitor Cells: Immunomodulatory Activity In Vitro and First Clinical Application in Tendon and Ligament Injuries In Vivo. Stem Cells Dev. 2013, 22, 3015–3024. [Google Scholar] [CrossRef]
- Lange-Consiglio, A.; Tassan, S.; Corradetti, B.; Meucci, A.; Perego, R.; Bizzaro, D.; Cremonesi, F. Investigating the efficacy of amnion-derived compared with bone marrow–derived mesenchymal stromal cells in equine tendon and ligament injuries. Cytotherapy 2013, 15, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Muttini, A.; Valbonetti, L.; Abate, M.; Colosimo, A.; Curini, V.; Mauro, A.; Berardinelli, P.; Russo, V.; Cocciolone, D.; Marchisio, M.; et al. Ovine amniotic epithelial cells: In vitro characterization and transplantation into equine superficial digital flexor tendon spontaneous defects. Res. Vet. Sci. 2013, 94, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Colosimo, A.; Curini, V.; Russo, V.; Mauro, A.; Bernabò, N.; Marchisio, M.; Alfonsi, M.; Muttini, A.; Mattioli, M.; Barboni, B. Characterization, GFP Gene Nucleofection, and Allotransplantation in Injured Tendons of Ovine Amniotic Fluid-Derived Stem Cells. Cell Transplant. 2013, 22, 99–117. [Google Scholar] [CrossRef]
- Russo, V.; El Khatib, M.; Di Marcantonio, L.; Ancora, M.; Wyrwa, R.; Mauro, A.; Walter, T.; Weisser, J.; Citeroni, M.R.; Lazzaro, F.; et al. Tendon Biomimetic Electrospun PLGA Fleeces Induce an Early Epithelial-Mesenchymal Transition and Tenogenic Differentiation on Amniotic Epithelial Stem Cells. Cells 2020, 9, 303. [Google Scholar] [CrossRef] [Green Version]
- Canciello, A.; Russo, V.; Berardinelli, P.; Bernabò, N.; Muttini, A.; Mattioli, M.; Barboni, B. Progesterone prevents epithelial-mesenchymal transition of ovine amniotic epithelial cells and enhances their immunomodulatory properties. Sci. Rep. 2017, 7, 3761. [Google Scholar] [CrossRef] [Green Version]
- Di Lollo, V.; Canciello, A.; Orsini, M.; Bernabò, N.; Ancora, M.; Di Federico, M.; Curini, V.; Mattioli, M.; Russo, V.; Mauro, A.; et al. Transcriptomic and computational analysis identified LPA metabolism, KLHL14 and KCNE3 as novel regulators of Epithelial-Mesenchymal Transition. Sci. Rep. 2020, 10, 4180. [Google Scholar] [CrossRef]
- Li, Y.; Liu, Z.; Jin, Y.; Zhu, X.; Wang, S.; Yang, J.; Ren, Y.; Fu, Q.; Xiong, H.; Zou, G.; et al. Differentiation of Human Amniotic Mesenchymal Stem Cells into Human Anterior Cruciate Ligament Fibroblast Cells by In Vitro Coculture. BioMed Res. Int. 2017, 2017, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Gulati, B.R.; Kumar, R.; Mohanty, N.; Kumar, P.; Somasundaram, R.K.; Yadav, P.S. Bone Morphogenetic Protein-12 Induces Tenogenic Differentiation of Mesenchymal Stem Cells Derived from Equine Amniotic Fluid. Cells Tissues Organs 2013, 198, 377–389. [Google Scholar] [CrossRef]
- Mohanty, N.; Gulati, B.R.; Kumar, R.; Gera, S.; Kumar, P.; Somasundaram, R.K.; Kumar, S. Immunophenotypic characterization and tenogenic differentiation of mesenchymal stromal cells isolated from equine umbilical cord blood. In Vitro Cell. Dev. Biol. Anim. 2014, 50, 538–548. [Google Scholar] [CrossRef]
- Ciardulli, M.C.; Marino, L.; Lamparelli, E.P.; Guida, M.; Forsyth, N.; Selleri, C.; Della Porta, G.; Maffulli, N. Dose-Response Tendon-Specific Markers Induction by Growth Differentiation Factor-5 in Human Bone Marrow and Umbilical Cord Mesenchymal Stem Cells. Int. J. Mol. Sci. 2020, 21, 5905. [Google Scholar] [CrossRef] [PubMed]
- Meirelles, L.D.S.; Chagastelles, P.C.; Nardi, N.B. Mesenchymal stem cells reside in virtually all post-natal organs and tissues. J. Cell Sci. 2006, 119, 2204–2213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Govoni, M.; Berardi, A.C.; Muscari, C.; Campardelli, R.; Bonafè, F.; Guarnieri, C.; Reverchon, E.; Giordano, E.; Maffulli, N.; Della Porta, G. An Engineered Multiphase Three-Dimensional Microenvironment to Ensure the Controlled Delivery of Cyclic Strain and Human Growth Differentiation Factor 5 for the Tenogenic Commitment of Human Bone Marrow Mesenchymal Stem Cells. Tissue Eng. Part A 2017, 23, 811–822. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.-L.; Ahmad, R.E.; Ahmad, T.S.; Merican, A.M.; Abbas, A.A.; Ng, W.M.; Kamarul, T. Effect of Growth Differentiation Factor 5 on the Proliferation and Tenogenic Differentiation Potential of Human Mesenchymal Stem Cells in vitro. Cells Tissues Organs 2012, 196, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.-W.; Chen, Z.-L.; Piao, Y.-J. Mesenchymal stem cells differentiate into tenocytes by bone morphogenetic protein (BMP) 12 gene transfer. J. Biosci. Bioeng. 2005, 100, 418–422. [Google Scholar] [CrossRef]
- Bottagisio, M.; Lopa, S.; Granata, V.; Talò, G.; Bazzocchi, C.; Moretti, M.; Lovati, A.B. Different combinations of growth factors for the tenogenic differentiation of bone marrow mesenchymal stem cells in monolayer culture and in fibrin-based three-dimensional constructs. Differentiation 2017, 95, 44–53. [Google Scholar] [CrossRef]
- Ciardulli, M.C.; Marino, L.; Lovecchio, J.; Giordano, E.; Forsyth, N.R.; Selleri, C.; Maffulli, N.; Della Porta, G. A study on Tendon and Cytokines Markers expression by Human Mesenchymal Stem Cells from Bone Marrow cultivated into a Hyaluronate/Fibrin 3D scaffold under cyclic strain. Cells 2020, 9, 1268. [Google Scholar] [CrossRef]
- Haramshahi, S.M.A.; Bonakdar, S.; Moghtadaei, M.; Kamguyan, K.; Thormann, E.; Tanbakooei, S.; Simorgh, S.; Milan, P.B.; Amini, N.; Latifi, N.A.; et al. Tenocyte-imprinted substrate: A topography-based inducer for tenogenic differentiation in adipose tissue-derived mesenchymal stem cells. Biomed. Mater. 2020, 15, 035014. [Google Scholar] [CrossRef]
- Dai, L.; Hu, X.; Zhang, X.; Zhu, J.; Zhang, J.; Fu, X.; Duan, X.; Ao, Y.; Zhou, C. Different tenogenic differentiation capacities of different mesenchymal stem cells in the presence of BMP-12. J. Transl. Med. 2015, 13, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Harris, M.; Butler, D.; Boivin, G.P.; Florer, J.; Schantz, E.; Wenstrup, R.J. Mesenchymal stem cells used for rabbit tendon repair can form ectopic bone and express alkaline phosphatase activity in constructs. J. Orthop. Res. 2004, 22, 998–1003. [Google Scholar] [CrossRef]
- Zarychta-Wiśniewska, W.; Burdzinska, A.; Kulesza, A.; Gala, K.; Kaleta, B.; Zielniok, K.; Siennicka, K.; Sabat, M.; Pączek, L. Bmp-12 activates tenogenic pathway in human adipose stem cells and affects their immunomodulatory and secretory properties. BMC Cell Biol. 2017, 18, 13. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Rothrauff, B.B.; Lin, H.; Yu, S.; Tuan, R.S. Tendon-Derived Extracellular Matrix Enhances Transforming Growth Factor-β3-Induced Tenogenic Differentiation of Human Adipose-Derived Stem Cells. Tissue Eng. Part A 2017, 23, 166–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, H.; Domingues, R.M.A.; Mithieux, S.M.; Pires, R.A.; Gonçalves, A.I.; Gómez-Florit, M.; Reis, R.L.; Weiss, A.S.; Gomes, M.E. Tropoelastin-Coated Tendon Biomimetic Scaffolds Promote Stem Cell Tenogenic Commitment and Deposition of Elastin-Rich Matrix. ACS Appl. Mater. Interfaces 2019, 11, 19830–19840. [Google Scholar] [CrossRef] [PubMed]
- Neo, P.Y.; See, E.Y.-S.; Toh, S.L.; Goh, J.C.-H. Temporal profiling of the growth and multi-lineage potentiality of adipose tissue-derived mesenchymal stem cells cell-sheets. J. Tissue Eng. Regen. Med. 2013, 10, 564–579. [Google Scholar] [CrossRef] [PubMed]
- Sheyn, D.; Mizrahi, O.; Benjamin, S.; Gazit, Z.; Pelled, G.; Gazit, D. Genetically modified cells in regenerative medicine and tissue engineering. Adv. Drug Deliv. Rev. 2010, 62, 683–698. [Google Scholar] [CrossRef]
- Li, B.; Qi, Y.; Ni, Q.; Jha, R.K.; Wang, H.; Chen, B.; Wang, H. Early cellular responses of BMSCs genetically modified with bFGF/BMP2 co-cultured with ligament fibroblasts in a three-dimensional model in vitro. Int. J. Mol. Med. 2016, 38, 1578–1586. [Google Scholar] [CrossRef]
- Hsieh, C.-F.; Alberton, P.; Loffredo-Verde, E.; Volkmer, E.; Pietschmann, M.; Müller, P.; Schieker, M.; Docheva, D. Scaffold-free Scleraxis-programmed tendon progenitors aid in significantly enhanced repair of full-size Achilles tendon rupture. Nanomedicine 2016, 11, 1153–1167. [Google Scholar] [CrossRef] [Green Version]
- Kohler, J.; Popov, C.; Klotz, B.; Alberton, P.; Prall, W.C.; Haasters, F.; Müller-Deubert, S.; Ebert, R.; Klein-Hitpass, L.; Jakob, F.; et al. Uncovering the cellular and molecular changes in tendon stem/progenitor cells attributed to tendon aging and degeneration. Aging Cell 2013, 12, 988–999. [Google Scholar] [CrossRef] [Green Version]
- Ni, M.; Rui, Y.F.; Tan, Q.; Liu, Y.; Xu, L.L.; Chan, K.-M.; Wang, Y.; Li, G. Engineered scaffold-free tendon tissue produced by tendon-derived stem cells. Biomaterials 2013, 34, 2024–2037. [Google Scholar] [CrossRef]
- Guo, J.; Chan, K.-M.; Zhang, J.-F.; Li, G. Tendon-derived stem cells undergo spontaneous tenogenic differentiation. Exp. Cell Res. 2016, 341, 1–7. [Google Scholar] [CrossRef]
- Mazzocca, A.D.; Chowaniec, D.; McCarthy, M.B.; Beitzel, K.; Cote, M.P.; McKinnon, W.; Arciero, R. In vitro changes in human tenocyte cultures obtained from proximal biceps tendon: Multiple passages result in changes in routine cell markers. Knee Surg. Sports Traumatol. Arthrosc. 2011, 20, 1666–1672. [Google Scholar] [CrossRef]
- Yao, L.; Bestwick, C.; Bestwick, L.; Maffulli, N.; Aspden, R. Phenotypic Drift in Human Tenocyte Culture. Tissue Eng. 2006, 12, 1843–1849. [Google Scholar] [CrossRef] [PubMed]
- Abdollahi, H.; Harris, L.J.; Zhang, P.; McIlhenny, S.; Srinivas, V.; Tulenko, T.; DiMuzio, P.J. The Role of Hypoxia in Stem Cell Differentiation and Therapeutics. J. Surg. Res. 2011, 165, 112–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohyeldin, A.; Garzon-Muvdi, T.; Quiñones-Hinojosa, A. Oxygen in Stem Cell Biology: A Critical Component of the Stem Cell Niche. Cell Stem Cell 2010, 7, 150–161. [Google Scholar] [CrossRef] [Green Version]
- Mas-Bargues, C.; Sanz-Ros, J.; Román-Domínguez, A.; Inglés, M.; Gimeno-Mallench, L.; El Alami, M.; Viña-Almunia, J.; Gambini, J.; Viña, J.; Borras, C. Relevance of Oxygen Concentration in Stem Cell Culture for Regenerative Medicine. Int. J. Mol. Sci. 2019, 20, 1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keith, B.; Simon, M.C. Hypoxia-Inducible Factors, Stem Cells, and Cancer. Cell 2007, 129, 465–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollard, P.J.; Yang, M.; Su, H.; Soga, T.; Kranc, K.R. Prolyl hydroxylase domain enzymes: Important regulators of cancer metabolism. Hypoxia 2014, 2, 127–142. [Google Scholar] [CrossRef] [Green Version]
- Jewell, U.R.; Kvietikova, I.; Scheid, A.; Bauer, C.; Wenger, R.H.; Gassmann, M. Induction of HIF–1α in response to hypoxia is instantaneous. FASEB J. 2001, 15, 1312–1314. [Google Scholar] [CrossRef]
- Majmundar, A.J.; Wong, W.J.; Simon, M.C. Hypoxia-Inducible Factors and the Response to Hypoxic Stress. Mol. Cell 2010, 40, 294–309. [Google Scholar] [CrossRef] [Green Version]
- Prabhakar, N.R.; Semenza, G.L. Oxygen Sensing and Homeostasis. Physiology 2015, 30, 340–348. [Google Scholar] [CrossRef] [Green Version]
- Yin, Z.; Guo, J.; Wu, T.-Y.; Chen, X.; Xu, L.-L.; Lin, S.; Sun, Y.-X.; Chan, K.-M.; Ouyang, H.; Li, G. Stepwise Differentiation of Mesenchymal Stem Cells Augments Tendon-Like Tissue Formation and Defect Repair In Vivo. Stem Cells Transl. Med. 2016, 5, 1106–1116. [Google Scholar] [CrossRef]
- Ejtehadifar, M.; Shamsasenjan, K.; Movassaghpour, A.A.; Akbarzadehlaleh, P.; Dehdilani, N.; Abbasi, P.; Molaeipour, Z.; Saleh, M. The Effect of Hypoxia on Mesenchymal Stem Cell Biology. Adv. Pharm. Bull. 2015, 5, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Ivanovic, Z. Hypoxia or in situ normoxia: The stem cell paradigm. J. Cell. Physiol. 2009, 219, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Lennon, N.P.; Edmison, J.M.; Caplan, A.I. Cultivation of rat marrow-derived mesenchymal stem cells in reduced oxygen tension: Effects on in vitro and in vivo osteochondrogenesis. J. Cell. Physiol. 2001, 187, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Carreau, A.; El Hafny-Rahbi, B.; Matejuk, A.; Grillon, C.; Kieda, C. Why is the partial oxygen pressure of human tissues a crucial parameter? Small molecules and hypoxia. J. Cell. Mol. Med. 2011, 15, 1239–1253. [Google Scholar] [CrossRef] [Green Version]
- Simon, M.C.; Keith, B. The role of oxygen availability in embryonic development and stem cell function. Nat. Rev. Mol. Cell Biol. 2008, 9, 285–296. [Google Scholar] [CrossRef]
- Spencer, J.A.; Ferraro, F.; Roussakis, E.; Klein, A.; Wu, J.; Runnels, J.M.; Zaher, W.; Mortensen, L.; Alt, C.; Turcotte, R.; et al. Direct measurement of local oxygen concentration in the bone marrow of live animals. Nature 2014, 508, 269–273. [Google Scholar] [CrossRef] [Green Version]
- Harrison, J.; Rameshwar, P.; Chang, V.; Bandari, P.; Chang, V. Oxygen saturation in the bone marrow of healthy volunteers. Blood 2002, 99, 394. [Google Scholar] [CrossRef] [PubMed]
- Grimshaw, M.; Mason, R. Bovine articular chondrocyte function in vitro depends upon oxygen tension. Osteoarthr. Cartil. 2000, 8, 386–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortiz-Prado, E.; Dunn, J.F.; Vasconez, J.; Castillo, D.; Viscor, G. Partial pressure of oxygen in the human body: A general review. Am. J. Blood Res. 2019, 9, 1–14. [Google Scholar]
- Hirai, D.M.; Colburn, T.D.; Craig, J.C.; Hotta, K.; Kano, Y.; Musch, T.I.; Poole, D.C. Skeletal muscle interstitial O2 pressures: Bridging the gap between the capillary and myocyte. Microcirculation 2018, 26, e12497. [Google Scholar] [CrossRef]
- Shukunami, C.; Takimoto, A.; Miura, S.; Nishizaki, Y.; Hiraki, Y. Chondromodulin-I and tenomodulin are differentially expressed in the avascular mesenchyme during mouse and chick development. Cell Tissue Res. 2008, 332, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.W.; Lui, P.P.Y.; Rui, Y.F. Hypoxia-Mediated Efficient Expansion of Human Tendon–Derived Stem Cells In Vitro. Tissue Eng. Part A 2012, 18, 484–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spanoudes, K.; Gaspar, D.; Pandit, A.; Zeugolis, D.I. The biophysical, biochemical, and biological toolbox for tenogenic phenotype maintenance in vitro. Trends Biotechnol. 2014, 32, 474–482. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, B.; Zhang, W.J.; Zhou, G.; Cao, Y.; Liu, W. Enhanced proliferation capacity of porcine tenocytes in low O2 tension culture. Biotechnol. Lett. 2009, 32, 181–187. [Google Scholar] [CrossRef]
- D’Ippolito, G.; Diabira, S.; Howard, G.A.; Roos, B.A.; Schiller, P.C. Low oxygen tension inhibits osteogenic differentiation and enhances stemness of human MIAMI cells. Bone 2006, 39, 513–522. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, J.H.-C. Human Tendon Stem Cells Better Maintain Their Stemness in Hypoxic Culture Conditions. PLoS ONE 2013, 8, e61424. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Lin, L.; Zhou, Y.; Lu, X.; Shao, X.; Lin, C.; Yu, K.; Zhang, X.; Hong, J.; Chen, Y. Effect of Hypoxia on Self-Renewal Capacity and Differentiation in Human Tendon-Derived Stem Cells. Med. Sci. Monit. 2017, 23, 1334–1339. [Google Scholar] [CrossRef] [Green Version]
- Shojaee, A.; Parham, A. Strategies of tenogenic differentiation of equine stem cells for tendon repair: Current status and challenges. Stem Cell Res. Ther. 2019, 10, 1–13. [Google Scholar] [CrossRef]
- Chang, W.; Lee, C.Y.; Park, J.-H.; Park, M.-S.; Maeng, L.-S.; Yoon, C.S.; Lee, M.Y.; Hwang, K.-C.; Chung, Y.-A. Survival of hypoxic human mesenchymal stem cells is enhanced by a positive feedback loop involving miR-210 and hypoxia-inducible factor 1. J. Vet. Sci. 2013, 14, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Bavelloni, A.; Ramazzotti, G.; Poli, A.; Piazzi, M.; Focaccia, E.; Blalock, W.; Faenza, I. MiRNA-210: A Current Overview. Anticancer Res. 2017, 37. [Google Scholar] [CrossRef] [Green Version]
- Kelly, T.J.; Souza, A.L.; Clish, C.B.; Puigserver, P. A Hypoxia-Induced Positive Feedback Loop Promotes Hypoxia-Inducible Factor 1 Stability through miR-210 Suppression of Glycerol-3-Phosphate Dehydrogenase 1-Like. Mol. Cell. Biol. 2011, 31, 2696–2706. [Google Scholar] [CrossRef] [Green Version]
- Usman, M.A.; Nakasa, T.; Shoji, T.; Kato, T.; Kawanishi, Y.; Hamanishi, M.; Kamei, N.; Ochi, M. The effect of administration of double stranded MicroRNA-210 on acceleration of Achilles tendon healing in a rat model. J. Orthop. Sci. 2015, 20, 538–546. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.Y.; Chun, S.Y.; Ha, Y.-S.; Kim, D.H.; Kim, J.; Song, P.H.; Kim, H.T.; Yoo, E.S.; Kim, B.S.; Kwon, T.G. Hypoxia Enhances Cell Properties of Human Mesenchymal Stem Cells. Tissue Eng. Regen. Med. 2017, 14, 595–604. [Google Scholar] [CrossRef]
- Widowati, W.; Rihibiha, D.D.; Khiong, K.; Widodo, M.A.; Sumitro, S.B.; Bachtiar, I. Hypoxia in Mesenchymal Stem Cell. In Hypoxia and Human Diseases; Zheng, J., Zhou, C., Eds.; InTech: Vienna, Austria, 2017; ISBN 978-953-51-2895-3. [Google Scholar]
- Hung, S.-C.; Pochampally, R.R.; Chen, S.-C.; Hsu, S.-C.; Prockop, D.J. Angiogenic Effects of Human Multipotent Stromal Cell Conditioned Medium Activate the PI3K-Akt Pathway in Hypoxic Endothelial Cells to Inhibit Apoptosis, Increase Survival, and Stimulate Angiogenesis. Stem Cells 2007, 25, 2363–2370. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Xu, Y.; Zhao, J.; Zhang, Z.; Yang, R.; Xie, J.; Liu, X.; Qi, S. Conditioned Medium from Hypoxic Bone Marrow-Derived Mesenchymal Stem Cells Enhances Wound Healing in Mice. PLoS ONE 2014, 9, e96161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, C.-C.; Chen, Y.-J.; Yew, T.-L.; Chen, L.-L.; Wang, J.-Y.; Chiu, C.-H.; Hung, S.-C. Hypoxia inhibits senescence and maintains mesenchymal stem cell properties through down-regulation of E2A-p21 by HIF-TWIST. Blood 2011, 117, 459–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, T.-F.; Yew, T.-L.; Chiang, E.-R.; Ma, H.-L.; Hsu, C.-Y.; Hsu, S.; Hsu, Y.-T.; Hung, S.-C. Mesenchymal Stem Cells From a Hypoxic Culture Improve and Engraft Achilles Tendon Repair. Am. J. Sports Med. 2013, 41, 1117–1125. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Tang, Y.-L.; Liang, X.-H. EMT: A new vision of hypoxia promoting cancer progression. Cancer Biol. Ther. 2011, 11, 714–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joseph, J.P.; Harishankar, M.; Pillai, A.A.; Devi, A. Hypoxia induced EMT: A review on the mechanism of tumor progression and metastasis in OSCC. Oral Oncol. 2018, 80, 23–32. [Google Scholar] [CrossRef]
- Yeo, C.D.; Kang, N.; Choi, S.Y.; Na Kim, B.; Park, C.K.; Kim, J.W.; Kim, Y.K.; Kim, S.J. The role of hypoxia on the acquisition of epithelial-mesenchymal transition and cancer stemness: A possible link to epigenetic regulation. Korean J. Intern. Med. 2017, 32, 589–599. [Google Scholar] [CrossRef]
- Zaidi, M.; Fu, F.; Cojocari, D.; McKee, T.D.; Wouters, B. Quantitative Visualization of Hypoxia and Proliferation Gradients Within Histological Tissue Sections. Front. Bioeng. Biotechnol. 2019, 7, 397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buravkova, L.B.; Andreeva, E.R.; Gogvadze, V.; Zhivotovsky, B. Mesenchymal stem cells and hypoxia: Where are we? Mitochondrion 2014, 19, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Banes, A.J.; Tsuzaki, M.; Yamamoto, J.; Fischer, T.; Brigman, B.; Brown, T.; Miller, L. Mechanoreception at the cellular level: The detection, interpretation, and diversity of responses to mechanical signals. Biochem. Cell Biol. 1995, 73, 349–365. [Google Scholar] [CrossRef] [PubMed]
- Lavagnino, M.; Arnoczky, S.P.; Tian, T.; Vaupel, Z. Effect of Amplitude and Frequency of Cyclic Tensile Strain on the Inhibition of MMP-1 mRNA Expression in Tendon Cells: An In Vitro Study. Connect. Tissue Res. 2003, 44, 181–187. [Google Scholar] [CrossRef]
- Screen, H.R.C.; Shelton, J.C.; Bader, D.L.; Lee, D.A. Cyclic tensile strain upregulates collagen synthesis in isolated tendon fascicles. Biochem. Biophys. Res. Commun. 2005, 336, 424–429. [Google Scholar] [CrossRef]
- Govoni, M.; Muscari, C.; Lovecchio, J.; Guarnieri, C.; Giordano, E. Mechanical Actuation Systems for the Phenotype Commitment of Stem Cell-Based Tendon and Ligament Tissue Substitutes. Stem Cell Rev. Rep. 2015, 12, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Hannafin, J.A.; Arnoczky, S.P.; Hoonjan, A.; Torzilli, P.A. Effect of stress deprivation and cyclic tensile loading on the material and morphologic properties of canine flexor digitorum profundus tendon: Anin vitro study. J. Orthop. Res. 1995, 13, 907–914. [Google Scholar] [CrossRef]
- Archambault, J.M.; Wiley, J.P.; Bray, R.C. Exercise Loading of Tendons and the Development of Overuse Injuries. Sports Med. 1995, 20, 77–89. [Google Scholar] [CrossRef]
- Arnoczky, S.P.; Lavagnino, M.; Egerbacher, M. The mechanobiological aetiopathogenesis of tendinopathy: Is it the over-stimulation or the under-stimulation of tendon cells? Int. J. Exp. Pathol. 2007, 88, 217–226. [Google Scholar] [CrossRef]
- Magnusson, S.P.; Langberg, H.; Kjaer, M. The pathogenesis of tendinopathy: Balancing the response to loading. Nat. Rev. Rheumatol. 2010, 6, 262–268. [Google Scholar] [CrossRef]
- Wang, T.; Gardiner, B.S.; Lin, Z.; Rubenson, J.; Kirk, T.B.; Wang, A.; Xu, J.; Smith, D.W.; Lloyd, D.G.; Zheng, M.H. Bioreactor Design for Tendon/Ligament Engineering. Tissue Eng. Part B Rev. 2012, 19, 133–146. [Google Scholar] [CrossRef] [Green Version]
- Abousleiman, R.I.; Reyes, Y.; McFetridge, P.; Sikavitsas, V. Tendon Tissue Engineering Using Cell-Seeded Umbilical Veins Cultured in a Mechanical Stimulator. Tissue Eng. Part A 2009, 15, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Lin, Z.; Day, R.E.; Gardiner, B.S.; Landao-Bassonga, E.; Rubenson, J.; Kirk, T.B.; Smith, D.W.; Lloyd, D.G.; Hardisty, G.; et al. Programmable mechanical stimulation influences tendon homeostasis in a bioreactor system. Biotechnol. Bioeng. 2013, 110, 1495–1507. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.D.; Liang, R.; Woo, S.L.-Y.; Burton, S.D.; Wu, C.; Almarza, A.; Sacks, M.S.; Abramowitch, S. Effects of Cell Seeding and Cyclic Stretch on the Fiber Remodeling in an Extracellular Matrix–Derived Bioscaffold. Tissue Eng. Part A 2009, 15, 957–963. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Im, H.-J.; Wang, J.H.-C. Repetitive mechanical stretching modulates IL-1beta induced COX-2, MMP-1 expression, and PGE2 production in human patellar tendon fibroblasts. Gene 2005, 363, 166–172. [Google Scholar] [CrossRef] [Green Version]
- Webb, K.; Hitchcock, R.W.; Smeal, R.M.; Li, W.; Gray, S.D.; Tresco, P.A. Cyclic strain increases fibroblast proliferation, matrix accumulation, and elastic modulus of fibroblast-seeded polyurethane constructs. J. Biomech. 2006, 39, 1136–1144. [Google Scholar] [CrossRef]
- Cribb, A.M.; Scott, J.E. Tendon response to tensile stress: An ultrastructural investigation of collagen:proteoglycan interactions in stressed tendon. J. Anat. 1995, 187, 423–428. [Google Scholar]
- Juncosa-Melvin, N.; Shearn, J.T.; Boivin, G.P.; Gooch, C.; Galloway, M.T.; West, J.R.; Nirmalanandhan, V.S.; Bradica, G.; Butler, D. Effects of Mechanical Stimulation on the Biomechanics and Histology of Stem Cell–Collagen Sponge Constructs for Rabbit Patellar Tendon Repair. Tissue Eng. 2006, 12, 2291–2300. [Google Scholar] [CrossRef]
- Woon, C.Y.; Kraus, A.; Raghavan, S.S.; Pridgen, B.C.; Megerle, K.; Pham, H.; Chang, J. Three-Dimensional-Construct Bioreactor Conditioning in Human Tendon Tissue Engineering. Tissue Eng. Part A 2011, 17, 2561–2572. [Google Scholar] [CrossRef]
- Saber, S.; Zhang, A.Y.; Ki, S.H.; Lindsey, D.P.; Smith, R.L.; Riboh, J.; Pham, H.; Chang, J. Flexor Tendon Tissue Engineering: Bioreactor Cyclic Strain Increases Construct Strength. Tissue Eng. Part A 2010, 16, 2085–2090. [Google Scholar] [CrossRef]
- Butler, D.; Hunter, S.A.; Chokalingam, K.; Cordray, M.J.; Shearn, J.; Juncosa-Melvin, N.; Nirmalanandhan, S.; Jain, A. Using Functional Tissue Engineering and Bioreactors to Mechanically Stimulate Tissue-Engineered Constructs. Tissue Eng. Part A 2009, 15, 741–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberton, P.; Popov, C.; Prägert, M.; Kohler, J.; Shukunami, C.; Schieker, M.; Docheva, D. Conversion of Human Bone Marrow-Derived Mesenchymal Stem Cells into Tendon Progenitor Cells by Ectopic Expression of Scleraxis. Stem Cells Dev. 2012, 21, 846–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagchi, R.; Roche, P.; Aroutiounova, N.; Espira, L.; Abrenica, B.; Schweitzer, R.; Czubryt, M.P. The transcription factor scleraxis is a critical regulator of cardiac fibroblast phenotype. BMC Biol. 2016, 14, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, A.; Danielson, P.; Abraham, T.; Fong, G.; Sampaio, A.V.; Underhill, T.M. Mechanical force modulates scleraxis expression in bioartificial tendons. J. Musculoskelet. Neuronal. Interact. 2011, 11, 124–132. [Google Scholar]
- Maeda, T.; Sakabe, T.; Sunaga, A.; Sakai, K.; Rivera, A.L.; Keene, D.R.; Sasaki, T.; Stavnezer, E.; Iannotti, J.; Schweitzer, R.; et al. Conversion of Mechanical Force into TGF-β-Mediated Biochemical Signals. Curr. Biol. 2011, 21, 933–941. [Google Scholar] [CrossRef] [Green Version]
- Doroski, D.M.; Levenston, M.E.; Temenoff, J.S. Cyclic Tensile Culture Promotes Fibroblastic Differentiation of Marrow Stromal Cells Encapsulated in Poly(Ethylene Glycol)-Based Hydrogels. Tissue Eng. Part A 2010, 16, 3457–3466. [Google Scholar] [CrossRef] [Green Version]
- Burk, J.; Plenge, A.; Brehm, W.; Heller, S.; Pfeiffer, B.; Kasper, C. Induction of Tenogenic Differentiation Mediated by Extracellular Tendon Matrix and Short-Term Cyclic Stretching. Stem Cells Int. 2016, 2016, 7342379. [Google Scholar] [CrossRef] [Green Version]
- Im, G.-I. Clinical use of stem cells in orthopaedics. Eur. Cells Mater. 2017, 33, 183–196. [Google Scholar] [CrossRef]
- Deniz, P.; Guler, S.; Celik, E.; Hosseinian, P.; Aydin, H.M. Use of cyclic strain bioreactor for the upregulation of key tenocyte gene expression on Poly(glycerol-sebacate) (PGS) sheets. Mater. Sci. Eng. C 2020, 106, 110293. [Google Scholar] [CrossRef]
- Nam, H.Y.; Pingguan-Murphy, B.; Abbas, A.A.; Merican, A.M.; Kamarul, T. Uniaxial Cyclic Tensile Stretching at 8% Strain Exclusively Promotes Tenogenic Differentiation of Human Bone Marrow-Derived Mesenchymal Stromal Cells. Stem Cells Int. 2019, 2019, 9723025. [Google Scholar] [CrossRef] [Green Version]
- Cao, D.; Liu, W.; Wei, X.; Xu, F.; Cui, L.; Cao, Y. In Vitro Tendon Engineering with Avian Tenocytes and Polyglycolic Acids: A Preliminary Report. Tissue Eng. 2006, 12, 1369–1377. [Google Scholar] [CrossRef] [PubMed]
- Barkhausen, T.; Van Griensven, M.; Zeichen, J.; Bosch, U. Modulation of cell functions of human tendon fibroblasts by different repetitive cyclic mechanical stress patterns. Exp. Toxicol. Pathol. 2003, 55, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Waldorff, E.I.; Zhang, N.; Ryaby, J.T. Pulsed electromagnetic field applications: A corporate perspective. J. Orthop. Transl. 2017, 9, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Tucker, J.J.; Cirone, J.M.; Morris, T.R.; Nuss, C.A.; Huegel, J.; Waldorff, E.I.; Zhang, N.; Ryaby, J.T.; Soslowsky, L.J. Pulsed electromagnetic field therapy improves tendon-to-bone healing in a rat rotator cuff repair model. J. Orthop. Res. 2016, 35, 902–909. [Google Scholar] [CrossRef]
- Liu, M.; Lee, C.; Laron, D.; Zhang, N.; Waldorff, E.I.; Ryaby, J.T.; Feeley, B.; Liu, X. Role of pulsed electromagnetic fields (PEMF) on tenocytes and myoblasts-potential application for treating rotator cuff tears. J. Orthop. Res. 2017, 35, 956–964. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.-J. Extracorporeal shockwave therapy in musculoskeletal disorders. J. Orthop. Surg. Res. 2012, 7, 11. [Google Scholar] [CrossRef] [Green Version]
- Ioppolo, F.; Rompe, J.D.; Furia, J.P.; Cacchio, A. Clinical application of shock wave therapy (SWT) in musculoskeletal disorders. Eur. J. Phys. Rehabil. Med. 2014, 50, 217–230. [Google Scholar]
- Furia, J.P.; Rompe, J.-D.; Maffulli, N.; Cacchio, A.; Schmitz, C. Radial Extracorporeal Shock Wave Therapy Is Effective and Safe in Chronic Distal Biceps Tendinopathy. Clin. J. Sport Med. 2017, 27, 430–437. [Google Scholar] [CrossRef]
- Frizziero, A.; Vittadini, F.; Barazzuol, M.; Gasparre, G.; Finotti, P.; Meneghini, A.; Maffulli, N.; Masiero, S. Extracorporeal shockwaves therapy versus hyaluronic acid injection for the treatment of painful non-calcific rotator cuff tendinopathies: Preliminary results. J. Sports Med. Phys. Fit. 2017. [Google Scholar] [CrossRef]
- Notarnicola, A.; Moretti, B. The biological effects of extracorporeal shock wave therapy (eswt) on tendon tissue. Muscle Ligaments Tendons J. 2012, 2, 33–37. [Google Scholar]
- Cricchio, V.; Best, M.; Reverchon, E.; Maffulli, N.; Phillips, G.; Santin, M.; Della Porta, G. Novel Superparamagnetic Microdevices Based on Magnetized PLGA/PLA Microparticles Obtained by Supercritical Fluid Emulsion and Coating by Carboxybetaine-Functionalized Chitosan Allowing the Tuneable Release of Therapeutics. J. Pharm. Sci. 2017, 106, 2097–2105. [Google Scholar] [CrossRef]
- Matos, A.M.; Gonçalves, A.I.; El Haj, A.J.; Gomes, M.E. Magnetic biomaterials and nano-instructive tools as mediators of tendon mechanotransduction. Nanoscale Adv. 2020, 2, 140–148. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, A.I.; Miranda, M.S.; Rodrigues, M.T.; Reis, R.L.; Gomes, M.E. Magnetic responsive cell-based strategies for diagnostics and therapeutics. Biomed. Mater. 2018, 13, 054001. [Google Scholar] [CrossRef] [PubMed]
- Santo, V.E.; Rodrigues, M.T.; Gomes, M.E. Contributions and future perspectives on the use of magnetic nanoparticles as diagnostic and therapeutic tools in the field of regenerative medicine. Expert Rev. Mol. Diagn. 2013, 13, 553–566. [Google Scholar] [CrossRef] [PubMed]
- Sapir-Lekhovitser, Y.; Rotenberg, M.Y.; Jopp, J.; Friedman, G.; Polyak, B.; Cohen, S. Magnetically actuated tissue engineered scaffold: Insights into mechanism of physical stimulation. Nanoscale 2016, 8, 3386–3399. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, A.I.; Rodrigues, M.T.; Gomes, M.E. Tissue-engineered magnetic cell sheet patches for advanced strategies in tendon regeneration. Acta Biomater. 2017, 63, 110–122. [Google Scholar] [CrossRef]
- Tomás, A.R.; Gonçalves, A.I.; Paz, E.; Freitas, P.P.; Domingues, R.M.A.; Gomes, M.E. Magneto-mechanical actuation of magnetic responsive fibrous scaffolds boosts tenogenesis of human adipose stem cells. Nanoscale 2019, 11, 18255–18271. [Google Scholar] [CrossRef]
- Cartmell, S.H.; Keramane, A.; Kirkham, G.R.; Verschueren, S.B.; Magnay, J.L.; El Haj, A.J.; Dobson, J. Use of magnetic particles to apply mechanical forces for bone tissue engineering purposes. J. Phys. Conf. Ser. 2005, 17, 77–80. [Google Scholar] [CrossRef]
- Markides, H.; McLaren, J.S.; El Haj, A.J. Overcoming translational challenges—The delivery of mechanical stimuli in vivo. Int. J. Biochem. Cell Biol. 2015, 69, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.; Reis, R.L.; Gomes, M.E. Harnessing magnetic-mechano actuation in regenerative medicine and tissue engineering. Trends Biotechnol. 2015, 33, 471–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, R.; Markides, H.; Morris, R.H.; Richards, P.; El Haj, A.J.; Sottile, V. Autonomous magnetic labelling of functional mesenchymal stem cells for improved traceability and spatial control in cell therapy applications. J. Tissue Eng. Regen. Med. 2016, 11, 2333–2348. [Google Scholar] [CrossRef]
- Harrison, R.; Leija, H.A.L.; Strohbuecker, S.; Crutchley, J.; Marsh, S.; Denning, C.; El Haj, A.; Sottile, V. Development and validation of broad-spectrum magnetic particle labelling processes for cell therapy manufacturing. Stem Cell Res. Ther. 2018, 9, 248. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, A.I.; Rotherham, M.; Markides, H.; Rodrigues, M.T.; Reis, R.; Gomes, M.E.; El Haj, A. Triggering the activation of Activin A type II receptor in human adipose stem cells towards tenogenic commitment using mechanomagnetic stimulation. Nanomed. Nanotechnol. Biol. Med. 2018, 14, 1149–1159. [Google Scholar] [CrossRef]
- Rinella, L.; Marano, F.; Paletto, L.; Fraccalvieri, M.; Annaratone, L.; Castellano, I.; Fortunati, N.; Bargoni, A.; Berta, L.; Frairia, R.; et al. Extracorporeal shock waves trigger tenogenic differentiation of human adipose-derived stem cells. Connect. Tissue Res. 2018, 59, 1–13. [Google Scholar] [CrossRef]
- Leone, L.; Vetrano, M.; Ranieri, D.; Raffa, S.; Vulpiani, M.C.; Ferretti, A.; Torrisi, M.R.; Visco, V. Extracorporeal Shock Wave Treatment (ESWT) Improves In Vitro Functional Activities of Ruptured Human Tendon-Derived Tenocytes. PLoS ONE 2012, 7, e49759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leone, L.; Raffa, S.; Vetrano, M.; Ranieri, D.; Malisan, F.; Scrofani, C.; Vulpiani, M.C.; Ferretti, A.; Torrisi, M.R.; Visco, V. Extracorporeal Shock Wave Treatment (ESWT) enhances the in vitro-induced differentiation of human tendon-derived stem/progenitor cells (hTSPCs). Oncotarget 2016, 7, 6410–6423. [Google Scholar] [CrossRef] [Green Version]
- Carleton, J.B.; Rodin, G.J.; Sacks, M.S. Layered Elastomeric Fibrous Scaffolds: An In-Silico Study of the Achievable Range of Mechanical Behaviors. ACS Biomater. Sci. Eng. 2017, 3, 2907–2921. [Google Scholar] [CrossRef]
- Chen, J.; Xu, J.; Wang, A.; Zheng, M. Scaffolds for tendon and ligament repair: Review of the efficacy of commercial products. Expert Rev. Med Devices 2009, 6, 61–73. [Google Scholar] [CrossRef]
- Moshiri, A.; Oryan, A. Role of tissue engineering in tendon reconstructive surgery and regenerative medicine: Current concepts, approaches and concerns. Hard Tissue 2012, 1. [Google Scholar] [CrossRef] [Green Version]
- Longo, U.G.; Lamberti, A.; Petrillo, S.; Maffulli, N.; Denaro, V. Scaffolds in Tendon Tissue Engineering. Stem Cells Int. 2012, 2012, 517165. [Google Scholar] [CrossRef] [Green Version]
- Kishore, V.; Bullock, W.; Sun, X.; Van Dyke, W.S.; Akkus, O. Tenogenic differentiation of human MSCs induced by the topography of electrochemically aligned collagen threads. Biomaterials 2012, 33, 2137–2144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AlShomer, F.; Chaves, C.; Kalaskar, D.M. Advances in Tendon and Ligament Tissue Engineering: Materials Perspective. J. Mater. 2018, 2018, 9868151. [Google Scholar] [CrossRef] [Green Version]
- Dong, C.; Lv, Y. Application of Collagen Scaffold in Tissue Engineering: Recent Advances and New Perspectives. Polymers 2016, 8, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhi, Y.; Liu, W.; Zhang, P.; Jiang, J.; Chen, S. Electrospun silk fibroin mat enhances tendon-bone healing in a rabbit extra-articular model. Biotechnol. Lett. 2016, 38, 1827–1835. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.-Y.; Chung, J.-W.; Park, H.-J.; Jiang, Y.-Y.; Park, J.-K.; Seo, Y.-K. Silk and collagen scaffolds for tendon reconstruction. Proc. Inst. Mech. Eng. Part H J. Eng. Med. 2014, 228, 388–396. [Google Scholar] [CrossRef]
- Seo, Y.-K.; Kim, J.-H.; Eo, S.-R. Co-effect of silk and amniotic membrane for tendon repair. J. Biomater. Sci. Polym. Ed. 2016, 27, 1232–1247. [Google Scholar] [CrossRef]
- Maghdouri-White, Y.; Petrova, S.; Sori, N.; Polk, S.; Wriggers, H.; Ogle, R.; Ogle, R.; Francis, M. Electrospun silk–collagen scaffolds and BMP-13 for ligament and tendon repair and regeneration. Biomed. Phys. Eng. Express 2018, 4, 025013. [Google Scholar] [CrossRef]
- Sahoo, S.; Toh, S.L.; Goh, J.C. A bFGF-releasing silk/PLGA-based biohybrid scaffold for ligament/tendon tissue engineering using mesenchymal progenitor cells. Biomaterials 2010, 31, 2990–2998. [Google Scholar] [CrossRef]
- Chen, E.; Yang, L.; Ye, C.; Zhang, W.; Ran, J.; Xue, D.; Wang, Z.; Pan, Z.; Hu, Q. An asymmetric chitosan scaffold for tendon tissue engineering: In vitro and in vivo evaluation with rat tendon stem/progenitor cells. Acta Biomater. 2018, 73, 377–387. [Google Scholar] [CrossRef]
- Rodríguez-Vázquez, M.; Vega-Ruiz, B.; Ramos-Zúñiga, R.; Saldaña-Koppel, D.A.; Quiñones-Olvera, L.F. Chitosan and Its Potential Use as a Scaffold for Tissue Engineering in Regenerative Medicine. BioMed Res. Int. 2015, 2015, 821279. [Google Scholar] [CrossRef] [Green Version]
- Leung, M.; Jana, S.; Tsao, C.-T.; Zhang, M. Tenogenic differentiation of human bone marrow stem cells via a combinatory effect of aligned chitosan-poly-caprolactone nanofibers and TGF-β3. J. Mater. Chem. B 2013, 1, 6516–6524. [Google Scholar] [CrossRef] [PubMed]
- Beldjilali-Labro, M.; García, A.G.; Farhat, F.; Bedoui, F.; Grosset, J.-F.; Dufresne, M.; Legallais, C. Biomaterials in Tendon and Skeletal Muscle Tissue Engineering: Current Trends and Challenges. Materials 2018, 11, 1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.J.; Kasper, F.K.; Mikos, A.G. Biomaterials for Tissue Engineering. Ann. Biomed. Eng. 2013, 42, 323–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, W.L.; Liau, L.L.; Ng, M.H.; Chowdhury, S.R.; Law, J.X. Current Progress in Tendon and Ligament Tissue Engineering. Tissue Eng. Regen. Med. 2019, 16, 549–571. [Google Scholar] [CrossRef] [PubMed]
- Shearn, J.T.; Kinneberg, K.R.; Dyment, N.A.; Galloway, M.T.; Kenter, K.; Wylie, C.; Butler, D.L. Tendon tissue engineering: Progress, challenges, and translation to the clinic. J. Musculoskelet. Neuronal. Interact. 2011, 11, 163–173. [Google Scholar]
- El Khatib, M.; Mauro, A.; Wyrwa, R.; Di Mattia, M.; Turriani, M.; Di Giacinto, O.; Kretzschmar, B.S.; Seemann, T.; Valbonetti, L.; Berardinelli, P.; et al. Fabrication and Plasma Surface Activation of Aligned Electrospun PLGA Fiber Fleeces with Improved Adhesion and Infiltration of Amniotic Epithelial Stem Cells Maintaining their Teno-inductive Potential. Molecules 2020, 25, 3176. [Google Scholar] [CrossRef]
- El Khatib, M.; Mauro, A.; Di Mattia, M.; Wyrwa, R.; Schweder, M.; Ancora, M.; Lazzaro, F.; Berardinelli, P.; Valbonetti, L.; Di Giacinto, O.; et al. Electrospun PLGA Fiber Diameter and Alignment of Tendon Biomimetic Fleece Potentiate Tenogenic Differentiation and Immunomodulatory Function of Amniotic Epithelial Stem Cells. Cells 2020, 9, 1207. [Google Scholar] [CrossRef]
- Bashur, C.A.; Shaffer, R.D.; Dahlgren, L.A.; Guelcher, S.A.; Goldstein, A.S. Effect of Fiber Diameter and Alignment of Electrospun Polyurethane Meshes on Mesenchymal Progenitor Cells. Tissue Eng. Part A 2009, 15, 2435–2445. [Google Scholar] [CrossRef]
- Cardwell, R.D.; Dahlgren, L.A.; Goldstein, A.S. Electrospun fibre diameter, not alignment, affects mesenchymal stem cell differentiation into the tendon/ligament lineage. J. Tissue Eng. Regen. Med. 2012, 8, 937–945. [Google Scholar] [CrossRef]
- Lee, C.H.; Shin, H.J.; Cho, I.H.; Kang, Y.-M.; Kim, I.A.; Park, K.-D.; Shin, J.-W. Nanofiber alignment and direction of mechanical strain affect the ECM production of human ACL fibroblast. Biomaterials 2005, 26, 1261–1270. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, X.; Zhang, E.; Yang, L.; Yuan, H.; Tu, W.; Zhang, H.; Yin, Z.; Shen, W.L.; Chen, X.; et al. An epigenetic bioactive composite scaffold with well-aligned nanofibers for functional tendon tissue engineering. Acta Biomater. 2018, 66, 141–156. [Google Scholar] [CrossRef] [PubMed]
- Nitti, P.; Gallo, N.; Natta, L.; Scalera, F.; Palazzo, B.; Sannino, A.; Gervaso, F. Influence of Nanofiber Orientation on Morphological and Mechanical Properties of Electrospun Chitosan Mats. J. Healthc. Eng. 2018, 2018, 3651480. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yuan, H.; Liu, H.; Chen, X.; Lu, P.; Zhu, T.; Yang, L.; Yin, Z.; Heng, B.C.; Zhang, Y.; et al. Well-aligned chitosan-based ultrafine fibers committed teno-lineage differentiation of human induced pluripotent stem cells for Achilles tendon regeneration. Biomaterials 2015, 53, 716–730. [Google Scholar] [CrossRef] [PubMed]
- Müller, S.A.; Dürselen, L.; Heisterbach, P.; Evans, C.; Majewski, M. Effect of a Simple Collagen Type I Sponge for Achilles Tendon Repair in a Rat Model. Am. J. Sports Med. 2016, 44, 1998–2004. [Google Scholar] [CrossRef]
- Zhang, B.; Luo, Q.; Deng, B.; Morita, Y.; Ju, Y.; Song, G. Construction of tendon replacement tissue based on collagen sponge and mesenchymal stem cells by coupled mechano-chemical induction and evaluation of its tendon repair abilities. Acta Biomater. 2018, 74, 247–259. [Google Scholar] [CrossRef]
- Gabler, C.; Saß, J.-O.; Gierschner, S.; Lindner, T.; Bader, R.; Tischer, T. In Vivo Evaluation of Different Collagen Scaffolds in an Achilles Tendon Defect Model. BioMed Res. Int. 2018, 2018, 6432742. [Google Scholar] [CrossRef]
- Qian, S.; Wang, Z.; Zheng, Z.; Ran, J.; Zhu, J.; Chen, W. A Collagen and Silk Scaffold for Improved Healing of the Tendon and Bone Interface in a Rabbit Model. Med Sci. Monit. 2019, 25, 269–278. [Google Scholar] [CrossRef]
- Della Porta, G.; Reverchon, E.; Maffulli, N. Biomaterials and Supercritical Fluid Technologies: Which Perspectives to Fabricate Artificial Extracellular Matrix? Curr. Pharm. Des. 2017, 23, 3759–3771. [Google Scholar] [CrossRef]
- Della Porta, G.; Ciardulli, M.C.; Maffulli, N. Microcapsule Technology for Controlled Growth Factor Release in Musculoskeletal Tissue Engineering. Sports Med. Arthrosc. Rev. 2018, 26, e2–e9. [Google Scholar] [CrossRef]
- Zafar, A.; Rushton, N. Extruded collagen fibres and biphasic collagen sponge for rotator cuff tendon repair application: An ovine in vitro study. Muscle Ligaments Tendons J. 2019, 8, 544. [Google Scholar] [CrossRef] [Green Version]
- Younesi, M.; Islam, A.; Kishore, V.; Anderson, J.M.; Akkus, O. Tenogenic Induction of Human MSCs by Anisotropically Aligned Collagen Biotextiles. Adv. Funct. Mater. 2014, 24, 5762–5770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, A.; Mbimba, T.; Younesi, M.; Akkus, O. Effects of substrate stiffness on the tenoinduction of human mesenchymal stem cells. Acta Biomater. 2017, 58, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Full, S.M.; Delman, C.; Gluck, J.M.; Abdmaulen, R.; Shemin, R.J.; Heydarkhan-Hagvall, S. Effect of fiber orientation of collagen-based electrospun meshes on human fibroblasts for ligament tissue engineering applications. J. Biomed. Mater. Res. Part B Appl. Biomater. 2014, 103, 39–46. [Google Scholar] [CrossRef]
- Meimandi-Parizi, A.; Oryan, A.; Moshiri, A. Role of tissue engineered collagen based tridimensional implant on the healing response of the experimentally induced large Achilles tendon defect model in rabbits: A long term study with high clinical relevance. J. Biomed. Sci. 2013, 20, 28. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, C.M.; Holmes, B.; Faucett, S.; Zhang, L.G. Three-Dimensional Printing of Nanomaterial Scaffolds for Complex Tissue Regeneration. Tissue Eng. Part B Rev. 2014, 21, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Wu, S.; Kuss, M.; Kong, Y.; Shi, W.; Streubel, P.N.; Li, T.; Duan, B. 3D printing of multilayered scaffolds for rotator cuff tendon regeneration. Bioact. Mater. 2020, 5, 636–643. [Google Scholar] [CrossRef]
- Touré, A.B.R.; Mele, E.; Christie, J.K. Multi-layer Scaffolds of Poly(caprolactone), Poly(glycerol sebacate) and Bioactive Glasses Manufactured by Combined 3D Printing and Electrospinning. Nanomaterials 2020, 10, 626. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.; Lee, H.; Lee, J.; Atala, A.; Yoo, J.J.; Lee, S.J.; Kim, G.H. Efficient myotube formation in 3D bioprinted tissue construct by biochemical and topographical cues. Biomaterials 2019, 230, 119632. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, Z.; Fuh, J.Y.H.; Wong, Y.S.; Wang, W.; Thian, E.S. Direct E-jet printing of three-dimensional fibrous scaffold for tendon tissue engineering. J. Biomed. Mater. Res. Part B Appl. Biomater. 2015, 105, 616–627. [Google Scholar] [CrossRef]
- Do, A.-V.; Khorsand, B.; Geary, S.M.; Salem, A.K. 3D Printing of Scaffolds for Tissue Regeneration Applications. Adv. Healthc. Mater. 2015, 4, 1742–1762. [Google Scholar] [CrossRef] [Green Version]
- Merceron, T.K.; Burt, M.; Seol, Y.-J.; Kang, H.-W.; Lee, S.J.; Yoo, J.J.; Atala, A. A 3D bioprinted complex structure for engineering the muscle–tendon unit. Biofabrication 2015, 7, 035003. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, S.; Kuss, M.A.; Streubel, P.N.; Duan, B. Effects of Hydroxyapatite and Hypoxia on Chondrogenesis and Hypertrophy in 3D Bioprinted ADMSC Laden Constructs. ACS Biomater. Sci. Eng. 2017, 3, 826–835. [Google Scholar] [CrossRef]
- Khorshidi, S.; Solouk, A.; Mirzadeh, H.; Mazinani, S.; Lagarón, J.M.; Sharifi, S.; Ramakrishna, S. A review of key challenges of electrospun scaffolds for tissue-engineering applications. J. Tissue Eng. Regen. Med. 2015, 10, 715–738. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Wu, T.; Dai, Y.; Xia, Y. Electrospinning and Electrospun Nanofibers: Methods, Materials, and Applications. Chem. Rev. 2019, 119, 5298–5415. [Google Scholar] [CrossRef] [PubMed]
- Repanas, A.; Andriopoulou, S.; Glasmacher, B. The significance of electrospinning as a method to create fibrous scaffolds for biomedical engineering and drug delivery applications. J. Drug Deliv. Sci. Technol. 2016, 31, 137–146. [Google Scholar] [CrossRef]
- Jiang, T.; Carbone, E.J.; Lo, K.W.-H.; Laurencin, C.T. Electrospinning of polymer nanofibers for tissue regeneration. Prog. Polym. Sci. 2015, 46, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Ding, X.; Zhou, G.; Li, P.; Wei, X.; Fan, Y. Electrospinning of Nanofibers for Tissue Engineering Applications. J. Nanomater. 2013, 2013, 495708. [Google Scholar] [CrossRef] [Green Version]
- Lannutti, J.J.; Reneker, D.; Ma, T.; Tomasko, D.; Farson, D. Electrospinning for tissue engineering scaffolds. Mater. Sci. Eng. C 2007, 27, 504–509. [Google Scholar] [CrossRef]
- Chen, M.; Patra, P.K.; Warner, S.B.; Bhowmick, S. Optimization of Electrospinning Process Parameters for Tissue Engineering Scaffolds. Biophys. Rev. Lett. 2006, 1, 153–178. [Google Scholar] [CrossRef]
- Rothrauff, B.B.; Lauro, B.B.; Yang, G.; Debski, R.E.; Musahl, V.; Tuan, R.S. Braided and Stacked Electrospun Nanofibrous Scaffolds for Tendon and Ligament Tissue Engineering. Tissue Eng. Part A 2017, 23, 378–389. [Google Scholar] [CrossRef]
- Orr, S.B.; Chainani, A.; Hippensteel, K.J.; Kishan, A.; Gilchrist, C.; Garrigues, N.W.; Ruch, D.S.; Guilak, F.; Little, D. Aligned multilayered electrospun scaffolds for rotator cuff tendon tissue engineering. Acta Biomater. 2015, 24, 117–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Z.; Chen, X.; Chen, J.; Shen, W.-L.; Nguyen, T.M.H.; Gao, L.; Ouyang, H.W. The regulation of tendon stem cell differentiation by the alignment of nanofibers. Biomaterials 2010, 31, 2163–2175. [Google Scholar] [CrossRef] [PubMed]
- Moffat, K.L.; Kwei, A.S.-P.; Spalazzi, J.P.; Doty, S.B.; Levine, W.N.; Lu, H.H. Novel Nanofiber-Based Scaffold for Rotator Cuff Repair and Augmentation. Tissue Eng. Part A 2009, 15, 115–126. [Google Scholar] [CrossRef] [Green Version]
- Sensini, A.; Gualandi, C.; Zucchelli, A.; Boyle, L.A.; Kao, A.P.; Reilly, G.C.; Tozzi, G.; Cristofolini, L.; Focarete, M.L. Tendon Fascicle-Inspired Nanofibrous Scaffold of Polylactic acid/Collagen with Enhanced 3D-Structure and Biomechanical Properties. Sci. Rep. 2018, 8, 17167. [Google Scholar] [CrossRef]
- Zheng, Z.; Ran, J.; Chen, W.; Hu, Y.; Zhu, T.; Chen, X.; Yin, Z.; Heng, B.C.; Feng, G.; Le, H.; et al. Alignment of collagen fiber in knitted silk scaffold for functional massive rotator cuff repair. Acta Biomater 2017, 51, 317–329. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Wang, Y.; Streubel, P.N.; Duan, B. Living nanofiber yarn-based woven biotextiles for tendon tissue engineering using cell tri-culture and mechanical stimulation. Acta Biomater. 2017, 62, 102–115. [Google Scholar] [CrossRef] [PubMed]
- Czaplewski, S.K.; Tsai, T.-L.; Duenwald-Kuehl, S.; Vanderby, R.; Li, W.-J. Tenogenic differentiation of human induced pluripotent stem cell-derived mesenchymal stem cells dictated by properties of braided submicron fibrous scaffolds. Biomaterials 2014, 35, 6907–6917. [Google Scholar] [CrossRef]
- Erisken, C.; Zhang, X.; Moffat, K.L.; Levine, W.N.; Lu, H.H. Scaffold Fiber Diameter Regulates Human Tendon Fibroblast Growth and Differentiation. Tissue Eng. Part A 2012, 19, 519–528. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.H.; Kim, M.J.; Ryu, S.J.; Ki, C.-S.; Park, Y.H. Effect of fiber diameter on surface morphology, mechanical property, and cell behavior of electrospun poly(ε-caprolactone) mat. Fibers Polym. 2016, 17, 1033–1042. [Google Scholar] [CrossRef]
- Subramony, S.D.; Dargis, B.R.; Castillo, M.; Azeloglu, E.U.; Tracey, M.S.; Su, A.; Lu, H.H. The guidance of stem cell differentiation by substrate alignment and mechanical stimulation. Biomaterials 2013, 34, 1942–1953. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.M.; Erisken, C.; Iskratsch, T.; Sheetz, M.; Levine, W.N.; Lu, H.H. Polymer fiber-based models of connective tissue repair and healing. Biomaterials 2016, 112, 303–312. [Google Scholar] [CrossRef] [Green Version]
- Longo, U.G.; Franceschi, F.; Berton, A.; Maffulli, N.; Droena, V. Conservative Treatment and Rotator Cuff Tear Progression. In Medicine and Sport Science; Maffulli, N., Ed.; KARGER: Basel, Switzerland, 2011; Volume 57, pp. 90–99. ISBN 978-3-8055-9815-6. [Google Scholar]
- Liu, Y.; Suen, C.-W.; Zhang, J.-F.; Li, G. Current concepts on tenogenic differentiation and clinical applications. J. Orthop. Transl. 2017, 9, 28–42. [Google Scholar] [CrossRef]
- Yan, Z.; Yin, H.; Nerlich, M.; Pfeifer, C.G.; Docheva, D. Boosting tendon repair: Interplay of cells, growth factors and scaffold-free and gel-based carriers. J. Exp. Orthop. 2018, 5, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, M.; Angele, P.; Järvinen, T.A.; Docheva, D. Rescue plan for Achilles: Therapeutics steering the fate and functions of stem cells in tendon wound healing. Adv. Drug Deliv. Rev. 2018, 129, 352–375. [Google Scholar] [CrossRef]
- James, R.; Kesturu, G.; Balian, G.; Chhabra, A.B. Tendon: Biology, Biomechanics, Repair, Growth Factors, and Evolving Treatment Options. J. Hand Surg. 2008, 33, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-J.; Chen, X.; Li, G.; Chan, K.-M.; Heng, B.C.; Yin, Z.; Ouyang, H. Concise Review: Stem Cell Fate Guided By Bioactive Molecules for Tendon Regeneration. Stem Cells Transl. Med. 2018, 7, 404–414. [Google Scholar] [CrossRef]
- Gaut, L.; Duprez, D. Tendon development and diseases. Wiley Interdiscip. Rev. Dev. Biol. 2015, 5, 5–23. [Google Scholar] [CrossRef] [Green Version]
- Jones, E.R.; Jones, G.C.; Legerlotz, K.; Riley, G.P. Corrigendum to “Cyclical strain modulates metalloprotease and matrix gene expression in human tenocytes via activation of TGFβ” [Biochim. Biophys. Acta (2013) 2596–2607]. Biochim. Biophys. Acta (BBA) Bioenerg. 2013, 1833, 3445–3446. [Google Scholar] [CrossRef]
- Holladay, C.A.; Abbah, A.A.; O’Dowd, C.; Pandit, A.; Zeugolis, D.I. Preferential tendon stem cell response to growth factor supplementation. J. Tissue Eng. Regen. Med. 2014, 10, 783–798. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.P.; Galassi, T.V.; Stoppato, M.; Schiele, N.R.; Kuo, C.K. Comparative analysis of mesenchymal stem cell and embryonic tendon progenitor cell response to embryonic tendon biochemical and mechanical factors. Stem Cell Res. Ther. 2015, 6, 89. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, A.I.; Rodrigues, M.T.; Lee, S.J.; Atala, A.; Yoo, J.J.; Reis, R.L.; Gomes, M.E. Understanding the Role of Growth Factors in Modulating Stem Cell Tenogenesis. PLoS ONE 2013, 8, e83734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, M.-T.; Yang, H.-W.; Chen, T.-H.; Lee, O.K.-S. Modulation of Proliferation and Differentiation of Human Anterior Cruciate Ligament–Derived Stem Cells by Different Growth Factors. Tissue Eng. Part A 2009, 15, 3979–3989. [Google Scholar] [CrossRef] [PubMed]
- Schneider, P.R.A.; Buhrmann, C.; Mobasheri, A.; Matis, U.; Shakibaei, M. Three-dimensional high-density co-culture with primary tenocytes induces tenogenic differentiation in mesenchymal stem cells. J. Orthop. Res. 2011, 29, 1351–1360. [Google Scholar] [CrossRef] [PubMed]
- Theiss, F.; Mirsaidi, A.; Mhanna, R.; Kümmerle, J.M.; Glanz, S.; Bahrenberg, G.; Tiaden, A.; Richards, P.J. Use of biomimetic microtissue spheroids and specific growth factor supplementation to improve tenocyte differentiation and adaptation to a collagen-based scaffold in vitro. Biomaterials 2015, 69, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Feng, L.; Xu, J.; Yang, Z.; Wu, T.; Zhang, J.; Shi, L.; Zhu, D.; Zhang, J.-F.; Li, G. MiR-378a suppresses tenogenic differentiation and tendon repair by targeting at TGF-β2. Stem Cell Res. Ther. 2019, 10, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orfei, C.P.; Viganò, M.; Pearson, J.R.; Colombini, A.; De Luca, P.; Ragni, E.; Santos-Ruiz, L.; De Girolamo, L. In Vitro Induction of Tendon-Specific Markers in Tendon Cells, Adipose- and Bone Marrow-Derived Stem Cells is Dependent on TGFβ3, BMP-12 and Ascorbic Acid Stimulation. Int. J. Mol. Sci. 2019, 20, 149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferdous, Z.; Wei, V.M.; Iozzo, R.V.; Höök, M.; Grande-Allen, K.J. Decorin-transforming Growth Factor-β Interaction Regulates Matrix Organization and Mechanical Characteristics of Three-dimensional Collagen Matrices. J. Biol. Chem. 2007, 282, 35887–35898. [Google Scholar] [CrossRef] [Green Version]
- Barsby, T.; Guest, D.; Guest, D. Transforming Growth Factor Beta3 Promotes Tendon Differentiation of Equine Embryo-Derived Stem Cells. Tissue Eng. Part A 2013, 19, 2156–2165. [Google Scholar] [CrossRef]
- Kapacee, Z.; Yeung, C.-Y.C.; Lu, Y.; Crabtree, D.; Holmes, D.F.; Kadler, K.E. Synthesis of embryonic tendon-like tissue by human marrow stromal/mesenchymal stem cells requires a three-dimensional environment and transforming growth factor β3. Matrix Biol. 2010, 29, 668–677. [Google Scholar] [CrossRef]
- Jiang, K.; Chun, G.; Du, Q.; Wang, Z.; Wang, A.; Xiong, Y. Effect of transforming growth factor-?3 on the expression of Smad3 and Smad7 in tenocytes. Mol. Med. Rep. 2016, 13, 3567–3573. [Google Scholar] [CrossRef]
- Lee, C.H.; Lee, F.Y.; Tarafder, S.; Kao, K.; Jun, Y.; Yang, G.; Mao, J.J. Harnessing endogenous stem/progenitor cells for tendon regeneration. J. Clin. Investig. 2015, 125, 2690–2701. [Google Scholar] [CrossRef]
- Hoffmann, A.; Pelled, G.; Turgeman, G.; Eberle, P.; Zilberman, Y.; Shinar, H.; Keinan-Adamsky, K.; Winkel, A.; Shahab, S.; Navon, G.; et al. Neotendon formation induced by manipulation of the Smad8 signalling pathway in mesenchymal stem cells. J. Clin. Investig. 2006, 116, 940–952. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Zhou, Z.; Taub, P.J.; Ramcharan, M.; Li, Y.; Akinbiyi, T.; Maharam, E.R.; Leong, D.J.; Laudier, D.M.; Ruike, T.; et al. BMP-12 Treatment of Adult Mesenchymal Stem Cells In Vitro Augments Tendon-Like Tissue Formation and Defect Repair In Vivo. PLoS ONE 2011, 6, e17531. [Google Scholar] [CrossRef]
- Chai, W.; Ni, M.; Rui, Y.-F.; Zhang, K.-Y.; Zhang, Q.; Xu, L.-L.; Chan, K.M.; Li, G.; Wang, Y. Effect of growth and differentiation factor 6 on the tenogenic differentiation of bone marrow-derived mesenchymal stem cells. Chin. Med. J. 2013, 126, 1509–1516. [Google Scholar]
- Violini, S.; Ramelli, P.; Pisani, L.F.; Gorni, C.; Mariani, P. Horse bone marrow mesenchymal stem cells express embryo stem cell markers and show the ability for tenogenic differentiation by in vitro exposure to BMP-12. BMC Cell Biol. 2009, 10, 29. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.; Gao, P.; Zhang, Y.; Yang, S. Combined effects of engineered tendon matrix and GDF-6 on bone marrow mesenchymal stem cell-based tendon regeneration. Biotechnol. Lett. 2016, 38, 885–892. [Google Scholar] [CrossRef] [PubMed]
- Ozasa, Y.; Gingery, A.; Thoreson, A.R.; An, K.-N.; Zhao, C.; Amadio, P.C. A Comparative Study of the Effects of Growth and Differentiation Factor 5 on Muscle-Derived Stem Cells and Bone Marrow Stromal Cells in an In Vitro Tendon Healing Model. J. Hand Surg. 2014, 39, 1706–1713. [Google Scholar] [CrossRef] [Green Version]
- Park, A.; Hogan, M.V.; Kesturu, G.S.; James, R.; Balian, G.; Chhabra, A.B. Adipose-Derived Mesenchymal Stem Cells Treated with Growth Differentiation Factor-5 Express Tendon-Specific Markers. Tissue Eng. Part A 2010, 16, 2941–2951. [Google Scholar] [CrossRef] [Green Version]
- Berasi, S.P.; Varadarajan, U.; Archambault, J.; Cain, M.; Souza, T.A.; Abouzeid, A.; Li, J.; Brown, C.T.; Dorner, A.J.; Seeherman, H.J.; et al. Divergent activities of osteogenic BMP2, and tenogenic BMP12 and BMP13 independent of receptor binding affinities. Growth Factors 2011, 29, 128–139. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.; Gelberman, R.H.; Silva, M.J.; Sakiyama-Elbert, S.E.; Thomopoulos, S. BMP12 induces tenogenic differentiation of adipose-derived stromal cells. PLoS ONE 2013, 8, e77613. [Google Scholar] [CrossRef] [Green Version]
- Tan, S.-L.; Ahmad, T.S.; Ng, W.-M.; Azlina, A.A.; Azhar, M.M.; Selvaratnam, L.; Kamarul, T. Identification Pathways Mediating Growth Differentiation Factor5-Induced Tenogenic Differentiation in Human Bone Marrow Stromal Cells. PLoS ONE 2015, 10, e0140869. [Google Scholar] [CrossRef] [Green Version]
- Moussad, E.E.-D.A.; Brigstock, D.R. Connective Tissue Growth Factor: What’s in a Name? Mol. Genet. Metab. 2000, 71, 276–292. [Google Scholar] [CrossRef]
- Yuda, A.; Maeda, H.; Fujii, S.; Monnouchi, S.; Yamamoto, N.; Wada, N.; Koori, K.; Tomokiyo, A.; Hamano, S.; Hasegawa, D.; et al. Effect of CTGF/CCN2 on Osteo/Cementoblastic and Fibroblastic Differentiation of a Human Periodontal Ligament Stem/Progenitor Cell Line. J. Cell. Physiol. 2014, 230, 150–159. [Google Scholar] [CrossRef]
- Li, X.; Pongkitwitoon, S.; Lu, H.; Lee, C.; Gelberman, R.; Thomopoulos, S. CTGF induces tenogenic differentiation and proliferation of adipose-derived stromal cells. J. Orthop. Res. 2019, 37, 574–582. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Tao, X.; Chen, L.; Han, W.; Zhou, Y.; Tang, K. CTGF Positively Regulates BMP12 Induced Tenogenic Differentiation of Tendon Stem Cells and Signaling. Cell. Physiol. Biochem. 2015, 35, 1831–1845. [Google Scholar] [CrossRef]
- Viganò, M.; Orfei, C.P.; Colombini, A.; Stanco, D.; Randelli, P.; Sansone, V.; De Girolamo, L. Different culture conditions affect the growth of human tendon stem/progenitor cells (TSPCs) within a mixed tendon cells (TCs) population. J. Exp. Orthop. 2017, 4, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hankemeier, S.; Keus, M.; Zeichen, J.; Jagodzinski, M.; Barkhausen, T.; Bosch, U.; Krettek, C.; Van Griensven, M. Modulation of Proliferation and Differentiation of Human Bone Marrow Stromal Cells by Fibroblast Growth Factor 2: Potential Implications for Tissue Engineering of Tendons and Ligaments. Tissue Eng. 2005, 11, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Kraus, T.M.; Imhoff, F.B.; Reinert, J.; Wexel, G.; Wolf, A.; Hirsch, D.; Hofmann, A.; Stöckle, U.; Buchmann, S.; Tischer, T.; et al. Stem cells and bFGF in tendon healing: Effects of lentiviral gene transfer and long-term follow-up in a rat Achilles tendon defect model. BMC Musculoskelet. Disord. 2016, 17, 148. [Google Scholar] [CrossRef] [Green Version]
- Kraus, T.M.; Imhoff, F.B.; Wexel, G.; Wolf, A.; Hirsch, D.; Lenz, L.; Stockle, U.; Buchmann, S.; Tischer, T.; Milz, S.; et al. Stem Cells and Basic Fibroblast Growth Factor Failed to Improve Tendon Healing. J. Bone Jt. Surg. Am. Vol. 2014, 96, 761–769. [Google Scholar] [CrossRef]
- Tokunaga, T.; Shukunami, C.; Okamoto, N.; Taniwaki, T.; Oka, K.; Sakamoto, H.; Ide, J.; Mizuta, H.; Hiraki, Y. FGF-2 Stimulates the Growth of Tenogenic Progenitor Cells to Facilitate the Generation of Tenomodulin -Positive Tenocytes in a Rat Rotator Cuff Healing Model. Am. J. Sports Med. 2015, 43, 2411–2422. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.P.; Finley, V.G.; Kuo, C.K. Embryonic mechanical and soluble cues regulate tendon progenitor cell gene expression as a function of developmental stage and anatomical origin. J. Biomech. 2013, 47, 214–222. [Google Scholar] [CrossRef] [Green Version]
- Murphy, D.J.; Nixon, A.J. Biochemical and site-specific effects of insulin-like growth factor I on intrinsic tenocyte activity in equine flexor tendons. Am. J. Vet. Res. 1997, 58, 103–109. [Google Scholar] [PubMed]
- Schnabel, L.V.; Lynch, M.E.; Van Der Meulen, M.C.; Yeager, A.E.; Kornatowski, M.A.; Nixon, A.J.; Van Der Meulen, M.C.H. Mesenchymal stem cells and insulin-like growth factor-I gene-enhanced mesenchymal stem cells improve structural aspects of healing in equine flexor digitorum superficialis tendons. J. Orthop. Res. 2009, 27, 1392–1398. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chen, L.; Zhou, Y.; Liu, X.; Tang, K. Insulin-Like Growth Factor-1 and Bone Morphogenetic Protein-2 Jointly Mediate Prostaglandin E2-Induced Adipogenic Differentiation of Rat Tendon Stem Cells. PLoS ONE 2014, 9, e85469. [Google Scholar] [CrossRef] [PubMed]
- Petersen, W.; Pufe, T.; Unterhauser, F.; Zantop, T.; Mentlein, R.; Weiler, A. The splice variants 120 and 164 of the angiogenic peptide vascular endothelial cell growth factor (VEGF) are expressed during Achilles tendon healing. Arch. Orthop. Trauma Surg. 2003, 123, 475–480. [Google Scholar] [CrossRef]
- Öhberg, L.; Lorentzon, R.; Alfredson, H. Neovascularisation in Achilles tendons with painful tendinosis but not in normal tendons: An ultrasonographic investigation. Knee Surg. Sports Traumatol. Arthrosc. 2001, 9, 233–238. [Google Scholar] [CrossRef]
- Boyer, M.I.; Watson, J.T.; Lou, J.; Manske, P.R.; Gelberman, R.H.; Cai, S.R. Quantitative variation in vascular endothelial growth factor mRNA expression during early flexor tendon healing: An investigation in a canine model. J. Orthop. Res. 2001, 19, 869–872. [Google Scholar] [CrossRef]
- Ferrara, N.; Adamis, A.P. Ten years of anti-vascular endothelial growth factor therapy. Nat. Rev. Drug Discov. 2016, 15, 385–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Liu, H.; Stile, F.; Lei, M.-P.; Pang, Y.; Oswald, T.M.; Beck, J.; Dorsett-Martin, W.; Lineaweaver, W.C. Effect of Vascular Endothelial Growth Factor on Rat Achilles Tendon Healing. Plast. Reconstr. Surg. 2003, 112, 1613–1619. [Google Scholar] [CrossRef]
- Petersen, W.; Unterhauser, F.; Pufe, T.; Zantop, T.; Südkamp, N.P.; Weiler, A. The angiogenic peptide vascular endothelial growth factor (VEGF) is expressed during the remodeling of free tendon grafts in sheep. Arch. Orthop. Trauma Surg. 2003, 123, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Kraus, A.; Sattler, D.; Wehland, M.; Luetzenberg, R.; Abuagela, N.; Infanger, M. Vascular Endothelial Growth Factor Enhances Proliferation of Human Tenocytes and Promotes Tenogenic Gene Expression. Plast. Reconstr. Surg. 2018, 142, 1240–1247. [Google Scholar] [CrossRef]
- Heldin, C. Structural and functional studies on platelet-derived growth factor. EMBO J. 1992, 11, 4251–4259. [Google Scholar] [CrossRef]
- Yoshikawa, Y.; Abrahamsson, S.-O. Dose-related cellular effects of platelet-derived growth factor-BB differ in various types of rabbit tendons in vitro. Acta Orthop. Scand. 2001, 72, 287–292. [Google Scholar] [CrossRef]
- Perikamana, S.K.M.; Lee, J.; Ahmad, T.; Kim, E.-M.; Byun, H.; Lee, S.; Shin, H. Harnessing biochemical and structural cues for tenogenic differentiation of adipose derived stem cells (ADSCs) and development of an in vitro tissue interface mimicking tendon-bone insertion graft. Biomaterials 2018, 165, 79–93. [Google Scholar] [CrossRef]
- Evrova, O.; Kellenberger, D.; Calcagni, M.; Vogel, V.; Buschmann, J. Supporting Cell-Based Tendon Therapy: Effect of PDGF-BB and Ascorbic Acid on Rabbit Achilles Tenocytes In Vitro. Int. J. Mol. Sci. 2020, 21, 458. [Google Scholar] [CrossRef] [Green Version]
- Roth, S.P.; Schubert, S.; Scheibe, P.; Groß, C.; Brehm, W.; Burk, J. Growth Factor-Mediated Tenogenic Induction of Multipotent Mesenchymal Stromal Cells Is Altered by the Microenvironment of Tendon Matrix. Cell Transplant. 2018, 27, 1434–1450. [Google Scholar] [CrossRef] [Green Version]
- Rajpar, I.; Barrett, J.G. Optimizing growth factor induction of tenogenesis in three-dimensional culture of mesenchymal stem cells. J. Tissue Eng. 2019, 10, 204173141984877. [Google Scholar] [CrossRef]
- Paschos, N.K.; Brown, W.E.; Eswaramoorthy, R.; Hu, J.C.; Athanasiou, K.A. Advances in tissue engineering through stem cell-based co-culture. J. Tissue Eng. Regen. Med. 2014, 9, 488–503. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Liu, Y.; Wang, B.; Sun, Y.; Xu, J.; Yuk-Wai, L.W.; Xu, L.; Zhang, J.-F.; Li, G. The Use of Cocultured Mesenchymal Stem Cells with Tendon-Derived Stem Cells as a Better Cell Source for Tendon Repair. Tissue Eng. Part A 2016, 22, 1229–1240. [Google Scholar] [CrossRef]
- Luo, Q.; Song, G.; Song, Y.; Xu, B.; Qin, J.; Shi, Y. Indirect co-culture with tenocytes promotes proliferation and mRNA expression of tendon/ligament related genes in rat bone marrow mesenchymal stem cells. Cytotechnology 2009, 61, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Canseco, J.A.; Kojima, K.; Penvose, A.R.; Ross, J.D.; Obokata, H.; Gomoll, A.H.; Vacanti, C.A. Effect on Ligament Marker Expression by Direct-Contact Co-culture of Mesenchymal Stem Cells and Anterior Cruciate Ligament Cells. Tissue Eng. Part A 2012, 18, 2549–2558. [Google Scholar] [CrossRef] [Green Version]
- Kraus, A.; Woon, C.Y.; Raghavan, S.; Megerle, K.; Pham, H.; Chang, J. Co-Culture of Human Adipose-Derived Stem Cells with Tenocytes Increases Proliferation and Induces Differentiation into a Tenogenic Lineage. Plast. Reconstr. Surg. 2013, 132, 754e–766e. [Google Scholar] [CrossRef]
- Yu, Y.; Zhou, Y.; Cheng, T.; Lu, X.; Yu, K.; Zhou, Y.; Hong, J.; Chen, Y. Hypoxia enhances tenocyte differentiation of adipose-derived mesenchymal stem cells by inducing hypoxia-inducible factor-1α in a co-culture system. Cell Prolif. 2016, 49, 173–184. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhou, Y.; Zhang, X.; Chen, Y.; Zheng, X.; Cheng, T.; Wang, C.; Hu, X.; Hong, J. Effects of hypoxia on differentiation of menstrual blood stromal stem cells towards tenogenic cells in a co-culture system with Achilles tendon cells. Exp. Ther. Med. 2017, 13, 3195–3202. [Google Scholar] [CrossRef]
- Wunderli, S.L.; Blache, U.; Snedecker, J. Tendon explant models for physiologically relevant in vitro study of tissue biology—A perspective. Connect. Tissue Res. 2020, 61, 262–277. [Google Scholar] [CrossRef] [Green Version]
- Lovati, A.B.; Corradetti, B.; Cremonesi, F.; Bizzaro, D.; Consiglio, A.L. Tenogenic differentiation of equine mesenchymal progenitor cells under indirect co-culture. Int. J. Artif. Organs 2012, 35, 996–1005. [Google Scholar] [CrossRef]
- Costa-Almeida, R.; Berdecka, D.; Rodrigues, M.T.; Reis, R.L.; Gomes, M.E. Tendon explant cultures to study the communication between adipose stem cells and native tendon niche. J. Cell. Biochem. 2018, 119, 3653–3662. [Google Scholar] [CrossRef]
- Wu, S.; Peng, H.; Li, X.; Streubel, P.N.; Liu, Y.; Duan, B. Effect of scaffold morphology and cell co-culture on tenogenic differentiation of HADMSC on centrifugal melt electrospun poly (L-lactic acid) fibrous meshes. Biofabrication 2017, 9, 044106. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, W.; Liu, Z.Y.; Heng, B.C.; Ouyang, H.W.; Dai, X.S. Physical regulation of stem cells differentiation into teno-lineage: Current strategies and future direction. Cell Tissue Res. 2014, 360, 195–207. [Google Scholar] [CrossRef]
- Testa, S.; Costantini, M.; Fornetti, E.; Bernardini, S.; Trombetta, M.; Seliktar, D.; Cannata, S.; Rainer, A.; Gargioli, C. Combination of biochemical and mechanical cues for tendon tissue engineering. J. Cell. Mol. Med. 2017, 21, 2711–2719. [Google Scholar] [CrossRef]
- Rinoldi, C.; Fallahi, A.; Yazdi, I.K.; Paras, J.C.; Kijeńska-Gawrońska, E.; Santiago, G.T.-D.; Tuoheti, A.; Demarchi, D.; Annabi, N.; Khademhosseini, A.; et al. Mechanical and Biochemical Stimulation of 3D Multilayered Scaffolds for Tendon Tissue Engineering. ACS Biomater. Sci. Eng. 2019, 5, 2953–2964. [Google Scholar] [CrossRef]












{kind=link}
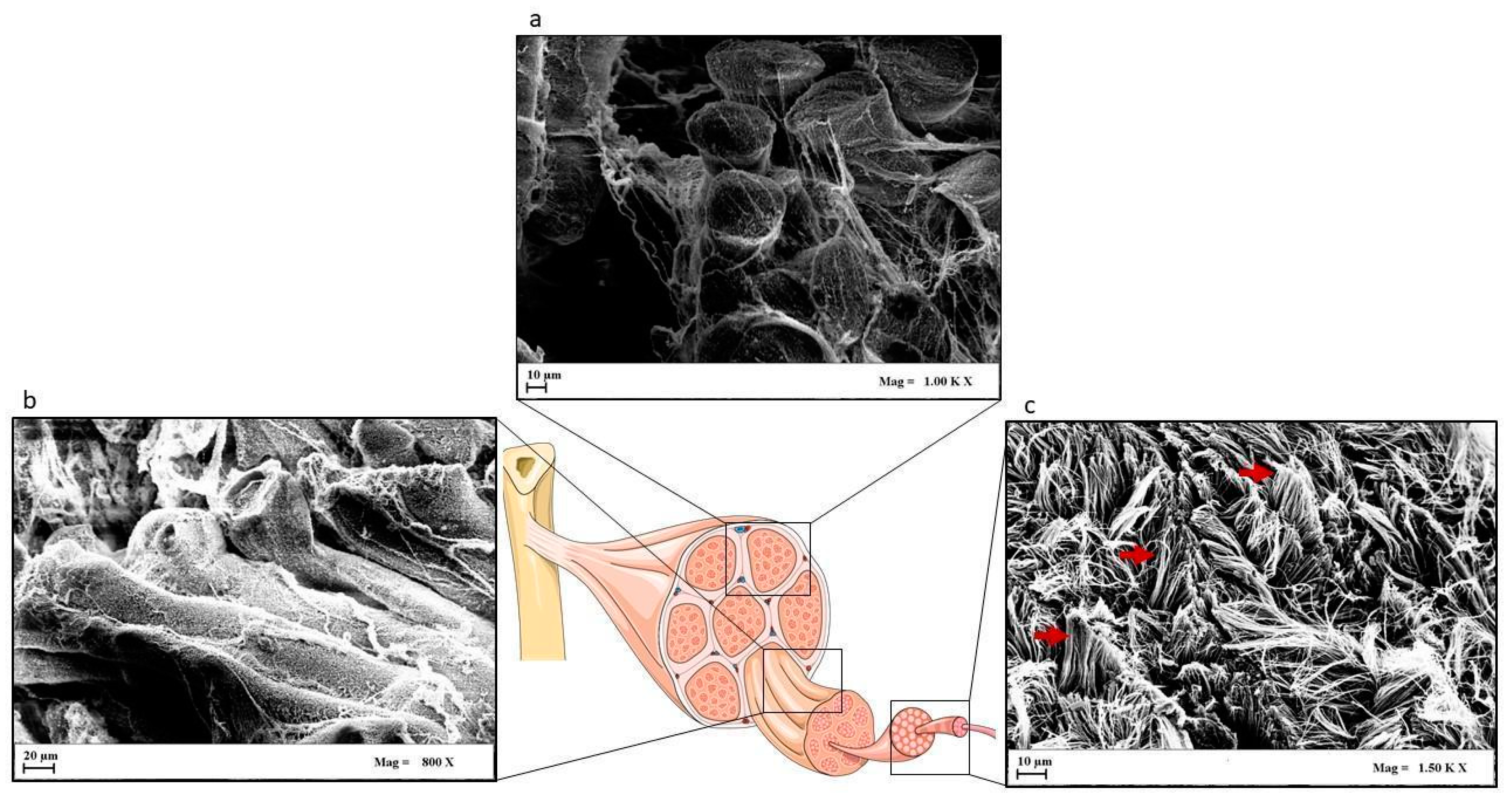{kind=link}
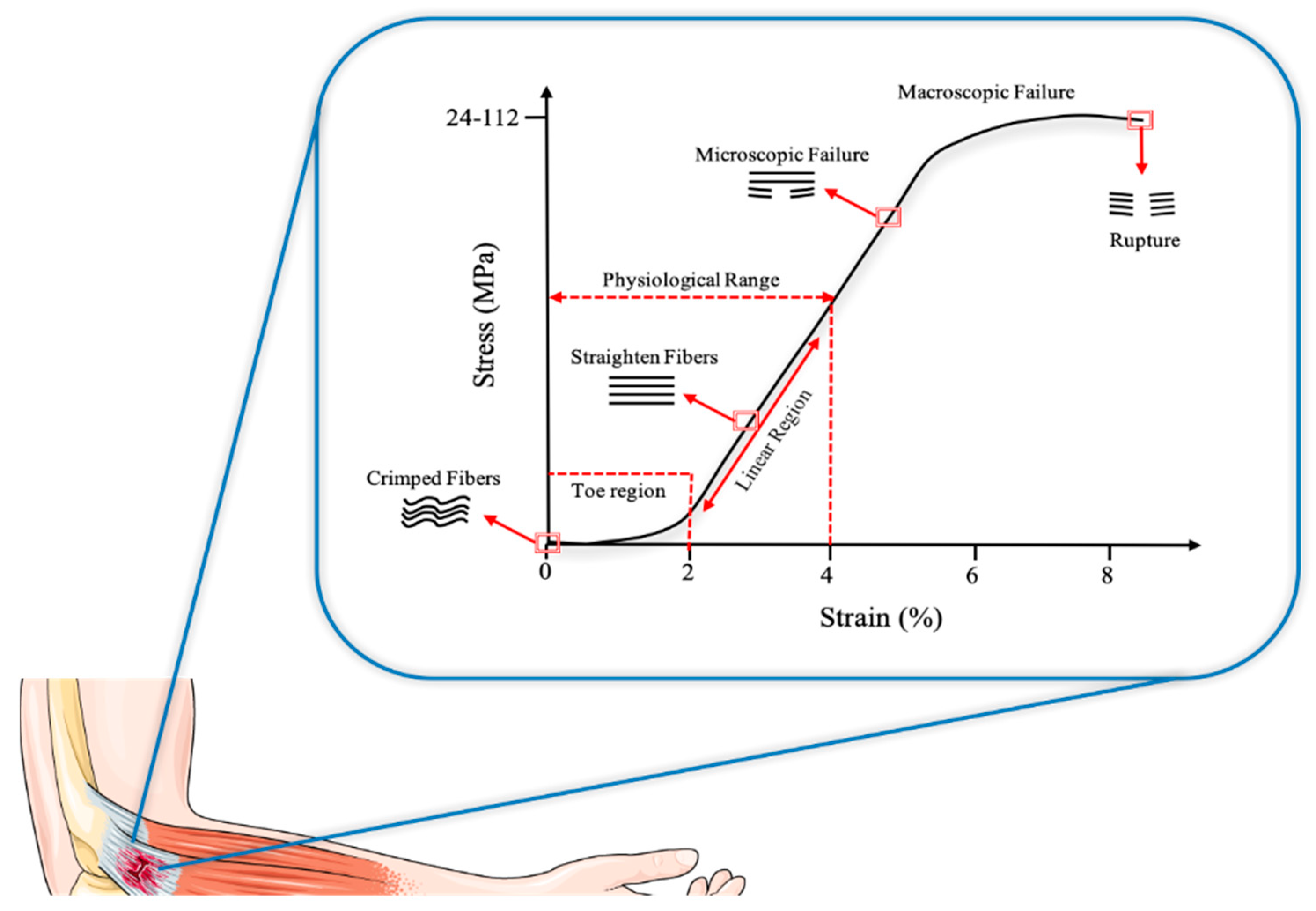{kind=link}
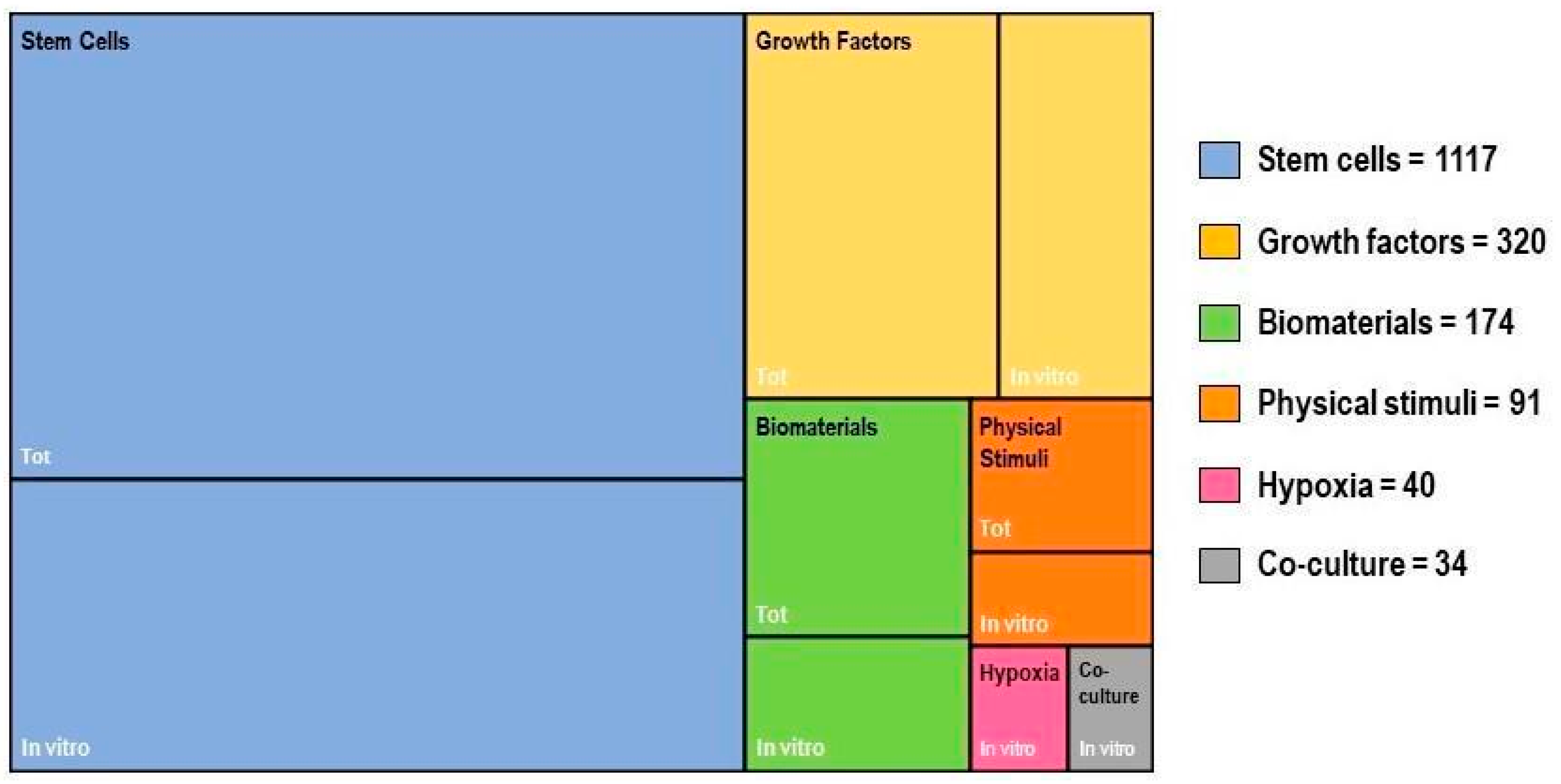{kind=link}
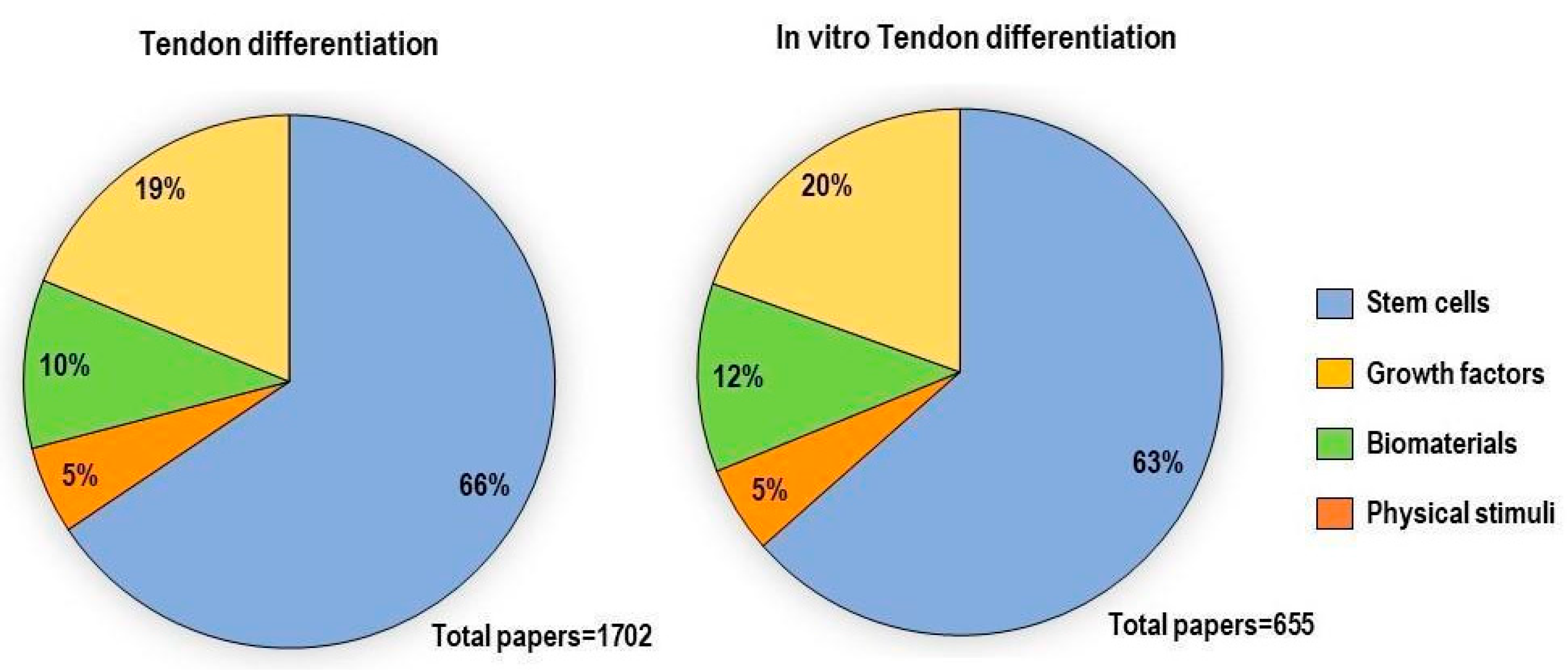{kind=link}
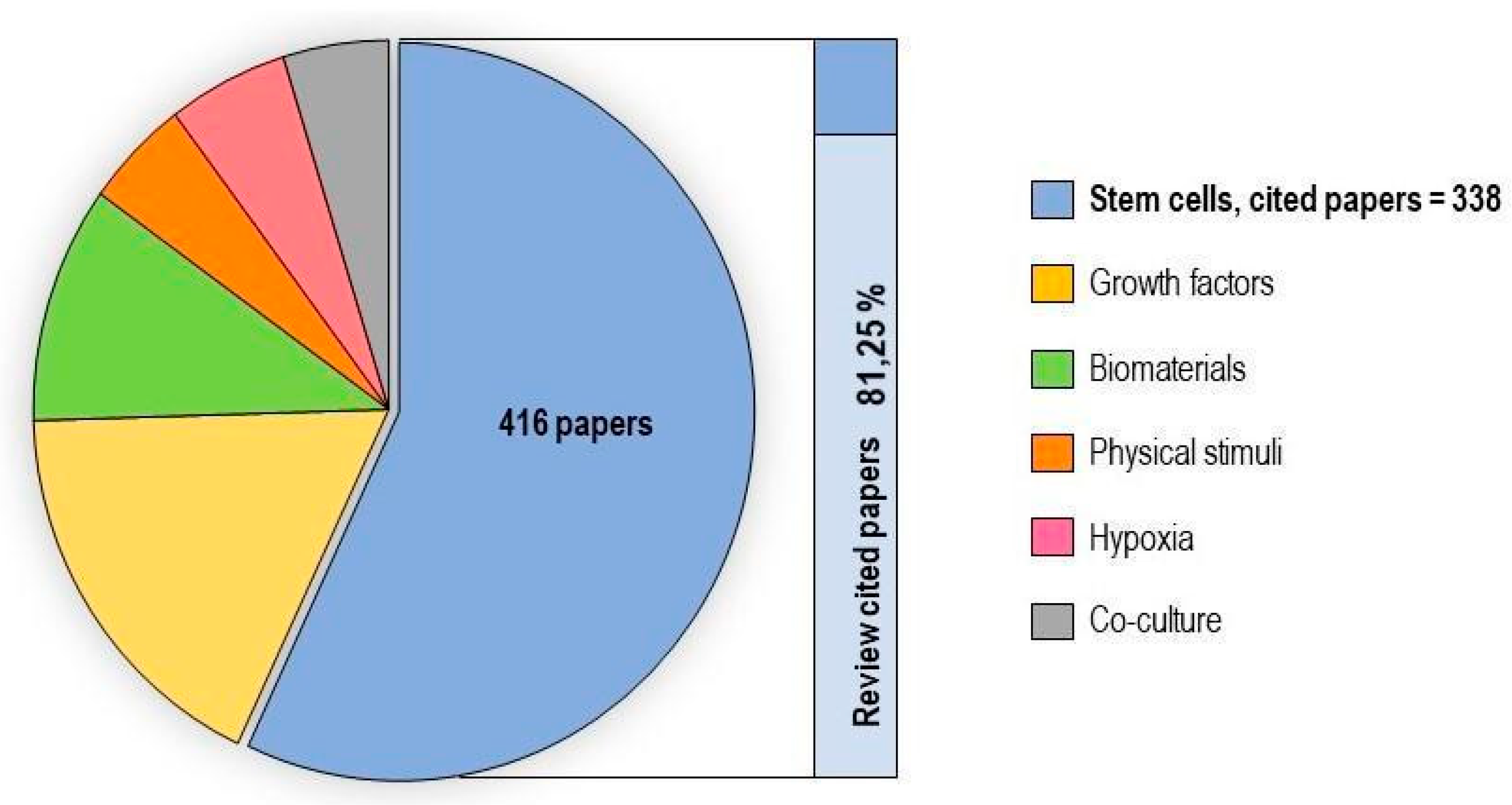{kind=link}
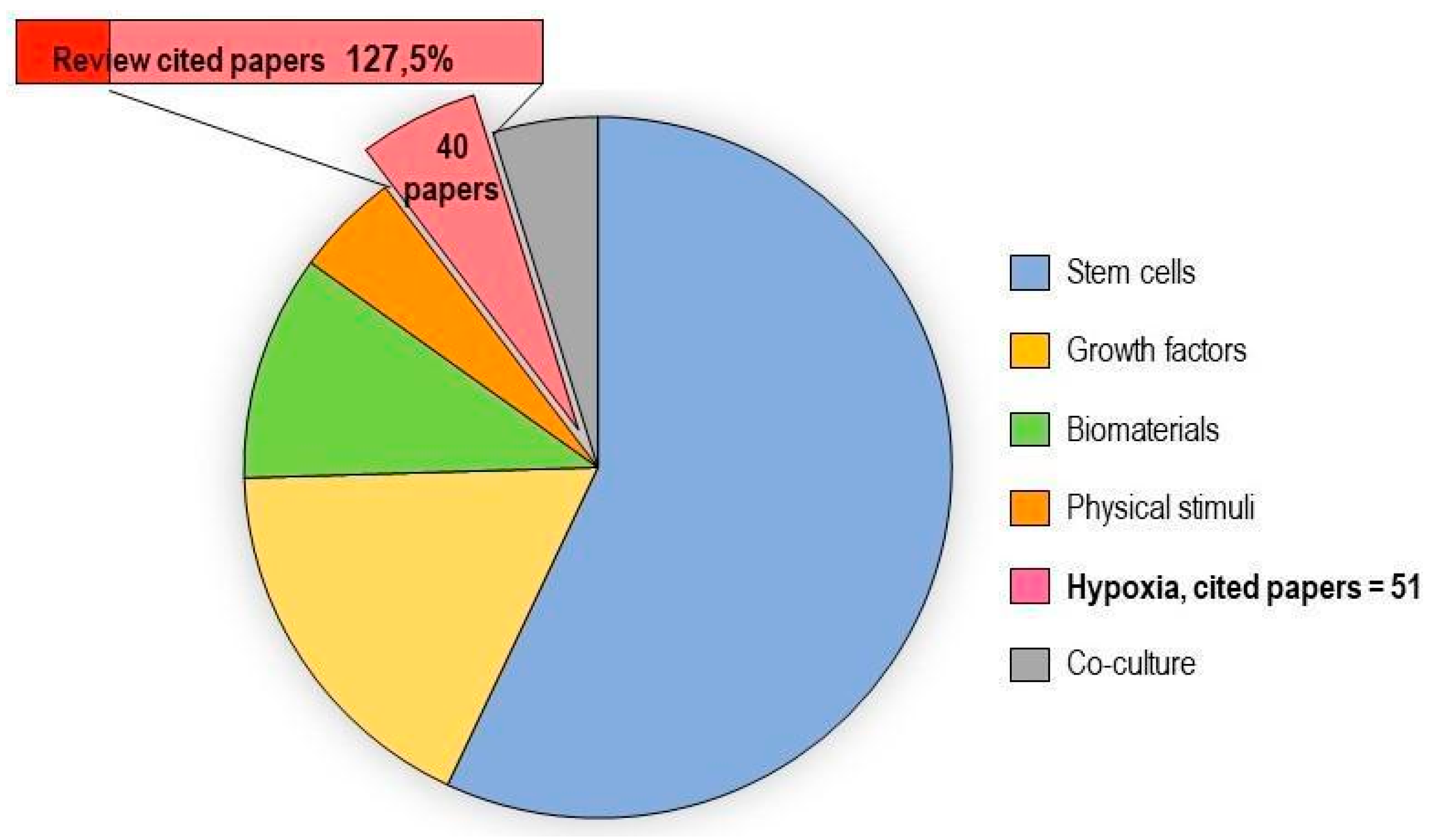{kind=link}
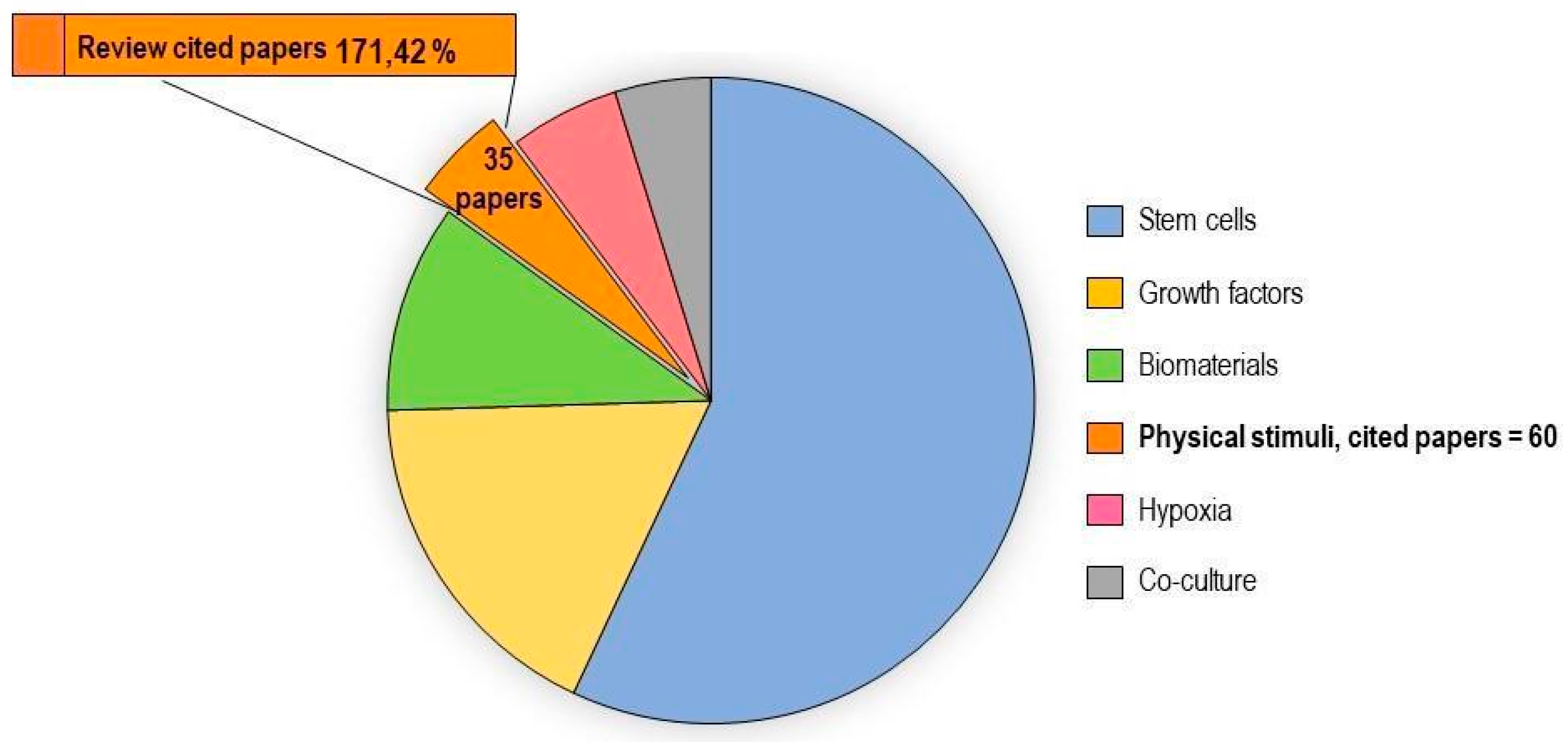{kind=link}
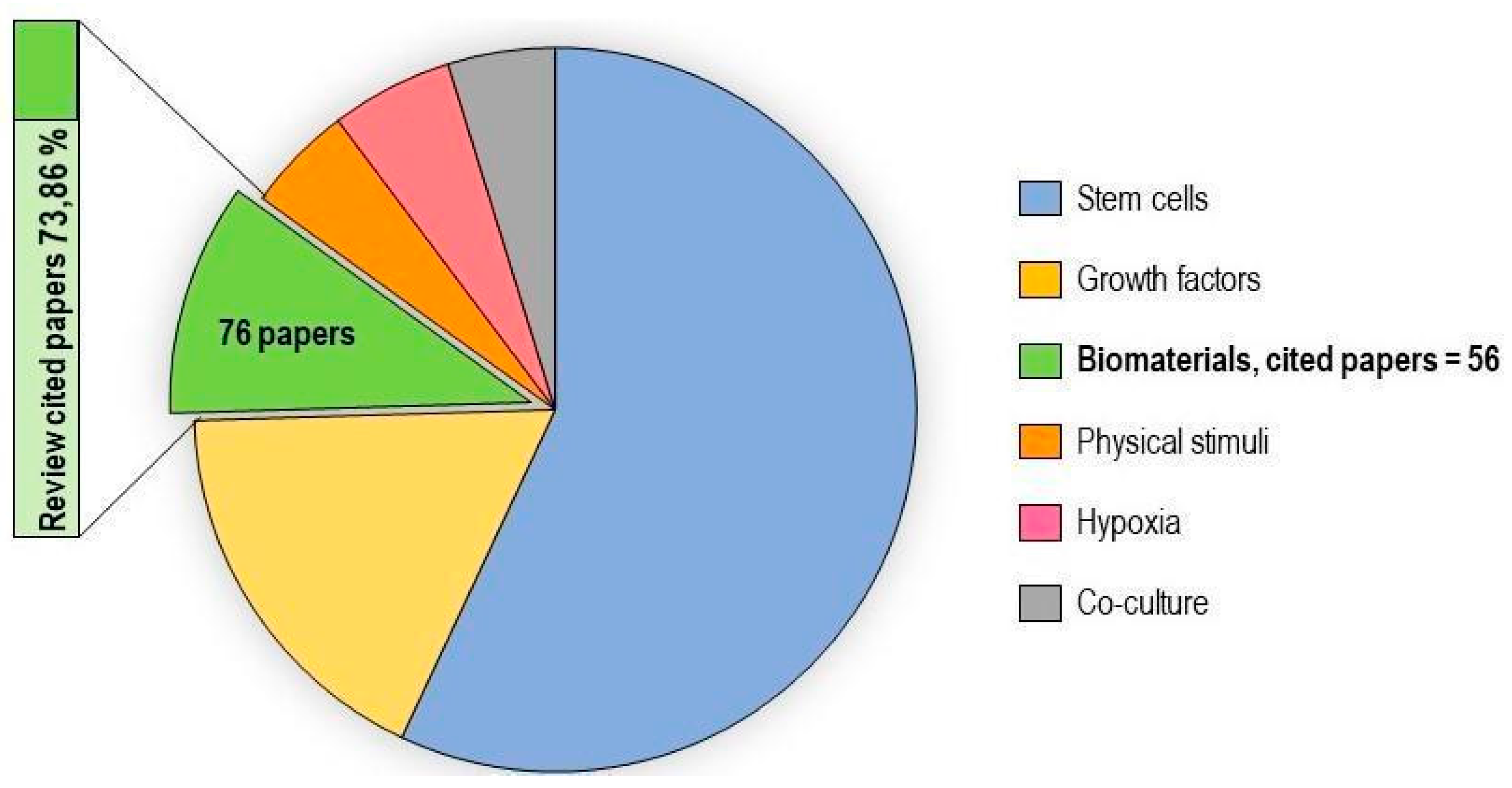{kind=link}
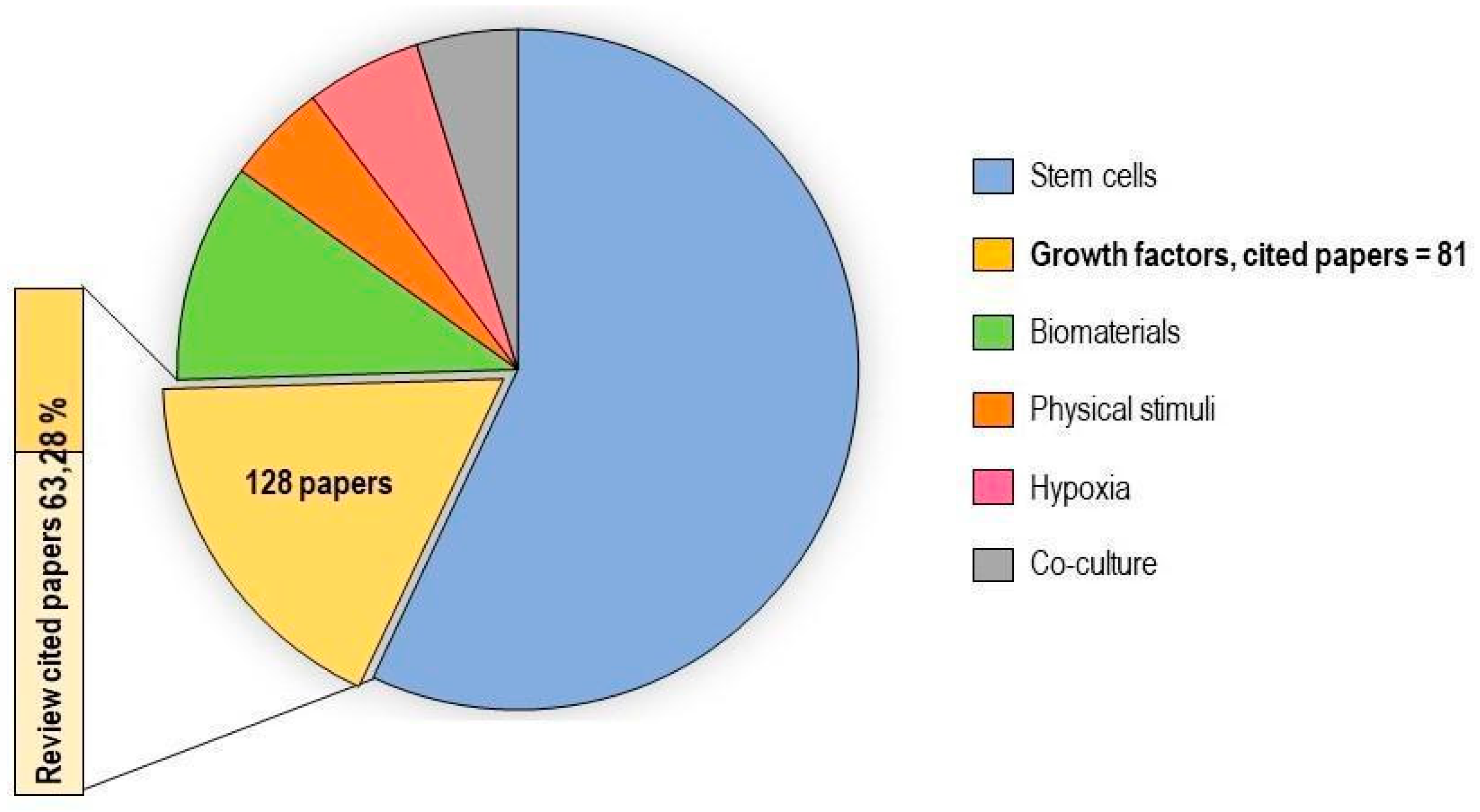{kind=link}
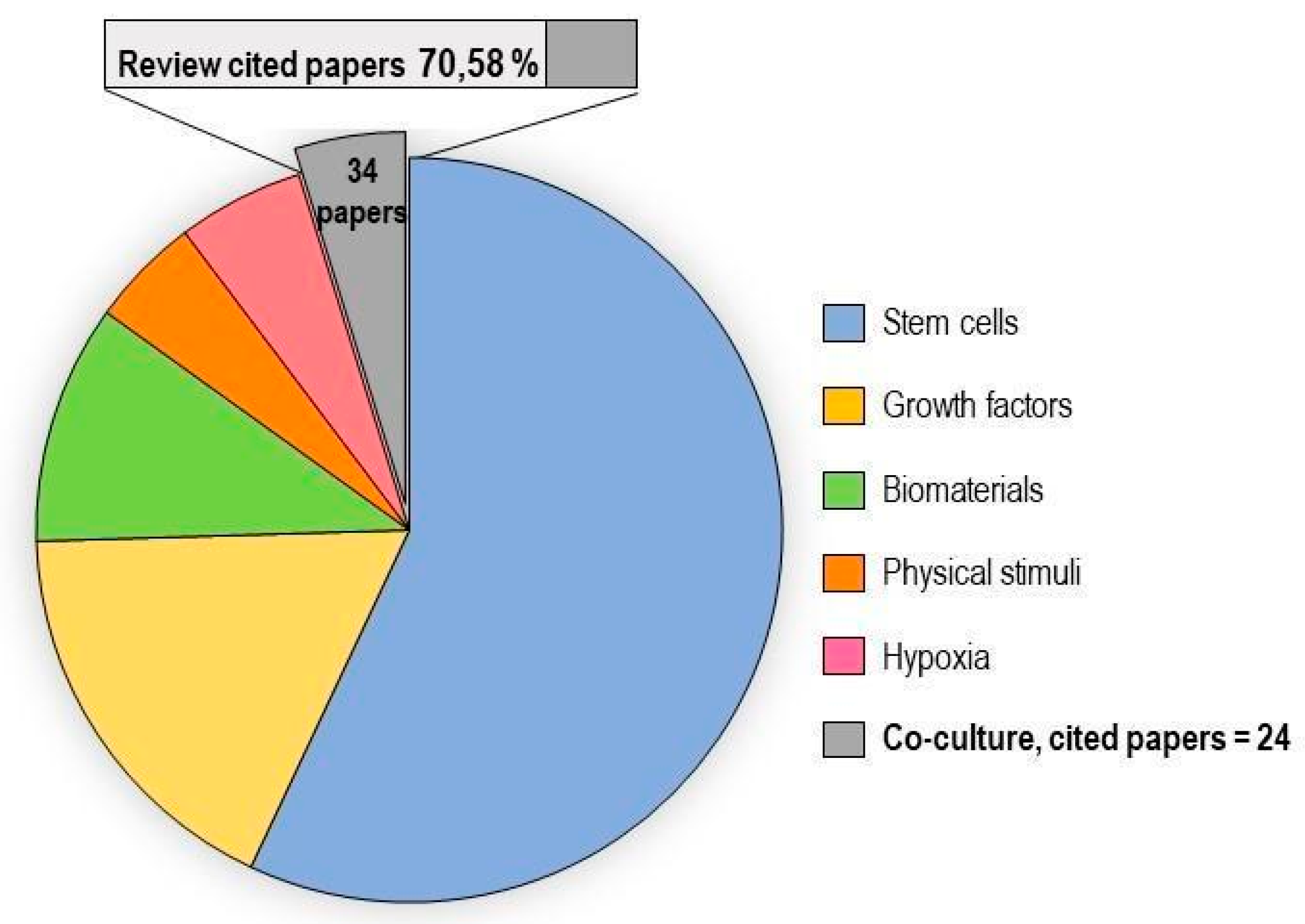{kind=link}
| Materials | Shape and Structure of the Scaffold | Mechanical Properties of the Scaffold | Ref. |
|---|---|---|---|
| Silk fibroin–collagen | As-spun scaffold → diameter = 1.15 μm; pore size = 43.79 μm2 Cross-linked scaffold with ethanol → diameter = 0.76 μm; pore size = 21.39 μm2 Cross-linked scaffold with methanol → diameter = 0.91 μm; pore size = 10.81 μm2 | Cross-linked scaffold with ethanol → Stress = 1.2 MPa; Modulus of elasticity = 4.2 MPa Cross-linked scaffold with methanol → Stress = 2.92 MPa; Modulus of elasticity = 9.78 MPa | [324] |
| PLGA–Silk fibroin | Knitted silk scaffold (3 yarns, diameter = 10 μm) coated with electrospun PLGA nanofibers diameter = 200–700 nm Thickness of the knitted scaffolds = 0.6–1 mm Thickness of the coated hybrid scaffolds = 0.8–1.4 mm | Hybrid scaffold → failure load = 70.8 N; stiffness = 4.29 N/mm | [325] |
| CS–PCL | Random nanofibers = 215.79 nm Aligned nanofibers = 175.82 nm | n.d. | [328] |
| PEUUR | Four randomly oriented meshes with diameter: 0.28, 0.72, 0.82, 2.3 μm Two meshes with aligned fibers with diameter: 0.46 and 0.53 μm | n.d. | [335] |
| PEUUR | Aligned and randomly oriented fibers with different diameter size <1 μm, >1–2 μm<, and >2 μm | n.d. | [336] |
| PU | Aligned and random nanofibers with diameter of ≈657 nm; porosity = 85%; total pore area = 61.8 m2/g | Aligned nanofibers → Young’s modulus = 2500 kPa; Ultimate strength = 3520 kPa; Max strain = 1.08 Random nanofibers → Young’s modulus = 540 kPa; Ultimate strength = 1130 kPa; Max strain = 1.03 | [337] |
| PLLA/PEO loaded with TSA | PLLA/PEO random mats → diameter = 1.51 μm PLLA/PEO/TSA random mats → diameter = 1.44 μm PLLA/PEO aligned mats → diameter = 1.73 μm PLLA/PEO/TSA aligned mats → diameter = 1.63 μm | PLLA/PEO/TSA random mats → Young’s modulus = 513.09 MPa; Tensile strength = 12.5 MPa PLLA/PEO/TSA aligned mats → Young’s Modulus = 51.59 MPa; Tensile strength = 2.70 MPa | [338] |
| PLGA (85:15)/COL/PU | Randomly oriented scaffolds with average fiber diameter size of 1.92 μm and pore size of 174.7 μm2 Aligned fiber scaffold with average fiber diameter size of 0.712 μm and pore size of 6.75 μm2 | Random fiber 1.92 μm → Young’s Modulus = 20.76 MPa; Ultimate Tensile Strain = 56.16%; Tensile Stress = 1.17 MPa Aligned fiber 0.712 μm → Young’s Modulus = 38.11 MPa; Ultimate Tensile Strain = 82.99%; Tensile Stress = 1.80 MPa | [350] |
| PLGA (50:50)/COL1/PU | Randomly oriented scaffolds with average fiber diameter size of 2.82 μm and pore size of 211.3 μm2 Aligned fiber scaffold with average fiber diameter size of 0.759 μm and 4.67 μm2 | Random fiber 2.82 μm → Young’s Modulus = 75.4 MPa; Ultimate Tensile Strain = 28.55%; Tensile Stress = 0.86 MPa Aligned fiber 0.759 μm → Young’s Modulus = 52.46 MPa; Ultimate Tensile Strain = 79.48%; Tensile Stress = 2.46 MPa | |
| PLGA (85:15) | Aligned fleeces → diameter = 2.5 μm Random fleeces → diameter = 2.1 μm | Aligned fibers → stress = 26.02 MPa and strain = 344% Random fibers → stress = 15 MPa and strain = 240% | [190] |
| PCL and PLLA | Sheets of aligned PCL → diameter = 898 nm; pore size = 14.3 μm Sheet of aligned PLLA → diameter = 869 μm; pore size = 21 μm | Braided PCL scaffolds → Young’s Modulus = 45.96 MPa; Ultimate stress = 19.99 MPa; Ultimate Strain = 0.62 mm/mm; Stiffness = 11.25 N/mm; Stacked PCL scaffolds → Young’s Modulus = 66.48 MPa; Ultimate stress = 8.73 MPa; Ultimate Strain = 0.24 mm/mm; Stiffness = 36.51 N/mm; Braided PLLA scaffolds → Young’s Modulus = 45.57 MPa; Ultimate stress = 5.74 MPa; Ultimate Strain = 0.50 mm/mm; Stiffness = 5.94 N/mm; Stacked PLLA scaffolds → Young’s Modulus = 118.47 MPa; Ultimate stress = 5.76 MPa; Ultimate Strain = 0.22 mm/mm; Stiffness = 24.31 N/mm; | [367] |
| PCL | Random multilayer scaffolds → diameter = 1.76 μm; thickness = 0.75 mm Aligned multilayer scaffolds → diameter = 1.57 μm; thickness = 0.43 mm | Random multilayer → Elastic Modulus = 12 MPa; Yield Strength = 0.5 MPa Aligned multilayer → Elastic Modulus = 18 MPa; Yield Strength = 1.5 MPa | [368] |
| PLLA | Mat thickness ranged between 0.14 and 0.17 mm Aligned fibers = 430 nm Random fibers = 450 nm | Aligned fibers → Stiffness = 3.48 N/mm; Failure force = 1.88 N; Young’s Modulus = 22.76 MPa Random fibers → Stiffness = 0.07 N/mm; Failure force = 0.17 N; Young’s Modulus = 0.63 MPa | [369] |
| PLGA (85:15) | Aligned scaffold →thickness = 0.22 mm; diameter = 615 nm; pore diameter = 4.228 μm; porosity = 80.745%; permeability = 7.87 × 10−12 m4/N s Random scaffold → thickness = 0.19 mm; diameter = 568 nm; pore diameter = 4.914 μm; porosity = 81.760%; permeability = 5.72 × 10−12 m4/N s | Aligned scaffolds → Elastic Modulus = 341 MPa; Yield Strength = 9.8 MPa; Ultimate Stress = 12 MPa and Strain = 8% Random scaffolds → Elastic Modulus = 107 MPa; Yield Strength = 2.5 MPa; Ultimate Stress = 3.7 MPa and Strain = 80%. | [370] |
| PLLA/COL1 (75:25) | Individual aligned fiber with diameter = 0.36 μm and bundle diameter = 624.9 μm | Bundle PLLA/Col1 (75:25) → failure stress = 11.3 MPa; work to failure = 0.225 J/mm3; | [371] |
| PLLA/COL1 (50:50) | Individual aligned fiber with diameter = 0.39 μm and bundle diameter = 643.1 μm | Bundle PLLA/Col1 (50::50) → failure stress = 6 MPa; work to failure = 0.208 J/mm3; | |
| PCL | PCL yarns with diameter = 208.5 μm Yarn diameter size = 460.2 nm, pore size = 12.2 μm Random PCL diameter = 484.5 nm, pore size = 2.1 μm Aligned PCL diameter = 452.3 nm, pore size = 1.4 μm | Yarn PCL woven fabrics → Young’s modulus = 70 MPa; Ultimate tensile strength = 10.7 MPa; Elongation at failure = 48% Random mesh → Young’s modulus = 5.2 MPa; Ultimate tensile strength = 2.2 MPa; Elongation at failure = 240% Aligned mesh → Young’s modulus = 13.6 MPa; Ultimate tensile strength = 5 MPa; Elongation at failure = 45% | [373] |
| PCL/DT-NPs | PCL/DT-NP scaffold PCL and PCL/DT-NP twisted yarn using 12 threads Diameter range 313–346 μm | PCL → Young’s modulus = 12 MPa; Strain at break = 3.4 mm·mm−1; Stress = 2.9 MPa PCL/DT-NP2.5 → Young’s modulus = 18 MPa; Strain at break = 3.9 mm·mm−1; Stress = 4.2 MPa PCL/DT-NP5 → Young’s modulus = 22 MPa; Strain at break = 4.2 mm·mm−1; Stress = 4.8 MPa | [304] |
| PLLA/PCL | Braided aligned PLLA and PCL nanofibers with diameter ≈990 and 945 nm, respectively. Varying number of stitches (SPI) (8, 12, 16, 20 and 24) →↑ braid angle from 47° to 67° | PCL/PLLA (100/0) → Ultimate strength = 50.57 MPa; ultimate strain = 1.01 mm/mm; Young’s modulus = 121.21 MPa PCL/PLLA (75/25) → Ultimate strength = 20.30 MPa; ultimate strain = 2.30 mm/mm; Young’s modulus = 78.71 MPa PCL/PLLA (50/50) → Ultimate strength = 4.64 MPa; ultimate strain = 0.75 mm/mm; Young’s modulus = 14.79 MPa PCL/PLLA (25/75) → Ultimate strength = 2.78 MPa; ultimate strain = 1.85 mm/mm; Young’s modulus = 5.25 MPa PCL/PLLA (0/100) → Ultimate strength = 13.47 MPa; ultimate strain = 0.75 mm/mm; Young’s modulus = 48.50 MPa Different braiding angles →↑ strength, Young’s Modulus and yield strength and ↓ yield strain with the braiding angles of 67° (8 SPI) compared to 47° (24 SPI) | [374] |
| PLGA (85:15) | Aligned fibers with diameters 320 nm, 680 nm, and 1.80 μm | The UTS and yield strength of the scaffolds remained unchanged whether is the fiber diameter size. The tensile modulus increased by increasing fiber diameter size while elongation at break and ductility decreased when the fiber diameter size in increased. | [375] |
| PCL | Four PCL mats with randomly oriented fiber and different diameter size; 0.11, 0.78, 1.88, and 3.43 μm | fiber diameters (range of 0.1–3.4 μm) → Young’s modulus = 7.6–30.6 MPa; ultimate tensile strength = 0.9–6.3 MPa; strain at break = 49–442% | [376] |
| PLGA (85:15) | Aligned nanofibers → diameter = 615 nm; pore size = 4.23 μm; porosity = 80.75% Random nanofibers → diameter = 667 nm; pore size = 4.91 μm; porosity = 81.76% | n.d. | [377] |
| PLGA (85:15) | Meshes of randomly oriented fiber with diameter nano-1 (390 nm), nano-2 (740 nm), micro (1.42 μm) Meshes of aligned fibers with diameter nano-2 (740 nm) | n.d. | [378] |
| Cells | Mechanical Properties of the Bio-Hybrid In Vitro | Major In-Vitro Outcome | Ref. |
|---|---|---|---|
| BMSCs | After 21 days of culture Acellular scaffold → failure load = 61.5 N; stiffness = 5.92 N/mm Bio-hybrid scaffolds FGF (−) → failure load = 68.2 N; stiffness = 5.53 N/mm Bio-hybrid scaffolds FGF (+) → failure load = 82.7 N; stiffness = 6.97 N/mm | Bio-hybrid scaffolds (FGF +/−) → viable cells on the surface and in the depths of the scaffolds with higher viability on scaffolds FGF + ↑ mRNA expression Col I, Col III, fibronectin and biglycan and collagen ECM content at day 14 and 21, respectively, in scaffold FGF (+) respect to FGF (−). | [325] |
| Human primary BMSCs | n.d. | Aligned/random scaffolds: No difference in cell adhesion, and fibroblast morphology was observed onto both scaffold Cells aligned parallel to the direction of the nanofiber orientation. No difference has been detected for Tnmd gene expression on the aligned/random scaffold while Col I and Col III was upregulated on aligned scaffold. | [328] |
| Rat BMSCs | n.d. | Cells acquired spindle-like morphology on aligned fibers respect to random ones. ↓ cellular density by increasing fiber diameter size. ↑ cellular aspect ratio by increasing fiber diameter size and alignment ↓ mRNA ColI a1, Tnmd and decorin by increasing fiber diameter and alignment while ↑ mRNA Scx by increasing fiber diameter and decreasing fiber alignment | [335] |
| Multipotent fibroblastic C3H10T1/2 cells | n.d. | ↓ cell density and ↓ mRNA decorin and Col I gene expression by increasing fiber diameter size after 7 days culture while ↑ mRNA Col I gene expression by decreasing fiber diameter after 14 days culture | [336] |
| Human LF | n.d. | Aligned/random nanofibers → no difference in cell proliferation and adhesion while ↑ collagen content in aligned nanofibers respect to random ones. By applying 5% uniaxial strain for 24 h at a frequency of 12 cycles/min → no histological difference between aligned nanofibers under static and dynamic conditions, ↑ collagen ECM in the aligned nanofibers under dynamic conditions. Random nanofibers → cells acquired a spindle-like morphology under only dynamic conditions. | [337] |
| Mouse tail TSPCs | n.d. | Aligned/random (+/− TSA) →↑ cell elongation and ↓ nuclear shape Aligned/random (+/− TSA) → no differences in cell proliferation and adhesion Aligned-TSA →↑ tenogenesis protein (Col I, Col V, Tnmd and Epha4), ↑ ScxGFP protein expression and mRNA expression of Scx, Mkx, Eya1, Eya2, Six2, HoxA11 and Egr1, and ↑ HADC 3 and 4 and ↓ HDAC 1 compared to other groups, | [338] |
| Human iPSCs from HFF (human foreskin fibroblast) | n.d. | Aligned/random nanofibers: no differences in cell proliferation rate and adhesion. Cells seeded onto aligned nanofibers present a fibroblastic phenotype while those onto random nanofibers show a stellate-patterned morphology. Aligned nanofibers: ↑ mRNA of tendon-related genes (Scx, Mkx, Tnmd, HoxA11, Epha4, Col Ia1) and mRNA of integrin a1, a2, a5, b1 and myosin II B. | [340] |
| AECs | n.d. | Aligned/random →↓ DNA quantity and cell proliferation Aligned/random →↑ mRNA Snail and Vimentin, and ↑ α-SMA and ↓ Cytokeratin-8 protein expression Aligned/random →↑ mRNA Tnmd and Col I after 48 h culture. | [190] |
| Human BMSCs | n.d. | Cells were homogenously distributed and showed an elongated morphology on the stacked scaffold compared to the braided ones. Braided scaffolds: ↓ cell infiltration and ↓ cell distribution homogeneity. Stacked scaffolds: ↑ cell proliferation and Col I ECM deposition with an enhanced deposition in the case of PLLA scaffolds. Both braided and stacked PCL and PLLA scaffolds upregulated the expression of Scx with a strong enhancement on braided PLLA scaffolds at day 7 of culture. Braided and stacked scaffolds: No differences were seen in the expression of tenogenic transcription factor Mkx and ECM glycoprotein Tnc. Stacked scaffolds downregulate the expression of Col Ia1 and Col IIIa1 compared to the braided ones. | [367] |
| Human ADSCs | Aligned scaffolds showed significant increase in Young’s modulus and yield stress along the axis of fiber alignment compared to random one after 28 days of culture. | Aligned/random scaffolds → no difference in cell proliferation, GAGs and Col I and Col III content. Both scaffolds expressed tendon related genes markers with ↑ mRNA Tnmd and Col3A1 in aligned scaffolds. | [368] |
| Human tendon Progenitor Stem Cells | n.d. | Aligned fibers →↑ tenogenic markers scleraxis, eya2, Col I, Col III, Col XIV, elastin, integrin α1, α5, β1 and myosin II. Random fibers: ↑ mRNA Ocn and Alp gene expression compared to aligned fibers. | [369] |
| Human rotator cuff fibroblast-like cells | Aligned cellular → Elastic modulus = 350 MPa; Ultimate stress = 6 MPa; Yield strength = 6 MPa Random cellular → Elastic modulus = 120 MPa; Ultimate stress = 1 MPa; Yield strength = 1 MPa | Aligned/random scaffolds: no difference in cell proliferation and adhesion. Cells acquired an elongated shape on the aligned scaffolds while maintaining their polygonal shape on the random ones. Aligned scaffolds: ↑ mRNA integrin α2 and Col I while similar mRNA gene expression for α5, β1 and Col III respect to random fibers. Similar matrix deposition in terms of Col I and Col III was seen with oriented collagen matrix along the aligned fiber. | [370] |
| Human tenocytes and human ADSCs | n.d. | Human tenocytes → cells elongated along the aligned fibers. All scaffolds types (random, aligned woven fabrics) expressed Tnmd and Col I. ↑ mRNA tendon-related genes (Tnc, Col III, Col II and Tnmd) in woven fabrics compared to aligned and random groups. Human ADSCs →↑ proliferation rate, cell infiltration, and ↑ mRNA of Scx, Col I, Tnmd gene expression on woven fabrics compared to aligned and random groups. Co-culture/tri-culture →↑ mRNA expression Scx, Tnc, Tnmd, A-VEGFA and ANGPT2 in the tri-culture system compared to other groups Dynamic culture by applying a 4% strain at frequency of 0.5 Hz for 2 h per day Dynamic/static culture →↑ Tnmd and Col protein and ↑ tendon related gene expression under dynamic stretch with tri-culture system compared to static condition. | [373] |
| Human ADSCs | n.d. | Aligned PCL/DT-NP5 yarns under static and magnetic stimulation conditions → no differences in cell activity while ↑ in cell alignment and elongation along the longitudinal direction of the fibers under magnetic stimulation. ↑ mRNA Dcn, Col Ia1, Col IIIa1, Tnc under both conditions while ↑ mRNA Tnmd and Scx and ↓ mRNA osteogenic marker (RUNX2) only under magnetic stimulation. | [304] |
| Human iPSC-MSCs | n.d. | Three days under static condition then for 7 days under 3% strain at 0.25 Hz for 2 h/day. PLLA/PCL scaffolds →↑ cell adhesion in PLLA compared to PCL. Cells were more elongated on PLLA scaffolds with 8 SPI compared to other groups (PCL 8 SPI, PLLA, and PCL 24 SPI). PLLA/PCL scaffolds → no difference in Scx and Tnmd mRNA expression while ↑ Col I, Col III, RUNX2, Ocn mRNA expressions and ↑ in Col I and Tnmd protein expressions in PLLA respect to PCL. Static/dynamic conditions →↓ RUNX2 and Ocn mRNA expression under cyclic condition compared to static one. Different braiding angles → cells showed elongated morphology on PLLA with 8 SPI while those seeded on PLLA with 24 SPI showed a rounder morphology. No difference in SCX and TNMD between PLLA and PCL with 8 and 24 stitches, ↓ Col I, Col III, RUNX2, and Ocn mRNA expression on PLLA with 8 SPI compared to 24 SPI after 10 days culture under cyclic condition. | [374] |
| Human rotator cuff fibroblast | n.d. | The cells were more aligned and elongated in the fibers with larger fiber diameter size. Smaller fiber diameter size →↑ cell proliferation and ↑ in collagen and proteoglycans synthesis. Larger fiber diameter size →↑ mRNA expression of Col I, Col III, Col V and Tnmd. | [375] |
| Human MSCs | n.d. | 1% strain at 1 Hz for 90 min twice a day Aligned/random scaffolds →↑ cell proliferation on random scaffolds after 28 days of dynamic culture with compared to other groups. Aligned/random → cells acquired an elongated morphology on aligned scaffolds (static and dynamic conditions) and on random scaffolds under dynamic conditions while those on random scaffolds remained cuboidal. Static/dynamic culture →↑ Col I ECM content in aligned (static) and random (dynamic) scaffolds while ↑ Col I and Col III ECM content on only aligned (dynamic) Aligned scaffolds under static/dynamic culture → no change in Col I and Tnmd mRNA expression between groups; ↑ Col III, fibronectin, and Tenascin-C and ↓ Scx mRNA expression under dynamic culture compared to static one after 28 days culture. ↑ integrin α2, α5, β1 expression on aligned scaffolds under dynamic condition. | [377] |
| Human rotator fibroblast | n.d. | Nano-/micro-fibers: ↑ cell adhesion, spreading and elongation by ↑ fiber diameter size. No differences in cell viability and proliferation. Nano-/micro-fibers: ↑ collagen content in nano-1 and nano-2 compared to micro scaffolds while ↑ Col I and Col III in the micron scaffold compared to the nano ones. ↓ mRNA expression α2 and ↑ mRNA expression β1 and ↑ mRNA expression RhoA and Rac1 on the nanofibers compared to microfibers after 7 days culture Aligned/random fibers: ↑ cell adhesion and alignment and ↓ cell proliferation onto aligned fibers compared to random ones. | [378] |
| Animal Model, Tissue Site, and Duration of Implantation | Mechanical Properties of the Scaffold Following Implantation | Biological Outcomes | Ref. |
|---|---|---|---|
| Rat Achilles Tendon model, 2 and 4 weeks | Aligned-TSA vs. Aligned vs. Random-TSA vs. Random Stiffness = 29 vs. 25 vs. 20 vs. 19 MPa Failure Force = 38 vs. 32 vs. 32 vs. 31 N Stress at Failure = 6.6 vs. 5 vs. 5.5 vs. 4.2 MPa Young’s Modulus = 51 vs. 33 vs. 31 vs. 32 MPa |
| [336] |
| Rat Achilles Tendon model, 2 and 4 weeks | Aligned vs. random Stiffness = 32.08 vs. 20.95 N/mm Failure Force = 50.47 vs. 42.85 N Stress at Failure = 5.91 vs. 4.90 MPa Young’s Modulus = 20.24 vs. 14.40 MPa | Aligned nanofiber: ↑ mRNA of tendon-related ECM gene markers Col Ia1, Col Va1 and Bgn, Scx, HoxA11, Tnmd and Fmod and ↓ mRNA of Ocn and RUNX2. ↑ deposition of ECM (Col I and Dcn) after 4 weeks implantation. | [340] |
| Mouse skeletal muscle, 1 and 6 weeks Mouse skin, 1 week | n.d. |
| [369] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Citeroni, M.R.; Ciardulli, M.C.; Russo, V.; Della Porta, G.; Mauro, A.; El Khatib, M.; Di Mattia, M.; Galesso, D.; Barbera, C.; Forsyth, N.R.; et al. In Vitro Innovation of Tendon Tissue Engineering Strategies. Int. J. Mol. Sci. 2020, 21, 6726. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186726
Citeroni MR, Ciardulli MC, Russo V, Della Porta G, Mauro A, El Khatib M, Di Mattia M, Galesso D, Barbera C, Forsyth NR, et al. In Vitro Innovation of Tendon Tissue Engineering Strategies. International Journal of Molecular Sciences. 2020; 21(18):6726. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186726
Chicago/Turabian StyleCiteroni, Maria Rita, Maria Camilla Ciardulli, Valentina Russo, Giovanna Della Porta, Annunziata Mauro, Mohammad El Khatib, Miriam Di Mattia, Devis Galesso, Carlo Barbera, Nicholas R. Forsyth, and et al. 2020. "In Vitro Innovation of Tendon Tissue Engineering Strategies" International Journal of Molecular Sciences 21, no. 18: 6726. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186726