Role of Alternatively Spliced Messenger RNA (mRNA) Isoforms of the Insulin-Like Growth Factor 1 (IGF1) in Selected Human Tumors


Abstract
:1. Introduction
2. Alternative Splicing (AS)—A Phenomenon in Physiology and Pathophysiology
3. IGF1 Gene Structure and Signaling
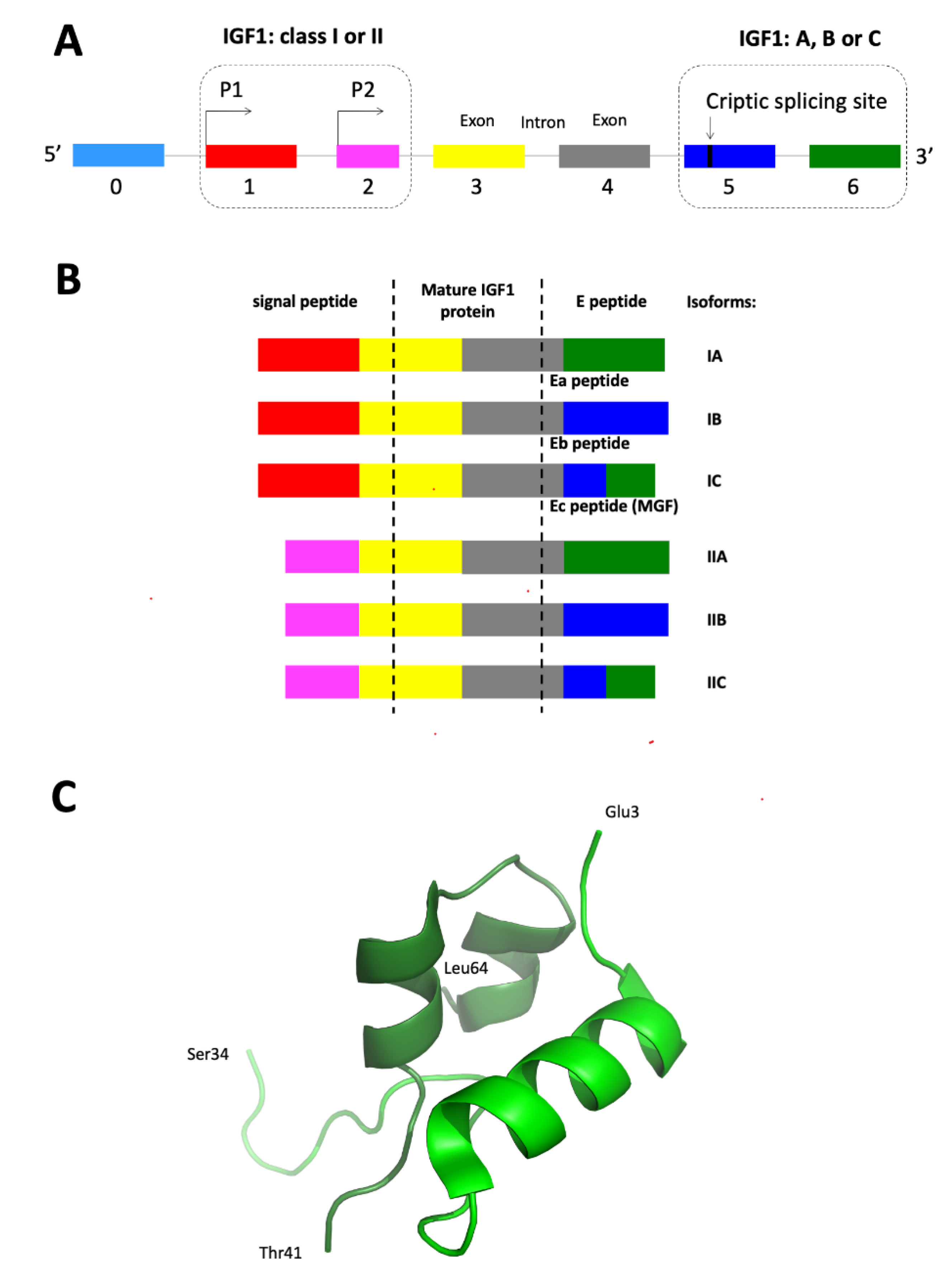
3.1. IGF1 Gene Promoters
3.2. IGF1 Gene Alternative Splicing and Promoter Usage
3.3. IGF1 Protein Processing
3.4. Regulating Mechanisms of IGF1 Gene Alternative Splicing
3.5. Phylogeny of the IGF1 Gene
4. IGF1 Isoforms and Their Function in Major Body Tissues
4.1. Epithelial Tissue
4.2. Connective Tissue
4.3. Muscle Tissue
4.3.1. Skeletal Muscle
4.3.2. Cardiac Muscle
4.4. Nervous Tissue
5. IGF1 Gene Alternative Splicing and Carcinogenesis
5.1. IGF1 Isoform Expression (mRNAs, Proteins) in Selected Tumors
5.1.1. Breast Cancer (BC)
5.1.2. Colorectal Cancer (CRC)
5.1.3. Endometrial Cancer (EC)
5.1.4. Epithelial Cervical Cancer (CC)
5.1.5. Hepatocellular Carcinoma (HCC)
5.1.6. Lung Cancer (LC)
5.1.7. Osteosarcoma
5.1.8. Prostate Cancer (PC)
5.1.9. Other Cancers
6. Aberrant IGF1 Isoform Expression (mRNAs, Proteins) and Cancer Therapy–Possible or Impossible Implications?
7. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| aa | amino acids |
| AKT | Serine-threonine Protein Kinase (now called AKT1) |
| AS | Alternative Splicing |
| BC | Breast Cancer |
| CC | Cervical Cancer |
| CD147 | Cluster of Differentiation 147 |
| c/n | Copy Number |
| CRC | Colorectal Cancer |
| CRISPR-Cas9 | Clustered Regularly-Interspaced Short Palindromic Repeats-CRISPR associated protein 9 |
| Dexa | Dexamethason |
| DHT | Dihydrotestosterone |
| E2 | Estradiol |
| EC | Endometrial Cancer |
| ECM | Extracellular Matrix |
| EMT | Epithelial-Mesenchymal Transition |
| ER | Estrogen Receptor |
| ERK1/2 | Extracellular Signal-regulated Kinase 1/2 |
| ESE | Exonic Splicing Enhancer |
| FOX2 | AS regulator, RNA binding motif protein 9 (RMB9) |
| GH | Growth Hormone |
| HCC | Hepatocellular Carcinoma |
| HCV | Hepatitis C Virus |
| H-SIL | High-grade Squamous Intraepithelial Lesions |
| IGF1, 2 | Insulin-like Growth Factor 1, 2 |
| IGF1R/2R | IGF1 Receptor type I/type II |
| INS | Insulin |
| INSR | INS Receptor |
| IHC | Immunohistochemistry |
| IL | Interleukin |
| LC | Lung Cancer |
| L-SIL | Low-grade Squamous Intraepithelial Lesions |
| mIGF1 | “muscle” specific IGF1 (IGF1Ea isoform) |
| MIR-b | Mammalian Interspersed Repetitive-b Element |
| MAPK | Mitogen-activated Protein Kinase (originally called ERK) |
| MGF | Mechano-growth factor |
| MMP-9 | Matrix Metalloproteinase 9 |
| MSCs | Mesenchymal Stem Cells |
| NPY | Neuropeptide Y |
| PC | Prostate Cancer |
| PGE2 | Prostaglandin E2 |
| PI3K | Phosphatidylinositol 3-Kinase |
| PKA, B (AKT), C | Protein Kinase A, B (AKT), C |
| ROS | Reactive Oxygen Species |
| SCID | Severe Combined Immunodeficiency |
| SP1 | Multiple Specificity Protein 1, a Transcription Factor |
| SRSF1 | Serine-arginine Protein Splicing Factor-2/Alternate Splicing Factor |
| TNF-α | Tumor Necrosis Factor α |
| TNM | T—tumor; N—lymph nodes; M—metastasis |
| UTR | Untranslated Region |
| VEGF | Vascular Endothelial Growth Factor |
| wt | wild type |
| ZEB1 | Zinc Finger E-box-binding Homeobox 1 |
References
- Yu, H.; Rohan, T. Role of the insulin-like growth factor family in cancer development and progression. J. Natl. Cancer Inst. 2000, 92, 1472–1489. [Google Scholar] [CrossRef]
- Redwan, E.M.; Linjawi, M.H.; Uversky, V.N. Looking at the carcinogenicity of human insulin analogues via the intrinsic disorder prism. Sci. Rep. 2016, 6, 23320. [Google Scholar] [CrossRef] [Green Version]
- Weinstein, D.; Simon, M.; Yehezkel, E.; Laron, Z.; Werner, H. Insulin analogues display IGF-I-like mitogenic and anti-apoptotic activities in cultured cancer cells. Diabetes Metab. Res. Rev. 2009, 25, 41–49. [Google Scholar] [CrossRef]
- Rotwein, P. Two insulin-like growth factor I messenger RNAs are expressed in human liver. Proc. Natl. Acad. Sci. USA 1986, 83, 77–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pell, J.M.; Saunders, J.C.; Gilmour, R.S. Differential regulation of transcription initiation from insulin-like growth factor-I (IGF-I) leader exons and of tissue IGF-I expression in response to changed growth hormone and nutritional status in sheep. Endocrinology 1993, 132, 1797–1807. [Google Scholar] [CrossRef]
- Rotwein, P. Mapping the growth hormone--Stat5b--IGF-I transcriptional circuit. Trends Endocrinol. Metab. 2012, 23, 186–193. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, A.; Alzhanov, D.; Rotwein, P. Defining human insulin-like growth factor I gene regulation. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E519–E529. [Google Scholar] [CrossRef]
- Le Roith, D.; Bondy, C.; Yakar, S.; Liu, J.L.; Butler, A. The somatomedin hypothesis: 2001. Endocr. Rev. 2001, 22, 53–74. [Google Scholar] [CrossRef]
- Barton, E.R. The ABCs of IGF-I isoforms: Impact on muscle hypertrophy and implications for repair. Appl. Physiol. Nutr. Metab. 2006, 31, 791–797. [Google Scholar] [CrossRef]
- Barton, E.R.; DeMeo, J.; Lei, H. The insulin-like growth factor (IGF)-I E-peptides are required for isoform-specific gene expression and muscle hypertrophy after local IGF-I production. J. Appl. Physiol. 2010, 108, 1069–1076. [Google Scholar] [CrossRef] [Green Version]
- Kasprzak, A.; Kwasniewski, W.; Adamek, A.; Gozdzicka-Jozefiak, A. Insulin-like growth factor (IGF) axis in cancerogenesis. Mutat. Res. Rev. Mutat. Res. 2017, 772, 78–104. [Google Scholar] [CrossRef] [PubMed]
- Durzyńska, J.; Philippou, A.; Brisson, B.K.; Nguyen-McCarty, M.; Barton, E.R. The pro-forms of insulin-like growth factor I (IGF-I) are predominant in skeletal muscle and alter IGF-I receptor activation. Endocrinology 2013, 154, 1215–1224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakuno, F.; Takahashi, S.I. IGF1 receptor signaling pathways. J. Mol. Endocrinol. 2018, 61, T69–T86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philippou, A.; Armakolas, A.; Panteleakou, Z.; Pissimissis, N.; Nezos, A.; Theos, A.; Kaparelou, M.; Armakolas, N.; Pneumaticos, S.G.; Koutsilieris, M. IGF1Ec expression in MG-63 human osteoblast-like osteosarcoma cells. Anticancer Res. 2011, 31, 4259–4265. [Google Scholar]
- Shang, J.; Fan, X.; Liu, H. The role of mechano-growth factor E peptide in the regulation of osteosarcoma. Oncol. Lett. 2015, 10, 697–704. [Google Scholar] [CrossRef] [Green Version]
- De Santi, M.; Annibalini, G.; Barbieri, E.; Villarini, A.; Vallorani, L.; Contarelli, S.; Berrino, F.; Stocchi, V.; Brandi, G. Human IGF1 pro-forms induce breast cancer cell proliferation via the IGF1 receptor. Cell Oncol. 2016, 39, 149–159. [Google Scholar] [CrossRef]
- Armakolas, A.; Dimakakos, A.; Loukogiannaki, C.; Armakolas, N.; Antonopoulos, A.; Florou, C.; Tsioli, P.; Papageorgiou, E.; Alexandrou, T.P.; Stathaki, M.; et al. IL-6 is associated to IGF-1Ec upregulation and Ec peptide secretion, from prostate tumors. Mol. Med. 2018, 24, 6. [Google Scholar] [CrossRef]
- Papageorgiou, E.; Philippou, A.; Armakolas, A.; Christopoulos, P.F.; Dimakakos, A.; Koutsilieris, M. The human Ec peptide: The active core of a progression growth factor with species-specific mode of action. Hormones 2016, 15, 423–434. [Google Scholar] [CrossRef]
- Yang, H.; Adamo, M.L.; Koval, A.P.; McGuinness, M.C.; Ben-Hur, H.; Yang, Y.; LeRoith, D.; Roberts, C.T., Jr. Alternative leader sequences in insulin-like growth factor I mRNAs modulate translational efficiency and encode multiple signal peptides. Mol. Endocrinol. 1995, 9, 1380–1395. [Google Scholar] [CrossRef] [Green Version]
- Matheny, R.W., Jr.; Nindl, B.C.; Adamo, M.L. Minireview: Mechano-growth factor: A putative product of IGF-I gene expression involved in tissue repair and regeneration. Endocrinology 2010, 151, 865–875. [Google Scholar] [CrossRef] [Green Version]
- Oberbauer, A.M. The Regulation of IGF-1 Gene Transcription and Splicing During Development and Aging. Front. Endocrinol. 2013, 4, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brisson, B.K.; Barton, E.R. New Modulators for IGF-I Activity within IGF-I Processing Products. Front. Endocrinol. 2013, 4, 42. [Google Scholar] [CrossRef] [Green Version]
- Philippou, A.; Maridaki, M.; Pneumaticos, S.; Koutsilieris, M. The complexity of the IGF1 gene splicing, posttranslational modification and bioactivity. Mol. Med. 2014, 20, 202–214. [Google Scholar] [CrossRef] [PubMed]
- Shavlakadze, T.; Winn, N.; Rosenthal, N.; Grounds, M.D. Reconciling data from transgenic mice that overexpress IGF-I specifically in skeletal muscle. Growth Horm. IGF Res. 2005, 15, 4–18. [Google Scholar] [CrossRef]
- Pohl, M.; Bortfeldt, R.H.; Grützmann, K.; Schuster, S. Alternative splicing of mutually exclusive exons—A review. Biosystems 2013, 114, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Tobin, G.; Yee, D.; Brünner, N.; Rotwein, P. A novel human insulin-like growth factor I messenger RNA is expressed in normal and tumor cells. Mol. Endocrinol. 1990, 4, 1914–1920. [Google Scholar] [CrossRef] [PubMed]
- Rotwein, P. Diversification of the insulin-like growth factor 1 gene in mammals. PLoS ONE 2017, 12, e0189642. [Google Scholar] [CrossRef]
- Rotwein, P. Variation in the Insulin-Like Growth Factor 1 Gene in Primates. Endocrinology 2017, 158, 804–814. [Google Scholar] [CrossRef] [Green Version]
- Smith, P.J.; Spurrell, E.L.; Coakley, J.; Hinds, C.J.; Ross, R.J.; Krainer, A.R.; Chew, S.L. An exonic splicing enhancer in human IGF-I pre-mRNA mediates recognition of alternative exon 5 by the serine-arginine protein splicing factor-2/alternative splicing factor. Endocrinology 2002, 143, 146–154. [Google Scholar] [CrossRef]
- Philippou, A.; Armakolas, A.; Koutsilieris, M. Evidence for the Possible Biological Significance of the igf-1 Gene Alternative Splicing in Prostate Cancer. Front. Endocrinol. 2013, 4, 31. [Google Scholar] [CrossRef] [Green Version]
- Durzyńska, J.; Barton, E. IGF expression in HPV-related and HPV-unrelated human cancer cells. Oncol. Rep. 2014, 32, 893–900. [Google Scholar] [CrossRef] [Green Version]
- Alexandraki, K.I.; Philippou, A.; Boutzios, G.; Kyriapoulos, G.; Xydakis, A.; Papaioannou, T.G.; Kolomodi, D.; Kaltsas, G.; Grossman, A.B. IGF-IEc expression is increased in secondary compared to primary foci in neuroendocrine neoplasms. Oncotarget 2017, 8, 79003–79011. [Google Scholar] [CrossRef]
- Mourmouras, N.; Philippou, A.; Christopoulos, P.; Kostoglou, K.; Grivaki, C.; Konstantinidis, C.; Serafetinides, E.; Delakas, D.; Koutsilieris, M. Differential Expression of IGF-I Transcripts in Bladder Cancer. Anticancer Res. 2018, 38, 3453–3459. [Google Scholar] [CrossRef]
- Alagaratnam, S.; Yang, S.Y.; Loizidou, M.; Fuller, B.; Ramesh, B. Mechano-Growth Factor Expression in Colorectal Cancer Investigated with Fluorescent Gold Nanoparticles. Anticancer Res. 2019, 39, 1705–1710. [Google Scholar] [CrossRef]
- Karagiannis, A.K.; Philippou, A.; Tseleni-Balafouta, S.; Zevolis, E.; Nakouti, T.; Tsopanomichalou-Gklotsou, M.; Psarras, V.; Koutsilieris, M. IGF-IEc Expression Is Associated with Advanced Differentiated Thyroid Cancer. Anticancer Res. 2019, 39, 2811–2819. [Google Scholar] [CrossRef]
- Black, D.L. Protein diversity from alternative splicing: A challenge for bioinformatics and post-genome biology. Cell 2000, 103, 367–370. [Google Scholar] [CrossRef] [Green Version]
- Roy, B.; Haupt, L.M.; Griffiths, L.R. Review: Alternative Splicing (AS) of Genes as an Approach for Generating Protein Complexity. Curr. Genom. 2013, 14, 182–194. [Google Scholar] [CrossRef] [Green Version]
- Fiszbein, A.; Kornblihtt, A.R. Alternative splicing switches: Important players in cell differentiation. Bioessays 2017, 39. [Google Scholar] [CrossRef]
- Tress, M.L.; Abascal, F.; Valencia, A. Alternative Splicing May Not Be the Key to Proteome Complexity. Trends Biochem. Sci. 2017, 42, 98–110. [Google Scholar] [CrossRef] [Green Version]
- Tress, M.L.; Abascal, F.; Valencia, A. Most Alternative Isoforms Are Not Functionally Important. Trends Biochem. Sci. 2017, 42, 408–410. [Google Scholar] [CrossRef] [Green Version]
- Singh, B.; Eyras, E. The role of alternative splicing in cancer. Transcription 2017, 8, 91–98. [Google Scholar] [CrossRef] [Green Version]
- Urbanski, L.M.; Leclair, N.; Anczuków, O. Alternative-splicing defects in cancer: Splicing regulators and their downstream targets, guiding the way to novel cancer therapeutics. Wiley Interdiscip. Rev. RNA 2018, 9, e1476. [Google Scholar] [CrossRef] [PubMed]
- Blencowe, B.J. Alternative splicing: New insights from global analyses. Cell 2006, 126, 37–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Montiel, N.; Rosas-Murrieta, N.H.; Anaya Ruiz, M.; Monjaraz-Guzman, E.; Martinez-Contreras, R. Alternative Splicing as a Target for Cancer Treatment. Int. J. Mol. Sci. 2018, 19, 545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- David, C.J.; Manley, J.L. Alternative pre-mRNA splicing regulation in cancer: Pathways and programs unhinged. Genes Dev. 2010, 24, 2343–2364. [Google Scholar] [CrossRef] [Green Version]
- Anczuków, O.; Krainer, A.R. Splicing-factor alterations in cancers. RNA 2016, 22, 1285–1301. [Google Scholar] [CrossRef] [Green Version]
- Climente-González, H.; Porta-Pardo, E.; Godzik, A.; Eyras, E. The Functional Impact of Alternative Splicing in Cancer. Cell Rep. 2017, 20, 2215–2226. [Google Scholar] [CrossRef] [Green Version]
- Farina, A.R.; Cappabianca, L.; Sebastiano, M.; Zelli, V.; Guadagni, S.; Mackay, A.R. Hypoxia-induced alternative splicing: The 11th Hallmark of Cancer. J. Exp. Clin. Cancer Res. 2020, 39, 110. [Google Scholar] [CrossRef]
- Koczorowska, M.M.; Kwasniewska, A.; Gozdzicka-Jozefiak, A. IGF1 mRNA isoform expression in the cervix of HPV-positive women with pre-cancerous and cancer lesions. Exp. Ther. Med. 2011, 2, 149–156. [Google Scholar] [CrossRef]
- Höppener, J.W.; de Pagter-Holthuizen, P.; Geurts van Kessel, A.H.; Jansen, M.; Kittur, S.D.; Antonarakis, S.E.; Lips, C.J.; Sussenbach, J.S. The human gene encoding insulin-like growth factor I is located on chromosome 12. Hum. Genet. 1985, 69, 157–160. [Google Scholar] [CrossRef]
- Gene: IGF1. Available online: www.ensemble.org (accessed on 3 August 2020).
- Jansen, M.; van Schaik, F.M.; Ricker, A.T.; Bullock, B.; Woods, D.E.; Gabbay, K.H.; Nussbaum, A.L.; Sussenbach, J.S.; Van den Brande, J.L. Sequence of cDNA encoding human insulin-like growth factor I precursor. Nature 1983, 306, 609–611. [Google Scholar] [CrossRef] [PubMed]
- Sussenbach, J.S.; Steenbergh, P.H.; Jansen, E.; Holthuizen, P.; Meinsma, D.; van Dijk, M.A.; Gloudemans, T. Structural and regulatory aspects of the human genes encoding IGF-I and -II. Adv. Exp. Med. Biol. 1991, 293, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Wallis, M. Characterization of a novel alternatively-spliced 5′ exon in the human insulin-like growth factor I (IGF-I) gene, expressed in liver and some cancers. Growth Horm. IGF Res. 2019, 46–47, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Adamo, M.L.; Ben-Hur, H.; LeRoith, D.; Roberts, C.T., Jr. Transcription initiation in the two leader exons of the rat IGF-I gene occurs from disperse versus localized sites. Biochem. Biophys. Res. Commun. 1991, 176, 887–893. [Google Scholar] [CrossRef]
- Jansen, E.; Steenbergh, P.H.; LeRoith, D.; Roberts, C.T., Jr.; Sussenbach, J.S. Identification of multiple transcription start sites in the human insulin-like growth factor-I gene. Mol. Cell Endocrinol. 1991, 78, 115–125. [Google Scholar] [CrossRef]
- Hall, L.J.; Kajimoto, Y.; Bichell, D.; Kim, S.W.; James, P.L.; Counts, D.; Nixon, L.J.; Tobin, G.; Rotwein, P. Functional analysis of the rat insulin-like growth factor I gene and identification of an IGF-I gene promoter. DNA Cell Biol. 1992, 11, 301–313. [Google Scholar] [CrossRef]
- Steenbergh, P.H.; Jansen, E.; van Schaik, F.M.; Sussenbach, J.S. Functional analysis of the human IGF-I gene promoters. Mol. Reprod. Dev. 1993, 35, 365–367. [Google Scholar] [CrossRef]
- Mittanck, D.W.; Kim, S.W.; Rotwein, P. Essential promoter elements are located within the 5′ untranslated region of human insulin-like growth factor-I exon I. Mol. Cell Endocrinol. 1997, 126, 153–163. [Google Scholar] [CrossRef]
- Wang, X.; Talamantez, J.L.; Adamo, M.L. A CACCC box in the proximal exon 2 promoter of the rat insulin-like growth factor I gene is required for basal promoter activity. Endocrinology 1998, 139, 1054–1066. [Google Scholar] [CrossRef]
- Wang, L.; Wang, X.; Adamo, M.L. Two putative GATA motifs in the proximal exon 1 promoter of the rat insulin-like growth factor I gene regulate basal promoter activity. Endocrinology 2000, 141, 1118–1126. [Google Scholar] [CrossRef]
- Krishna, A.Y.; Pao, C.I.; Thulé, P.M.; Villafuerte, B.C.; Phillips, L.S. Transcription initiation of the rat insulin-like growth factor-I gene in hepatocyte primary culture. J. Endocrinol. 1996, 151, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Ohtsuki, T.; Otsuki, M.; Murakami, Y.; Maekawa, T.; Yamamoto, T.; Akasaka, K.; Takeuchi, S.; Takahashi, S. Organ-specific and age-dependent expression of insulin-like growth factor-I (IGF-I) mRNA variants: IGF-IA and IB mRNAs in the mouse. Zoolog. Sci. 2005, 22, 1011–1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotwein, P. Quantifying Promoter-Specific Insulin-Like Growth Factor 1 Gene Expression by Interrogating Public Databases. Physiol. Rep. 2019, 7, e13970. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.W.; Lajara, R.; Rotwein, P. Structure and function of a human insulin-like growth factor-I gene promoter. Mol. Endocrinol. 1991, 5, 1964–1972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen, E.; Steenbergh, P.H.; van Schaik, F.M.; Sussenbach, J.S. The human IGF-I gene contains two cell type-specifically regulated promoters. Biochem. Biophys. Res. Commun. 1992, 187, 1219–1226. [Google Scholar] [CrossRef]
- Chia, D.J.; Young, J.J.; Mertens, A.R.; Rotwein, P. Distinct alterations in chromatin organization of the two IGF-I promoters precede growth hormone-induced activation of IGF-I gene transcription. Mol. Endocrinol. 2010, 24, 779–789. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, D.C.; Szestak, T.A.; Pell, J.M. Regulation of IGF-I mRNA by GH: Putative functions for class 1 and 2 message. Am. J. Physiol. Endocrinol. Metab. 2002, 283, E251–E258. [Google Scholar] [CrossRef]
- Wang, Y.; Price, S.E.; Jiang, H. Cloning and characterization of the bovine class 1 and class 2 insulin-like growth factor-I mRNAs. Domest. Anim. Endocrinol. 2003, 25, 315–328. [Google Scholar] [CrossRef]
- Lin, W.W.; Oberbauer, A.M. Alternative splicing of insulin-like growth factor I mRNA is developmentally regulated in the rat and mouse with preferential exon 2 usage in the mouse. Growth Horm. IGF Res. 1998, 8, 225–233. [Google Scholar] [CrossRef]
- McCarthy, T.L.; Yun, Z.; Madri, J.A.; Centrella, M. Stratified control of IGF-I expression by hypoxia and stress hormones in osteoblasts. Gene 2014, 539, 141–151. [Google Scholar] [CrossRef] [Green Version]
- Bell, G.I.; Stempien, M.M.; Fong, N.M.; Rall, L.B. Sequences of liver cDNAs encoding two different mouse insulin-like growth factor I precursors. Nucleic Acids Res. 1986, 14, 7873–7882. [Google Scholar] [CrossRef] [PubMed]
- Rotwein, P.; Pollock, K.M.; Didier, D.K.; Krivi, G.G. Organization and sequence of the human insulin-like growth factor I gene. Alternative RNA processing produces two insulin-like growth factor I precursor peptides. J. Biol. Chem. 1986, 261, 4828–4832. [Google Scholar] [PubMed]
- Temmerman, L.; Slonimsky, E.; Rosenthal, N. Class 2 IGF-1 isoforms are dispensable for viability, growth and maintenance of IGF-1 serum levels. Growth Horm. IGF Res. 2010, 20, 255–263. [Google Scholar] [CrossRef]
- Chew, S.L.; Lavender, P.; Clark, A.J.; Ross, R.J. An alternatively spliced human insulin-like growth factor-I transcript with hepatic tissue expression that diverts away from the mitogenic IBE1 peptide. Endocrinology 1995, 136, 1939–1944. [Google Scholar] [CrossRef]
- Nagaoka, I.; Someya, A.; Iwabuchi, K.; Yamashita, T. Expression of insulin-like growth factor-IA and factor-IB mRNA in human liver, hepatoma cells, macrophage-like cells and fibroblasts. FEBS Lett. 1991, 280, 79–83. [Google Scholar] [CrossRef] [Green Version]
- Simmons, J.G.; Van Wyk, J.J.; Hoyt, E.C.; Lund, P.K. Multiple transcription start sites in the rat insulin-like growth factor-I gene give rise to IGF-I mRNAs that encode different IGF-I precursors and are processed differently in vitro. Growth Factors 1993, 9, 205–221. [Google Scholar] [CrossRef]
- Tahimic, C.G.; Wang, Y.; Bikle, D.D. Anabolic effects of IGF-1 signaling on the skeleton. Front. Endocrinol. 2013, 4, 6. [Google Scholar] [CrossRef] [Green Version]
- RCSB PDB. Available online: https://www.rcsb.org/structure/1IMX0 (accessed on 3 August 2020).
- Steenbergh, P.H.; Koonen-Reemst, A.M.; Cleutjens, C.B.; Sussenbach, J.S. Complete nucleotide sequence of the high molecular weight human IGF-I mRNA. Biochem. Biophys. Res. Commun. 1991, 175, 507–514. [Google Scholar] [CrossRef]
- Tan, D.S.; Cook, A.; Chew, S.L. Nucleolar localization of an isoform of the IGF-I precursor. BMC Cell Biol. 2002, 3, 17. [Google Scholar] [CrossRef]
- Lowe, W.L., Jr.; Lasky, S.R.; LeRoith, D.; Roberts, C.T., Jr. Distribution and regulation of rat insulin-like growth factor I messenger ribonucleic acids encoding alternative carboxyterminal E-peptides: Evidence for differential processing and regulation in liver. Mol. Endocrinol. 1988, 2, 528–535. [Google Scholar] [CrossRef]
- Wallis, M. New insulin-like growth factor (IGF)-precursor sequences from mammalian genomes: The molecular evolution of IGFs and associated peptides in primates. Growth Horm. IGF Res. 2009, 19, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Siegfried, J.M.; Kasprzyk, P.G.; Treston, A.M.; Mulshine, J.L.; Quinn, K.A.; Cuttitta, F. A mitogenic peptide amide encoded within the E peptide domain of the insulin-like growth factor IB prohormone. Proc. Natl. Acad. Sci. USA 1992, 89, 8107–8111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velloso, C.P.; Harridge, S.D. Insulin-like growth factor-I E peptides: Implications for aging skeletal muscle. Scand. J. Med. Sci. Sports. 2010, 20, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Brisson, B.K.; Barton, E.R. Insulin-like growth factor-I E-peptide activity is dependent on the IGF-I receptor. PLoS ONE 2012, 7, e45588. [Google Scholar] [CrossRef]
- Vassilakos, G.; Philippou, A.; Koutsilieris, M. Identification of the IGF-1 processing product human Ec/rodent Eb peptide in various tissues: Evidence for its differential regulation after exercise-induced muscle damage in humans. Growth Horm. IGF Res. 2017, 32, 22–28. [Google Scholar] [CrossRef]
- Annibalini, G.; Contarelli, S.; De Santi, M.; Saltarelli, R.; Di Patria, L.; Guescini, M.; Villarini, A.; Brandi, G.; Stocchi, V.; Barbieri, E. The intrinsically disordered E-domains regulate the IGF-1 prohormones stability, subcellular localisation and secretion. Sci. Rep. 2018, 8, 8919. [Google Scholar] [CrossRef]
- Pfeffer, L.A.; Brisson, B.K.; Lei, H.; Barton, E.R. The insulin-like growth factor (IGF)-I E-peptides modulate cell entry of the mature IGF-I protein. Mol. Biol. Cell. 2009, 20, 3810–3817. [Google Scholar] [CrossRef] [Green Version]
- Durzyńska, J.; Wardziński, A.; Koczorowska, M.; Goździcka-Józefiak, A.; Barton, E.R. Human Eb peptide: Not just a by-product of pre-pro-IGF1b processing? Horm. Metab. Res. 2013, 45, 415–422. [Google Scholar] [CrossRef] [Green Version]
- Poreba, E.; Durzynska, J. Nuclear localization and actions of the insulin-like growth factor 1 (IGF-1) system components: Transcriptional regulation and DNA damage response. Mutat. Res. 2020, 784, 108307. [Google Scholar] [CrossRef]
- Cheema, U.; Brown, R.; Mudera, V.; Yang, S.Y.; McGrouther, G.; Goldspink, G. Mechanical signals and IGF-I gene splicing in vitro in relation to development of skeletal muscle. J. Cell Physiol. 2005, 202, 67–75. [Google Scholar] [CrossRef]
- Rotwein, P.; Bichell, D.; Kikuchi, K. Multifactorial regulation of IGF-I gene expression. Mol. Reprod. Dev. 1993, 35, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Whitehead, R.E., Jr.; Underwood, L.E. Effect of fasting on insulin-like growth factor (IGF)-IA and IGF-IB messenger ribonucleic acids and prehormones in rat liver. Endocrinology 1997, 138, 3112–3118. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chrysis, D.; Underwood, L.E. Reduction of hepatic insulin-like growth factor I (IGF-I) messenger ribonucleic acid (mRNA) during fasting is associated with diminished splicing of IGF-I pre-mRNA and decreased stability of cytoplasmic IGF-I mRNA. Endocrinology 1998, 139, 4523–4530. [Google Scholar] [CrossRef]
- Lin, W.W.; Oberbauer, A.M. Spatiotemporal expression of alternatively spliced IGF-I mRNA in the rat costochondral growth plate. J. Endocrinol. 1999, 160, 461–467. [Google Scholar] [CrossRef] [Green Version]
- Chia, D.J. Minireview: Mechanisms of growth hormone-mediated gene regulation. Mol. Endocrinol. 2014, 28, 1012–1025. [Google Scholar] [CrossRef]
- Bichell, D.P.; Kikuchi, K.; Rotwein, P. Growth hormone rapidly activates insulin-like growth factor I gene transcription in vivo. Mol. Endocrinol. 1992, 6, 1899–1908. [Google Scholar] [CrossRef]
- Chia, D.J.; Varco-Merth, B.; Rotwein, P. Dispersed Chromosomal Stat5b-binding elements mediate growth hormone-activated insulin-like growth factor-I gene transcription. J. Biol. Chem. 2010, 285, 17636–17647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, T.L.; Thomas, M.J.; Centrella, M.; Rotwein, P. Regulation of insulin-like growth factor I transcription by cyclic adenosine 3′,5′-monophosphate (cAMP) in fetal rat bone cells through an element within exon 1: Protein kinase A-dependent control without a consensus AMP response element. Endocrinology 1995, 136, 3901–3908. [Google Scholar] [CrossRef] [PubMed]
- Lowe, W.L., Jr.; Roberts, C.T., Jr.; Lasky, S.R.; LeRoith, D. Differential expression of alternative 5′ untranslated regions in mRNAs encoding rat insulin-like growth factor I. Proc. Natl. Acad. Sci. USA 1987, 84, 8946–8950. [Google Scholar] [CrossRef] [Green Version]
- Arkins, S.; Rebeiz, N.; Biragyn, A.; Reese, D.L.; Kelley, K.W. Murine macrophages express abundant insulin-like growth factor-I class I Ea and Eb transcripts. Endocrinology 1993, 133, 2334–2343. [Google Scholar] [CrossRef]
- Annibalini, G.; Bielli, P.; De Santi, M.; Agostini, D.; Guescini, M.; Sisti, D.; Contarelli, S.; Brandi, G.; Villarini, A.; Stocchi, V.; et al. MIR retroposon exonization promotes evolutionary variability and generates species-specific expression of IGF-1 splice variants. Biochim. Biophys. Acta. 2016, 1859, 757–768. [Google Scholar] [CrossRef] [PubMed]
- Kasprzak, A.; Szaflarski, W.; Szmeja, J.; Andrzejewska, M.; Przybyszewska, W.; Koczorowska, M.; Drews, M.; Kaczmarek, E. Expression of various insulin-like growth factor-1 mRNA isoforms in colorectal cancer. Contemp. Oncol. 2012, 6, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Kasprzak, A.; Szaflarski, W.; Szmeja, J.; Andrzejewska, M.; Przybyszewska, W.; Kaczmarek, E.; Koczorowska, M.; Kościński, T.; Zabel, M.; Drews, M. Differential expression of IGF-1 mRNA isoforms in colorectal carcinoma and normal colon tissue. Int. J. Oncol. 2013, 42, 305–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christopoulos, P.F.; Philippou, A.; Koutsilieris, M. Pattern of IGF-1 variants’ expression in human cancer cell lines using a novel q-RT-PCR approach. Anticancer Res. 2015, 5, 107–115. [Google Scholar]
- Milingos, D.S.; Philippou, A.; Armakolas, A.; Papageorgiou, E.; Sourla, A.; Protopapas, A.; Liapi, A.; Antsaklis, A.; Mastrominas, M.; Koutsilieris, M. Insulinlike growth factor-1Ec (MGF) expression in eutopic and ectopic endometrium: Characterization of the MGF E-peptide actions in vitro. Mol. Med. 2011, 17, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Armakolas, A.; Philippou, A.; Panteleakou, Z.; Nezos, A.; Sourla, A.; Petraki, C.; Koutsilieris, M. Preferential expression of IGF-1Ec (MGF) transcript in cancerous tissues of human prostate: Evidence for a novel and autonomous growth factor activity of MGF E peptide in human prostate cancer cells. Prostate 2010, 70, 1233–1242. [Google Scholar] [CrossRef] [PubMed]
- Savvani, A.; Petraki, C.; Msaouel, P.; Diamanti, E.; Xoxakos, I.; Koutsilieris, M. IGF-IEc expression is associated with advanced clinical and pathological stage of prostate cancer. Anticancer Res. 2013, 33, 2441–2445. [Google Scholar]
- Semenova, E.; Koegel, H.; Hasse, S.; Klatte, J.E.; Slonimsky, E.; Bilbao, D.; Paus, R.; Werner, S.; Rosenthal, N. Overexpression of mIGF-1 in keratinocytes improves wound healing and accelerates hair follicle formation and cycling in mice. Am. J. Pathol. 2008, 173, 1295–1310. [Google Scholar] [CrossRef] [Green Version]
- Gatto, M.; Drudi-Metalli, V.; Torrice, A.; Alpini, G.; Cantafora, A.; Blotta, I.; Alvaro, D. Insulin-like growth factor-1 isoforms in rat hepatocytes and cholangiocytes and their involvement in protection against cholestatic injury. Lab. Investig. 2008, 88, 986–994. [Google Scholar] [CrossRef] [Green Version]
- Kasprzak, A.; Adamek, A.; Przybyszewska, W.; Pyda, P.; Szmeja, J.; Seraszek-Jaros, A.; Lanzafame, A.; Surdacka, A.; Mozer-Lisewska, I.; Koczorowska, M. Insulin-like growth factor-1 mRNA isoforms and insulin-like growth factor-1 receptor mRNA expression in chronic hepatitis C. World J. Gastroenterol. 2015, 21, 3867–3875. [Google Scholar] [CrossRef]
- Hunziker, E.B.; Wagner, J.; Zapf, J. Differential effects of insulin-like growth factor I and growth hormone on developmental stages of rat growth plate chondrocytes in vivo. J. Clin. Investig. 1994, 93, 1078–1086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazowski, D.A.; Fraher, L.J.; Hodsman, A.; Steer, B.; Modrowski, D.; Han, V.K. Regional variation of insulin-like growth factor-I gene expression in mature rat bone and cartilage. Bone 1994, 15, 563–576. [Google Scholar] [CrossRef]
- Reinecke, M.; Schmid, A.C.; Heyberger-Meyer, B.; Hunziker, E.B.; Zapf, J. Effect of growth hormone and insulin-like growth factor I (IGF-I) on the expression of IGF-I messenger ribonucleic acid and peptide in rat tibial growth plate and articular chondrocytes in vivo. Endocrinology 2000, 141, 2847–2853. [Google Scholar] [CrossRef] [PubMed]
- Govoni, K.E.; Lee, S.K.; Chung, Y.S.; Behringer, R.R.; Wergedal, J.E.; Baylink, D.J.; Mohan, S. Disruption of insulin-like growth factor-I expression in type IIalphaI collagen-expressing cells reduces bone length and width in mice. Physiol. Genom. 2007, 30, 354–362. [Google Scholar] [CrossRef]
- Reijnders, C.M.; Bravenboer, N.; Tromp, A.M.; Blankenstein, M.A.; Lips, P. Effect of mechanical loading on insulin-like growth factor-I gene expression in rat tibia. J. Endocrinol. 2007, 192, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Armakolas, N.; Armakolas, A.; Antonopoulos, A.; Dimakakos, A.; Stathaki, M.; Koutsilieris, M. The role of the IGF-1 Ec in myoskeletal system and osteosarcoma pathophysiology. Crit. Rev. Oncol. Hematol. 2016, 108, 137–145. [Google Scholar] [CrossRef]
- Laugero, L.A.; Oberbauer, A.M. Stability of alternatively spliced IGF-I mRNA in growth plate chondrocytes. Connect. Tissue Res. 2000, 41, 165–174. [Google Scholar] [CrossRef]
- Tang, L.L.; Xian, C.Y.; Wang, Y.L. The MGF expression of osteoblasts in response to mechanical overload. Arch. Oral Biol. 2006, 51, 1080–1085. [Google Scholar] [CrossRef]
- Xin, C.; Bingbing, Z.; Yuanliang, W.; Chengyu, X.; Li, Y.; Moyuan, D.; Qin, P.; Yuxiao, L. Mechano-growth factor E peptide inhibits the differentiation and mineralization of osteoblasts. Arch. Oral Biol. 2012, 57, 720–727. [Google Scholar] [CrossRef]
- Cui, H.; Yi, Q.; Feng, J.; Yang, L.; Tang, L. Mechano growth factor E peptide regulates migration and differentiation of bone marrow mesenchymal stem cells. J. Mol. Endocrinol. 2014, 52, 111–120. [Google Scholar] [CrossRef] [Green Version]
- Schlegel, W.; Raimann, A.; Halbauer, D.; Scharmer, D.; Sagmeister, S.; Wessner, B.; Helmreich, M.; Haeusler, G.; Egerbacher, M. Insulin-like growth factor I (IGF-1) Ec/Mechano Growth factor--A splice variant of IGF-1 within the growth plate. PLoS ONE 2013, 8, e76133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armakolas, N.; Dimakakos, A.; Armakolas, A.; Antonopoulos, A.; Koutsilieris, M. Possible role of the Ec peptide of IGF-1Ec in cartilage repair. Mol. Med. Rep. 2016, 14, 3066–3072. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.S.; Kessler, C.B.; Shenoy, V.; Rosen, C.J.; Delany, A.M. IGF-I 3′ untranslated region: Strain-specific polymorphisms and motifs regulating IGF-I in osteoblasts. Endocrinology 2013, 154, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.; Kelly, B.J.; Wang, C.; Klingler, K.; Chan, A.; Eckert, G.J.; Trippel, S.B. Human IGF-I propeptide A promotes articular chondrocyte biosynthesis and employs glycosylation-dependent heparin binding. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 567–575. [Google Scholar] [CrossRef]
- Wei, W.; Liu, S.; Song, J.; Feng, T.; Yang, R.; Cheng, Y.; Li, H.; Hao, L. MGF-19E peptide promoted proliferation, differentiation and mineralization of MC3T3-E1 cell and promoted bone defect healing. Gene 2020, 749, 144703. [Google Scholar] [CrossRef]
- Roberston, M.J.; Raghunathan, S.; Potaman, V.N.; Zhang, F.; Stewart, M.D.; McConnell, B.K.; Schwartz, R.J. CRISPR-Cas9-induced IGF1 gene activation as a tool for enhancing muscle differentiation via multiple isoform expression. FASEB J. 2020, 34, 555–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vassilakos, G.; Lei, H.; Yang, Y.; Puglise, J.; Matheny, M.; Durzynska, J.; Ozery, M.; Bennett, K.; Spradlin, R.; Bonanno, H.; et al. Deletion of muscle IGF-I transiently impairs growth and progressively disrupts glucose homeostasis in male mice. FASEB J. 2019, 33, 181–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ascenzi, F.; Barberi, L.; Dobrowolny, G.; Villa Nova Bacurau, A.; Nicoletti, C.; Rizzuto, E.; Rosenthal, N.; Scicchitano, B.M.; Musarò, A. Effects of IGF-1 isoforms on muscle growth and sarcopenia. Aging Cell 2019, 18, e12954. [Google Scholar] [CrossRef]
- Yang, H.; Alnaqeeb, M.; Simpson, H.; Goldspink, G. Changes in muscle fibre type, muscle mass and IGF-I gene expression in rabbit skeletal muscle subjected to stretch. J. Anat. 1997, 190, 613–622. [Google Scholar] [CrossRef]
- McKoy, G.; Ashley, W.; Mander, J.; Yang, S.Y.; Williams, N.; Russell, B.; Goldspink, G. Expression of insulin growth factor-1 splice variants and structural genes in rabbit skeletal muscle induced by stretch and stimulation. J. Physiol. 1999, 516, 583–592. [Google Scholar] [CrossRef]
- Goldspink, G. Gene expression in skeletal muscle. Biochem. Soc. Trans. 2002, 30, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Philippou, A.; Maridaki, M.; Halapas, A.; Koutsilieris, M. The role of the insulin-like growth factor 1 (IGF-1) in skeletal muscle physiology. Vivo 2007, 21, 45–54. [Google Scholar]
- Hameed, M.; Orrell, R.W.; Cobbold, M.; Goldspink, G.; Harridge, S.D. Expression of IGF-I splice variants in young and old human skeletal muscle after high resistance exercise. J. Physiol. 2003, 547, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Barton, E.R. Viral expression of insulin-like growth factor-I isoforms promotes different responses in skeletal muscle. J. Appl. Physiol. 2006, 100, 1778–1784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Brisson, B.K.; Liu, M.; Spinazzola, J.M.; Barton, E.R. Mature IGF-I excels in promoting functional muscle recovery from disuse atrophy compared with pro-IGF-IA. J. Appl. Physiol. 2014, 116, 797–806. [Google Scholar] [CrossRef]
- Kandalla, P.K.; Goldspink, G.; Butler-Browne, G.; Mouly, V. Mechano Growth Factor E peptide (MGF-E), derived from an isoform of IGF-1, activates human muscle progenitor cells and induces an increase in their fusion potential at different ages. Mech. Ageing Dev. 2011, 132, 154–162. [Google Scholar] [CrossRef]
- Musarò, A.; McCullagh, K.; Paul, A.; Houghton, L.; Dobrowolny, G.; Molinaro, M.; Barton, E.R.; Sweeney, H.L.; Rosenthal, N. Localized Igf-1 transgene expression sustains hypertrophy and regeneration in senescent skeletal muscle. Nat. Genet. 2001, 27, 195–200. [Google Scholar] [CrossRef]
- Fornaro, M.; Hinken, A.C.; Needle, S.; Hu, E.; Trendelenburg, A.U.; Mayer, A.; Rosenstiel, A.; Chang, C.; Meier, V.; Billin, A.N.; et al. Mechano-growth factor peptide, the COOH terminus of unprocessed insulin-like growth factor 1, has no apparent effect on myoblasts or primary muscle stem cells. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E150–E156. [Google Scholar] [CrossRef] [Green Version]
- Philippou, A.; Papageorgiou, E.; Bogdanis, G.; Halapas, A.; Sourla, A.; Maridaki, M.; Pissimissis, N.; Koutsilieris, M. Expression of IGF-1 isoforms after exercise-induced muscle damage in humans: Characterization of the MGF E peptide actions in vitro. Vivo 2009, 23, 567–575. [Google Scholar]
- Yang, S.Y.; Goldspink, G. Different roles of the IGF-I Ec peptide (MGF) and mature IGF-I in myoblast proliferation and differentiation. FEBS Lett. 2002, 522, 156–160. [Google Scholar] [CrossRef] [Green Version]
- Ates, K.; Yang, S.Y.; Orrell, R.W.; Sinanan, A.C.; Simons, P.; Solomon, A.; Beech, S.; Goldspink, G.; Lewis, M.P. The IGF-I splice variant MGF increases progenitor cells in ALS, dystrophic, and normal muscle. FEBS Lett. 2007, 581, 2727–2732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, L.L.; Li, X.K.; Xu, J.; Mo, D.L.; Tong, X.; Pan, Z.C.; Li, J.Q.; Chen, Y.S.; Zhang, Z.; Wang, C.; et al. Mechano growth factor (MGF) promotes proliferation and inhibits differentiation of porcine satellite cells (PSCs) by down-regulation of key myogenic transcriptional factors. Mol. Cell Biochem. 2012, 370, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.; Wernig, A.; Goldspink, G. Muscle satellite (stem) cell activation during local tissue injury and repair. J. Anat. 2003, 203, 89–99. [Google Scholar] [CrossRef]
- Mills, P.; Lafrenière, J.F.; Benabdallah, B.F.; El Fahime, E.M.; Tremblay, J.P. A new pro-migratory activity on human myogenic precursor cells for a synthetic peptide within the E domain of the mechano growth factor. Exp. Cell Res. 2007, 313, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Imanaka, M.; Iida, K.; Murawaki, A.; Nishizawa, H.; Fukuoka, H.; Takeno, R.; Takahashi, Y.; Okimura, Y.; Kaji, H.; Chihara, K. Growth hormone stimulates mechano growth factor expression and activates myoblast transformation in C2C12 cells. Kobe J. Med. Sci. 2008, 54, E46–E54. [Google Scholar]
- Kravchenko, I.V.; Furalyov, V.A.; Popov, V.O. Stimulation of mechano-growth factor expression by myofibrillar proteins in murine myoblasts and myotubes. Mol. Cell Biochem. 2012, 363, 347–355. [Google Scholar] [CrossRef]
- Kravchenko, I.V.; Furalyov, V.A.; Chatziefthimiou, S.; Wilmanns, M.; Popov, V.O. Induction of insulin-like growth factor 1 splice forms by subfragments of myofibrillar proteins. Mol. Cell Endocrinol. 2015, 399, 69–77. [Google Scholar] [CrossRef]
- Brisson, B.K.; Spinazzola, J.; Park, S.; Barton, E.R. Viral expression of insulin-like growth factor I E-peptides increases skeletal muscle mass but at the expense of strength. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E965–E974. [Google Scholar] [CrossRef] [Green Version]
- Owino, V.; Yang, S.Y.; Goldspink, G. Age-related loss of skeletal muscle function and the inability to express the autocrine form of insulin-like growth factor-1 (MGF) in response to mechanical overload. FEBS Lett. 2001, 505, 259–263. [Google Scholar] [CrossRef]
- McKay, B.R.; O’Reilly, C.E.; Phillips, S.M.; Tarnopolsky, M.A.; Parise, G. Co-expression of IGF-1 family members with myogenic regulatory factors following acute damaging muscle-lengthening contractions in humans. J. Physiol. 2008, 586, 5549–5560. [Google Scholar] [CrossRef]
- Stavropoulou, A.; Halapas, A.; Sourla, A.; Philippou, A.; Papageorgiou, E.; Papalois, A.; Koutsilieris, M. IGF-1 expression in infarcted myocardium and MGF E peptide actions in rat cardiomyocytes in vitro. Mol. Med. 2009, 15, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Vinciguerra, M.; Santini, M.P.; Claycomb, W.C.; Ladurner, A.G.; Rosenthal, N. Local IGF-1 isoform protects cardiomyocytes from hypertrophic and oxidative stresses via SirT1 activity. Aging 2009, 2, 43–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mavrommatis, E.; Shioura, K.M.; Los, T.; Goldspink, P.H. The E-domain region of mechano-growth factor inhibits cellular apoptosis and preserves cardiac function during myocardial infarction. Mol. Cell Biochem. 2013, 381, 69–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shioura, K.; Pena, J.; Goldspink, P. Administration of a Synthetic Peptide Derived from the E-domain Region of Mechano-Growth Factor Delays Decompensation Following Myocardial Infarction. Int. J. Cardiovasc. Res. 2014, 3, 1000169. [Google Scholar] [CrossRef] [Green Version]
- Vassilakos, G.; Philippou, A.; Tsakiroglou, P.; Koutsilieris, M. Biological activity of the e domain of the IGF-1Ec as addressed by synthetic peptides. Hormones 2014, 13, 182–196. [Google Scholar] [CrossRef]
- Peña, J.R.; Pinney, J.R.; Ayala, P.; Desai, T.A.; Goldspink, P.H. Localized delivery of mechano-growth factor E-domain peptide via polymeric microstructures improves cardiac function following myocardial infarction. Biomaterials 2015, 46, 26–34. [Google Scholar] [CrossRef] [Green Version]
- Górecki, D.C.; Beresewicz, M.; Zabłocka, B. Neuroprotective Effects of Short Peptides Derived from the Insulin-Like Growth Factor 1. Neurochem. Int. 2007, 51, 451–458. [Google Scholar] [CrossRef]
- Lin, S.; Fan, L.W.; Pang, Y.; Rhodes, P.G.; Mitchell, H.J.; Cai, Z. IGF-1 protects oligodendrocyte progenitor cells and improves neurological functions following cerebral hypoxia-ischemia in the neonatal rat. Brain Res. 2005, 1063, 15–26. [Google Scholar] [CrossRef]
- Aperghis, M.; Johnson, I.P.; Cannon, J.; Yang, S.Y.; Goldspink, G. Different levels of neuroprotection by two insulin-like growth factor-I splice variants. Brain Res. 2004, 1009, 213–218. [Google Scholar] [CrossRef]
- Dluzniewska, J.; Sarnowska, A.; Beresewicz, M.; Johnson, I.; Srai, S.K.; Ramesh, B.; Goldspink, G.; Górecki, D.C.; Zabłocka, B. A strong neuroprotective effect of the autonomous C-terminal peptide of IGF-1 Ec (MGF) in brain ischemia. FASEB J. 2005, 19, 1896–1898. [Google Scholar] [CrossRef]
- Tang, J.J.; Podratz, J.L.; Lange, M.; Scrable, H.J.; Jang, M.H.; Windebank, A.J. Mechano growth factor, a splice variant of IGF-1, promotes neurogenesis in the aging mouse brain. Mol. Brain. 2017, 10, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quesada, A.; Ogi, J.; Schultz, J.; Handforth, A. C-terminal mechano-growth factor induces heme oxygenase-1-mediated neuroprotection of SH-SY5Y cells via the protein kinase Cϵ/Nrf2 pathway. J. Neurosci. Res. 2011, 89, 394–405, (published correction appears in J. Neurosci. Res. 2011, 89, 1142).. [Google Scholar] [CrossRef] [PubMed]
- Pickard, A.; Durzynska, J.; McCance, D.J.; Barton, E.R. The IGF axis in HPV associated cancers. Mutat Res. Rev. Mutat Res. 2017, 772, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Armakolas, A.; Kaparelou, M.; Dimakakos, A.; Papageorgiou, E.; Armakolas, N.; Antonopoulos, A.; Petraki, C.; Lekarakou, M.; Lelovas, P.; Stathaki, M.; et al. Oncogenic Role of the Ec Peptide of the IGF-1Ec Isoform in Prostate Cancer. Mol. Med. 2015, 21, 167–179. [Google Scholar] [CrossRef]
- Lo, J.H.; Chen, T.T. Production of bioactive recombinant human Eb-peptide of pro-IGF-I and identification of binding components from the plasma membrane of human breast cancer cells (MDA-MB-231). Exp. Cell Res. 2018, 362, 235–243. [Google Scholar] [CrossRef]
- Chen, M.J.; Chiou, P.P.; Lin, P.; Lin, C.M.; Siri, S.; Peck, K.; Chen, T.T. Suppression of growth and cancer-induced angiogenesis of aggressive human breast cancer cells (MDA-MB-231) on the chorioallantoic membrane of developing chicken embryos by E-peptide of pro-IGF-I. J. Cell Biochem. 2007, 101, 1316–1327. [Google Scholar] [CrossRef]
- Chen, M.J.; Kuo, Y.H.; Tian, X.C.; Chen, T.T. Novel biological activities of the fish pro-IGF-I E-peptides: Studies on effects of fish pro-IGF-I E-peptide on morphological change, anchorage-dependent cell division, and invasiveness in tumor cells. Gen. Comp. Endocrinol. 2002, 126, 342–351. [Google Scholar] [CrossRef]
- Christopoulos, P.F.; Papageorgiou, E.; Petraki, C.; Koutsilieris, M. The COOH-Terminus of the IGF-1Ec Isoform Enhances the Proliferation and Migration of Human MCF-7 Breast Cancer Cells. Anticancer Res. 2017, 37, 2899–2912. [Google Scholar] [CrossRef] [Green Version]
- Siri, S.; Chen, M.J.; Chen, T.T. Biological activity of rainbow trout Ea4-peptide of the pro-insulin-like growth factor (pro-IGF)-I on promoting attachment of breast cancer cells (MDA-MB-231) via alpha2- and beta1-integrin. J. Cell Biochem. 2006, 99, 1524–1535. [Google Scholar] [CrossRef]
- Siri, S.; Chen, M.J.; Chen, T.T. Inhibition of human breast cancer cell (MBA-MD-231) invasion by the Ea4-peptide of rainbow trout pro-IGF-I. J. Cell Biochem. 2006, 99, 1363–1373. [Google Scholar] [CrossRef]
- Yeh, Y.J.; Lin, C.M.; Chen, T.T. Human IGF-I Eb-peptide induces cell attachment and lamellipodia outspread of metastatic breast carcinoma cells (MDA-MB-231). Exp. Cell Res. 2017, 358, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Murphy, N.; Carreras-Torres, R.; Song, M.; Martin, R.M.; Tsilidis, K.K.; Smith-Byrne, K.; Fensom, G.; Perez-Cornago, A.; Travis, R.C.; Key, T.J.; et al. Circulating Levels of Insulin-Like Growth Factor 1 and Insulin-Like Growth Factor Binding Protein 3 Associate with Risk of Colorectal Cancer Based on Serologic and Mendelian Randomization Analyses. Gastroenterology 2020, 158, 1300–1312.e20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, N.; Oshima, T.; Yoshihara, K.; Aoyama, T.; Hayashi, T.; Yamada, T.; Sato, T.; Shiozawa, M.; Yoshikawa, T.; Morinaga, S.; et al. Clinicopathological significance and impact on outcomes of the gene expression levels of IGF-1, IGF-2 and IGF-1R, IGFBP-3 in patients with colorectal cancer: Overexpression of the IGFBP-3 gene is an effective predictor of outcomes in patients with colorectal cancer. Oncol. Lett. 2017, 13, 3958–3966. [Google Scholar] [CrossRef] [PubMed]
- Andrzejewska, M. Tkankowa Ekspresja Insulinopodobnego Czynnika Wzrostu 1 (IGF-1) w Nowotworach Jelita Grubego; In Polish; Poznan University of Medical Sciences: Poznan, Poland, 2015; Available online: http://www.wbc.poznan.pl/Content/393686/index.pdf (accessed on 8 July 2020).
- Zur Hausen, H. Papillomaviruses in the causation of human cancers—A brief historical account. Virology 2009, 384, 260–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doorbar, J. Model systems of human papillomavirus-associated disease. J. Pathol. 2016, 238, 166–179. [Google Scholar] [CrossRef] [Green Version]
- Durzyńska, J. IGF axis and other factors in HPV-related and HPV-unrelated carcinogenesis (review). Oncol. Rep. 2014, 32, 2295–2306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamek, A.; Kasprzak, A. Insulin-Like Growth Factor (IGF) System in Liver Diseases. Int. J. Mol. Sci. 2018, 19, 1308. [Google Scholar] [CrossRef] [Green Version]
- Zinevich, L.S.; Goncharova, N.O.; Uruvaeva, I.V.; Delone, G.V.; Mikaelian, A.S. Igf-1 and Its Isoform Expression in Hepatic Cell Tumors and the Surrounding Tissue in Mice Liver Carcinogenesis Induced by Diethylnitrozamine. Izv. Akad. Nauk. Ser. Biol. 2013, 6, 673–681, (Article in Russian). [Google Scholar]
- Pal, S.; Yadav, P.; Sainis, K.B.; Shankar, B.S. TNF-α and IGF-1 differentially modulate ionizing radiation responses of lung cancer cell lines. Cytokine 2018, 101, 89–98. [Google Scholar] [CrossRef]
- Sandberg-Nordqvist, A.C.; Ståhlbom, P.A.; Reinecke, M.; Collins, V.P.; von Holst, H.; Sara, V. Characterization of insulin-like growth factor 1 in human primary brain tumors. Cancer Res. 1993, 53, 2475–2478. [Google Scholar]
- Kuo, Y.H.; Chen, T.T. Novel activities of pro-IGF-I E peptides: Regulation of morphological differentiation and anchorage-independent growth in human neuroblastoma cells. Exp. Cell Res. 2002, 280, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Fagin, J.A.; Fernandez-Mejia, C.; Melmed, S. Pituitary insulin-like growth factor-I gene expression: Regulation by triiodothyronine and growth hormone. Endocrinology 1989, 25, 2385–2391. [Google Scholar] [CrossRef] [PubMed]
- Brahmkhatri, V.P.; Prasanna, C.; Atreya, H.S. Insulin-like growth factor system in cancer: Novel targeted therapies. Biomed. Res. Int. 2015, 2015, 538019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philippou, A.; Christopoulos, P.F.; Koutsilierism, D.M. Clinical studies in humans targeting the various components of the IGF system show lack of efficacy in the treatment of cancer. Mutat. Res. Rev. Mutat Res. 2017, 772, 105–122. [Google Scholar] [CrossRef] [PubMed]
- Salisbury, T.B.; Tomblin, J.K. Insulin/Insulin-like growth factors in cancer: New roles for the aryl hydrocarbon receptor, tumor resistance mechanisms, and new blocking strategies. Front. Endocrinol. 2015, 6, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, A.; Petnga, W.; Macaulay, V.M.; Weyer-Czernilofsky, U.; Bogenrieder, T. Insulin-Like Growth Factor (IGF) Pathway Targeting in Cancer: Role of the IGF Axis and Opportunities for Future Combination Studies. Target. Oncol. 2017, 12, 571–597. [Google Scholar] [CrossRef] [Green Version]
- Black, A.J.; Gamarra, J.R.; Giudice, J. More than a messenger: Alternative splicing as a therapeutic target. Biochim. Biophys. Acta Gene Regul. Mech. 2019, 1862, 194395. [Google Scholar] [CrossRef]


{kind=link}
| Human Cancer | Tissue Material/Technique | IGF1 mRNAs | IGF1 Propeptides/Peptides | No of ref. | ||||
|---|---|---|---|---|---|---|---|---|
| IGF1Ea (Class I and II) | IGF1Eb (Class I and II) | IGF1Ec (Class I and II) | Pro-Ea/Ea | Pro-Eb/Eb | Pro-Ec/Ec (MGF) | |||
| Brain cancer | n = 9 gliomas; IHC, RT-PCR | (+) | (+) | (-) | nt | [183] | ||
| Colorectal cancer | One colonic and one retro-sigmoidal NENs; IHC | nt | nt | (-) Ec peptide in both CRC | [32] | |||
| n = 13 pairs of CRC/CT; IHC; qRT-PCR | (+) ↑ vs. CT; ↑ vs. Eb&Ec; | (+) ↓ vs. CT; ↑ vs. Ec | (+) ≈ vs. CT | nt | [104] | |||
| ↓ class I/II vs. CT; ↑ class II vs. class I; | ||||||||
| n = 28 pairs of CRC/CT; IHC; qRT-PCR | (+); 82% of all transcripts; ≈ vs. Eb; ↑ vs. Ec | (+); 17% of all transcripts; ↑ vs. Ec | (+); ~1% of all transcripts | nt | [105] | |||
| ↓ all isoforms vs. CT; CRC: Class I - 59%; class II - 41%, in quantitative expression - NS | ||||||||
| CRC/polyps/CT; fluorescent gold nanoparticles | nt | nt | (+) Ec peptide (MGF); in C | [34] | ||||
| Epithelial Cervical cancer | One uterine cervical NEN; IHC | nt | nt | (+) Ec peptide; in C | [32] | |||
| n = 29 squamous CC (HPV+), n = 28 L-SIL (HPV+), n = 30 H-SIL (HPV+), n = 20 CT (HPV-); PCR, qRT-PCR | (+++) 85% in CC; 92% in CT | (+++) 14% in CC; 8% in CT | (+++) 1% in CC and CT | nt | [49] | |||
| Class I - 69–86%; class II - 14–31%; ↑ of all isoforms in pre-cancerous tissues vs. CC and CT, and a shift in the balance towards IGF1Eb in CC; (+) correlation between the FOX2 mRNA expression and Ea/Eb in precancerous and CC, and Ec in L-SIL and H-SIL | ||||||||
| Gastric cancer | n = 8 NENs; IHC | nt | nt | (+) Ec peptide in 37.5%; in C | [32] | |||
| Lung cancer | n = 2 NENs; IHC | nt | nt | (+) Ec peptide in one LC; in C | [32] | |||
| Pancreatic cancer | n = 17 NENs; IHC | nt | nt | (+) Ec peptide in 58.8%; in C | [32] | |||
| Prostate cancer | IHC, Western blot, RT-PCR | nt | nt | (+++) MGF; ↑ PC and PIN vs. CT | [108] | |||
| n = 83 patients; paraffin sections; IHC | nt | nt | (+) Ec peptide; in C; #, ↑ Ec peptide | [109] | ||||
| n = 78 patients; IHC; Western blot, qRT-PCR | nt | nt | (+) Ec peptide; #, ↑ Ec peptide | [166] | ||||
| Small intestine cancer | n = 9 NENs, IHC | nt | nt | (+) Ec in 44.4%; in C | [32] | |||
| Thyroid cancer | n = 92 of different types of TC; IHC, qRT-PCR | nt | (+) IGF1Ec; ↑in more aggressive vs. non-aggressive papillary TC | nt | (+) Ec peptide in papillary TC; in C; #, ↑ Ec | [35] | ||
| Urinary bladder cancer | n = 46 biopsies/CT; qRT-PCR | (+) marginally ↑ vs. CT | (+) marginally ↑ vs. CT | ↓ vs. CT; #, ↓IGF1Ec | nt | nt | [33] | |
| ↑ of all IGF1 mRNA isoforms in in situ carcinomas | ||||||||
| UPO | n = 4 NENs; IHC | nt | nt | (+) Ec peptide in 100%; in C | [32] | |||
| Others | NENs; appendiceal (n = 3), gallbladder (n = 1); IHC | nt | nt | (-) Ec peptide in 100% | [32] | |||
| IGF1 mRNA Isoforms | IGF1 Propetides/Peptides | |||||||
|---|---|---|---|---|---|---|---|---|
| Human Cancer | Human Cell Lines | IGF1Ea (Class I/II) | IGF1Eb (Class I/II) | IGF1Ec (Class I/II) | Pro-Ea/Ea | Pro-Eb/Eb | Pro-Ec/Ec (MGF) | No of ref. |
| Breast cancer | MSF7 | (+)↑ vs. Ec; class nt | (-) | (+); class nt | nt | (+) Ec peptide | [106] | |
| nt | nt | shEc - ↑cell proliferation and migration via ERK1/2 | [170] | |||||
| nt | nt | rEb peptide - anticancer activity | nt | [168,169] | ||||
| nt | all pro-forms - ↑cell proliferation via the IGF1R; less capable of phosphorylating the IGF1R vs. mature IGF1 | [16] | ||||||
| MDA-MB-231 | (+); class nt | (-) | (+)↑ vs. Ea; class nt | nt | (+) Ec peptide | [106] | ||
| nt | nt | shEc did not ↑cell proliferation | [170] | |||||
| nt | nt | rEb peptide - anticancer activity | nt | [168,169,171,172] | ||||
| T47D | nt | all pro-forms - ↑cell proliferation via the IGF1R signaling, less capable of phosphorylating the IGF1R vs. mature IGF1 | [16] | |||||
| ZR751 | nt | |||||||
| Colorectal cancer | DLD1 | (+) ≈ vs. Eb, ↑ vs. Ec; from class I/II | (+)↑ vs. Ec; from class I/II | (+) from class I/II | nt | (+) Ec peptide | [106] | |
| SW620 | nt | nt | (+) Ec (MGF) peptide | [34] | ||||
| HT29 | nt | nt | (+) Ec (MGF) peptide | [34] | ||||
| nt | nt | rEb peptide - anticancer activity | nt | [169] | ||||
| Endometrial cancer | KLE | (+) all mRNA isoforms in stromal cells of eutopic and ectopic endometrium; (+) IGF1Ec - in glandular cells of ectopic endometrium | nt | sEc peptide (MGF) - ↑cell growth via an IGF1R-, INSR-independent mechanism | [107] | |||
| (+)↑ vs. Eb; from class I/II | (+); from class I/II | (+)↑ vs. Ea&Eb; from class I/II | (+) pro-Ea; ↑ vs. other cells | nt | (+) Ec peptide; ↑ vs. other cells | [106] | ||
| Epithelial Cervical cancer | HeLa (HPV18+) | nt | nt | shEb peptide - ↑cell growth; in N | nt | [90] | ||
| (+)↑ vs. Eb&Ec; in C | (+)↑ vs. Ec | (+) | (+) pro-IGF1A; ↑ vs. other cells | (+) pro-IGF1B in N; (+) hEb peptide in N | (+) Ec - very low expression | [31] | ||
| (+) ≈ vs. Eb; from class II | (+) from class I | (+)↑ vs. Ea&Eb; from class I | nt | (+) Ec peptide; ↑ vs. other cells | [106] | |||
| Hepatocellular cancer | HepG2 | (++)↑ vs. Ec | (+++)↑ vs. Ea and Ec | (+) | (+) pro-IGF1A; (-) Ea peptide | (-) pro-IGF1B;(+) hEb peptide in N | (+) Ec - very low expression | [31] |
| nt | nt | rEb - anticancer activity | nt | [169] | ||||
| HuH7 | (+) ≈ vs. Eb; from class I/II | (+) from class I | (+)↑ vs. Ea&Eb; from class I | nt | (+) Ec peptide | [106] | ||
| Lung cancer | NCI-H345 | nt | nt | sEb peptide - ↑cell proliferation via an IGF1R-independent mechanism | nt | [84] | ||
| A549 | (+)↑ vs. Eb; from class I/II | (+) from class I | (+)↑ vs. Ea&Eb; from class I/II | nt | (+) Ec peptide | [106] | ||
| Melanoma malignum | SK-MEL28 | (+)↑ vs. Eb&Ec; from class I/II | (+)↑ vs. Ec; ↑ vs. other cells; from class I | (+); from class I | nt | (+) pro-IGF1Eb; ↑ vs. other cells | (+) Ec peptide; ↑ vs. other cells | [106] |
| Osteosarcoma | U2OS | (+)↑ vs. Ec | (+)↑ vs. Ea and Ec | (+) | (+) pro-IGF1A; (-) Ea peptide | (-) pro-IGF1B;(+) hEb in N | (+) Ec - very low expression | [31] |
| nt | nt | sEb peptide - ↑cell growth; in N | nt | [90] | ||||
| MG63 | (+) from class I | (+)↑ vs. Ea; from class II | (+)↑ vs. Ea&Eb; from class I | nt | (+) Ec peptide; ↑ vs. other cells | [106] | ||
| nt | (+) Ec - ↑ vs. MHos cells | nt | sEc peptide (MGF) - ↑cell proliferation and migration | [15] | ||||
| (+) | (-); (+) after exposure to DHT for 72 h | (+) | nt | sEc peptide - ↑cell growth via IGF1R/INSR/hybrid receptor-independent way | [14] | |||
| Prostate cancer | LnCaP | (+);↑ vs. Eb; ↑ vs. other cells; from class I/II | (+)↑ vs. other cells; from class I | (+)↑ vs. Ea&Eb; from class I | nt | (+) Ec peptide; ↑ vs. other cells | [106] | |
| nt | (+) | nt | (+) Ec peptide; sEc peptide - ↑cell growth via ERK1/2 and IGF1R/INSR/hybrid receptor-independent mechanism | [108] | ||||
| PC3 | nt | (+) | nt | (+) Ec peptide; sEc - ↑cell growth via ERK1/2 and IGF1R/INSR/hybrid receptor-independent mechanism | [108] | |||
| nt | (+) | nt | (+) Ec peptide; endogenous Ec peptide - ↑cell proliferation via ERK1/2 | [166] | ||||
| Myelogenous leukemia | K562 | (+++)↑ vs. Eb&Ec | (++)↑ vs. Ec | (+) | (+) pro-IGF1A; (-) Ea peptide | (-) pro-IGF1B; ↑ Eb peptide vs. other cells; (+) hEb in N | (+) Ec - very low expression | [31] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kasprzak, A.; Szaflarski, W. Role of Alternatively Spliced Messenger RNA (mRNA) Isoforms of the Insulin-Like Growth Factor 1 (IGF1) in Selected Human Tumors. Int. J. Mol. Sci. 2020, 21, 6995. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21196995
Kasprzak A, Szaflarski W. Role of Alternatively Spliced Messenger RNA (mRNA) Isoforms of the Insulin-Like Growth Factor 1 (IGF1) in Selected Human Tumors. International Journal of Molecular Sciences. 2020; 21(19):6995. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21196995
Chicago/Turabian StyleKasprzak, Aldona, and Witold Szaflarski. 2020. "Role of Alternatively Spliced Messenger RNA (mRNA) Isoforms of the Insulin-Like Growth Factor 1 (IGF1) in Selected Human Tumors" International Journal of Molecular Sciences 21, no. 19: 6995. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21196995