Functional and Pharmacological Comparison of Human, Mouse, and Rat Organic Cation Transporter 1 toward Drug and Pesticide Interaction

Abstract
:1. Introduction
2. Results
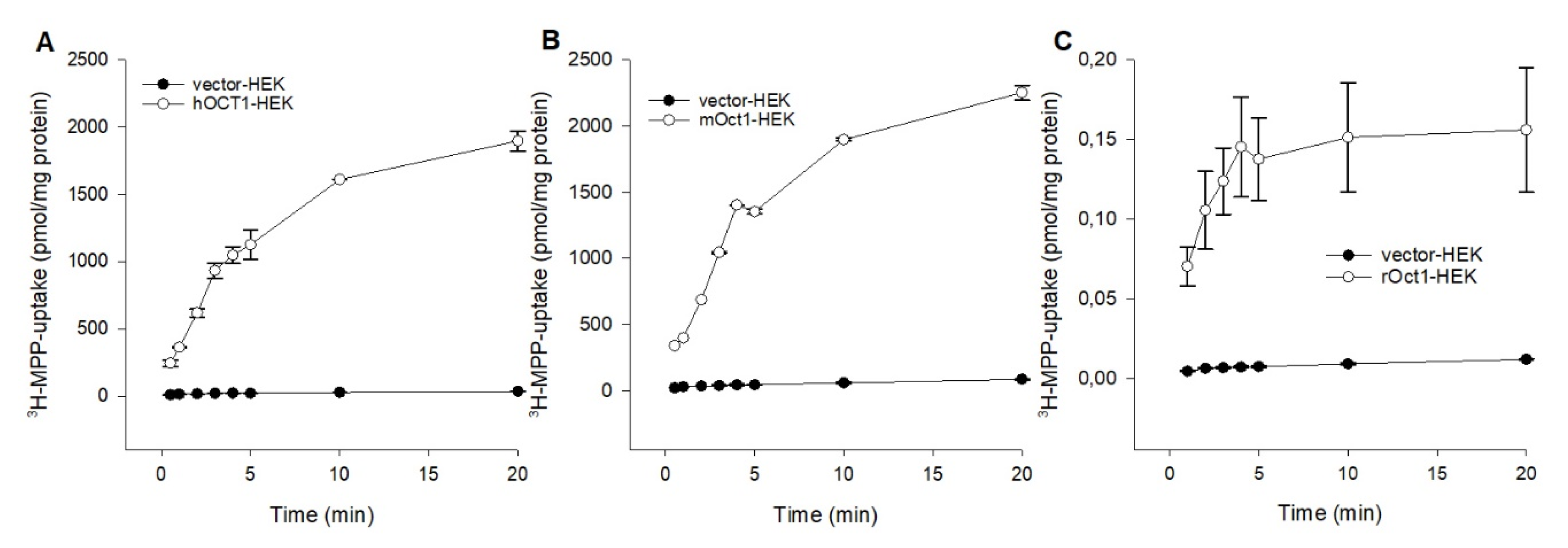
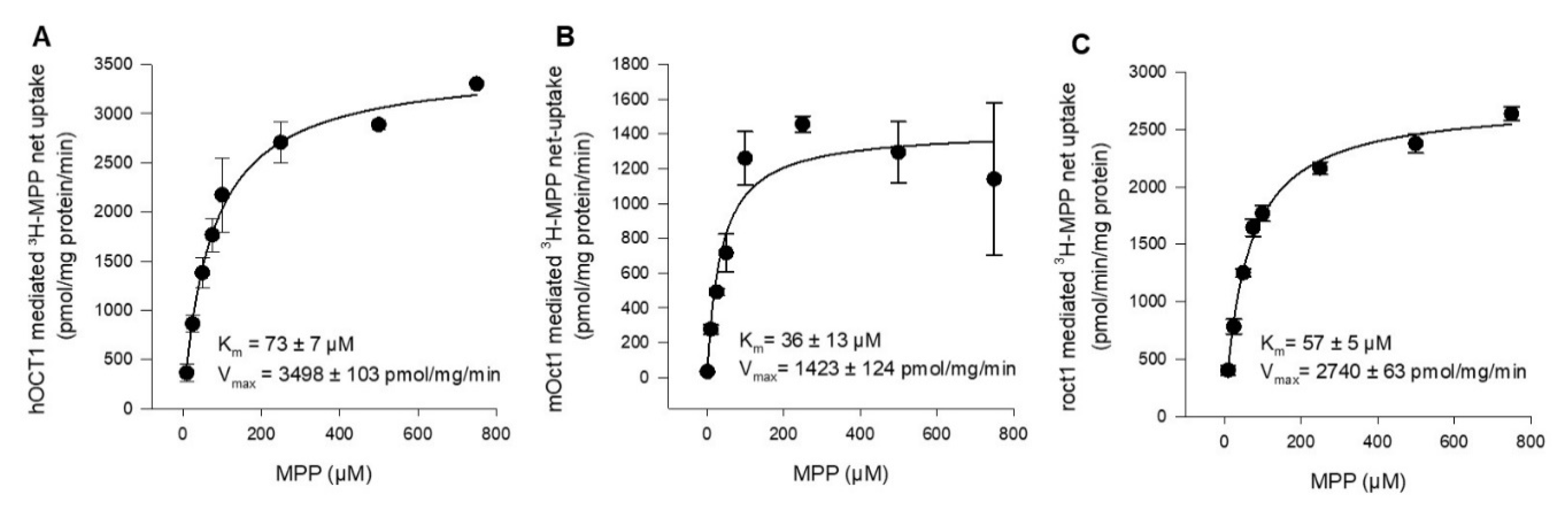
2.1. Functional Characterization of Human, Mouse, and Rat OCT1
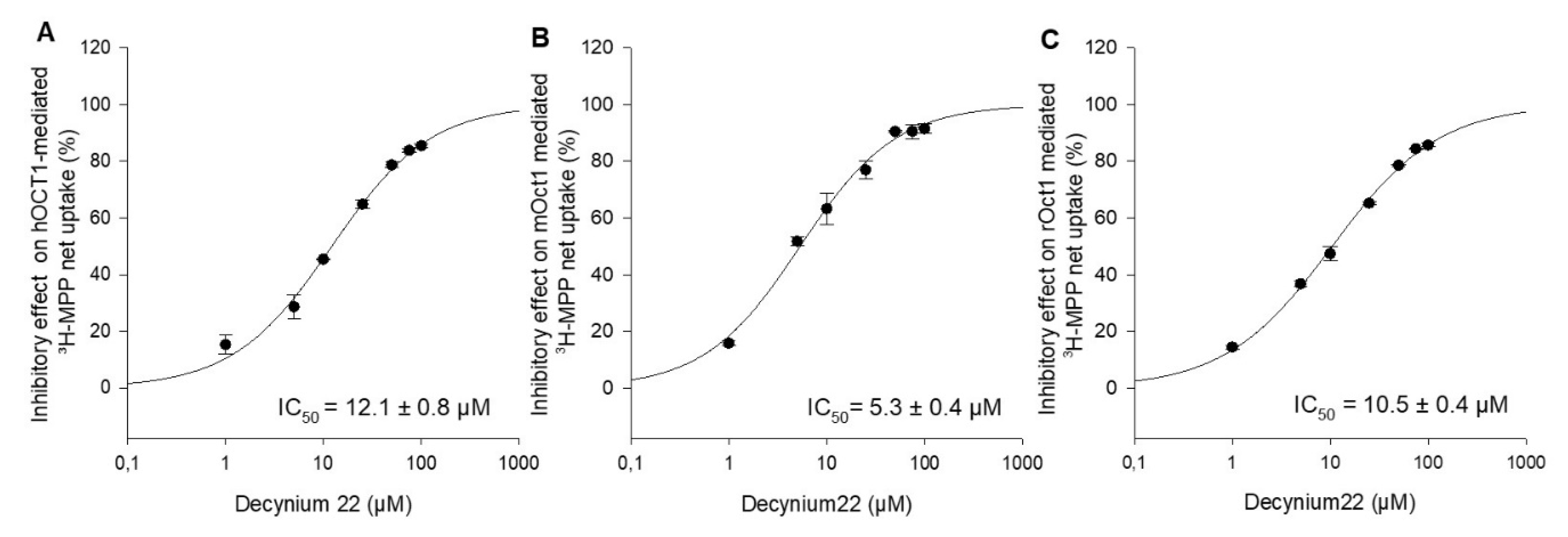
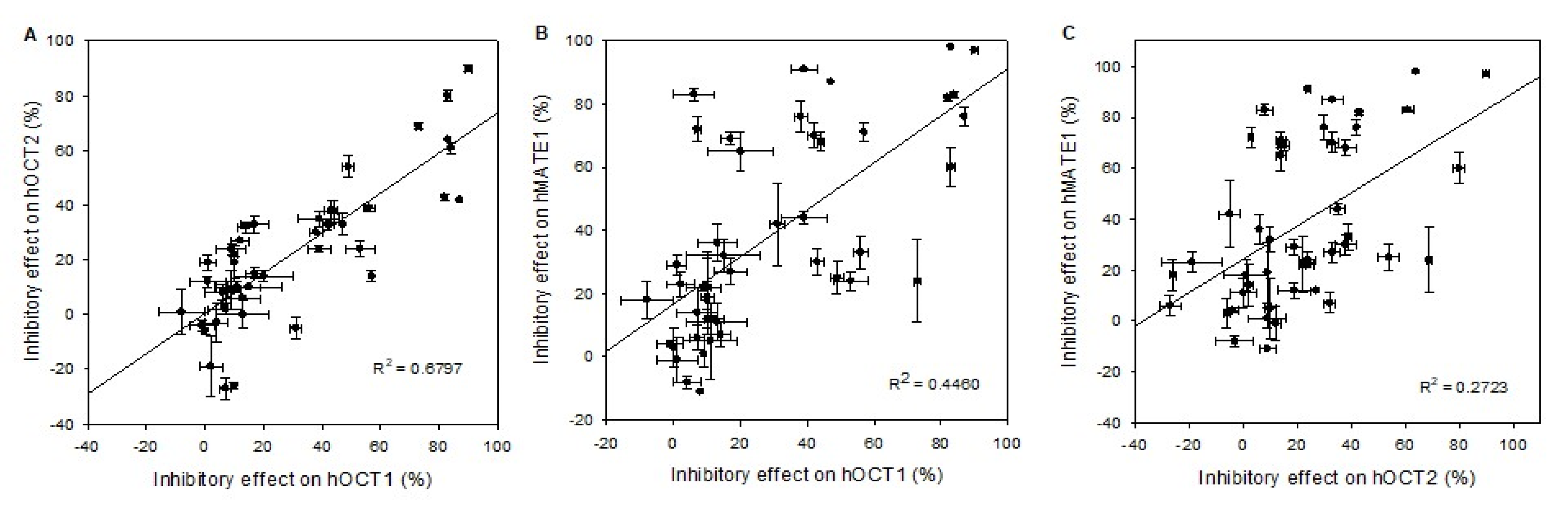
2.2. Comparison of the Interaction of hOCT1, mOct1, and rOct1 with Drugs and Pesticides
3. Discussion
4. Materials and Methods
4.1. Material
4.2. Transfection and Cell Culture
4.3. Transporter Mediated Uptake of Radiolabeled Substrates
4.4. Inhibition Experiments
4.5. Determination of Protein Concentration
4.6. Data Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gründemann, D.; Gorboulev, V.; Gambaryan, S.; Veyhl, M.; Koepsell, H. Drug excretion mediated by a new prototype of polyspecific transporter. Nature 1994, 372, 549–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Dresser, M.J.; Gray, A.T.; Yost, S.C.; Terashita, S.; Giacomini, K.M. Cloning and functional expression of a human liver organic cation transporter. Mol. Pharmacol. 1997, 51, 913–921. [Google Scholar] [CrossRef] [PubMed]
- Koepsell, H.; Schmitt, B.M.; Gorboulev, V. Organic cation transporters. Rev. Physiol. Biochem. Pharmacol. 2003, 150, 36–90. [Google Scholar] [CrossRef] [PubMed]
- Gorboulev, V.; Ulzheimer, J.C.; Akhoundova, A.; Ulzheimer-Teuber, I.; Karbach, U.; Quester, S.; Baumann, C.; Lang, F.; Busch, A.E.; Koepsell, H. Cloning and characterization of two human polyspecific organic cation transporters. DNA Cell Biol. 1997, 16, 871–881. [Google Scholar] [CrossRef] [PubMed]
- Nies, A.T.; Koepsell, H.; Winter, S.; Burk, O.; Klein, K.; Kerb, R.; Zanger, U.M.; Keppler, D.; Schwab, M.; Schaeffeler, E. Expression of organic cation transporters OCT1 (SLC22A1) and OCT3 (SLC22A3) is affected by genetic factors and cholestasis in human liver. Hepatol. 2009, 50, 1227–1240. [Google Scholar] [CrossRef] [PubMed]
- Koepsell, H.; Lips, K.; Volk, C. Polyspecific organic cation transporters: Structure, function, physiological roles, and biopharmaceutical implications. Pharm.Res. 2007, 24, 1227–1251. [Google Scholar] [CrossRef] [PubMed]
- Han, T.K.; Everett, R.S.; Proctor, W.R.; Ng, C.M.; Costales, C.L.; Brouwer, K.L.R.; Thakker, D.R. Organic cation transporter 1 (OCT1/mOct1) is localized in the apical membrane of Caco-2 cell monolayers and enterocytes. Mol. Pharmacol. 2013, 84, 182–189. [Google Scholar] [CrossRef] [Green Version]
- Koepsell, H. Organic Cation Transporters in Health and Disease. Pharmacol. Rev. 2020, 72, 253–319. [Google Scholar] [CrossRef]
- Tzvetkov, M.V.; dos Santos Pereira, J.N.; Meineke, I.; Saadatmand, A.R.; Stingl, J.C.; Brockmöller, J. Morphine is a substrate of the organic cation transporter OCT1 and polymorphisms in OCT1 gene affect morphine pharmacokinetics after codeine administration. Biochem. Pharmacol. 2013, 86, 666–678. [Google Scholar] [CrossRef] [Green Version]
- Koepsell, H. The SLC22 family with transporters of organic cations, anions and zwitterions. Mol. Asp. Med. 2013, 34, 413–435. [Google Scholar] [CrossRef]
- Matthaei, J.; Kuron, D.; Faltraco, F.; Knoch, T.; Dos Santos Pereira, J.N.; Abu Abed, M.; Prukop, T.; Brockmöller, J.; Tzvetkov, M.V. OCT1 mediates hepatic uptake of sumatriptan and loss-of-function OCT1 polymorphisms affect sumatriptan pharmacokinetics. Clin. Pharmacol. Ther. 2016, 99, 633–641. [Google Scholar] [CrossRef] [PubMed]
- Tu, M.; Sun, S.; Wang, K.; Peng, X.; Wang, R.; Li, L.; Zeng, S.; Zhou, H.; Jiang, H. Organic cation transporter 1 mediates the uptake of monocrotaline and plays an important role in its hepatotoxicity. Toxicology 2013, 311, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Giacomini, K.M.; Huang, S.M.; Tweedie, D.J.; Benet, L.Z.; Brouwer, K.L.; Chu, X.; Dahlin, A.; Evers, R.; Fischer, V.; Hillgren, K.M.; et al. Membrane transporters in drug development. Nat. Rev. Drug Discov. 2010, 9, 215–236. [Google Scholar] [PubMed]
- Lauer, B.; Tuschl, G.; Kling, M.; Mueller, S.O. Species-specific toxicity of diclofenac and troglitazone in primary human and rat hepatocytes. Chem. Biol. Interact. 2009, 179, 17–24. [Google Scholar] [CrossRef]
- Tuschl, G.; Lauer, B.; Mueller, S.O. Primary hepatocytes as a model to analyze species-specific toxicity and drug metabolism. Expert Opin. Drug Metab. Toxicol. 2008, 4, 855–870. [Google Scholar] [CrossRef]
- Lai, Y. Identification of interspecies difference in hepatobiliary transporters to improve extrapolation of human biliary secretion. Expert Opin. Drug Metab. Toxicol. 2009, 5, 1175–1187. [Google Scholar] [CrossRef]
- Dresser, M.J.; Gray, A.T.; Giacomini, K.M. Kinetic and selectivity differences between rodent, rabbit, and human organic cation transporters (OCT1). J. Pharmacol. Exp. Ther. 2000, 292, 1146–1152. [Google Scholar]
- Okuda, M.; Urakami, Y.; Saito, H.; Inui, K. Molecular mechanisms of organic cation transport in OCT2-expressing Xenopus oocytes. Biochim. Biophys. Acta 1999, 1417, 224–231. [Google Scholar] [CrossRef] [Green Version]
- Vermeer, L.M.M.; Isringhausen, C.D.; Ogilvie, B.W.; Buckley, D.B. Evaluation of Ketoconazole and Its Alternative Clinical CYP3A4/5 Inhibitors as Inhibitors of Drug Transporters: The In Vitro Effects of Ketoconazole, Ritonavir, Clarithromycin, and Itraconazole on 13 Clinically-Relevant Drug Transporters. Drug Metab. Dispos. 2016, 44, 453–459. [Google Scholar] [CrossRef] [Green Version]
- Panfen, E.; Chen, W.; Zhang, Y.; Sinz, M.; Marathe, P.; Gan, J.; Shen, H. Enhanced and Persistent Inhibition of Organic Cation Transporter 1 Activity by Preincubation of Cyclosporine A. Drug Metab. Dispos. 2019, 47, 1352–1360. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, S.; Sorani, M.; Giacomini, K.M. Transport of paraquat by human organic cation transporters and multidrug and toxic compound extrusion family. J. Pharmacol. Exp. Ther. 2007, 322, 695–700. [Google Scholar] [CrossRef]
- Meyer, M.J.; Seitz, T.; Brockmöller, J.; Tzvetkov, M.V. Effects of genetic polymorphisms on the OCT1 and OCT2-mediated uptake of ranitidine. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gründemann, D.; Hahne, C.; Berkels, R.; Schömig, E. Agmatine is efficiently transported by non-neuronal monoamine transporters extraneuronal monoamine transporter (EMT) and organic cation transporter 2 (OCT2). J. Pharmacol. Exp. Ther. 2003, 304, 810–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakehi, M.; Koyabu, N.; Nakamura, T.; Uchiumi, T.; Kuwano, M.; Ohtani, H.; Sawada, Y. Functional characterization of mouse cation transporter mOCT2 compared with mOCT1. Biochem. Biophys. Res. Commun. 2002, 296, 644–650. [Google Scholar] [CrossRef]
- Gorboulev, V.; Shatskaya, N.; Volk, C.; Koepsell, H. Subtype-specific affinity for corticosterone of rat organic cation transporters rOCT1 and rOCT2 depends on three amino acids within the substrate binding region. Mol. Pharmacol. 2005, 67, 1612–1619. [Google Scholar] [CrossRef] [Green Version]
- Martel, F.; Martins, M.J.; Azevedo, I. Inward transport of 3H-MPP+ in freshly isolated rat hepatocytes: Evidence for interaction with catecholamines. Naunyn Schmiedebergs Arch. Pharmacol. 1996, 354, 305–311. [Google Scholar] [CrossRef]
- Wright, S.H.; Dantzler, W.H. Molecular and cellular physiology of renal organic cation and anion transport. Physiol. Rev. 2004, 84, 987–1049. [Google Scholar] [CrossRef]
- Gorboulev, V.; Volk, C.; Arndt, P.; Akhoundova, A.; Koepsell, H. Selectivity of the polyspecific cation transporter rOCT1 is changed by mutation of aspartate 475 to glutamate. Mol. Pharmacol. 1999, 56, 1254–1261. [Google Scholar] [CrossRef]
- Seitz, T.; Stalmann, R.; Dalila, N.; Chen, J.; Pojar, S.; Dos Santos Pereira, J.N.; Krätzner, R.; Brockmöller, J.; Tzvetkov, M.V. Global genetic analyses reveal strong inter-ethnic variability in the loss of activity of the organic cation transporter OCT1. Genome Med. 2015, 7. [Google Scholar] [CrossRef] [Green Version]
- Tsiplakou, E.; Anagnostopoulos, C.J.; Liapis, K.; Haroutounian, S.A.; Zervas, G. Pesticides residues in milks and feedstuff of farm animals drawn from Greece. Chemosphere 2010, 80, 504–512. [Google Scholar] [CrossRef]
- Pirsaheb, M.; Limoee, M.; Namdari, F.; Khamutian, R. Organochlorine pesticides residue in breast milk: A systematic review. Med. J. Islam. Repub. Iran. 2015, 29, 228. [Google Scholar] [PubMed]
- Hamilton, D.; Ambrus, A.; Dieterle, R.; Felsot, A.; Harris, C.; Petersen, B.; Racke, K.; Wong, S.-S.; Gonzalez, R.; Tanaka, K.; et al. Pesticide residues in food--acute dietary exposure. Pest Manag. Sci. 2004, 60, 311–339. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D.M.; Buckley, N.A.; Mohamed, F.; Eddleston, M.; Goldstein, D.A.; Mehrsheikh, A.; Bleeke, M.S.; Dawson, A.H. A prospective observational study of the clinical toxicology of glyphosate-containing herbicides in adults with acute self-poisoning. Clin. Toxicol. (Phila.) 2010, 48, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Niemann, L.; Sieke, C.; Pfeil, R.; Solecki, R. A critical review of glyphosate findings in human urine samples and comparison with the exposure of operators and consumers. J. Verbr. Lebensm. 2015, 10, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Acquavella, J.F.; Alexander, B.H.; Mandel, J.S.; Gustin, C.; Baker, B.; Chapman, P.; Bleeke, M. Glyphosate biomonitoring for farmers and their families: Results from the Farm Family Exposure Study. Environ. Health Perspect. 2004, 112, 321–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, B.S.; Lazzaro, V.A.; Seale, J.P.; Duggin, G.G. The renal excretory mechanisms and the role of organic cations in modulating the renal handling of paraquat. Pharmacol. Ther. 1998, 79, 193–203. [Google Scholar] [CrossRef]
- Faniband, M.H.; Littorin, M.; Ekman, E.; Jönsson, B.A.G.; Lindh, C.H. LC-MS-MS Analysis of Urinary Biomarkers of Imazalil Following Experimental Exposures. J. Anal. Toxicol. 2015, 39, 691–697. [Google Scholar] [CrossRef]
- Jamin, E.L.; Bonvallot, N.; Tremblay-Franco, M.; Cravedi, J.-P.; Chevrier, C.; Cordier, S.; Debrauwer, L. Untargeted profiling of pesticide metabolites by LC-HRMS: An exposomics tool for human exposure evaluation. Anal. Bioanal. Chem. 2014, 406, 1149–1161. [Google Scholar] [CrossRef]
- Kuklenyik, Z.; Panuwet, P.; Jayatilaka, N.K.; Pirkle, J.L.; Calafat, A.M. Two-dimensional high performance liquid chromatography separation and tandem mass spectrometry detection of atrazine and its metabolic and hydrolysis products in urine. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2012, 901, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Tan, Y.; Guo, H. Simultaneous determination of amitraz, chlordimeform, formetanate and their main metabolites in human urine by high performance liquid chromatography-tandem mass spectrometry. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2017, 1052, 27–33. [Google Scholar] [CrossRef]
- Tao, Y.; Dong, F.; Xu, J.; Phung, D.; Liu, Q.; Li, R.; Liu, X.; Wu, X.; He, M.; Zheng, Y. Characteristics of neonicotinoid imidacloprid in urine following exposure of humans to orchards in China. Environ. Int. 2019, 132, 105079. [Google Scholar] [CrossRef] [PubMed]
- Needham, D.; Challis, I.R. The metabolism and excretion of prochloraz, an imidazole-based fungicide, in the rat. Xenobiotica 1991, 21, 1473–1482. [Google Scholar] [CrossRef] [PubMed]
- Halwachs, S.; Schäfer, I.; Kneuer, C.; Seibel, P.; Honscha, W. Assessment of ABCG2-mediated transport of pesticides across the rabbit placenta barrier using a novel MDCKII in vitro model. Toxicol. Appl. Pharmacol. 2016, 305, 66–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guéniche, N.; Bruyere, A.; Ringeval, M.; Jouan, E.; Huguet, A.; Hégarat, L.L.; Fardel, O. Differential interactions of carbamate pesticides with drug transporters. Xenobiotica 2020, 1–13. [Google Scholar] [CrossRef]
- Hagos, Y.; Hundertmark, P.; Shnitsar, V.; Marada, V.V.; Wulf, G.; Burckhardt, G. Renal human organic anion transporter 3 increases the susceptibility of lymphoma cells to bendamustine uptake. Am. J. Physiol. Renal. Physiol. 2015, 308, F330–F338. [Google Scholar] [CrossRef] [Green Version]
- Marada, V.V.; Florl, S.; Kuhne, A.; Burckhardt, G.; Hagos, Y. Interaction of human organic anion transporter polypeptides 1B1 and 1B3 with antineoplastic compounds. Eur. J. Med. Chem. 2015, 92, 723–731. [Google Scholar] [CrossRef]
- Hagos, Y.; Stein, D.; Ugele, B.; Burckhardt, G.; Bahn, A. Human Renal Organic Anion Transporter 4 Operates as an Asymmetric Urate Transporter. J. Am. Soc. Nephrol. 2007, 18, 430–439. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drugs (15) | Type of Drug | Charge at pH 7.4 | Inhibitory Effects (%) | |||||
|---|---|---|---|---|---|---|---|---|
| hOCT1 | mOct1 | rOct1 | ||||||
| 10 µM | 100 µM | 10 µM | 100 µM | 10 µM | 100 µM | |||
| Ketoconazol * | antifungal | 82% uncharged 18% cation | 47 | 83 | 46 | 74 | 43 | 75 |
| Clonidine | hypertension | 100% cation | 73 | 83 | 72 | 82 | 62 | 80 |
| Verapamil * | class IV antiarrhythmic agent | 100% cation | 39 | 82 | 29 | 64 | 29 | 64 |
| Quinine * | anti malaria | 100% cation | 20 | 59 | −7 | 50 | −1 | 44 |
| Elacridar | tumor drug resistance | 100% cation | 31 | 57 | 61 | 76 | 53 | 70 |
| Quinidine * | class I antiarrhythmic agent | 100% cation | 13 | 53 | −5 | 46 | 19 | 37 |
| Procainamide * | class I antiarrhythmic agent | 100% cation | 10 | 43 | −5 | 58 | 21 | 58 |
| Ritonavir * | antiretroviral HIV | 100% cation | 17 | 39 | 8 | −16 | 17 | 22 |
| Ranitidine * | H2 histamine receptor antagonist | 100% cation | 10 | 38 | −12 | 12 | 11 | 45 |
| Zosuquidar | antineoplastic drug | 37% uncharged 63% cation | 2 | 20 | −4 | −23 | 0 | 4 |
| Metformin * | type 2 diabetes | 100% cation | 11 | 13 | 12 | 9 | −19 | −4 |
| Amiodarone * | class III antiarrhythmic agent | 100% cation | 9 | 10 | −29 | 9 | −25 | 1 |
| Cimetidine * | H2 histamine receptor antagonist | 75% uncharged 25% cation | 8 | 9 | −25 | 21 | 19 | 20 |
| CyclosporinA * | immunsuppressant | 100% cation | 10 | 7 | −27 | 6 | −15 | 8 |
| Reserpine | hypertension | 70% uncharged 30% cation | 7 | 6 | −12 | 2 | −17 | 12 |
| Pesticides (n = 9) | Type of Pesticide | Charge at pH 7.4 | Inhibitory Effects (%) | |||||
|---|---|---|---|---|---|---|---|---|
| hOCT1 | mOct1 | rOct1 | ||||||
| 10 µM | 100 µM | 10 µM | 100 µM | 10 µM | 100 µM | |||
| Imazalil | fungicide | 81% uncharged 19% cation | 56 | 84 | 84 | 97 | 68 | 92 |
| Propamocarb | fungicide | 100% uncharged | 9 | 49 | 29 | 63 | 24 | 57 |
| Azoxystrobin | fungicide | 100% uncharged | 17 | 44 | −2 | 49 | 0 | 42 |
| Prochloraz | fungicide | 100% cation | 14 | 42 | 29 | 28 | 25 | 33 |
| Atrazin | herbicide | 100% uncharged | −8 | 15 | −1 | 1 | −65 | −33 |
| Amitraz | insecticide | 100% cation | 8 | 12 | −23 | 21 | −30 | 10 |
| Glyphosat | herbicide | 73% anion 27% ± charge | 1 | 7 | −9 | 22 | −52 | −19 |
| Imidacloprid | insecticide | 100% ± charge | 4 | 1 | 23 | −9 | −5 | −36 |
| Paraquat * | herbicide | 100% cation | 0 | −1 | 17 | −9 | 1 | −35 |
| Drugs (14) | Inhibitory Effects (%) | |||||
|---|---|---|---|---|---|---|
| hOCT1 | hOCT2 | hMATE1 | ||||
| 10 µM | 100 µM | 10 µM | 100 µM | 10 µM | 100 µM | |
| Decynium22 * | 87 | 90 | 42 | 90 | 76 | 97 |
| Clonidine | 73 | 83 | 69 | 80 | 24 | 60 |
| Ketoconazol * | 47 | 83 | 33 | 64 | 87 | 98 |
| Verapamil * | 39 | 82 | 35 | 43 | 44 | 82 |
| Elacridar | 31 | 57 | −5 | 14 | 42 | 71 |
| Quinidine * | 13 | 53 | 0 | 24 | 11 | 24 |
| Procainamide * | 10 | 43 | 22 | 38 | 22 | 30 |
| Ritonavir * | 17 | 39 | 15 | 24 | 69 | 91 |
| Ranitidine * | 10 | 38 | 9 | 30 | 19 | 76 |
| Zosuquidar | 2 | 20 | −19 | 14 | 23 | 65 |
| Metformin * | 11 | 13 | 10 | 6 | 5 | 36 |
| Amiodarone * | 9 | 10 | 9 | 19 | 1 | 12 |
| CyclosporinA | 10 | 7 | −26 | −27 | 18 | 6 |
| Reserpine | 7 | 6 | 3 | 8 | 72 | 83 |
| Pesticides (9) | Inhibitory Effects (%) | |||||
|---|---|---|---|---|---|---|
| OCT1 | OCT2 | MATE1 | ||||
| 10 µM | 100 µM | 10 µM | 100 µM | 10 µM | 100 µM | |
| Imazalil | 56 | 84 | 39 | 61 | 33 | 83 |
| Propamocarb | 9 | 49 | 24 | 54 | 22 | 25 |
| Azoxystrobin | 17 | 44 | 33 | 38 | 27 | 68 |
| Prochloraz | 14 | 42 | 32 | 33 | 7 | 70 |
| Atrazin | −8 | 15 | 1 | 10 | 18 | 32 |
| Amitraz | 8 | 12 | 9 | 27 | −11 | 12 |
| Glyphosat | 1 | 7 | 12 | 2 | −1 | 14 |
| Imidacloprid | 4 | 1 | −3 | 19 | −8 | 29 |
| Paraquat * | 0 | −1 | −6 | −4 | 3 | 4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Floerl, S.; Kuehne, A.; Hagos, Y. Functional and Pharmacological Comparison of Human, Mouse, and Rat Organic Cation Transporter 1 toward Drug and Pesticide Interaction. Int. J. Mol. Sci. 2020, 21, 6871. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186871
Floerl S, Kuehne A, Hagos Y. Functional and Pharmacological Comparison of Human, Mouse, and Rat Organic Cation Transporter 1 toward Drug and Pesticide Interaction. International Journal of Molecular Sciences. 2020; 21(18):6871. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186871
Chicago/Turabian StyleFloerl, Saskia, Annett Kuehne, and Yohannes Hagos. 2020. "Functional and Pharmacological Comparison of Human, Mouse, and Rat Organic Cation Transporter 1 toward Drug and Pesticide Interaction" International Journal of Molecular Sciences 21, no. 18: 6871. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186871