Semaphorin4A-Plexin D1 Axis Induces Th2 and Th17 While Represses Th1 Skewing in an Autocrine Manner

 , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Sema4A Is Induced During CD4+ T Cell Differentiation
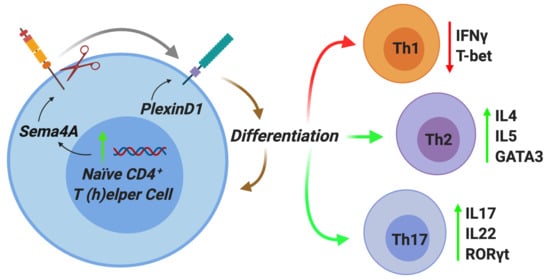
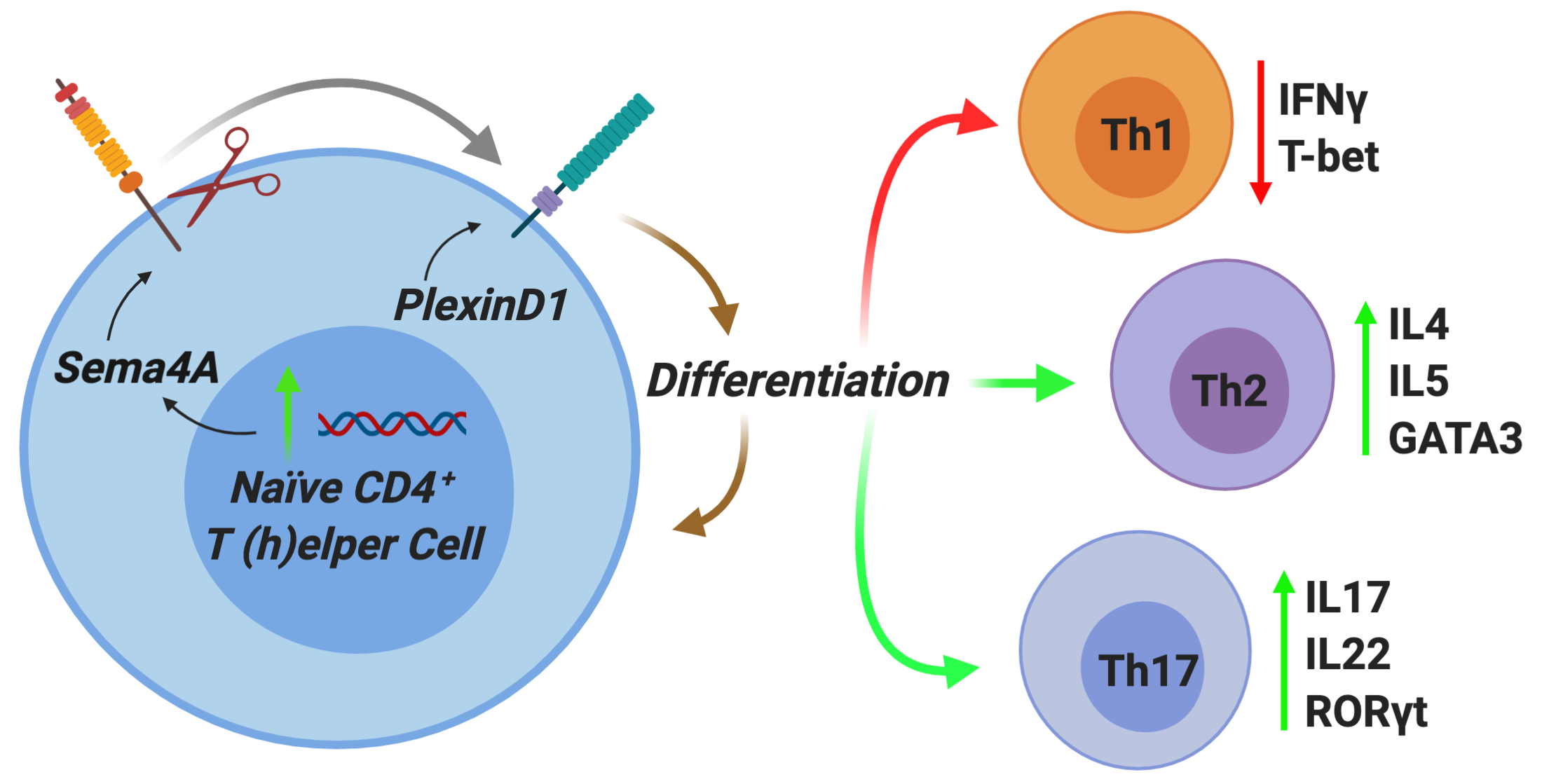
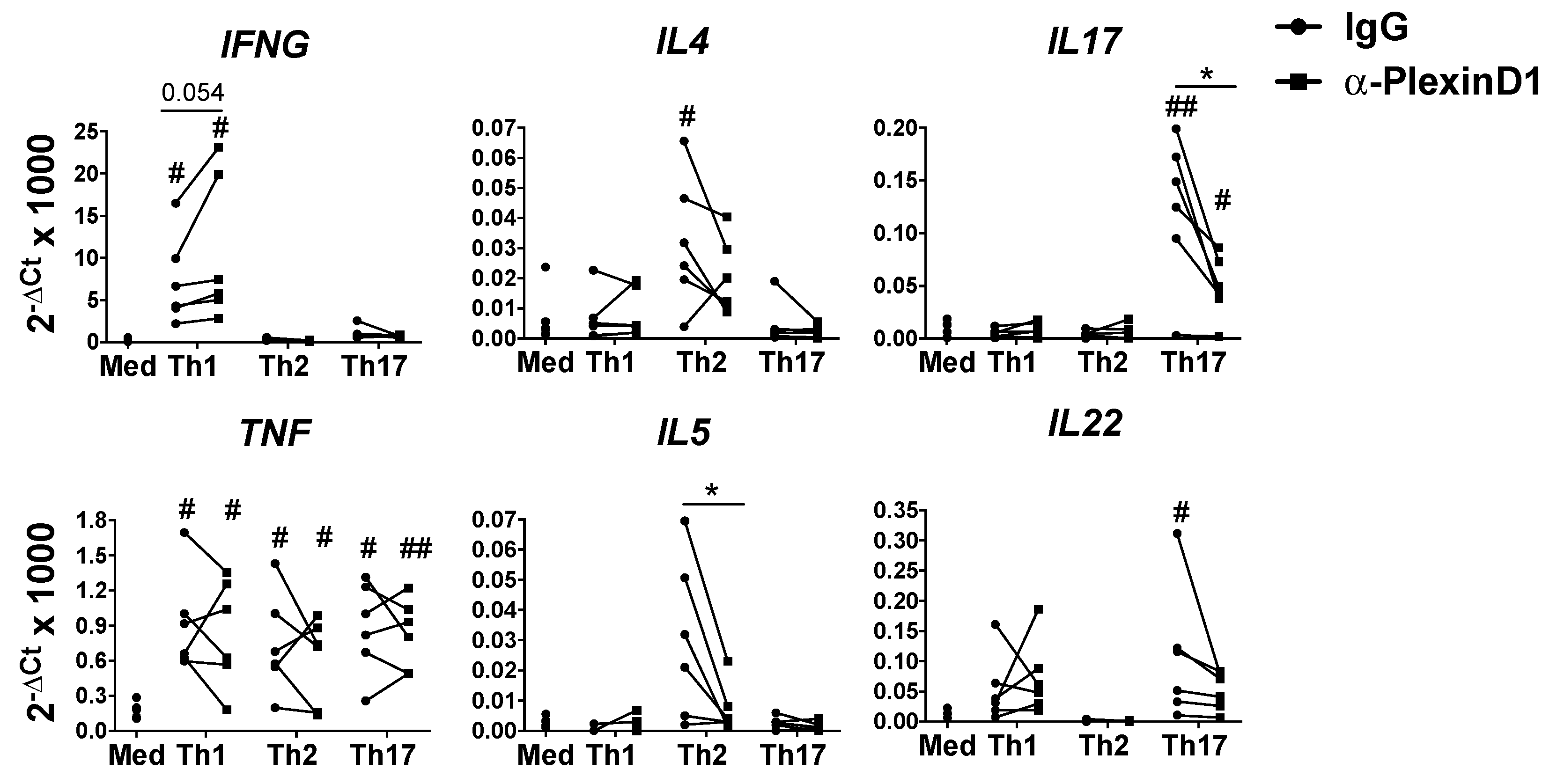
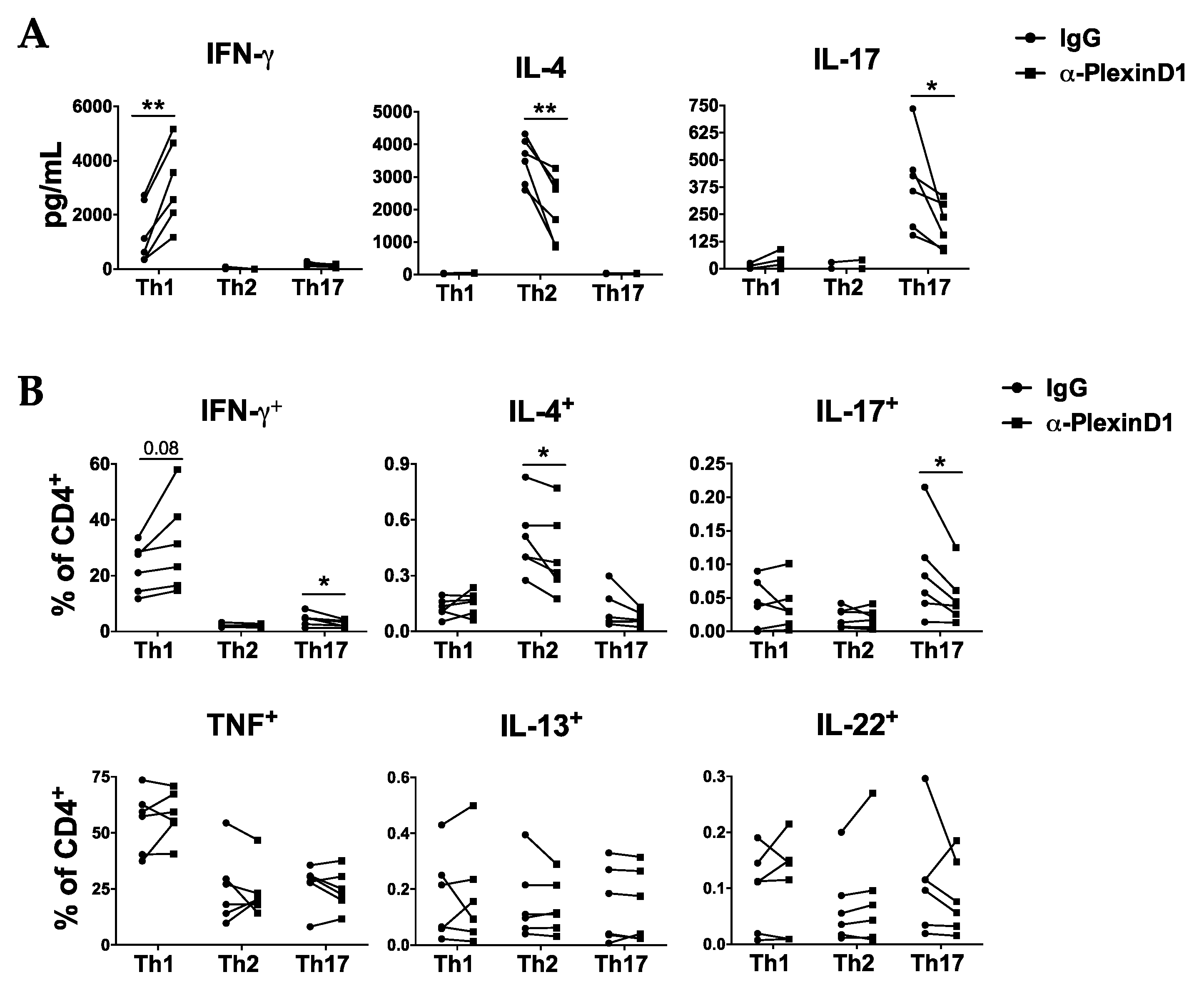
2.2. Autocrine Sema4A–plexinD1 Signaling Induces Th2 and Th17 Differentiation and Suppresses Th1 Skewing
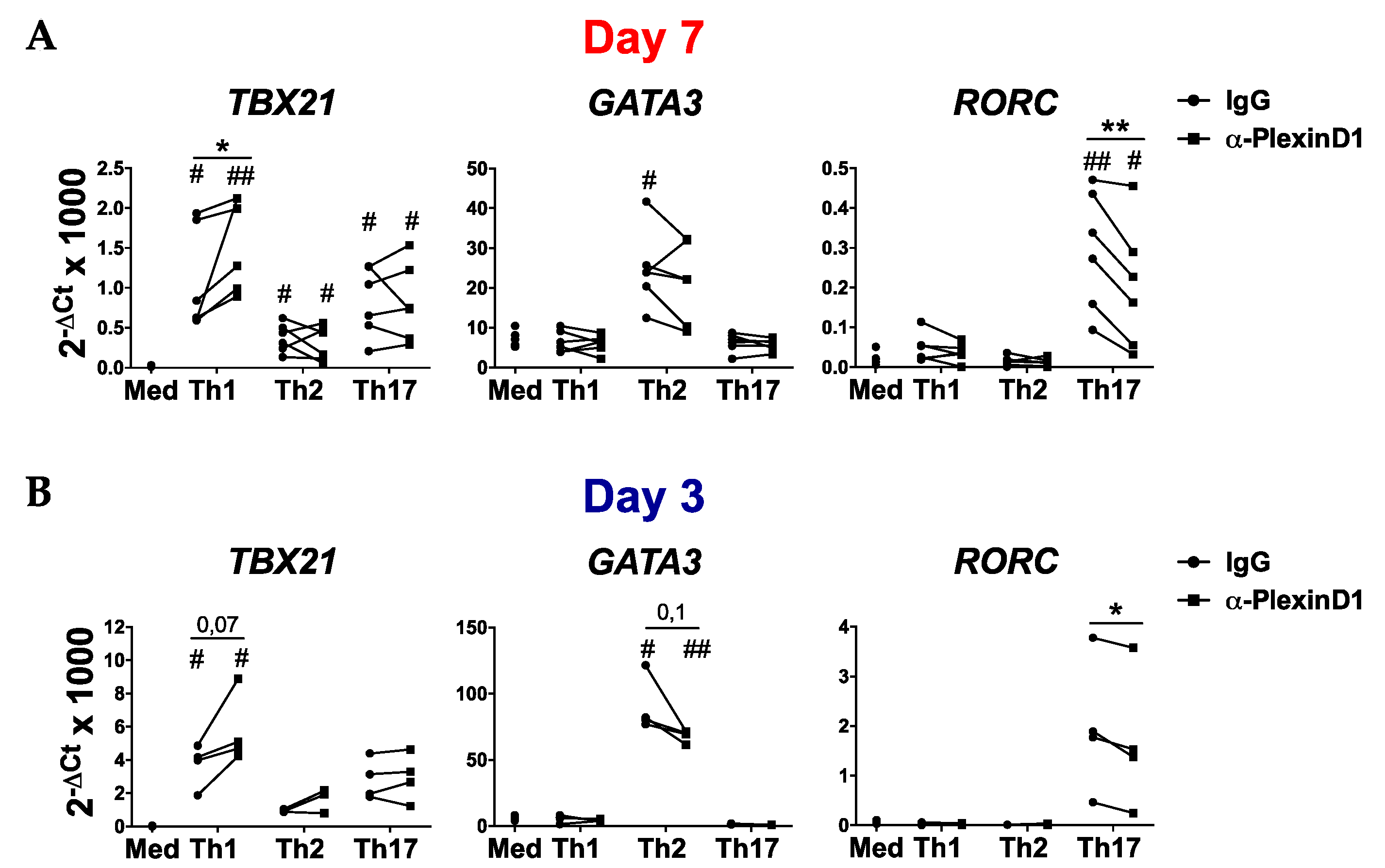
2.3. PlexinD1 Neutralization Regulates TBX21, GATA3 and RORC Expression
2.4. Sema4A-PlexinD1 Blockade Reduces PlexinB2 Expression during Th17 Cell Differentiation
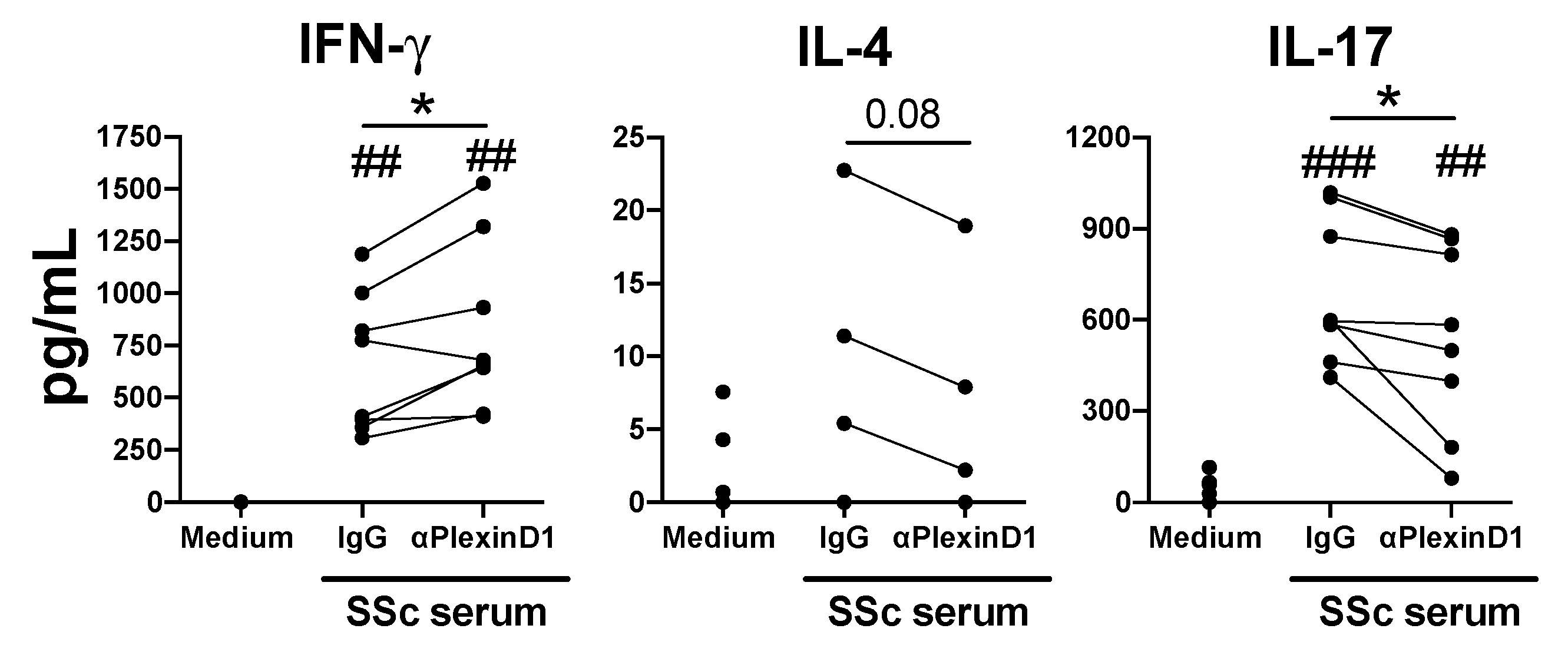
2.5. PlexinD1 Neutralization Regulates CD4+ T Cells Cytokine Production Induced by Systemic Sclerosis Patient Serum
3. Discussion
4. Materials and Methods
4.1. Naïve CD4+ T Cell Isolation
4.2. Naïve CD4+ T Cell Differentiation
4.3. CD4+ T Cell Stimulation
4.4. Flow Cytometry
4.5. Cytokine Measurement
4.6. RT-PCR and Quantitative qPCR
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Sema4A | Semaphorin4A |
| Th | T helper |
| IFN-γ | Interferon-γ |
| IL | Interleukin |
| ILT4 | Immunoglobulin-like transcript 4 |
| SSc | Systemic sclerosis |
| NRP-1 | Neuropilin-1 |
References
- Zhu, J. T helper cell differentiation, heterogeneity, and plasticity. Cold Spring Harb. Perspect. Biol. 2018, 10, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Yamane, H.; Paul, W.E. Differentiation of Effector CD4 T Cell Populations. Annu. Rev. Immunol. 2010, 28, 445–489. [Google Scholar] [CrossRef] [Green Version]
- Fang, D.; Zhu, J. Dynamic balance between master transcription factors determines the fates and functions of CD4 T cell and innate lymphoid cell subsets. J. Exp. Med. 2017, 214, 1861–1876. [Google Scholar] [CrossRef]
- Szabo, S.J.; Sullivan, B.M.; Peng, S.L.; Glimcher, L.H. M Olecular M Echanisms R Egulating T H 1 I Mmune R Esponses. Annu. Rev. Immunol. 2003, 21, 713–758. [Google Scholar] [CrossRef] [PubMed]
- Gaffen, S.L.; Jain, R.; Garg, A.V.; Cua, D.J. The IL-23-IL-17 immune axis: From mechanisms to therapeutic testing. Nat. Rev. Immunol. 2014, 14, 585–600. [Google Scholar] [CrossRef] [PubMed]
- Korn, T.; Bettelli, E.; Oukka, M.; Kuchroo, V.K. IL-17 and Th17 Cells. Annu. Rev. Immunol. 2009, 27, 485–517. [Google Scholar] [CrossRef]
- Murdaca, G.; Colombo, B.M.; Puppo, F. The role of Th17 lymphocytes in the autoimmune and chronic inflammatory diseases. Intern. Emerg. Med. 2011, 6, 487–495. [Google Scholar] [CrossRef]
- Palm, N.W.; Rosenstein, R.K.; Medzhitov, R. Allergic host defences. Nature 2012, 484, 465–472. [Google Scholar] [CrossRef]
- Licona-limón, P.; Kim, L.K.; Palm, N.W.; Flavell, R.A. REVIEW T H 2, allergy and group 2 innate lymphoid cells. Nat. Immunol. 2013, 14, 536–542. [Google Scholar] [CrossRef]
- Worzfeld, T.; Offermanns, S. Semaphorins and plexins as therapeutic targets. Nat. Rev. Drug Discov. 2014, 13, 603–621. [Google Scholar] [CrossRef]
- Nakatsuji, Y.; Okuno, T.; Moriya, M.; Sugimoto, T.; Kinoshita, M.; Takamatsu, H.; Nojima, S.; Kimura, T.; Kang, S.; Ito, D.; et al. Elevation of Sema4A Implicates Th Cell Skewing and the Efficacy of IFN- Therapy in Multiple Sclerosis. J. Immunol. 2012, 188, 4858–4865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, N.; Li, Y.; Zhang, Z.; Xing, J.; Sun, Y.; Yao, S.; Chen, L. Human Semaphorin-4A drives Th2 responses by binding to receptor ILT-4. Nat. Commun. 2018, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Carvalheiro, T.; Affandi, A.J.; Malvar-Fernández, B.; Dullemond, I.; Cossu, M.; Ottria, A.; Mertens, J.S.; Giovannone, B.; Bonte-Mineur, F.; Kok, M.R.; et al. Induction of Inflammation and Fibrosis by Semaphorin 4A in Systemic Sclerosis. Arthritis Rheumatol. 2019, 71, 1711–1722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumanogoh, A.; Marukawa, S.; Suzuki, K.; Takegahara, N.; Watanabe, C.; Ch’ng, E.S.; Ishida, I.; Fujimura, H.; Sakoda, S.; Yoshida, K.; et al. Class iv semaphorin sema4a enhances t-cell activation and interacts with tim-2. Nature 2002, 419, 629–633. [Google Scholar] [CrossRef] [PubMed]
- Toyofuku, T.; Yabuki, M.; Kamei, J.; Kamei, M.; Makino, N.; Kumanogoh, A.; Hori, M. Semaphorin-4A, an activator for T-cell-mediated immunity, suppresses angiogenesis via Plexin-D1. EMBO J. 2007, 26, 1373–1384. [Google Scholar] [CrossRef]
- Delgoffe, G.M.; Woo, S.-R.; Turnis, M.E.; Gravano, D.M.; Guy, C.; Overacre, A.E.; Bettini, M.L.; Vogel, P.; Finkelstein, D.; Bonnevier, J.; et al. Stability and function of regulatory T cells is maintained by a neuropilin-1–semaphorin-4a axis. Nature 2013, 501, 252–256. [Google Scholar] [CrossRef]
- Ito, D.; Nojima, S.; Nishide, M.; Okuno, T.; Takamatsu, H.; Kang, S.; Kimura, T.; Yoshida, Y.; Morimoto, K.; Maeda, Y.; et al. mTOR Complex Signaling through the SEMA4A–Plexin B2 Axis Is Required for Optimal Activation and Differentiation of CD8 + T Cells. J. Immunol. 2015, 195, 934–943. [Google Scholar] [CrossRef] [Green Version]
- Chapoval, S.P.; Hritzo, M.; Qi, X.; Tamagnone, L.; Golding, A.; Keegan, A.D. Semaphorin 4A Stabilizes Human Regulatory T Cell Phenotype via Plexin B1. ImmunoHorizons 2019, 3, 71–87. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Song, G.; Zheng, Y.; Tan, W.; Pan, J.; Zhao, Y.; Chang, X. Expression of Semaphorin 4A and its potential role in rheumatoid arthritis. Arthritis Res. Ther. 2015, 17, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Kumanogoh, A.; Shikina, T.; Suzuki, K.; Uematsu, S.; Yukawa, K.; Kashiwamura, S.I.; Tsutsui, H.; Yamamoto, M.; Takamatsu, H.; Ko-Mitamura, E.P.; et al. Nonredundant roles of Sema4A in the immune system: Defective T cell priming and Th1/Th2 regulation in Sema4A-deficient mice. Immunity 2005, 22, 305–316. [Google Scholar] [CrossRef] [Green Version]
- Nkyimbeng-Takwi, E.H.; Shanks, K.; Smith, E.; Iyer, A.; Lipsky, M.M.; Detolla, L.J.; Kikutani, H.; Keegan, A.D.; Chapoval, S.P. Neuroimmune semaphorin 4A downregulates the severity of allergic response. Mucosal Immunol. 2012, 5, 409–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morihana, T.; Goya, S.; Mizui, M.; Yasui, T.; Prasad, D.V.R.; Kumanogoh, A.; Tamura, M.; Shikina, T.; Maeda, Y.; Iwamoto, Y.; et al. An inhibitory role for Sema4A in antigen-specific allergic asthma. J. Clin. Immunol. 2013, 33, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Makino, N.; Toyofuku, T.; Takegahara, N.; Takamatsu, H.; Okuno, T.; Nakagawa, Y.; Kang, S.; Nojima, S.; Hori, M.; Kikutani, H.; et al. Involvement of Sema4A in the progression of experimental autoimmune myocarditis. FEBS Lett. 2008, 582, 3935–3940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koda, T.; Okuno, T.; Takata, K.; Honorat, J.A.; Kinoshita, M.; Tada, S.; Moriya, M.; Sakoda, S.; Mochizuki, H.; Kumanogoh, A.; et al. Sema4A inhibits the therapeutic effect of IFN-β in EAE. J. Neuroimmunol. 2014, 268, 43–49. [Google Scholar] [CrossRef]
- Gasparini, G.; Cozzani, E.; Parodi, A. Interleukin-4 and interleukin-13 as possible therapeutic targets in systemic sclerosis. Cytokine 2020, 125, 154799. [Google Scholar] [CrossRef]
- Allanore, Y.; Simms, R.; Distler, O.; Trojanowska, M.; Pope, J.; Denton, C.P.; Varga, J. Systemic sclerosis. Nat. Rev. Dis. Prim. 2015, 1, 1–21. [Google Scholar] [CrossRef]
- Radstake, T.R.D.J.; van Bon, L.; Broen, J.; Hussiani, A.; Hesselstrand, R.; Wuttge, D.M.; Deng, Y.; Simms, R.; Lubberts, E.; Lafyatis, R. The pronounced Th17 profile in systemic sclerosis (SSc) together with intracellular expression of TGF?? and IFN?? distinguishes SSc phenotypes. PLoS ONE 2009, 4, e5903. [Google Scholar] [CrossRef] [Green Version]
- Truchetet, M.E.; Brembilla, N.C.; Montanari, E.; Lonati, P.; Raschi, E.; Zeni, S.; Fontao, L.; Meroni, P.L.; Chizzolini, C. Interleukin-17A+ Cell Counts Are Increased in Systemic Sclerosis Skin and Their Number Is Inversely Correlated with the Extent of Skin Involvement. Arthritis Rheum. 2013, 65, 1347–1356. [Google Scholar] [CrossRef]
- Gonçalves, R.S.G.; Pereira, M.C.; Dantas, A.T.; de Almeida, A.R.; Marques, C.D.L.; Rego, M.J.B.M.; Pitta, I.R.; Duarte, A.L.B.P.; Pitta, M.G.R. IL-17 and related cytokines involved in systemic sclerosis: Perspectives. Autoimmunity 2018, 51, 1–9. [Google Scholar] [CrossRef]
- Kumanogoh, A.; Kikutani, H. Immunological functions of the neuropilins and plexins as receptors for semaphorins. Nat. Rev. Immunol. 2013, 13, 802–814. [Google Scholar] [CrossRef]
- Suzuki, K.; Kumanogoh, A.; Kikutani, H. Semaphorins and their receptors in immune cell interactions. Nat. Immunol. 2008, 9, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Hu, J.; Uemura, A.; Tetzlaff, F.; Augustin, H.G.; Fischer, A. Semaphorin-3C signals through Neuropilin-1 and PlexinD1 receptors to inhibit pathological angiogenesis. EMBO Mol. Med. 2015, 7, 1267–1284. [Google Scholar] [CrossRef]
- Liu, X.; Uemura, A.; Fukushima, Y.; Yoshida, Y.; Hirashima, M. Semaphorin 3G Provides a Repulsive Guidance Cue to Lymphatic Endothelial Cells via Neuropilin-2/PlexinD1. Cell Rep. 2016, 17, 2299–2311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamm, M.J.; Kirchmaier, B.C.; Herzog, W. Sema3d controls collective endothelial cell migration by distinct mechanisms via nrp1 and plxnD1. J. Cell Biol. 2016, 215, 415–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Singh, M.K.; Degenhardt, K.R.; Lu, M.M.; Bennett, J.; Yoshida, Y.; Epstein, J.A. Tie2Cre-mediated inactivation of plexinD1 results in congenital heart, vascular and skeletal defects. Dev. Biol. 2009, 325, 82–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bribián, A.; Nocentini, S.; Llorens, F.; Gil, V.; Mire, E.; Reginensi, D.; Yoshida, Y.; Mann, F.; Del Río, J.A. Sema3E/PlexinD1 regulates the migration of hem-derived Cajal-Retzius cells in developing cerebral cortex. Nat. Commun. 2014, 5, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Gu, C.; Yoshida, Y.; Livet, J.; Reimert, D.V.; Mann, F.; Merte, J.; Henderson, C.E.; Jessell, T.M.; Kolodkin, A.L.; Ginty, D.D. Semaphorin 3E and plexin-D1 control vascular pattern independently of neuropilins. Science 2005, 307, 265–268. [Google Scholar] [CrossRef]
- Mata, A.; Gil, V.; Pérez-Clausell, J.; Dasilva, M.; González-Calixto, M.C.; Soriano, E.; García-Verdugo, J.M.; Sanchez-Vives, M.V.; Del Río, J.A. New functions of Semaphorin 3E and its receptor PlexinD1 during developing and adult hippocampal formation. Sci. Rep. 2018, 8, 1–16. [Google Scholar] [CrossRef]
- Sawada, M.; Ohno, N.; Kawaguchi, M.; Huang, S.-H.; Hikita, T.; Sakurai, Y.; Bang Nguyen, H.; Quynh Thai, T.; Ishido, Y.; Yoshida, Y.; et al. PlexinD1 signaling controls morphological changes and migration termination in newborn neurons. EMBO J. 2018, 37, e97404. [Google Scholar] [CrossRef]
- Sandireddy, R.; Cibi, D.M.; Gupta, P.; Singh, A.; Tee, N.; Uemura, A.; Epstein, J.A.; Singh, M.K. Semaphorin 3E/PlexinD1 signaling is required for cardiac ventricular compaction. JCI Insight 2019, 4, e125908. [Google Scholar] [CrossRef]
- Van Den Hoogen, F.; Khanna, D.; Fransen, J.; Johnson, S.R.; Baron, M.; Tyndall, A.; Matucci-Cerinic, M.; Naden, R.P.; Medsger, T.A.; Carreira, P.E.; et al. 2013 classification criteria for systemic sclerosis: An american college of rheumatology/European league against rheumatism collaborative initiative. Arthritis Rheum. 2013, 65, 2737–2747. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carvalheiro, T.; Rafael-Vidal, C.; Malvar-Fernandez, B.; Lopes, A.P.; Pego-Reigosa, J.M.; Radstake, T.R.D.J.; Garcia, S. Semaphorin4A-Plexin D1 Axis Induces Th2 and Th17 While Represses Th1 Skewing in an Autocrine Manner. Int. J. Mol. Sci. 2020, 21, 6965. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186965
Carvalheiro T, Rafael-Vidal C, Malvar-Fernandez B, Lopes AP, Pego-Reigosa JM, Radstake TRDJ, Garcia S. Semaphorin4A-Plexin D1 Axis Induces Th2 and Th17 While Represses Th1 Skewing in an Autocrine Manner. International Journal of Molecular Sciences. 2020; 21(18):6965. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186965
Chicago/Turabian StyleCarvalheiro, Tiago, Carlos Rafael-Vidal, Beatriz Malvar-Fernandez, Ana P. Lopes, Jose M. Pego-Reigosa, Timothy R. D. J. Radstake, and Samuel Garcia. 2020. "Semaphorin4A-Plexin D1 Axis Induces Th2 and Th17 While Represses Th1 Skewing in an Autocrine Manner" International Journal of Molecular Sciences 21, no. 18: 6965. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186965