Dissecting the Effect of a 3D Microscaffold on the Transcriptome of Neural Stem Cells with Computational Approaches: A Focus on Mechanotransduction

 ,
,  , , , ,
, , , ,  and
and
Abstract
:
1. Introduction
2. Results
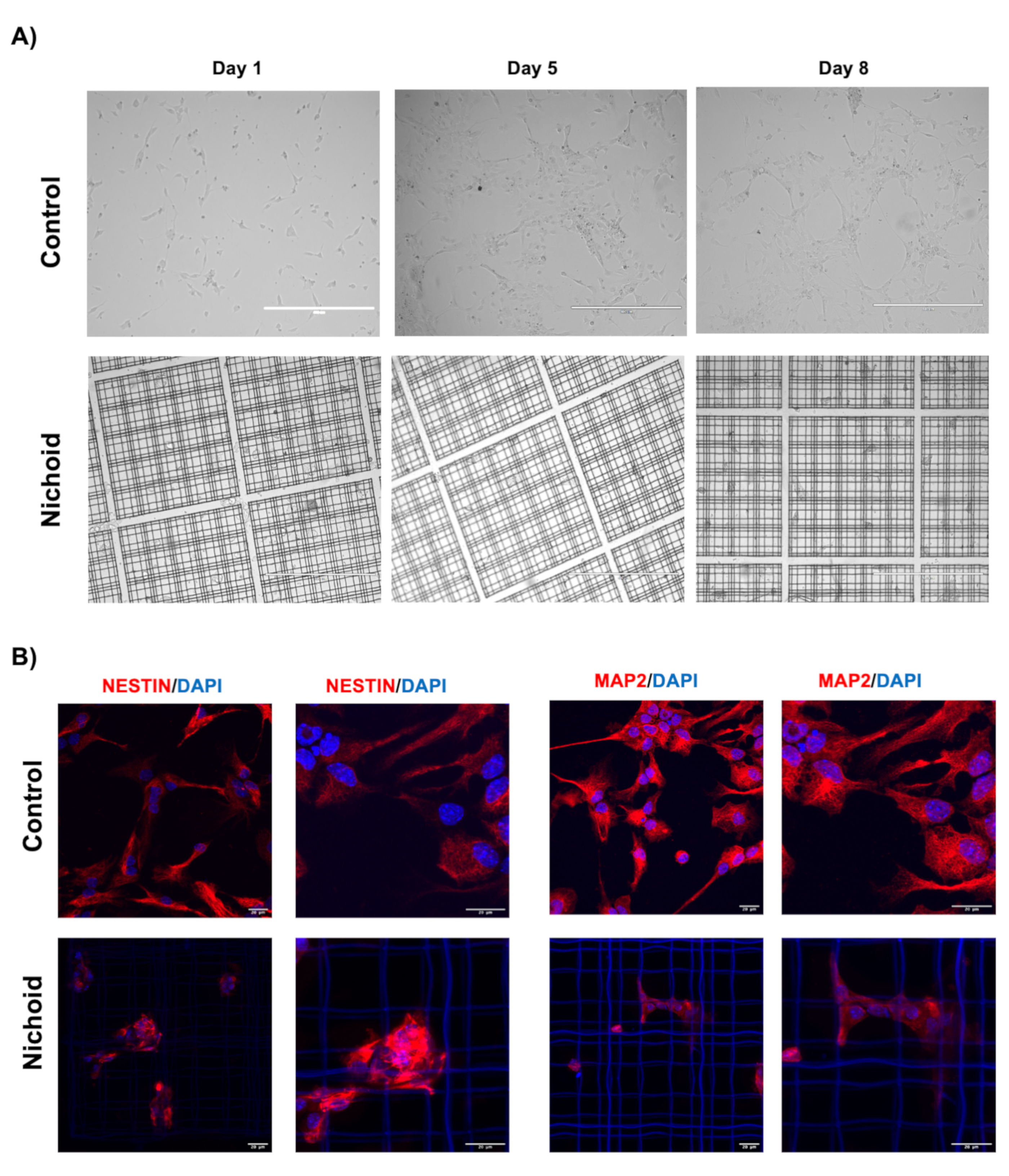
2.1. Neural Precursor Cells (NPCs) Expanded inside the Nichoid Present with Different Cellular Organization and Growing Capabilities
2.2. Deep Sequencing RNAs Expression Profiles in NPCs Grown inside the NICHOID vs. Control Conditions
2.3. Role of the Noncoding Transcriptome
2.4. Pathway Analysis of Deregulated Transcripts
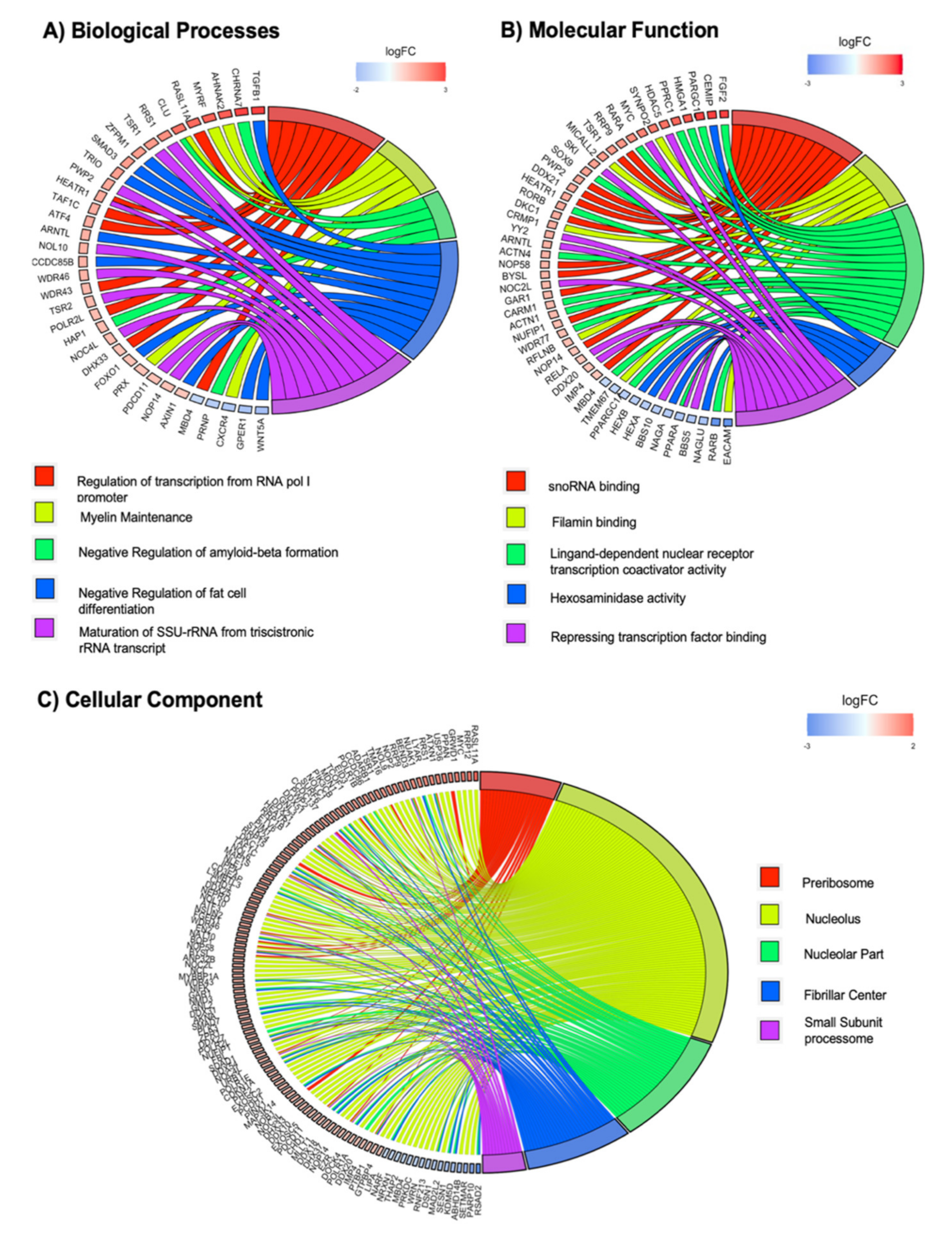
2.5. GO Terms Enrichment Show Significant Deregulation of Genes Implicated in Mechanotransduction via the Nichoid Scaffold
2.6. NPCs Grown inside Nichoid Differently Express Cytoskeleton and Membrane Organization-Related Genes
2.7. Cellular Growth inside the Nichoid Influences Genes Implicated in Signaling Transduction Pathways of Mechanotransduction
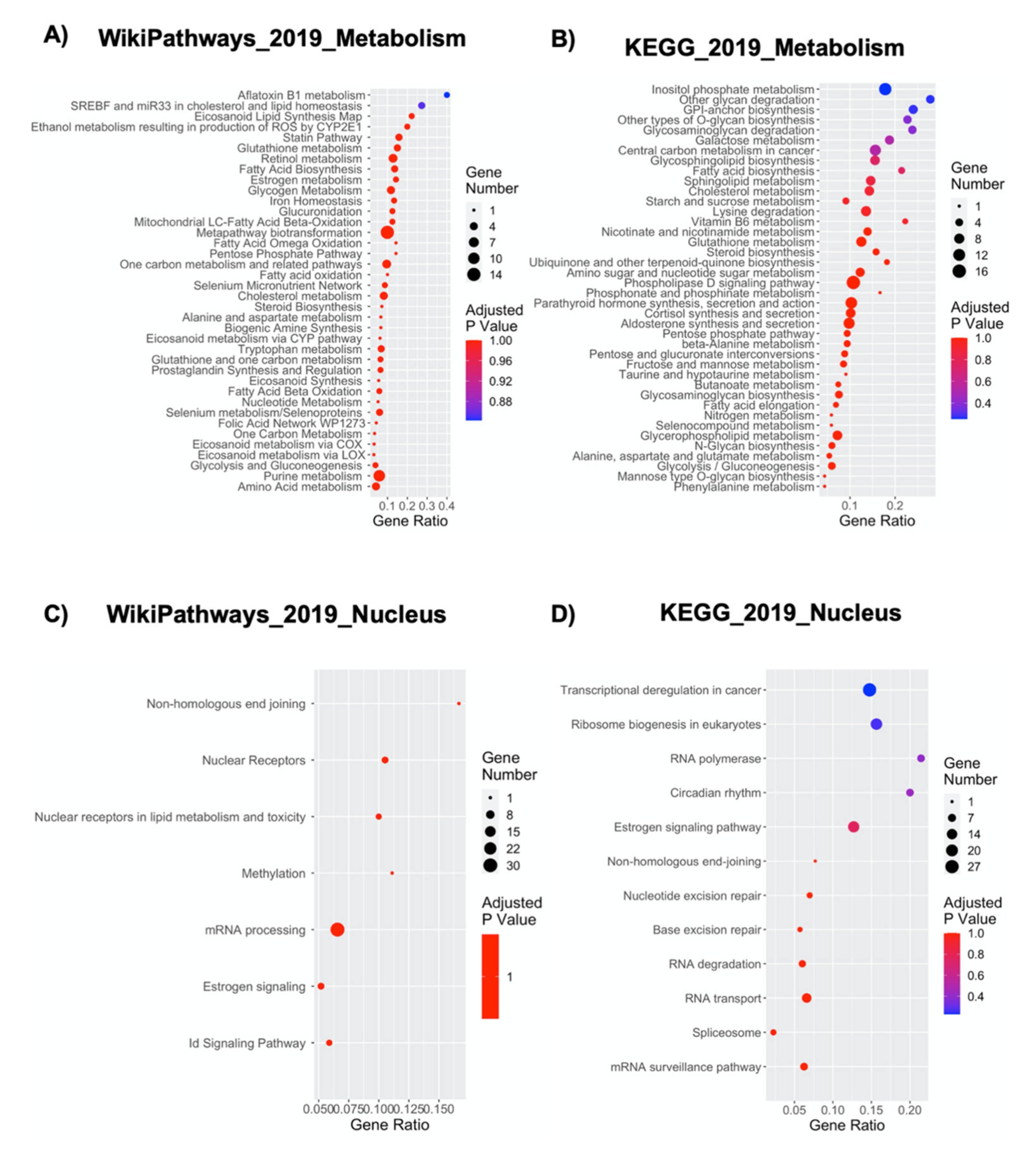
2.8. NPCs’ Growth inside the Nichoid Significantly Impacts Cellular Metabolism-Related Genes
2.9. The Nichoid Scaffold Influences Nuclear Compartment Processes through Alteration of Related Gene Expression
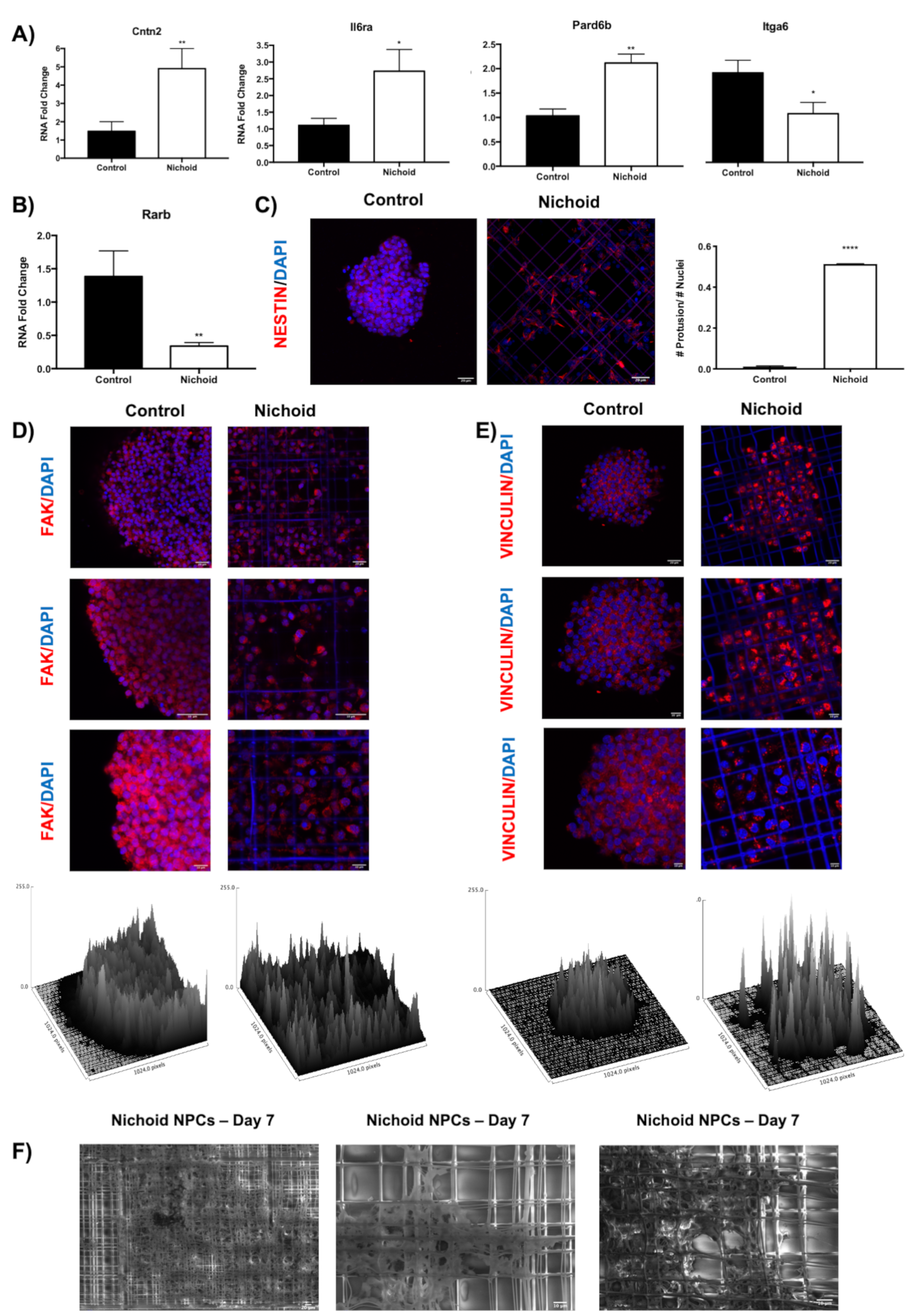
2.10. Influence of the Nichoid on Cytoskeletal Organization and Adhesion Complexes Formation
2.11. Deregulation of Genes Involved in Regeneration and Neural-Related Processes Highlights Potential Translational Applications
2.12. Nichoid Grown NPCs Are Able to Differentiate to a Neuronal Phenotype
2.13. Deep Sequencing RNAs Expression Profiles and Pathways Analysis in NPCs Replated after Nichoid-Expansion vs. Control Conditions
3. Discussion
4. Materials and Methods
4.1. Nichoid Microfabrication
4.2. Substrate Preparation
4.3. Primary Cells Isolation and Culture
4.4. Cells’ Seeding in the Nichoid
4.5. Cells’ Detachment from the Nichoid and Proliferation Assay
4.6. Environmental Scanning Electron Microscopy
4.7. RNA Extraction
4.8. Libraries Preparation for RNA-Seq and Bioinformatic Data Analysis
4.9. Pathway and Coexpression Analysis
4.10. Real-Time PCR
4.11. Primary Cells Isolation and Differentiation
4.12. Immunocytochemistry Analysis for Differentiated Cells
4.13. Immunocytochemistry Analysis for NPCs in Standard Conditions
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 3D | Three-dimensional |
| AMPK | AMP-activated protein kinase |
| ceRNA | Competing endogenous RNA |
| DE RNAs | Differentially expressed coding and noncoding RNAs |
| ECM | Extracellular matrix |
| ESEM | Environmental scanning electron microscopy |
| GO | Gene Ontology |
| GPCRs | G-protein coupled receptors |
| KEGG | Kyoto Encyclopedia of Genes and Genome |
| lncRNA | Long noncoding RNA |
| NPCs | Neural precursors stem cells |
| S1P | Sphingosine-1-phosphate |
| TGF-β | Transforming growth factor-beta |
Appendix A
References
- Kumar, G.; Tison, C.K.; Chatterjee, K.; Pine, P.S.; McDaniel, J.H.; Salit, M.L.; Young, M.F.; Simon, C.G. The determination of stem cell fate by 3D scaffold structures through the control of cell shape. Biomaterials 2011, 32, 9188–9196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, O.Z.; Khademhosseini, A.; Langer, R.; Peppas, N.A. Bioinspired materials for controlling stem cell fate. Acc. Chem. Res. 2010, 43, 419–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kshitiz; Park, J.; Kim, P.; Helen, W.; Engler, A.J.; Levchenko, A.; Kim, D.H. Control of stem cell fate and function by engineering physical microenvironments. Integr. Biol. 2012, 4, 1008–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, M.; Li, W.; Dong, X.; Yuan, X.; Midgley, A.C.; Chang, H.; Wang, Y.; Wang, H.; Wang, K.; Ma, P.X.; et al. In vivo engineered extracellular matrix scaffolds with instructive niches for oriented tissue regeneration. Nat. Commun. 2019, 10, 4620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkins, T.L.; Little, D. Synthetic scaffolds for musculoskeletal tissue engineering: Cellular responses to fiber parameters. NPJ Regen. Med. 2019, 4, 15. [Google Scholar] [CrossRef]
- Persson, M.; Lehenkari, P.P.; Berglin, L.; Turunen, S.; Finnilä, M.A.J.; Risteli, J.; Skrifvars, M.; Tuukkanen, J. Osteogenic Differentiation of Human Mesenchymal Stem cells in a 3D Woven Scaffold. Sci. Rep. 2018, 8, 10457. [Google Scholar] [CrossRef]
- Rey, F.; Barzaghini, B.; Nardini, A.; Bordoni, M.; Zuccotti, G.V.; Cereda, C.; Raimondi, M.T.; Carelli, S. Advances in Tissue Engineering and Innovative Fabrication Techniques for 3-D-Structures: Translational Applications in Neurodegenerative Diseases. Cells 2020, 9, 1636. [Google Scholar] [CrossRef]
- Bordoni, M.; Scarian, E.; Rey, F.; Gagliardi, S.; Carelli, S.; Pansarasa, O.; Cereda, C. Biomaterials in Neurodegenerative Disorders: A Promising Therapeutic Approach. Int. J. Mol. Sci. 2020, 21, 3243. [Google Scholar] [CrossRef]
- Nava, M.M.; Piuma, A.; Figliuzzi, M.; Cattaneo, I.; Bonandrini, B.; Zandrini, T.; Cerullo, G.; Osellame, R.; Remuzzi, A.; Raimondi, M.T. Two-photon polymerized “nichoid” substrates maintain function of pluripotent stem cells when expanded under feeder-free conditions. Stem Cell Res. Ther. 2016, 7, 132. [Google Scholar] [CrossRef] [Green Version]
- Raimondi, M.T.; Eaton, S.M.; Laganà, M.; Aprile, V.; Nava, M.M.; Cerullo, G.; Osellame, R. Three-dimensional structural niches engineered via two-photon laser polymerization promote stem cell homing. Acta Biomater. 2013, 9, 4579–4584. [Google Scholar] [CrossRef]
- Raimondi, M.T.; Eaton, S.M.; Nava, M.M.; Laganà, M.; Cerullo, G.; Osellame, R. Two-photon laser polymerization: From fundamentals to biomedical application in tissue engineering and regenerative medicine. J. Appl. Biomater. Funct. Mater. 2012, 10, 55–65. [Google Scholar] [CrossRef]
- Zandrini, T.; Shan, O.; Parodi, V.; Cerullo, G.; Raimondi, M.T.; Osellame, R. Multi-foci laser microfabrication of 3D polymeric scaffolds for stem cell expansion in regenerative medicine. Sci. Rep. 2019, 9, 11761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raimondi, M.T.; Nava, M.M.; Eaton, S.M.; Bernasconi, A.; Vishnubhatla, K.C.; Cerullo, G.; Osellame, R. Optimization of Femtosecond Laser Polymerized Structural Niches to Control Mesenchymal Stromal Cell Fate in Culture. Micromachines 2014, 5, 341–358. [Google Scholar] [CrossRef] [Green Version]
- Nava, M.M.; Raimondi, M.T.; Pietrabissa, R. Controlling self-renewal and differentiation of stem cells via mechanical cues. J. Biomed. Biotechnol. 2012, 2012, 797410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricci, D.; Nava, M.M.; Zandrini, T.; Cerullo, G.; Raimondi, M.T.; Osellame, R. Scaling-Up Techniques for the Nanofabrication of Cell Culture Substrates via Two-Photon Polymerization for Industrial-Scale Expansion of Stem Cells. Materials 2017, 10, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruitt, B.L.; Dunn, A.R.; Weis, W.I.; Nelson, W.J. Mechano-transduction: From molecules to tissues. PLoS Biol. 2014, 12, e1001996. [Google Scholar] [CrossRef] [PubMed]
- Boeri, L.; Albani, D.; Raimondi, M.T.; Jacchetti, E. Mechanical regulation of nucleocytoplasmic translocation in mesenchymal stem cells: Characterization and methods for investigation. Biophys. Rev. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kjell, J.; Fischer-Sternjak, J.; Thompson, A.J.; Friess, C.; Sticco, M.J.; Salinas, F.; Cox, J.; Martinelli, D.C.; Ninkovic, J.; Franze, K.; et al. Defining the Adult Neural Stem Cell Niche Proteome Identifies Key Regulators of Adult Neurogenesis. Cell Stem Cell 2020, 26, 277–293.e278. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liao, K.; Li, C.; Lai, A.C.K.; Foo, J.J.; Chan, V. Progress in Integrative Biomaterial Systems to Approach Three-Dimensional Cell Mechanotransduction. Bioengineering 2017, 4, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geiger, B.; Spatz, J.P.; Bershadsky, A.D. Environmental sensing through focal adhesions. Nat. Rev. Mol. Cell Biol. 2009, 10, 21–33. [Google Scholar] [CrossRef]
- Wang, N. Review of Cellular Mechanotransduction. J. Phys. D Appl. Phys. 2017, 50. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Tytell, J.D.; Ingber, D.E. Mechanotransduction at a distance: Mechanically coupling the extracellular matrix with the nucleus. Nat. Rev. Mol. Cell Biol. 2009, 10, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Lin, J.; Roy, K. Effect of 3D scaffold and dynamic culture condition on the global gene expression profile of mouse embryonic stem cells. Biomaterials 2006, 27, 5978–5989. [Google Scholar] [CrossRef]
- Salvi, A.M.; DeMali, K.A. Mechanisms linking mechanotransduction and cell metabolism. Curr. Opin. Cell Biol. 2018, 54, 114–120. [Google Scholar] [CrossRef]
- Sun, Z.; Guo, S.S.; Fässler, R. Integrin-mediated mechanotransduction. J. Cell Biol. 2016, 215, 445–456. [Google Scholar] [CrossRef]
- Guilak, F.; Cohen, D.M.; Estes, B.T.; Gimble, J.M.; Liedtke, W.; Chen, C.S. Control of stem cell fate by physical interactions with the extracellular matrix. Cell Stem Cell 2009, 5, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Ortinau, S.; Schmich, J.; Block, S.; Liedmann, A.; Jonas, L.; Weiss, D.G.; Helm, C.A.; Rolfs, A.; Frech, M.J. Effect of 3D-scaffold formation on differentiation and survival in human neural progenitor cells. Biomed. Eng. Online 2010, 9, 70. [Google Scholar] [CrossRef] [Green Version]
- Marfia, G.; Madaschi, L.; Marra, F.; Menarini, M.; Bottai, D.; Formenti, A.; Bellardita, C.; Di Giulio, A.M.; Carelli, S.; Gorio, A. Adult neural precursors isolated from post mortem brain yield mostly neurons: An erythropoietin-dependent process. Neurobiol. Dis. 2011, 43, 86–98. [Google Scholar] [CrossRef]
- Carelli, S.; Giallongo, T.; Gombalova, Z.; Rey, F.; Gorio, M.C.F.; Mazza, M.; Di Giulio, A.M. Counteracting neuroinflammation in experimental Parkinson’s disease favors recovery of function: Effects of Er-NPCs administration. J. Neuroinflamm. 2018, 15, 333. [Google Scholar] [CrossRef]
- Carelli, S.; Giallongo, T.; Viaggi, C.; Latorre, E.; Gombalova, Z.; Raspa, A.; Mazza, M.; Vaglini, F.; Di Giulio, A.M.; Gorio, A. Recovery from experimental parkinsonism by intrastriatal application of erythropoietin or EPO-releasing neural precursors. Neuropharmacology 2017, 119, 76–90. [Google Scholar] [CrossRef] [PubMed]
- Carelli, S.; Giallongo, T.; Gombalova, Z.; Merli, D.; Di Giulio, A.M.; Gorio, A. EPO-releasing neural precursor cells promote axonal regeneration and recovery of function in spinal cord traumatic injury. Restor. Neurol. Neurosci. 2017, 35, 583–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carelli, S.; Giallongo, T.; Viaggi, C.; Gombalova, Z.; Latorre, E.; Mazza, M.; Vaglini, F.; Di Giulio, A.M.; Gorio, A. Grafted Neural Precursors Integrate Into Mouse Striatum, Differentiate and Promote Recovery of Function Through Release of Erythropoietin in MPTP-Treated Mice. ASN Neuro 2016, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carelli, S.; Giallongo, T.; Marfia, G.; Merli, D.; Ottobrini, L.; Degrassi, A.; Basso, M.D.; Di Giulio, A.M.; Gorio, A. Exogenous adult postmortem neural precursors attenuate secondary degeneration and promote myelin sparing and functional recovery following experimental spinal cord injury. Cell Transplant. 2015, 24, 703–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carelli, S.; Giallongo, T.; Latorre, E.; Caremoli, F.; Gerace, C.; Basso, D.M.; Di Giulio, A.M.; Gorio, A. Adult Mouse Post Mortem Neural Precursors Survive, Differentiate, Counteract Cytokine Production and Promote Functional Recovery After Transplantation in Experimental Traumatic Spinal Cord Injury. Stem Cell Res. Transplant. 2014, 1, 1008. [Google Scholar]
- Carelli, S.; Giallongo, T.; Gerace, C.; De Angelis, A.; Basso, M.D.; Di Giulio, A.M.; Gorio, A. Neural stem cell transplantation in experimental contusive model of spinal cord injury. J. Vis. Exp. 2014. [Google Scholar] [CrossRef] [Green Version]
- Madrazo, I.; Kopyov, O.; Ávila-Rodríguez, M.A.; Ostrosky, F.; Carrasco, H.; Kopyov, A.; Avendaño-Estrada, A.; Jiménez, F.; Magallón, E.; Zamorano, C.; et al. Transplantation of Human Neural Progenitor Cells (NPC) into Putamina of Parkinsonian Patients: A Case Series Study, Safety and Efficacy Four Years after Surgery. Cell Transplant. 2019, 28, 269–285. [Google Scholar] [CrossRef]
- Kim, D.H.; Marinov, G.K.; Pepke, S.; Singer, Z.S.; He, P.; Williams, B.; Schroth, G.P.; Elowitz, M.B.; Wold, B.J. Single-cell transcriptome analysis reveals dynamic changes in lncRNA expression during reprogramming. Cell Stem Cell 2015, 16, 88–101. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Yan, J.; Liu, Q.; Xie, Z.; Jiang, H. LncRNA Rik-203 contributes to anesthesia neurotoxicity via microRNA-101a-3p and GSK-3β-mediated neural differentiation. Sci. Rep. 2019, 9, 6822. [Google Scholar] [CrossRef]
- Chen, Z.; Bu, N.; Qiao, X.; Zuo, Z.; Shu, Y.; Liu, Z.; Qian, Z.; Chen, J.; Hou, Y. Forkhead Box M1 Transcriptionally Regulates the Expression of Long Noncoding RNAs Snhg8 and Gm26917 to Promote Proliferation and Survival of Muscle Satellite Cells. Stem Cells 2018, 36, 1097–1108. [Google Scholar] [CrossRef] [Green Version]
- Marín-Béjar, O.; Marchese, F.P.; Athie, A.; Sánchez, Y.; González, J.; Segura, V.; Huang, L.; Moreno, I.; Navarro, A.; Monzó, M.; et al. Pint lincRNA connects the p53 pathway with epigenetic silencing by the Polycomb repressive complex 2. Genome Biol. 2013, 14, R104. [Google Scholar] [CrossRef] [Green Version]
- Huarte, M.; Guttman, M.; Feldser, D.; Garber, M.; Koziol, M.J.; Kenzelmann-Broz, D.; Khalil, A.M.; Zuk, O.; Amit, I.; Rabani, M.; et al. A large intergenic noncoding RNA induced by p53 mediates global gene repression in the p53 response. Cell 2010, 142, 409–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carelli, S.; Giallongo, T.; Rey, F.; Latorre, E.; Bordoni, M.; Mazzucchelli, S.; Gorio, M.C.; Pansarasa, O.; Provenzani, A.; Cereda, C.; et al. HuR interacts with lincBRN1a and lincBRN1b during neuronal stem cells differentiation. RNA Biol. 2019, 16, 1471–1485. [Google Scholar] [CrossRef] [PubMed]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruce, D.L.; Macartney, T.; Yong, W.; Shou, W.; Sapkota, G.P. Protein phosphatase 5 modulates SMAD3 function in the transforming growth factor-β pathway. Cell. Signal. 2012, 24, 1999–2006. [Google Scholar] [CrossRef] [Green Version]
- Qin, Y.; Garrison, B.S.; Ma, W.; Wang, R.; Jiang, A.; Li, J.; Mistry, M.; Bronson, R.T.; Santoro, D.; Franco, C.; et al. A Milieu Molecule for TGF-β Required for Microglia Function in the Nervous System. Cell 2018, 174, 156–171.e116. [Google Scholar] [CrossRef] [Green Version]
- Rys, J.P.; Monteiro, D.A.; Alliston, T. Mechanobiology of TGFβ signaling in the skeleton. Matrix Biol. 2016, 52, 413–425. [Google Scholar] [CrossRef] [Green Version]
- Hinz, B. The extracellular matrix and transforming growth factor-β1: Tale of a strained relationship. Matrix Biol. 2015, 47, 54–65. [Google Scholar] [CrossRef]
- Chen, X.D.; Su, M.Y.; Chen, T.T.; Hong, H.Y.; Han, A.D.; Li, W.S. Oxidative stress affects retinal pigment epithelial cell survival through epidermal growth factor receptor/AKT signaling pathway. Int. J. Ophthalmol. 2017, 10, 507–514. [Google Scholar] [CrossRef]
- Rosa, J.P.; Raslova, H.; Bryckaert, M. Filamin A: Key actor in platelet biology. Blood 2019, 134, 1279–1288. [Google Scholar] [CrossRef]
- Louro, R.; Nakaya, H.I.; Paquola, A.C.; Martins, E.A.; da Silva, A.M.; Verjovski-Almeida, S.; Reis, E.M. RASL11A, member of a novel small monomeric GTPase gene family, is down-regulated in prostate tumors. Biochem. Biophys. Res. Commun. 2004, 316, 618–627. [Google Scholar] [CrossRef]
- Burridge, K.; Monaghan-Benson, E.; Graham, D.M. Mechanotransduction: From the cell surface to the nucleus via RhoA. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20180229. [Google Scholar] [CrossRef] [PubMed]
- Alenghat, F.J.; Ingber, D.E. Mechanotransduction: All signals point to cytoskeleton, matrix, and integrins. Sci. STKE 2002, 2002, pe6. [Google Scholar] [CrossRef] [PubMed]
- Canovas Nunes, S.; Manzoni, M.; Pizzi, M.; Mandato, E.; Carrino, M.; Quotti Tubi, L.; Zambello, R.; Adami, F.; Visentin, A.; Barilà, G.; et al. The small GTPase RhoU lays downstream of JAK/STAT signaling and mediates cell migration in multiple myeloma. Blood Cancer J. 2018, 8, 20. [Google Scholar] [CrossRef]
- Leszczynska, K.; Kaur, S.; Wilson, E.; Bicknell, R.; Heath, V.L. The role of RhoJ in endothelial cell biology and angiogenesis. Biochem. Soc. Trans. 2011, 39, 1606–1611. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.A.; DeSimone, D.W. Cell adhesion receptors in mechanotransduction. Curr. Opin. Cell Biol. 2008, 20, 551–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mui, K.L.; Chen, C.S.; Assoian, R.K. The mechanical regulation of integrin-cadherin crosstalk organizes cells, signaling and forces. J. Cell Sci. 2016, 129, 1093–1100. [Google Scholar] [CrossRef] [Green Version]
- Young, N.; Van Brocklyn, J.R. Signal transduction of sphingosine-1-phosphate G protein-coupled receptors. Sci. World J. 2006, 6, 946–966. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Chen, Y.J.; Ono, T.; Wang, J.J. Crosstalk between integrin and G protein pathways involved in mechanotransduction in mandibular condylar chondrocytes under pressure. Arch. Biochem. Biophys. 2008, 474, 102–108. [Google Scholar] [CrossRef]
- Morrison, D.K. MAP kinase pathways. Cold Spring Harb. Perspect. Biol. 2012, 4. [Google Scholar] [CrossRef]
- Shaul, Y.D.; Seger, R. The MEK/ERK cascade: From signaling specificity to diverse functions. Biochim. Biophys. Acta 2007, 1773, 1213–1226. [Google Scholar] [CrossRef] [Green Version]
- Komiya, Y.; Habas, R. Wnt signal transduction pathways. Organogenesis 2008, 4, 68–75. [Google Scholar] [CrossRef] [Green Version]
- Chrzanowska-Wodnicka, M. Rap1 in endothelial biology. Curr. Opin. Hematol. 2017, 24, 248–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercurio, A.M.; Rabinovitz, I.; Shaw, L.M. The alpha 6 beta 4 integrin and epithelial cell migration. Curr. Opin. Cell Biol. 2001, 13, 541–545. [Google Scholar] [CrossRef]
- Issaeva, N. p53 Signaling in Cancers. Cancers 2019, 11, 332. [Google Scholar] [CrossRef] [Green Version]
- DeBerardinis, R.J.; Thompson, C.B. Cellular metabolism and disease: What do metabolic outliers teach us? Cell 2012, 148, 1132–1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ata, R.; Antonescu, C.N. Integrins and Cell Metabolism: An Intimate Relationship Impacting Cancer. Int. J. Mol. Sci. 2017, 18, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romani, P.; Brian, I.; Santinon, G.; Pocaterra, A.; Audano, M.; Pedretti, S.; Mathieu, S.; Forcato, M.; Bicciato, S.; Manneville, J.B.; et al. Extracellular matrix mechanical cues regulate lipid metabolism through Lipin-1 and SREBP. Nat. Cell Biol. 2019, 21, 338–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Head, B.P.; Patel, H.H.; Insel, P.A. Interaction of membrane/lipid rafts with the cytoskeleton: Impact on signaling and function: Membrane/lipid rafts, mediators of cytoskeletal arrangement and cell signaling. Biochim. Biophys. Acta 2014, 1838, 532–545. [Google Scholar] [CrossRef] [Green Version]
- Boulter, E.; Estrach, S.; Tissot, F.S.; Hennrich, M.L.; Tosello, L.; Cailleteau, L.; de la Ballina, L.R.; Pisano, S.; Gavin, A.C.; Féral, C.C. Cell metabolism regulates integrin mechanosensing via an SLC3A2-dependent sphingolipid biosynthesis pathway. Nat. Commun. 2018, 9, 4862. [Google Scholar] [CrossRef] [PubMed]
- Bays, J.L.; Campbell, H.K.; Heidema, C.; Sebbagh, M.; DeMali, K.A. Linking E-cadherin mechanotransduction to cell metabolism through force-mediated activation of AMPK. Nat. Cell Biol. 2017, 19, 724–731. [Google Scholar] [CrossRef] [Green Version]
- Hardie, D.G. AMPK—Sensing energy while talking to other signaling pathways. Cell Metab. 2014, 20, 939–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahl, K.N.; Ribeiro, A.J.; Lammerding, J. Nuclear shape, mechanics, and mechanotransduction. Circ. Res. 2008, 102, 1307–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, S.; Irianto, J.; Discher, D.E. Mechanosensing by the nucleus: From pathways to scaling relationships. J. Cell Biol. 2017, 216, 305–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guilluy, C.; Burridge, K. Nuclear mechanotransduction: Forcing the nucleus to respond. Nucleus 2015, 6, 19–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Booth-Gauthier, E.A.; Alcoser, T.A.; Yang, G.; Dahl, K.N. Force-induced changes in subnuclear movement and rheology. Biophys. J. 2012, 103, 2423–2431. [Google Scholar] [CrossRef] [Green Version]
- Camozzi, D.; Capanni, C.; Cenni, V.; Mattioli, E.; Columbaro, M.; Squarzoni, S.; Lattanzi, G. Diverse lamin-dependent mechanisms interact to control chromatin dynamics. Focus on laminopathies. Nucleus 2014, 5, 427–440. [Google Scholar] [CrossRef] [Green Version]
- Ivorra, C.; Kubicek, M.; González, J.M.; Sanz-González, S.M.; Alvarez-Barrientos, A.; O’Connor, J.E.; Burke, B.; Andrés, V. A mechanism of AP-1 suppression through interaction of c-Fos with lamin A/C. Genes Dev. 2006, 20, 307–320. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, K.S.; Tapley, E.C.; Cruz, V.E.; Li, Q.; Aung, K.; Hart, K.C.; Schwartz, T.U.; Starr, D.A.; Engebrecht, J. LINC complexes promote homologous recombination in part through inhibition of nonhomologous end joining. J. Cell Biol. 2016, 215, 801–821. [Google Scholar] [CrossRef]
- Maynard, S.; Keijzers, G.; Akbari, M.; Ezra, M.B.; Hall, A.; Morevati, M.; Scheibye-Knudsen, M.; Gonzalo, S.; Bartek, J.; Bohr, V.A. Lamin A/C promotes DNA base excision repair. Nucleic Acids Res. 2019, 47, 11709–11728. [Google Scholar] [CrossRef]
- Uhler, C.; Shivashankar, G.V. Regulation of genome organization and gene expression by nuclear mechanotransduction. Nat. Rev. Mol. Cell Biol. 2017, 18, 717–727. [Google Scholar] [CrossRef]
- González, S.; Aguilera, S.; Urzúa, U.; Quest, A.F.; Molina, C.; Alliende, C.; Hermoso, M.; González, M.J. Mechanotransduction and epigenetic control in autoimmune diseases. Autoimmun. Rev 2011, 10, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Mendez, M.G.; Janmey, P.A. Transcription factor regulation by mechanical stress. Int. J. Biochem. Cell Biol. 2012, 44, 728–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, D.H.; Stuwe, T.; Schilbach, S.; Rundlet, E.J.; Perriches, T.; Mobbs, G.; Fan, Y.; Thierbach, K.; Huber, F.M.; Collins, L.N.; et al. Architecture of the symmetric core of the nuclear pore. Science 2016, 352, aaf1015. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Hamilla, S.; Cam, M.; Aranda-Espinoza, H.; Mili, S. Extracellular matrix stiffness and cell contractility control RNA localization to promote cell migration. Nat. Commun. 2017, 8, 896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirby, T.J. Mechanosensitive pathways controlling translation regulatory processes in skeletal muscle and implications for adaptation. J. Appl. Physiol. 2019, 127, 608–618. [Google Scholar] [CrossRef]
- Liu, H.; Tang, L. Mechano-regulation of alternative splicing. Curr. Genom. 2013, 14, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Soheilypour, M.; Mofrad, M.R.K. Quality control of mRNAs at the entry of the nuclear pore: Cooperation in a complex molecular system. Nucleus 2018, 9, 202–211. [Google Scholar] [CrossRef] [Green Version]
- Masuda, T. Contactin-2/TAG-1, active on the front line for three decades. Cell Adhes. Migr. 2017, 11, 524–531. [Google Scholar] [CrossRef]
- Walsh, F.S.; Doherty, P. Glycosylphosphatidylinositol anchored recognition molecules that function in axonal fasciculation, growth and guidance in the nervous system. Cell Biol. Int. Rep. 1991, 15, 1151–1166. [Google Scholar] [CrossRef]
- Gennarini, G.; Bizzoca, A.; Picocci, S.; Puzzo, D.; Corsi, P.; Furley, A.J.W. The role of Gpi-anchored axonal glycoproteins in neural development and neurological disorders. Mol. Cell Neurosci. 2017, 81, 49–63. [Google Scholar] [CrossRef]
- Nishimoto, N.; Kishimoto, T. Interleukin 6: From bench to bedside. Nat. Clin. Pract. Rheumatol. 2006, 2, 619–626. [Google Scholar] [CrossRef]
- Hao, Y.; Wang, Y.; Liu, X.; Yang, X.; Wang, P.; Tian, Q.; Bai, Q.; Chen, X.; Li, Z.; Wu, J.; et al. The Kinase Complex mTOR Complex 2 Promotes the Follicular Migration and Functional Maturation of Differentiated Follicular Helper CD4. Front. Immunol. 2018, 9, 1127. [Google Scholar] [CrossRef]
- Doe, C.Q. Cell polarity: The PARty expands. Nat. Cell Biol. 2001, 3, E7–E9. [Google Scholar] [CrossRef]
- Cunliffe, H.E.; Jiang, Y.; Fornace, K.M.; Yang, F.; Meltzer, P.S. PAR6B is required for tight junction formation and activated PKCζ localization in breast cancer. Am. J. Cancer Res. 2012, 2, 478–491. [Google Scholar]
- Wang, S.; Cha, S.W.; Zorn, A.M.; Wylie, C. Par6b regulates the dynamics of apicobasal polarity during development of the stratified Xenopus epidermis. PLoS ONE 2013, 8, e76854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchetti, G.; De Arcangelis, A.; Pfister, V.; Georges-Labouesse, E. α6 integrin subunit regulates cerebellar development. Cell Adhes. Migr. 2013, 7, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Ivanovska, I.L.; Shin, J.W.; Swift, J.; Discher, D.E. Stem cell mechanobiology: Diverse lessons from bone marrow. Trends Cell Biol. 2015, 25, 523–532. [Google Scholar] [CrossRef] [Green Version]
- Bernal, A.; Arranz, L. Nestin-expressing progenitor cells: Function, identity and therapeutic implications. Cell. Mol. Life Sci. 2018, 75, 2177–2195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gritti, A.; Bonfanti, L.; Doetsch, F.; Caille, I.; Alvarez-Buylla, A.; Lim, D.A.; Galli, R.; Verdugo, J.M.; Herrera, D.G.; Vescovi, A.L. Multipotent neural stem cells reside into the rostral extension and olfactory bulb of adult rodents. J. Neurosci. 2002, 22, 437–445. [Google Scholar] [CrossRef] [Green Version]
- Castillo, A.B.; Blundo, J.T.; Chen, J.C.; Lee, K.L.; Yereddi, N.R.; Jang, E.; Kumar, S.; Tang, W.J.; Zarrin, S.; Kim, J.B.; et al. Focal adhesion kinase plays a role in osteoblast mechanotransduction in vitro but does not affect load-induced bone formation in vivo. PLoS ONE 2012, 7, e43291. [Google Scholar] [CrossRef]
- Paillard, T.; Rolland, Y.; de Souto Barreto, P. Protective Effects of Physical Exercise in Alzheimer’s Disease and Parkinson’s Disease: A Narrative Review. J. Clin. Neurol. 2015, 11, 212–219. [Google Scholar] [CrossRef] [Green Version]
- Reichardt, L.F. Neurotrophin-regulated signalling pathways. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006, 361, 1545–1564. [Google Scholar] [CrossRef] [Green Version]
- Chédotal, A. Roles of axon guidance molecules in neuronal wiring in the developing spinal cord. Nat. Rev. Neurosci. 2019, 20, 380–396. [Google Scholar] [CrossRef]
- Liu, E.Y.; Cali, C.P.; Lee, E.B. RNA metabolism in neurodegenerative disease. Dis. Model. Mech. 2017, 10, 509–518. [Google Scholar] [CrossRef] [Green Version]
- Andersen, R.E.; Lim, D.A. Forging our understanding of lncRNAs in the brain. Cell Tissue Res. 2018, 371, 55–71. [Google Scholar] [CrossRef]
- Gagliardi, S.; Zucca, S.; Pandini, C.; Diamanti, L.; Bordoni, M.; Sproviero, D.; Arigoni, M.; Olivero, M.; Pansarasa, O.; Ceroni, M.; et al. Long non-coding and coding RNAs characterization in Peripheral Blood Mononuclear Cells and Spinal Cord from Amyotrophic Lateral Sclerosis patients. Sci. Rep. 2018, 8, 2378. [Google Scholar] [CrossRef]
- Humphrey, J.D.; Dufresne, E.R.; Schwartz, M.A. Mechanotransduction and extracellular matrix homeostasis. Nat. Rev. Mol. Cell Biol. 2014, 15, 802–812. [Google Scholar] [CrossRef] [Green Version]
- Zignego, D.L.; Hilmer, J.K.; June, R.K. Mechanotransduction in primary human osteoarthritic chondrocytes is mediated by metabolism of energy, lipids, and amino acids. J. Biomech. 2015, 48, 4253–4261. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, A.; Kurosawa, S.; Shishido, T.; Pejchal, J.; Yokota, Y.; Futami, Y.; Yoshikawa, A. Fast and High-Energy-Resolution Oxide Scintillator: Ce-Doped (La,Gd) 2Si2O7. Appl. Phys. Express 2012, 5, 102601. [Google Scholar] [CrossRef]
- Malinauskas, M.; Farsari, M.; Piskarskas, A.; Juodkazis, S. Ultrafast laser nanostructuring of photopolymers: A decade of advances. Phys. Rep. 2013, 533, 14452–14459. [Google Scholar] [CrossRef]
- LaFratta, C.N.; Fourkas, J.T.; Baldacchini, T.; Farrer, R.A. Multiphoton fabrication. Angew. Chem. Int. Ed. Engl. 2007, 46, 6238–6258. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leng, N.; Dawson, J.A.; Thomson, J.A.; Ruotti, V.; Rissman, A.I.; Smits, B.M.; Haag, J.D.; Gould, M.N.; Stewart, R.M.; Kendziorski, C. EBSeq: An empirical Bayes hierarchical model for inference in RNA-seq experiments. Bioinformatics 2013, 29, 1035–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrara, M.; Lum, J.; Cordero, F.; Beccuti, M.; Poidinger, M.; Donatelli, S.; Calogero, R.A.; Zolezzi, F. Alternative splicing detection workflow needs a careful combination of sample prep and bioinformatics analysis. BMC Bioinform. 2015, 16 (Suppl. 9), S2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CTR vs. NIC | ||
|---|---|---|
| mRNAs | ncRNAs | |
| Upregulated | 927 | 147 |
| Downregulated | 650 | 210 |
| Total | 1577 | 357 |
| Gene Name | log2FC | Gene Description |
|---|---|---|
| Clcf1 | 7.45 | Protein that in complexation with CRLF1 forms a heterodimeric neurotropic cytokine that plays a crucial role during neuronal development. Stimulates B-cells. Binds to and activates the ILST/gp130 receptor. |
| Ceacam1 | −7.25 | Cell adhesion protein that mediates homophilic cell adhesion in a calcium-independent manner. Plays a role as coinhibitory receptor in immune response and insulin action, and is an activator of angiogenesis. Downregulates cell growth in response to EGF through its interaction with SHC1. Inhibits cell migration and cell scattering through interaction with FLNA, interfering with FLNA’s interaction with RALA. |
| St18 | 7.21 | Repressor that binds to DNA sequences containing a bipartite element consisting of a direct repeat of the sequence 5′-AAAGTTT-3′ separated by 2–9 nucleotides. |
| Slc38a1 | −7.19 | Functions as a sodium-dependent amino acid transporter. May supply glutamatergic and GABAergic neurons with glutamine, which is required for the synthesis of the neurotransmitters, e.g., glutamate and GABA |
| Sema3e | 7.05 | Plays an important role in signaling via the cell surface receptor PLXND1. Mediates reorganization of the actin cytoskeleton, leading to the retraction of cell projections. Promotes focal adhesion disassembly and inhibits adhesion of endothelial cells to the extracellular matrix. |
| Actl7b | 6.35 | Actin-like protein 7B, belongs to the actin family |
| Scrt2 | 6.14 | Transcriptional repressor scratch 2; plays a role in DNA binding and negative regulation of transcription |
| Aldh1a3 | 5.69 | NAD-dependent aldehyde dehydrogenase that catalyzes the formation of retinoic acid |
| Fgl1 | −5.64 | Immune suppressive molecule that inhibits antigen-specific T-cell activation by acting as a major ligand of LAG3 |
| Lhx6 | 5.63 | Probable transcription factor required for the expression of a subset of genes involved in interneurons migration and development. Functions in the specification of cortical interneuron subtypes and in the migration of GABAergic interneuron precursors from the subpallium to the cerebral cortex |
| ncRNAs | |||
|---|---|---|---|
| Upregulated | Downregulated | Total | |
| Antisense | 11 | 16 | 27 |
| Bidirectional promoter lncRNA | 0 | 3 | 3 |
| lincRNA | 20 | 40 | 60 |
| miRNA | 2 | 1 | 3 |
| miscRNA | 1 | 0 | 1 |
| Processed pseudogene | 11 | 14 | 25 |
| Processed transcript | 17 | 28 | 45 |
| rRNA | 1 | 0 | 1 |
| Sense intronic | 9 | 3 | 12 |
| snoRNA | 1 | 31 | 32 |
| snRNA | 3 | 6 | 9 |
| TEC | 67 | 51 | 118 |
| Transcribed unprocessed pseudogene | 3 | 6 | 9 |
| Unitary pseudogene | 0 | 3 | 3 |
| Unprocessed pseudogene | 1 | 8 | 9 |
| 147 | 210 | 357 | |
| Gene Name | log2FC | Gene Description |
|---|---|---|
| Megf6 | 1.88 | Multiple epidermal growth factor-like domains protein 6; multiple EGF-like-domains 6 |
| Adgrb1 | 1.62 | Cell adhesion protein that mediates homophilic cell adhesion in a calcium-independent manner. Plays a role as coinhibitory receptor in immune response and insulin action and is an activator of angiogenesis. Downregulates cell growth in response to EGF through its interaction with SHC1. Inhibits cell migration and cell scattering through interaction with FLNA, interfering with FLNA’s interaction with RALA |
| Unc80 | 1.45 | Protein unc-80 homolog; component of the NALCN sodium channel complex, required for channel regulation. This complex is a cation channel activated by neuropeptides substance P, neurotensin, and extracellular calcium that regulates neuronal excitability by controlling the sizes of NALCN-dependent sodium-leak current. UNC80 is essential for NALCN sensitivity to extracellular calcium; Belongs to the unc-80 family |
| Cad | 1.05 | CAD protein; this protein is a “fusion” protein encoding four enzymatic activities of the pyrimidine pathway (GATase, CPSase, ATCase, and DHOase) |
| Cald1 | −1.11 | Caldesmon 1 |
| Maf | −1.13 | Transcription factor Maf; acts as a transcriptional activator or repressor. When overexpressed, represses antioxidant response element (ARE)-mediated transcription. Involved either as an oncogene or as a tumor suppressor, depending on the cell context. Involved in embryonic lens fiber cell development. Recruits the transcriptional coactivators CREBBP and/or EP300 to crystallin promoters leading to upregulation of crystallin gene during lens fiber cell differentiation |
| Syt7 | −1.28 | Synaptotagmin-7; Ca(2+) sensor involved in Ca(2+)-dependent exocytosis of secretory and synaptic vesicles through Ca(2+) and phospholipid binding to the C2 domain. Ca(2+) induces binding of the C2 domains to phospholipid membranes and to assembled SNARE-complexes; both actions contribute to triggering exocytosis. SYT7 binds Ca(2+) with high affinity and slow kinetics compared to other synaptotagmins |
| Ifi35 | −1.47 | Interferon-induced protein 35 |
| Ccser1 | −1.70 | Serine-rich coiled-coil domain-containing protein 1; coiled-coil serine rich 1 |
| Txlnb | −1.79 | Beta-taxilin; promotes motor nerve regeneration. May be involved in intracellular vesicle traffic |
| Cdk6 | −1.82 | Cyclin-dependent kinase 6; serine/threonine-protein kinase involved in the control of the cell cycle and differentiation; promotes G1/S transition. Phosphorylates pRB/RB1 and NPM1. Interacts with D-type G1 cyclins during interphase at G1 to form a pRB/RB1 kinase and controls the entrance into the cell cycle. Involved in initiation and maintenance of cell cycle exit during cell differentiation; prevents cell proliferation and regulates negatively cell differentiation, but is required for the proliferation of specific cell types (e.g., erythroid and hematopoietic cells) |
| Gbp9 | −1.95 | Guanylate-binding protein 9 |
| Cp | −2.00 | Ceruloplasmin; ceruloplasmin is a blue, copper-binding (6–7 atoms per molecule) glycoprotein. It has ferroxidase activity oxidizing Fe(2+) to Fe(3+) without releasing radical oxygen species. It is involved in iron transport across the cell membrane. Provides Cu(2+) ions for the ascorbate-mediated deaminase degradation of the heparan sulfate chains of GPC1. May also play a role in fetal lung development or pulmonary antioxidant defense |
| S100b | −2.16 | Protein S100-B; weakly binds calcium but binds zinc very tightly—distinct binding sites with different affinities exist for both ions on each monomer. Physiological concentrations of potassium ion antagonize the binding of both divalent cations, especially affecting high-affinity calcium-binding sites. Binds to and initiates the activation of STK38 by releasing autoinhibitory intramolecular interactions within the kinase. Interaction with AGER after myocardial infarction may play a role in myocyte apoptosis by activating ERK1/2 and p53/TP53 signaling |
| Serpinb8 | −2.25 | Serpin B8; has an important role in epithelial desmosome-mediated cell–cell adhesion |
| Il33 | −2.29 | Interleukin-33; cytokine that binds to and signals through the IL1RL1/ST2 receptor, which in turn activates NF- κ -B and MAPK signaling pathways in target cells. Involved in the maturation of Th2 cells inducing the secretion of T-helper type 2-associated cytokines. It is also involved in activation of mast cells, basophils, eosinophils, and natural killer cells. Acts as a chemoattractant for Th2 cells, and may function as an “alarmin,” which amplifies immune responses during tissue injury |
| Thrsp | −2.50 | Thyroid hormone-inducible hepatic protein; plays a role in the regulation of lipogenesis, especially in lactating mammary gland. Important for the biosynthesis of triglycerides with medium-length fatty acid chains. May modulate lipogenesis by interacting with MID1IP1 and preventing its interaction with ACACA. May function as transcriptional coactivator |
| Cmbl | −2.53 | Carboxymethylenebutenolidase homolog; cysteine hydrolase |
| Nupr1 | −2.60 | Nuclear protein 1; chromatin-binding protein that converts stress signals into a program of gene expression that empowers cells with resistance to the stress induced by a change in their microenvironment. Interacts with MSL1 and inhibits its activity on histone H4 ‘Lys-16′ acetylation (H4K16ac). Binds the RELB promoter and activates its transcription, leading to the transactivation of IER3. |
| Thrb | −2.81 | Thyroid hormone receptor beta; nuclear hormone receptor that can act as a repressor or activator of transcription. High-affinity receptor for thyroid hormones, including triiodothyronine and thyroxine |
| Olfr287 | −3.00 | Olfactory receptor 287 |
| Srl | −3.16 | Sarcalumenin; may be involved in the regulation of calcium transport |
| Scara5 | −3.27 | Scavenger receptor class A member 5; ferritin receptor that mediates nontransferrin-dependent delivery of iron. Mediates cellular uptake of ferritin-bound iron by stimulating ferritin endocytosis from the cell surface with consequent iron delivery within the cell. Delivery of iron to cells by ferritin is required for the development of specific cell types, suggesting the existence of cell type-specific mechanisms of iron traffic in organogenesis, which alternatively utilize transferrin or nontransferrin iron delivery pathways |
| Ip6k3 | −3.33 | Inositol hexakisphosphate kinase 3; converts inositol hexakisphosphate (InsP6) to diphosphoinositol pentakisphosphate (InsP7/PP-InsP5). Converts 1,3,4,5,6-pentakisphosphate (InsP5) to PP-InsP4 (by similarity) |
| Pygm | −3.45 | Glycogen phosphorylase, muscle form; phosphorylase is an important allosteric enzyme in carbohydrate metabolism |
| Otoa | −3.55 | Otoancorin; may act as an adhesion molecule; belongs to the stereocilin family |
| Bace2 | −3.72 | Beta-secretase 2; responsible for the proteolytic processing of the amyloid precursor protein (APP). Cleaves APP, between residues 690 and 691, leading to the generation and extracellular release of beta-cleaved soluble APP, and a corresponding cell-associated C-terminal fragment, which is later released by gamma-secretase |
| Myh11 | −3.78 | Myosin-11; muscle contraction; belongs to the TRAFAC class myosin-kinesin ATPase superfamily. Myosin family |
| Grin2a | −4.45 | Glutamate receptor ionotropic, NMDA 2A; component of NMDA receptor complexes that function as heterotetrameric, ligand-gated ion channels with high calcium permeability and voltage-dependent sensitivity to magnesium. Channel activation requires binding of the neurotransmitter glutamate to the epsilon subunit, glycine binding to the zeta subunit, plus membrane depolarization to eliminate channel inhibition by Mg(2+) |
| Ccdc187 | −4.53 | Coiled-coil domain-containing protein 187; RIKEN cDNA 4932418E24 gene |
| Wif1 | −5.10 | Wnt inhibitory factor 1; binds to WNT proteins and inhibits their activities. May be involved in mesoderm segmentation |
| Nnmt | −5.15 | Nicotinamide N-methyltransferase; catalyzes the N-methylation of nicotinamide and other pyridines to form pyridinium ions. This activity is important for biotransformation of many drugs and xenobiotic compounds; belongs to the class I-like SAM-binding methyltransferase superfamily. NNMT/PNMT/TEMT family |
| Gm14406 | −6.56 | Predicted gene 14406 |
| Defb41 | −8.64 | Beta-defensin 41; has bactericidal activity; belongs to the beta-defensin family |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rey, F.; Pandini, C.; Barzaghini, B.; Messa, L.; Giallongo, T.; Pansarasa, O.; Gagliardi, S.; Brilli, M.; Zuccotti, G.V.; Cereda, C.; et al. Dissecting the Effect of a 3D Microscaffold on the Transcriptome of Neural Stem Cells with Computational Approaches: A Focus on Mechanotransduction. Int. J. Mol. Sci. 2020, 21, 6775. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186775
Rey F, Pandini C, Barzaghini B, Messa L, Giallongo T, Pansarasa O, Gagliardi S, Brilli M, Zuccotti GV, Cereda C, et al. Dissecting the Effect of a 3D Microscaffold on the Transcriptome of Neural Stem Cells with Computational Approaches: A Focus on Mechanotransduction. International Journal of Molecular Sciences. 2020; 21(18):6775. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186775
Chicago/Turabian StyleRey, Federica, Cecilia Pandini, Bianca Barzaghini, Letizia Messa, Toniella Giallongo, Orietta Pansarasa, Stella Gagliardi, Matteo Brilli, Gian Vincenzo Zuccotti, Cristina Cereda, and et al. 2020. "Dissecting the Effect of a 3D Microscaffold on the Transcriptome of Neural Stem Cells with Computational Approaches: A Focus on Mechanotransduction" International Journal of Molecular Sciences 21, no. 18: 6775. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186775