A Novel Cis-Acting RNA Structural Element Embedded in the Core Coding Region of the Hepatitis C Virus Genome Directs Internal Translation Initiation of the Overlapping Core+1 ORF

 and
and
Abstract
:1. Introduction
2. Results
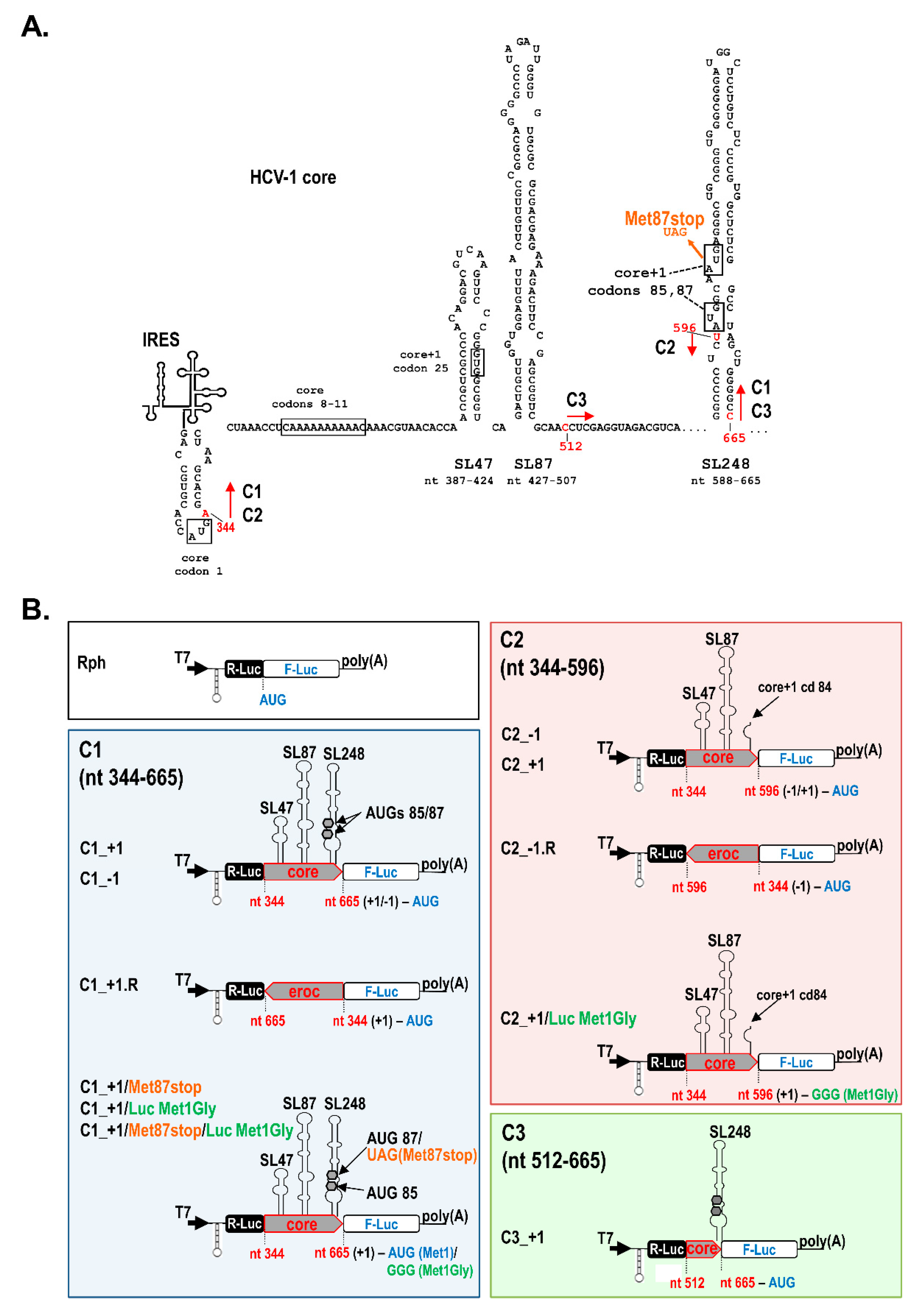
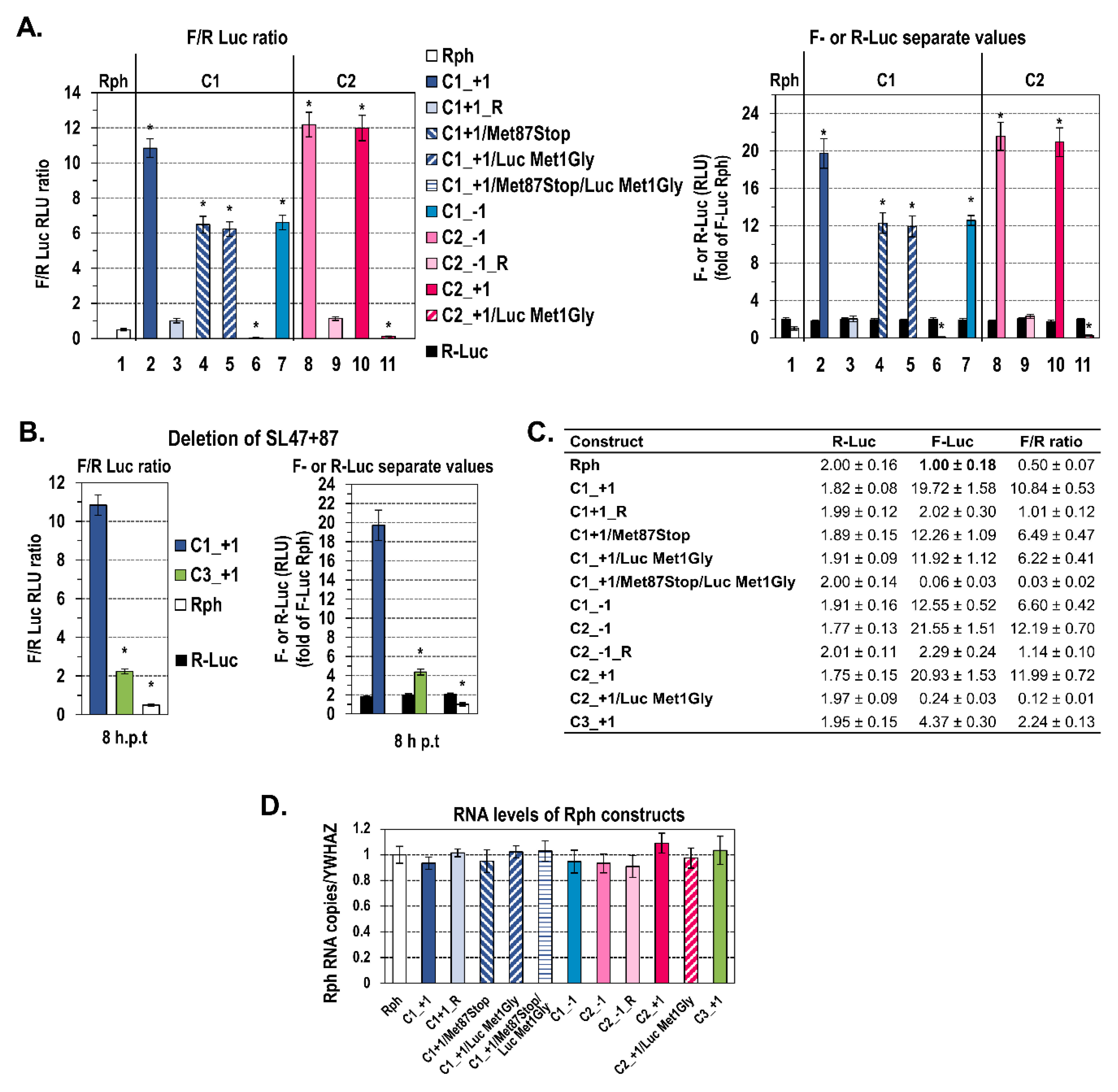
2.1. A Novel RNA Structural Element in the Core Coding Region Drives Internal Translation Initiation of the Core+1 ORF
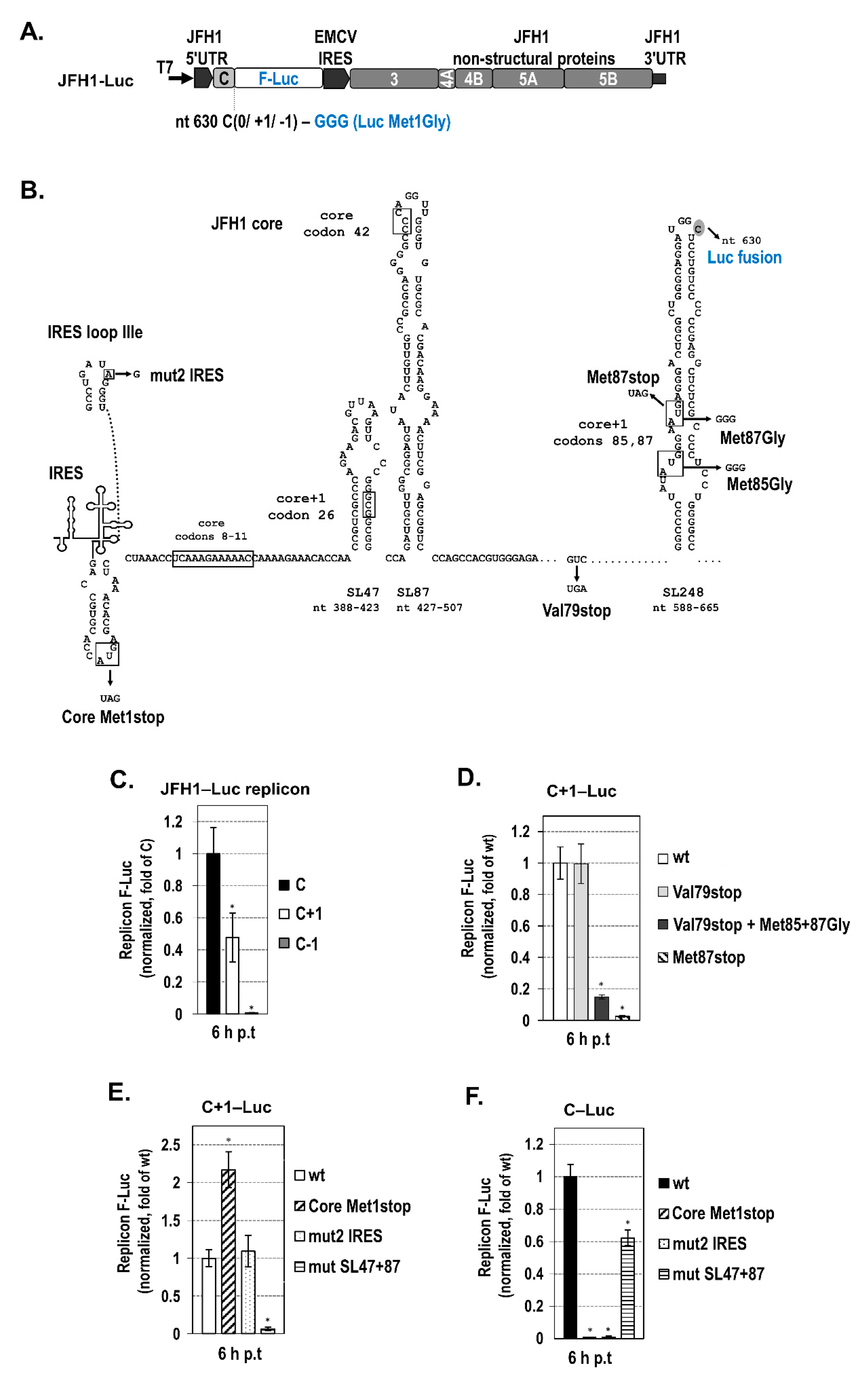
2.2. The Presence of Intact SL47 (nt 387–424) and SL87 (nt 427–507) but Not SL248 (nt 588–665) Is Necessary for Internal Translation Initiation at Codons 85/87 of Core+1 ORF
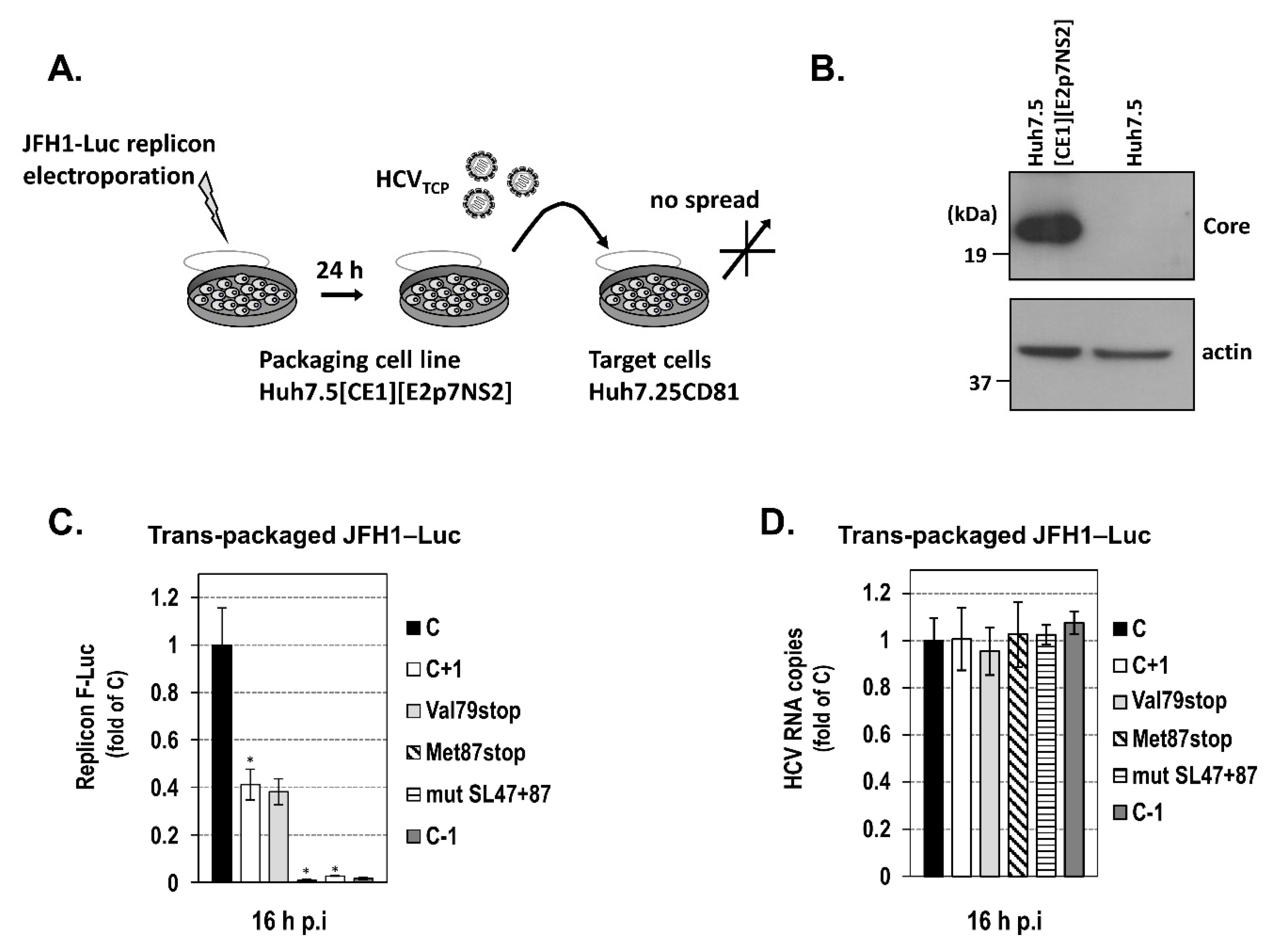
2.3. The Intact Core RNA SL47 and SL87 Structures Support Core+1/S Expression from the JFH1 Replicon
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Plasmid Construction and Site-Directed Mutagenesis
4.3. In Vitro Transcription
4.4. Transfection with In Vitro Transcribed RNA
4.5. Preparation of Trans-Complemented HCV Particles ([HCVTCP]Luc-NS3-5B) and Infection Assays
4.6. Luciferase and Bradford Assays
4.7. Total RNA Extraction and Quantification of Rph RNAs and Viral Replicons
4.8. Gel Electrophoresis and Western Blot Analysis
4.9. Polyribosome Fractionation and Quantification of Ribosome Associated RNAs
4.10. Statistical Analysis
4.11. RNA Secondary Structure Prediction
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chen, S.L.; Morgan, T.R. The natural history of hepatitis C virus (HCV) infection. Int. J. Med. Sci. 2006, 3, 47–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilder, J.; Patel, K. A Review of the Natural History of Chronic Hepatitis C Infection. N. Am. J. Med. Sci. 2014, 7, 1–7. [Google Scholar] [CrossRef]
- Hoofnagle, J.H. Course and outcome of hepatitis C. Hepatology 2002, 36, S21–S29. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Hepatitis C. Available online: https://www.who.int/en/news-room/fact-sheets/detail/hepatitis-c. (accessed on 31 July 2020).
- Duncan, J.D.; Urbanowicz, R.A.; Tarr, A.W.; Ball, J.K. Hepatitis C Virus Vaccine: Challenges and Prospects. Vaccines 2020, 8, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asselah, T.; Marcellin, P.; Schinazi, R.F. Treatment of hepatitis C virus infection with direct-acting antiviral agents: 100% cure? Liver Int. Off. J. Int. Assoc. Study Liver 2018, 38, 7–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terrault, N.A. Hepatitis C elimination: Challenges with under-diagnosis and under-treatment. F1000 Res. 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Bartenschlager, R.; Lohmann, V.; Penin, F. The molecular and structural basis of advanced antiviral therapy for hepatitis C virus infection. Nat. Rev. Microbiol. 2013, 11, 482–496. [Google Scholar] [CrossRef] [Green Version]
- Moradpour, D.; Penin, F.; Rice, C.M. Replication of hepatitis C virus. Nat. Rev. Microbiol. 2007, 5, 453–463. [Google Scholar] [CrossRef]
- Niepmann, M. Hepatitis C virus RNA translation. Curr. Top. Microbiol. Immunol. 2013, 369, 143–166. [Google Scholar] [CrossRef]
- Vassilaki, N.; Mavromara, P. The HCV ARFP/F/core+1 protein: Production and functional analysis of an unconventional viral product. IUBMB Life 2009, 61, 739–752. [Google Scholar] [CrossRef]
- Branch, A.D.; Stump, D.D.; Gutierrez, J.A.; Eng, F.; Walewski, J.L. The hepatitis C virus alternate reading frame (ARF) and its family of novel products: The alternate reading frame protein/F-protein, the double-frameshift protein, and others. Semin. Liver Dis. 2005, 25, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Boumlic, A.; Vassilaki, N.; Dalagiorgou, G.; Kochlios, E.; Kakkanas, A.; Georgopoulou, U.; Markoulatos, P.; Orfanoudakis, G.; Mavromara, P. Internal translation initiation stimulates expression of the ARF/core+1 open reading frame of HCV genotype 1b. Virus Res. 2011, 155, 213–220. [Google Scholar] [CrossRef]
- Vassilaki, N.; Mavromara, P. Two alternative translation mechanisms are responsible for the expression of the HCV ARFP/F/core+1 coding open reading frame. J. Biol. Chem. 2003, 278, 40503–40513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vassilaki, N.; Boleti, H.; Mavromara, P. Expression studies of the core+1 protein of the hepatitis C virus 1a in mammalian cells. The influence of the core protein and proteasomes on the intracellular levels of core+1. FEBS J. 2007, 274, 4057–4074. [Google Scholar] [CrossRef] [PubMed]
- Vassilaki, N.; Boleti, H.; Mavromara, P. Expression studies of the HCV-1a core+1 open reading frame in mammalian cells. Virus Res. 2008, 133, 123–135. [Google Scholar] [CrossRef]
- Kaul, A.; Woerz, I.; Meuleman, P.; Leroux-Roels, G.; Bartenschlager, R. Cell culture adaptation of hepatitis C virus and in vivo viability of an adapted variant. J. Virol. 2007, 81, 13168–13179. [Google Scholar] [CrossRef] [Green Version]
- Kotta-Loizou, I.; Karakasiliotis, I.; Vassilaki, N.; Sakellariou, P.; Bartenschlager, R.; Mavromara, P. Expression of the novel hepatitis C virus core+1/ARF protein in the context of JFH1-based replicons. J. Virol. 2015, 89, 5164–5170. [Google Scholar] [CrossRef] [Green Version]
- Baril, M.; Brakier-Gingras, L. Translation of the F protein of hepatitis C virus is initiated at a non-AUG codon in a +1 reading frame relative to the polyprotein. Nucleic Acids Res. 2005, 33, 1474–1486. [Google Scholar] [CrossRef] [Green Version]
- Cohen, M.; Bachmatov, L.; Ben-Ari, Z.; Rotman, Y.; Tur-Kaspa, R.; Zemel, R. Development of specific antibodies to an ARF protein in treated patients with chronic HCV infection. Dig. Dis. Sci. 2007, 52, 2427–2432. [Google Scholar] [CrossRef]
- Komurian-Pradel, F.; Rajoharison, A.; Berland, J.L.; Khouri, V.; Perret, M.; Van Roosmalen, M.; Pol, S.; Negro, F.; Paranhos-Baccala, G. Antigenic relevance of F protein in chronic hepatitis C virus infection. Hepatology 2004, 40, 900–909. [Google Scholar] [CrossRef]
- Varaklioti, A.; Vassilaki, N.; Georgopoulou, U.; Mavromara, P. Alternate translation occurs within the core coding region of the hepatitis C viral genome. J. Biol. Chem. 2002, 277, 17713–17721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walewski, J.L.; Keller, T.R.; Stump, D.D.; Branch, A.D. Evidence for a new hepatitis C virus antigen encoded in an overlapping reading frame. RNA 2001, 7, 710–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Choi, J.; Yen, T.S.; Lu, W.; Strohecker, A.; Govindarajan, S.; Chien, D.; Selby, M.J.; Ou, J. Synthesis of a novel hepatitis C virus protein by ribosomal frameshift. EMBO J. 2001, 20, 3840–3848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuang, W.C.; Allain, J.P. Differential reactivity of putative genotype 2 hepatitis C virus F protein between chronic and recovered infections. J. Gen. Virol. 2008, 89, 1890–1900. [Google Scholar] [CrossRef] [PubMed]
- Karamitros, T.; Kakkanas, A.; Katsoulidou, A.; Sypsa, V.; Dalagiorgou, G.; Mavromara, P.; Hatzakis, A. Detection of specific antibodies to HCV-ARF/CORE+1 protein in patients treated with pegylated interferon plus ribavirin. J. Viral Hepat. 2012, 19, 182–188. [Google Scholar] [CrossRef]
- Ajorloo, M.; Bamdad, T.; Hashempour, T.; Alborzi, A.M.; Mozhgani, S.H.; Asadi, R.; Haj-sheykholeslami, A.; Merat, S. Detection of Specific Antibodies to HCV-ARF/CORE+1 Protein in Cirrhotic and Non-Cirrhotic Patients with Hepatitis C: A Possible Association with Progressive Fibrosis. Arch. Iran. Med. 2015, 18, 304–307. [Google Scholar] [CrossRef]
- Morice, Y.; Ratinier, M.; Miladi, A.; Chevaliez, S.; Germanidis, G.; Wedemeyer, H.; Laperche, S.; Lavergne, J.P.; Pawlotsky, J.M. Seroconversion to hepatitis C virus alternate reading frame protein during acute infection. Hepatology 2009, 49, 1449–1459. [Google Scholar] [CrossRef] [Green Version]
- Shehat, M.G.; Bahey-El-Din, M.; Kassem, M.A.; Farghaly, F.A.; Abdul-Rahman, M.H.; Fanaki, N.H. Recombinant expression of the alternate reading frame protein (ARFP) of hepatitis C virus genotype 4a (HCV-4a) and detection of ARFP and anti-ARFP antibodies in HCV-infected patients. Arch. Virol. 2015, 160, 1939–1952. [Google Scholar] [CrossRef]
- Tobler, L.H.; Stramer, S.L.; Chien, D.Y.; Lin, S.; Arcangel, P.; Phelps, B.H.; Cooper, S.L.; Busch, M.P. Antibodies to a novel antigen in acute hepatitis C virus infections. Vox Sang. 2007, 92, 1–7. [Google Scholar] [CrossRef]
- Gao, D.Y.; Zhang, X.X.; Hou, G.; Jin, G.D.; Deng, Q.; Kong, X.F.; Zhang, D.H.; Ling, Y.; Yu, D.M.; Gong, Q.M.; et al. Assessment of specific antibodies to F protein in serum samples from Chinese hepatitis C patients treated with interferon plus ribavarin. J. Clin. Microbiol. 2008, 46, 3746–3751. [Google Scholar] [CrossRef] [Green Version]
- Bain, C.; Parroche, P.; Lavergne, J.P.; Duverger, B.; Vieux, C.; Dubois, V.; Komurian-Pradel, F.; Trepo, C.; Gebuhrer, L.; Paranhos-Baccala, G.; et al. Memory T-cell-mediated immune responses specific to an alternative core protein in hepatitis C virus infection. J. Virol. 2004, 78, 10460–10469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Troesch, M.; Jalbert, E.; Canobio, S.; Boulassel, M.R.; Routy, J.P.; Bernard, N.F.; Bruneau, J.; Lapointe, N.; Boucher, M.; Soudeyns, H. Characterization of humoral and cell-mediated immune responses directed against hepatitis C virus F protein in subjects co-infected with hepatitis C virus and HIV-1. AIDS 2005, 19, 775–784. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.Y.; Jin, G.D.; Yao, B.L.; Zhang, D.H.; Gu, L.L.; Lu, Z.M.; Gong, Q.; Lone, Y.C.; Deng, Q.; Zhang, X.X. Characterization of the specific CD4+ T cell response against the F protein during chronic hepatitis C virus infection. PLoS ONE 2010, 5, e14237. [Google Scholar] [CrossRef] [PubMed]
- Kassela, K.; Karakasiliotis, I.; Charpantidis, S.; Koskinas, J.; Mylopoulou, T.; Mimidis, K.; Sarrazin, C.; Grammatikos, G.; Mavromara, P. High prevalence of antibodies to core+1/ARF protein in HCV-infected patients with advanced cirrhosis. J. Gen. Virol. 2017, 98, 1713–1719. [Google Scholar] [CrossRef]
- Dalagiorgou, G.; Vassilaki, N.; Foka, P.; Boumlic, A.; Kakkanas, A.; Kochlios, E.; Khalili, S.; Aslanoglou, E.; Veletza, S.; Orfanoudakis, G.; et al. High levels of HCV core+1 antibodies in HCV patients with hepatocellular carcinoma. J. Gen. Virol. 2011, 92, 1343–1351. [Google Scholar] [CrossRef]
- Moustafa, S.; Karakasiliotis, I.; Mavromara, P. Hepatitis C Virus core+1/ARF Protein Modulates the Cyclin D1/pRb Pathway and Promotes Carcinogenesis. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [Green Version]
- Tuplin, A.; Evans, D.J.; Simmonds, P. Detailed mapping of RNA secondary structures in core and NS5B-encoding region sequences of hepatitis C virus by RNase cleavage and novel bioinformatic prediction methods. J. Gen. Virol. 2004, 85, 3037–3047. [Google Scholar] [CrossRef]
- Tuplin, A.; Wood, J.; Evans, D.J.; Patel, A.H.; Simmonds, P. Thermodynamic and phylogenetic prediction of RNA secondary structures in the coding region of hepatitis C virus. RNA 2002, 8, 824–841. [Google Scholar] [CrossRef] [Green Version]
- Kato, T.; Furusaka, A.; Miyamoto, M.; Date, T.; Yasui, K.; Hiramoto, J.; Nagayama, K.; Tanaka, T.; Wakita, T. Sequence analysis of hepatitis C virus isolated from a fulminant hepatitis patient. J. Med. Virol. 2001, 64, 334–339. [Google Scholar] [CrossRef]
- Wakita, T.; Pietschmann, T.; Kato, T.; Date, T.; Miyamoto, M.; Zhao, Z.; Murthy, K.; Habermann, A.; Krausslich, H.G.; Mizokami, M.; et al. Production of infectious hepatitis C virus in tissue culture from a cloned viral genome. Nat. Med. 2005, 11, 791–796. [Google Scholar] [CrossRef] [Green Version]
- Vassilaki, N.; Friebe, P.; Meuleman, P.; Kallis, S.; Kaul, A.; Paranhos-Baccala, G.; Leroux-Roels, G.; Mavromara, P.; Bartenschlager, R. Role of the hepatitis C virus core+1 open reading frame and core cis-acting RNA elements in viral RNA translation and replication. J. Virol. 2008, 82, 11503–11515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, Q.; Wang, Q.S.; Firth, A.E.; Chan, M.M.; Gouw, J.W.; Guarna, M.M.; Foster, L.J.; Atkins, J.F.; Jan, E. Alternative reading frame selection mediated by a tRNA-like domain of an internal ribosome entry site. Proc. Natl. Acad. Sci. USA 2012, 109, E630–E639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerr, C.H.; Wang, Q.S.; Moon, K.M.; Keatings, K.; Allan, D.W.; Foster, L.J.; Jan, E. IRES-dependent ribosome repositioning directs translation of a +1 overlapping ORF that enhances viral infection. Nucleic Acids Res. 2018, 46, 11952–11967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaag, H.M.; Kawchuk, L.; Rohde, W.; Fischer, R.; Emans, N.; Prufer, D. An unusual internal ribosomal entry site of inverted symmetry directs expression of a potato leafroll polerovirus replication-associated protein. Proc. Natl. Acad. Sci. USA 2003, 100, 8939–8944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lulla, V.; Dinan, A.M.; Hosmillo, M.; Chaudhry, Y.; Sherry, L.; Irigoyen, N.; Nayak, K.M.; Stonehouse, N.J.; Zilbauer, M.; Goodfellow, I.; et al. An upstream protein-coding region in enteroviruses modulates virus infection in gut epithelial cells. Nat. Microbiol. 2019, 4, 280–292. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Li, Y.; Liu, G.; Jiang, Y.; Shen, S.; Bi, R.; Huang, H.; Cheng, T.; Wang, C.; Wei, W. A second open reading frame in human enterovirus determines viral replication in intestinal epithelial cells. Nat. Commun. 2019, 10, 4066. [Google Scholar] [CrossRef] [Green Version]
- Andreev, D.E.; Fernandez-Miragall, O.; Ramajo, J.; Dmitriev, S.E.; Terenin, I.M.; Martinez-Salas, E.; Shatsky, I.N. Differential factor requirement to assemble translation initiation complexes at the alternative start codons of foot-and-mouth disease virus RNA. RNA 2007, 13, 1366–1374. [Google Scholar] [CrossRef] [Green Version]
- Chappell, S.A.; Edelman, G.M.; Mauro, V.P. A 9-nt segment of a cellular mRNA can function as an internal ribosome entry site (IRES) and when present in linked multiple copies greatly enhances IRES activity. Proc. Natl. Acad. Sci. USA 2000, 97, 1536–1541. [Google Scholar] [CrossRef] [Green Version]
- Owens, G.C.; Chappell, S.A.; Mauro, V.P.; Edelman, G.M. Identification of two short internal ribosome entry sites selected from libraries of random oligonucleotides. Proc. Natl. Acad. Sci. USA 2001, 98, 1471–1476. [Google Scholar] [CrossRef] [Green Version]
- Chappell, S.A.; Mauro, V.P. The internal ribosome entry site (IRES) contained within the RNA-binding motif protein 3 (Rbm3) mRNA is composed of functionally distinct elements. J. Biol. Chem. 2003, 278, 33793–33800. [Google Scholar] [CrossRef] [Green Version]
- Pinkstaff, J.K.; Chappell, S.A.; Mauro, V.P.; Edelman, G.M.; Krushel, L.A. Internal initiation of translation of five dendritically localized neuronal mRNAs. Proc. Natl. Acad. Sci. USA 2001, 98, 2770–2775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, S.R. So you want to know if your message has an IRES? Wiley Interdiscip. Rev. RNA 2012, 3, 697–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terenin, I.M.; Smirnova, V.V.; Andreev, D.E.; Dmitriev, S.E.; Shatsky, I.N. A researcher’s guide to the galaxy of IRESs. Cell. Mol. Life Sci. CMLS 2017, 74, 1431–1455. [Google Scholar] [CrossRef]
- Shatsky, I.N.; Dmitriev, S.E.; Terenin, I.M.; Andreev, D.E. Cap- and IRES-independent scanning mechanism of translation initiation as an alternative to the concept of cellular IRESs. Mol. Cells 2010, 30, 285–293. [Google Scholar] [CrossRef]
- Shulla, A.; Randall, G. Spatiotemporal analysis of hepatitis C virus infection. PLoS Pathog. 2015, 11, e1004758. [Google Scholar] [CrossRef] [PubMed]
- Kieft, J.S.; Zhou, K.; Jubin, R.; Doudna, J.A. Mechanism of ribosome recruitment by hepatitis C IRES RNA. RNA 2001, 7, 194–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lukavsky, P.J.; Otto, G.A.; Lancaster, A.M.; Sarnow, P.; Puglisi, J.D. Structures of two RNA domains essential for hepatitis C virus internal ribosome entry site function. Nat. Struct. Biol. 2000, 7, 1105–1110. [Google Scholar] [CrossRef]
- Psaridi, L.; Georgopoulou, U.; Varaklioti, A.; Mavromara, P. Mutational analysis of a conserved tetraloop in the 5’ untranslated region of hepatitis C virus identifies a novel RNA element essential for the internal ribosome entry site function. FEBS Lett. 1999, 453, 49–53. [Google Scholar] [CrossRef] [Green Version]
- Jaafar, Z.A.; Kieft, J.S. Viral RNA structure-based strategies to manipulate translation. Nat. Rev. Microbiol. 2019, 17, 110–123. [Google Scholar] [CrossRef]
- Buck, C.B.; Shen, X.; Egan, M.A.; Pierson, T.C.; Walker, C.M.; Siliciano, R.F. The human immunodeficiency virus type 1 gag gene encodes an internal ribosome entry site. J. Virol. 2001, 75, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Herbreteau, C.H.; Weill, L.; Decimo, D.; Prevot, D.; Darlix, J.L.; Sargueil, B.; Ohlmann, T. HIV-2 genomic RNA contains a novel type of IRES located downstream of its initiation codon. Nat. Struct. Mol. Biol. 2005, 12, 1001–1007. [Google Scholar] [CrossRef]
- Akuta, N.; Suzuki, F.; Kawamura, Y.; Yatsuji, H.; Sezaki, H.; Suzuki, Y.; Hosaka, T.; Kobayashi, M.; Kobayashi, M.; Arase, Y.; et al. Prediction of response to pegylated interferon and ribavirin in hepatitis C by polymorphisms in the viral core protein and very early dynamics of viremia. Intervirology 2007, 50, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Eng, F.J.; Walewski, J.L.; Klepper, A.L.; Fishman, S.L.; Desai, S.M.; McMullan, L.K.; Evans, M.J.; Rice, C.M.; Branch, A.D. Internal initiation stimulates production of p8 minicore, a member of a newly discovered family of hepatitis C virus core protein isoforms. J. Virol. 2009, 83, 3104–3114. [Google Scholar] [CrossRef] [Green Version]
- Jan, E.; Sarnow, P. Factorless ribosome assembly on the internal ribosome entry site of cricket paralysis virus. J. Mol. Biol. 2002, 324, 889–902. [Google Scholar] [CrossRef]
- Pirakitikulr, N.; Kohlway, A.; Lindenbach, B.D.; Pyle, A.M. The Coding Region of the HCV Genome Contains a Network of Regulatory RNA Structures. Mol. Cell 2016, 62, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Park, S.M.; Park, J.H.; Keum, S.J.; Jang, S.K. eIF2A mediates translation of hepatitis C viral mRNA under stress conditions. EMBO J. 2011, 30, 2454–2464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalliampakou, K.I.; Kalamvoki, M.; Mavromara, P. Hepatitis C virus (HCV) NS5A protein downregulates HCV IRES-dependent translation. J. Gen. Virol. 2005, 86, 1015–1025. [Google Scholar] [CrossRef]
- Gale, M., Jr.; Blakely, C.M.; Kwieciszewski, B.; Tan, S.L.; Dossett, M.; Tang, N.M.; Korth, M.J.; Polyak, S.J.; Gretch, D.R.; Katze, M.G. Control of PKR protein kinase by hepatitis C virus nonstructural 5A protein: Molecular mechanisms of kinase regulation. Mol. Cell. Biol. 1998, 18, 5208–5218. [Google Scholar] [CrossRef] [Green Version]
- Karamichali, E.; Foka, P.; Tsitoura, E.; Kalliampakou, K.; Kazazi, D.; Karayiannis, P.; Georgopoulou, U.; Mavromara, P. HCV NS5A co-operates with PKR in modulating HCV IRES-dependent translation. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2014, 26, 113–122. [Google Scholar] [CrossRef]
- Honda, M.; Rijnbrand, R.; Abell, G.; Kim, D.; Lemon, S.M. Natural variation in translational activities of the 5’ nontranslated RNAs of hepatitis C virus genotypes 1a and 1b: Evidence for a long-range RNA-RNA interaction outside of the internal ribosomal entry site. J. Virol. 1999, 73, 4941–4951. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.H.; Rijnbrand, R.C.; Lemon, S.M. Core protein-coding sequence, but not core protein, modulates the efficiency of cap-independent translation directed by the internal ribosome entry site of hepatitis C virus. J. Virol. 2000, 74, 11347–11358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.K.; Lee, S.H.; Kim, C.S.; Seol, S.K.; Jang, S.K. Long-range RNA-RNA interaction between the 5’ nontranslated region and the core-coding sequences of hepatitis C virus modulates the IRES-dependent translation. RNA 2003, 9, 599–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Paek, K.Y.; Ha, S.H.; Cho, S.; Choi, K.; Kim, C.S.; Ryu, S.H.; Jang, S.K. A cellular RNA-binding protein enhances internal ribosomal entry site-dependent translation through an interaction downstream of the hepatitis C virus polyprotein initiation codon. Mol. Cell. Biol. 2004, 24, 7878–7890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, B.; Lim, J.H.; Hahm, B.; Jang, S.K.; Lee, S.W. hnRNP L is required for the translation mediated by HCV IRES. Biochem. Biophys. Res. Commun. 2009, 378, 584–588. [Google Scholar] [CrossRef] [PubMed]
- Hahm, B.; Kim, Y.K.; Kim, J.H.; Kim, T.Y.; Jang, S.K. Heterogeneous nuclear ribonucleoprotein L interacts with the 3’ border of the internal ribosomal entry site of hepatitis C virus. J. Virol. 1998, 72, 8782–8788. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Lai, M.M. An internal polypyrimidine-tract-binding protein-binding site in the hepatitis C virus RNA attenuates translation, which is relieved by the 3’-untranslated sequence. Virology 1999, 254, 288–296. [Google Scholar] [CrossRef] [Green Version]
- Mauro, V.P.; Edelman, G.M. The ribosome filter hypothesis. Proc. Natl. Acad. Sci. USA 2002, 99, 12031–12036. [Google Scholar] [CrossRef] [Green Version]
- Mortimer, S.A.; Kidwell, M.A.; Doudna, J.A. Insights into RNA structure and function from genome-wide studies. Nat. Rev. Genet. 2014, 15, 469–479. [Google Scholar] [CrossRef]
- Firth, A.E.; Brierley, I. Non-canonical translation in RNA viruses. J. Gen. Virol. 2012, 93, 1385–1409. [Google Scholar] [CrossRef]
- Walsh, D.; Mathews, M.B.; Mohr, I. Tinkering with translation: Protein synthesis in virus-infected cells. Cold Spring Harb. Perspect. Biol. 2013, 5, a012351. [Google Scholar] [CrossRef]
- Wilson, T.M. Nucleocapsid disassembly and early gene expression by positive-strand RNA viruses. J. Gen. Virol. 1985, 66 Pt 6, 1201–1207. [Google Scholar] [CrossRef] [PubMed]
- Friebe, P.; Boudet, J.; Simorre, J.P.; Bartenschlager, R. Kissing-loop interaction in the 3’ end of the hepatitis C virus genome essential for RNA replication. J. Virol. 2005, 79, 380–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinmann, E.; Brohm, C.; Kallis, S.; Bartenschlager, R.; Pietschmann, T. Efficient trans-encapsidation of hepatitis C virus RNAs into infectious virus-like particles. J. Virol. 2008, 82, 7034–7046. [Google Scholar] [CrossRef] [Green Version]
- Akazawa, D.; Date, T.; Morikawa, K.; Murayama, A.; Miyamoto, M.; Kaga, M.; Barth, H.; Baumert, T.F.; Dubuisson, J.; Wakita, T. CD81 expression is important for the permissiveness of Huh7 cell clones for heterogeneous hepatitis C virus infection. J. Virol. 2007, 81, 5036–5045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalliampakou, K.I.; Psaridi-Linardaki, L.; Mavromara, P. Mutational analysis of the apical region of domain II of the HCV IRES. FEBS Lett. 2002, 511, 79–84. [Google Scholar] [CrossRef]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence | Template | Rph Plasmid (HCV nt seq) |
|---|---|---|---|
| C1,C2_F C1_+1_R | 5′-CCGGAATTCCGAGCACGAATCCTAAACCTC-3′ 5′-CGCCGGAATTCCGGGGCCCCAGCTAGGCCGAG-3′ | HCV-1 | C1_+1 (nt 344–665) |
| C1_+1.R (nt 665–344) | |||
| C1,C2_F C1_−1_R | 5′-CCGGAATTCCGAGCACGAATCCTAAACCTC-3′ 5΄-CGCCGGAATTCCGGCCCCAGCTAGGCCGAGAG-3΄ | HCV-1 | C1_−1 (nt 344–665) |
| C1,C2_F C2_−1_R | 5′-CCGGAATTCCGAGCACGAATCCTAAACCTC-3′ 5′-CTCGAATTCCAGAGGGGCCAAGGGTACCC-3′ | HCV-1 | C2_−1 (nt 344–596) |
| C2_−1.R (nt 596–344) | |||
| C1,C2_F C2_+1_R | 5′-CCGGAATTCCGAGCACGAATCCTAAACCTC-3′ 5′-CCGGAATTCCGAGGGGCCAAGGGTACCCG-3′ | HCV-1 | C2_+1 C2_+1/Luc Met1Gly (nt 344–596) |
| C3_F C1_+1_R | 5′- CGCCGGAATTCCTCGAGGTAGACGTCAGCC-3′ 5′-CGCCGGAATTCCGGGGCCCCAGCTAGGCCGAG-3′ | HCV-1 | C3_+1 (512–665) |
| C2_F_JFH1 C2_−1_R_ JFH1 | 5′-GCGCGCCGGAATTCCGAGCACAAATCCTAAACC TC-3′ 5′-CGCCGGAATTCCATAGGGGCCAGGGGCGACCTG-3′ | JFH1 | C2_−1.JFH1 (nt 344–596) |
| JFH1/mut SL47+87 [42] | C2_−1.JFH1/ mut SL47+87 | ||
| IRES_F IRES_R | 5′-CCGGAATTCCTGATGGGGGCGACACTCCACC-3′ 5′-CCGGAATTCCGGGACGTCCTGTGGGCGGCGGT TGGTG-3′ | pHPI-1046 [86] | 5′UTR IRES (nt 8–406) |
| Primer | Sequence | Template | Plasmid Constructed |
|---|---|---|---|
| Met85Gly F | 5′-GGCCCCTATGGGGGAATGAGG-3′ | C+1‒Luc (JFH1 replicon) | Met85Gly |
| Met85Gly R | 5′-CCTCATTCCCCCATAGGGGCC-3′ | ||
| Met87Gly F | 5′-CCTATATGGGAGGGAGGGACTCG-3′ | C+1‒Luc (JFH1 replicon) | Met87Gly |
| Met87Gly R | 5′-CGAGTCCCTCCCTCCCATATAGG-3′ | ||
| Core Met1stop F | 5′-GCACCTAGAGCACAAATCCTAAAC-3′ | C+1‒Luc C‒Luc (JFH1 replicon) | Core Met1stop |
| Core Met1stop R | 5′-GATTTGTGCTCTAGGTGCACGGTCTACG-3′ | ||
| mut2 IRES F | 5′-GGTACTGCCTGATGGGGCGCTTGCG-3′ | C+1‒Luc C‒Luc (JFH1 replicon) | mut2 IRES |
| mut2 IRES R | 5′-CGCAAGCGCCCCATCAGGCAGTACC-3′ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vassilaki, N.; Frakolaki, E.; Kalliampakou, K.I.; Sakellariou, P.; Kotta-Loizou, I.; Bartenschlager, R.; Mavromara, P. A Novel Cis-Acting RNA Structural Element Embedded in the Core Coding Region of the Hepatitis C Virus Genome Directs Internal Translation Initiation of the Overlapping Core+1 ORF. Int. J. Mol. Sci. 2020, 21, 6974. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186974
Vassilaki N, Frakolaki E, Kalliampakou KI, Sakellariou P, Kotta-Loizou I, Bartenschlager R, Mavromara P. A Novel Cis-Acting RNA Structural Element Embedded in the Core Coding Region of the Hepatitis C Virus Genome Directs Internal Translation Initiation of the Overlapping Core+1 ORF. International Journal of Molecular Sciences. 2020; 21(18):6974. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186974
Chicago/Turabian StyleVassilaki, Niki, Efseveia Frakolaki, Katerina I. Kalliampakou, Panagiotis Sakellariou, Ioly Kotta-Loizou, Ralf Bartenschlager, and Penelope Mavromara. 2020. "A Novel Cis-Acting RNA Structural Element Embedded in the Core Coding Region of the Hepatitis C Virus Genome Directs Internal Translation Initiation of the Overlapping Core+1 ORF" International Journal of Molecular Sciences 21, no. 18: 6974. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186974