Early Stimulation of TREK Channel Transcription and Activity Induced by Oxaliplatin-Dependent Cytosolic Acidification

 , , ,
, , ,  ,
,  ,
,
Abstract
:1. Introduction
2. Results
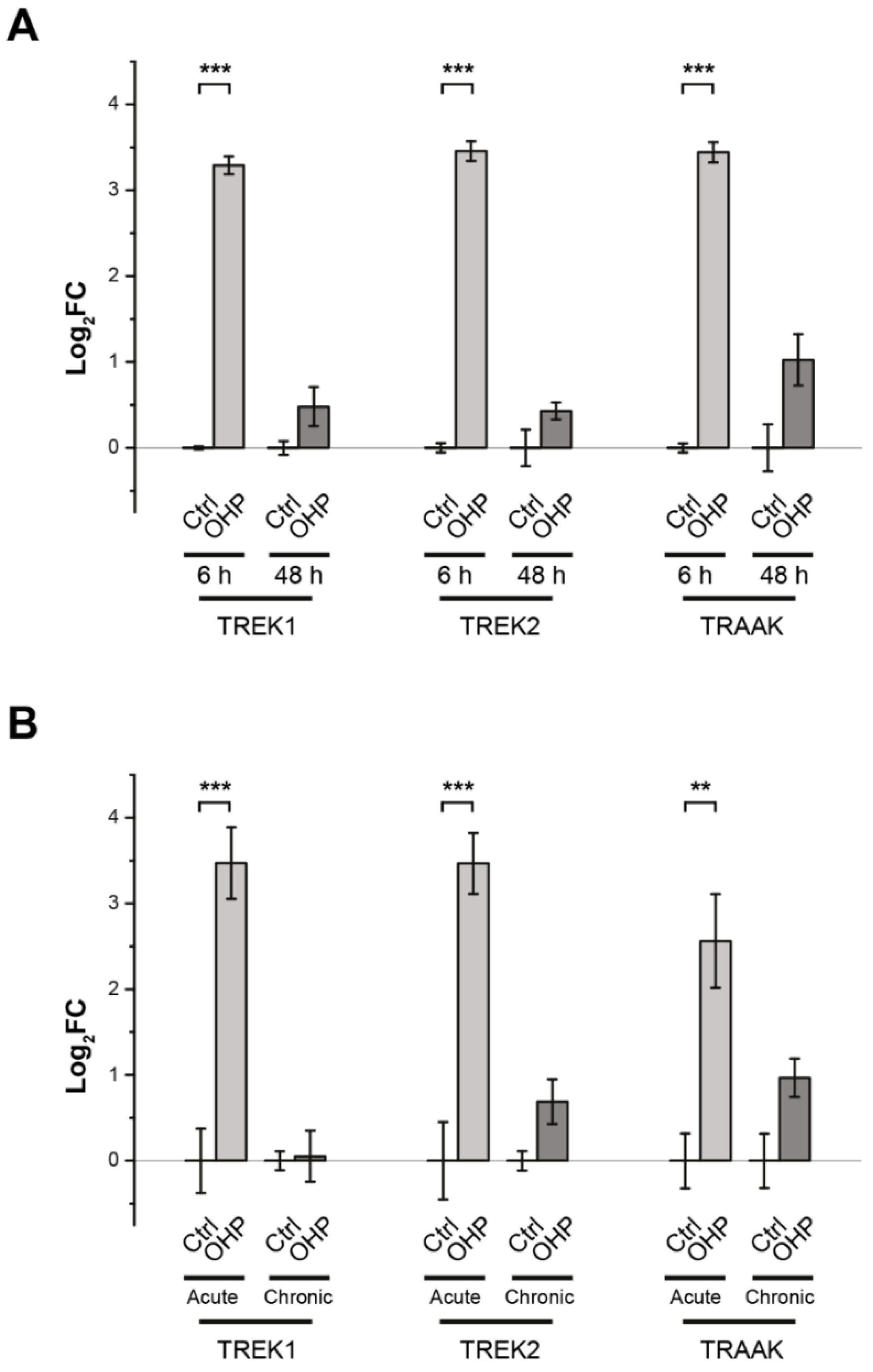
2.1. Effects of OHP on TREK Transcription in DRG Cells
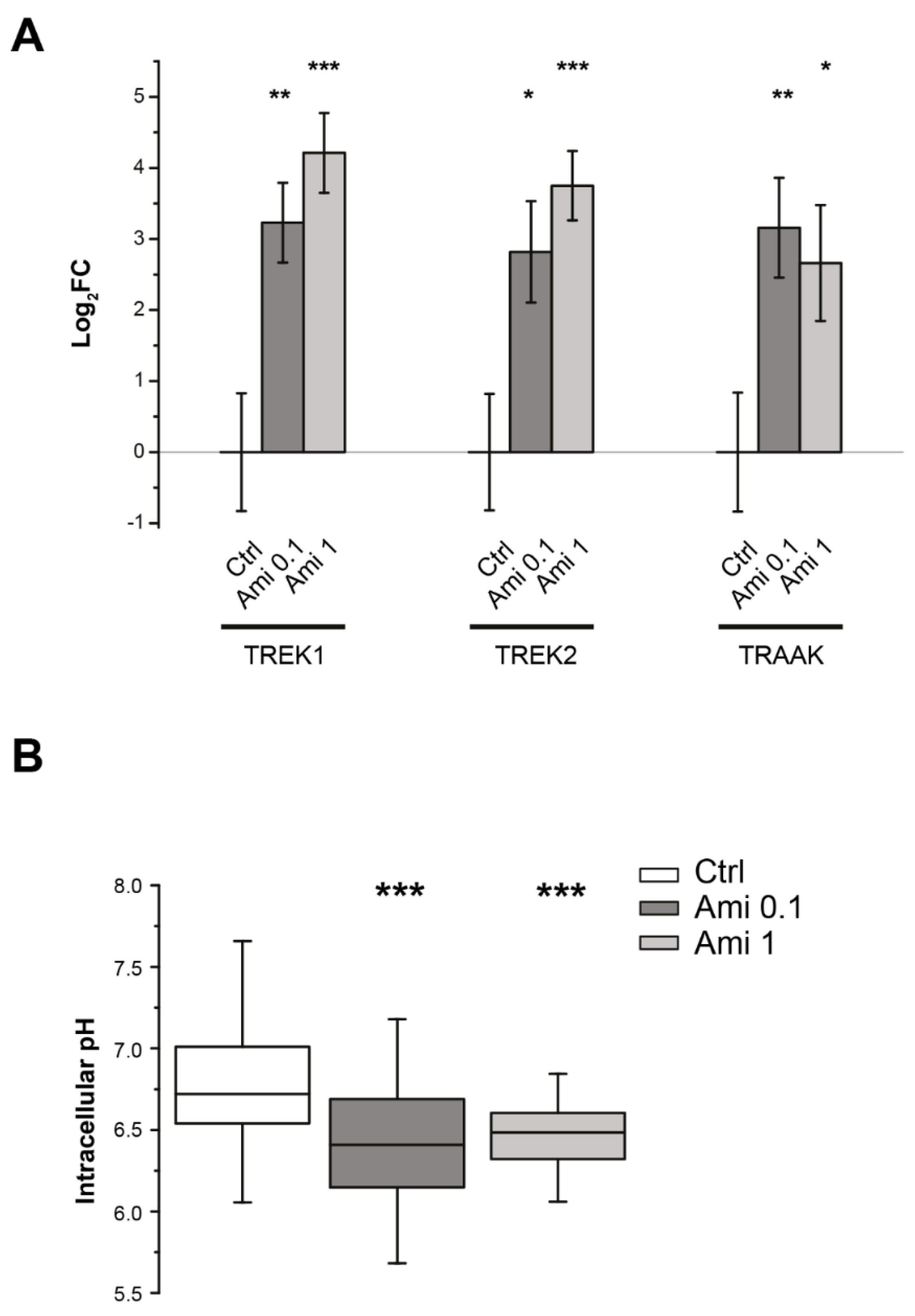
2.2. Effects of Amiloride on Intracellular pH and TREK Channel Transcription
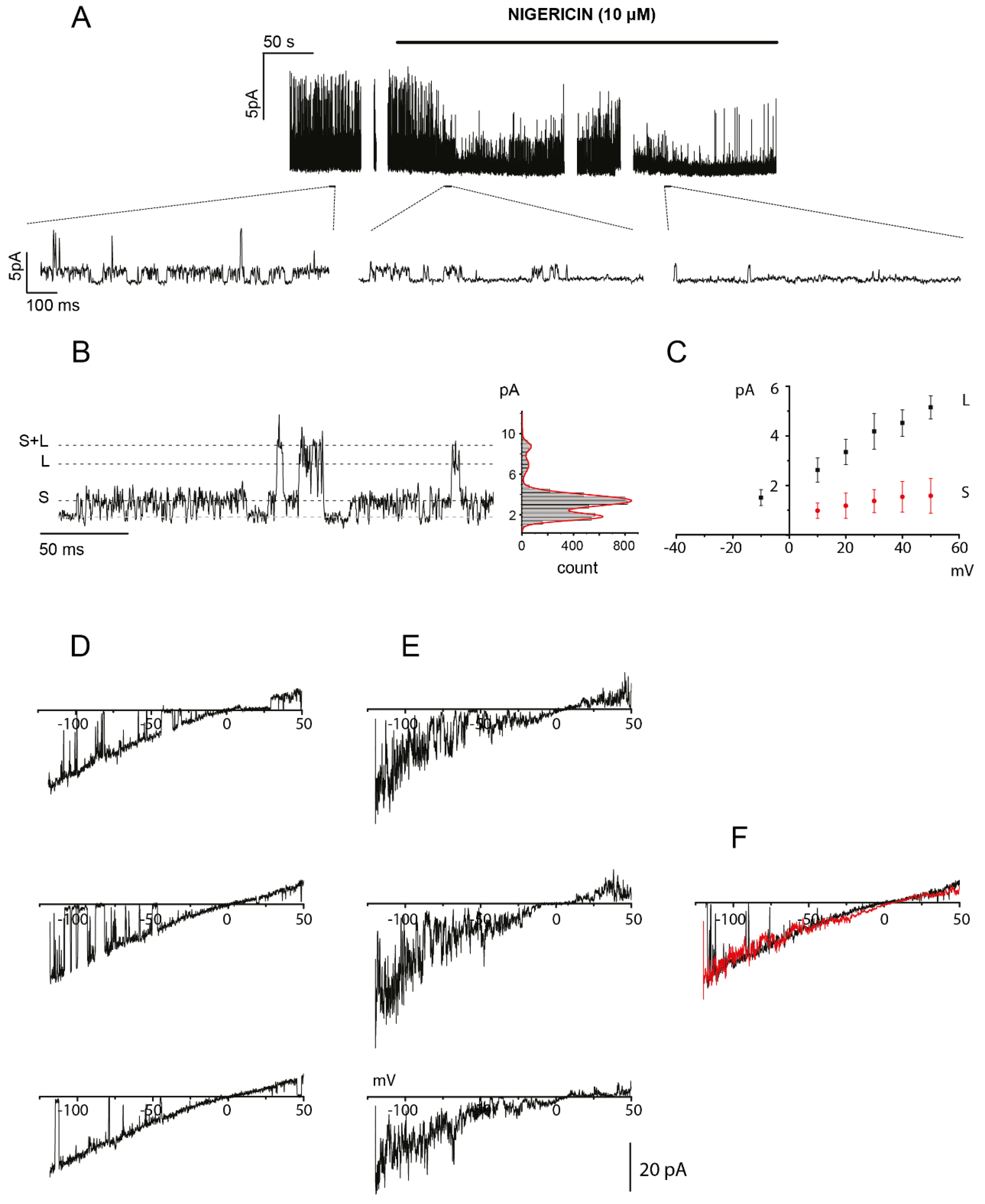
2.3. OHP-Induced Cytosolic Acidification Positively Regulates TREK-2 Channel Activity
2.4. Effects of OHP on Electrical Activity in DRG Neurons
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Chemicals
4.3. Isolation, Culture, and Treatment of Mouse DRG Neurons
4.4. Real-Time Quantitative PCR (RT-qPCR)
4.5. Measurement of Intracellular pH in DRG Cultures by Epifluorescence Microscopy with BCECF
4.6. Electrophysiology-Multielectrode Arrays (MEAs)
4.7. Electrophysiology—Patch–Clamp
4.8. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Grisold, W.; Cavaletti, G.; Windebank, A.J. Peripheral neuropathies from chemotherapeutics and targeted agents. Neuro Oncol. 2012, 14, 45–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Descoeur, J.; Pereira, V.; Pizzoccaro, A.; Francois, A.; Ling, B.; Maffre, V.; Couette, B.; Busserolles, J.; Courteix, C.; Noel, J.; et al. Oxaliplatin-induced cold hypersensitivity is due to remodelling of ion channel expression in nociceptors. EMBO Mol. Med. 2011, 3, 266–278. [Google Scholar] [CrossRef] [PubMed]
- Marmiroli, P.; Cavaletti, G.; Carozzi, V.; Riva, B.; Lim, D.; AGenazzani, A. Calcium-related Neurotoxicity of Oxaliplatin: Understanding the Mechanisms to Drive Therapy. Curr. Med. Chem. 2015, 22, 3682–3694. [Google Scholar] [CrossRef] [PubMed]
- Chiorazzi, A.; Semperboni, S.; Marmiroli, P. Current view in platinum drug mechanisms of peripheral neurotoxicity. Toxics 2015, 3, 304–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plant, L.D. A Role for K2P Channels in the Operation of Somatosensory Nociceptors. Front. Mol. Neurosci. 2012, 5, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamas, J.A.; Rueda-Ruzafa, L.; Herrera-Pérez, S. Ion Channels and Thermosensitivity: TRP, TREK, or Both? Int. J. Mol. Sci. 2019, 20, 2371. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Isami, K.; Nakamura, S.; Shirakawa, H.; Nakagawa, T.; Kaneko, S. Acute cold hypersensitivity characteristically induced by oxaliplatin is caused by the enhanced responsiveness of TRPA1 in mice. Mol. Pain 2012, 8, 55. [Google Scholar] [CrossRef] [Green Version]
- Alloui, A.; Zimmermann, K.; Mamet, J.; Duprat, F.; Noël, J.; Chemin, J.; Guy, N.; Blondeau, N.; Voilley, N.; Rubat-Coudert, C.; et al. TREK-1, a K+ channel involved in polymodal pain perception. EMBO J. 2006, 25, 2368–2376. [Google Scholar] [CrossRef] [Green Version]
- Noël, J.; Zimmermann, K.; Busserolles, J.; Deval, E.; Alloui, A.; Diochot, S.; Guy, N.; Borsotto, M.; Reeh, P.; Eschalier, A.; et al. The mechano-activated K±channels TRAAK and TREK-1 control both warm and cold perception. EMBO J. 2009, 28, 1308–1318. [Google Scholar] [CrossRef] [Green Version]
- Waxman, S.G.; Zamponi, G.W. Regulating excitability of peripheral afferents: Emerging ion channel targets. Nat. Neurosci. 2014, 17, 153–163. [Google Scholar] [CrossRef]
- Jin, P.; Jan, L.Y.; Jan, Y.-N. Mechanosensitive Ion Channels: Structural Features Relevant to Mechanotransduction Mechanisms. Annu. Rev. Neurosci. 2020, 43, 207–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gada, K.; Plant, L.D. Two-pore domain potassium channels: Emerging targets for novel analgesic drugs: IUPHAR Review 26. Br. J. Pharmacol. 2019, 176, 256–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riva, B.; Dionisi, M.; Potenzieri, A.; Chiorazzi, A.; Cordero-Sanchez, C.; Rigolio, R.; Carozzi, V.A.; Lim, D.; Cavaletti, G.; Marmiroli, P.; et al. Oxaliplatin induces pH acidification in dorsal root ganglia neurons. Sci. Rep. 2018, 8, 15084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potenzieri, A.; Riva, B.; Rigolio, R.; Chiorazzi, A.; Pozzi, E.; Ballarini, E.; Cavaletti, G.; Genazzani, A.A. Oxaliplatin-induced neuropathy occurs through impairment of haemoglobin proton buffering and is reversed by carbonic anhydrase inhibitors. Pain 2020, 161, 405–415. [Google Scholar] [CrossRef]
- Patel, A.J.; Honoré, E.; Maingret, F.; Lesage, F.; Fink, M.; Duprat, F.; Lazdunski, M. A mammalian two pore domain mechano-gated S-like K + channel. EMBO J. 1998, 17, 4283–4290. [Google Scholar] [CrossRef]
- Braun, G.; Lengyel, M.; Enyedi, P.; Czirják, G. Differential sensitivity of TREK-1, TREK-2 and TRAAK background potassium channels to the polycationic dye ruthenium red. Br. J. Pharmacol. 2015, 172, 1728–1738. [Google Scholar] [CrossRef] [Green Version]
- Lesage, F.; Terrenoire, C.; Romey, G.; Lazdunski, M. Human TREK2, a 2P Domain Mechano-sensitive K+Channel with Multiple Regulations by Polyunsaturated Fatty Acids, Lysophospholipids, and Gs, Gi, and GqProtein-coupled Receptors. J. Biol. Chem. 2000, 275, 28398–28405. [Google Scholar] [CrossRef] [Green Version]
- Pereira, V.; Busserolles, J.; Christin, M.; Devilliers, M.; Poupon, L.; Legha, W.; Alloui, A.; Aissouni, Y.; Bourinet, E.; Lesage, F.; et al. Role of the TREK2 potassium channel in cold and warm thermosensation and in pain perception. Pain 2014, 155, 2534–2544. [Google Scholar] [CrossRef]
- Steen, K.H.; Wegner, H.; Reeh, P.W. The pH response of rat cutaneous nociceptors correlates with extracellular [Na+] and is increased under amiloride. Eur. J. Neurosci. 1999, 11, 2783–2792. [Google Scholar] [CrossRef]
- Ruffin, V.A.; Salameh, A.I.; Boron, W.F.; Parker, M.D. Intracellular pH regulation by acid-base transporters in mammalian neurons. Front. Physiol. 2014, 5, 43. [Google Scholar] [CrossRef] [Green Version]
- Casey, J.R.; Grinstein, S.; Orlowski, J. Sensors and regulators of intracellular pH. Nat. Rev. Mol. Cell Biol. 2010, 11, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.; Choe, C.; Kim, D. Thermosensitivity of the two-pore domain K+ channels TREK-2 and TRAAK. J. Physiol. 2005, 564, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Acosta, C.; Djouhri, L.; Watkins, R.; Berry, C.; Bromage, K.; Lawson, S.N. TREK2 expressed selectively in IB4-binding C-fiber nociceptors hyperpolarizes their membrane potentials and limits spontaneous pain. J. Neurosci. 2014, 34, 1494–1509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noël, J.; Sandoz, G.; Lesage, F. Molecular regulations governing TREK and TRAAK channel functions. Channels 2011, 5, 402–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, D.; Choe, C.; Cavanaugh, E.; Kim, D. Properties of single two-pore domain TREK-2 channels expressed in mammalian cells. J. Physiol. 2007, 583, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Ładno, W.; Gawlak, M.; Szulczyk, P.; Nurowska, E. Kinetic properties and adrenergic control of TREK-2-like channels in rat medial prefrontal cortex (mPFC) pyramidal neurons. Brain Res. 2017, 1665, 95–104. [Google Scholar] [CrossRef]
- Cadaveira-Mosquera, A.; Ribeiro, S.J.; Reboreda, A.; Pérez, M.; Lamas, J.A. Activation of TREK currents by the neuroprotective agent riluzole in mouse sympathetic neurons. J. Neurosci. 2011, 31, 1375–1385. [Google Scholar] [CrossRef]
- Fernández-Fernández, D.; Cadaveira-Mosquera, A.; Rueda-Ruzafa, L.; Herrera-Pérez, S.; Veale, E.L.; Reboreda, A.; Mathie, A.; Antonio Lamas, J. Activation of TREK currents by riluzole in three subgroups of cultured mouse nodose ganglion neurons. PLoS ONE 2018, 13, e0199282. [Google Scholar] [CrossRef]
- Newberry, K.; Wang, S.; Hoque, N.; Kiss, L.; Ahlijanian, M.K.; Herrington, J.; Graef, J.D. Development of a spontaneously active dorsal root ganglia assay using multiwell multielectrode arrays. J. Neurophysiol. 2016, 115, 3217–3228. [Google Scholar] [CrossRef] [Green Version]
- Aromolaran, K.A.; Goldstein, P.A. Ion channels and neuronal hyperexcitability in chemotherapy-induced peripheral neuropathy: Cause and effect? Mol. Pain 2017, 13, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Zajaczkowską, R.; Kocot-Kępska, M.; Leppert, W.; Wrzosek, A.; Mika, J.; Wordliczek, J. Mechanisms of chemotherapy-induced peripheral neuropathy. Int. J. Mol. Sci. 2019, 20, 1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.B.; Zhang, H.X.; Li, L.L.; Wang, X.L. Enhanced expressions of arachidonic acid-sensitive tandem-pore domain potassium channels in rat experimental acute cerebral ischemia. Biochem. Biophys. Res. Commun. 2005, 327, 1163–1169. [Google Scholar] [CrossRef] [PubMed]
- Orlowski, J. Heterologous expression and functional properties of amiloride high affinity (NHE-1) and low affinity (NHE-3) isoforms of the rat Na/H exchanger. J. Biol. Chem. 1993, 268, 16369–16377. [Google Scholar] [PubMed]
- Causton, H.C.; Ren, B.; Koh, S.S.; Harbison, C.T.; Kanin, E.; Jennings, E.G.; Lee, T.L.; True, H.L.; Lander, E.S.; Young, R.A. Remodeling of yeast genome expression in response to environmental changes. Mol. Biol. Cell 2001, 12, 323–337. [Google Scholar] [CrossRef] [Green Version]
- Serrano, R.; Ruiz, A.; Bernal, D.; Chambers, J.R.; Ariño, J. The transcriptional response to alkaline pH in Saccharomyces cerevisiae: Evidence for calcium-mediated signalling. Mol. Microbiol. 2002, 46, 1319–1333. [Google Scholar] [CrossRef]
- Olson, E.R. Influence of pH on bacterial gene expression. Mol. Microbiol. 1993, 8, 5–14. [Google Scholar] [CrossRef]
- Putney, L.K.; Barber, D.L. Expression profile of genes regulated by activity of the Na-H exchanger NHE1. BMC Genom. 2004, 5, 46. [Google Scholar] [CrossRef] [Green Version]
- Torres-López, J.E.; Guzmán-Priego, C.G.; Rocha-González, H.I.; Granados-Soto, V. Role of NHE1 in nociception. Pain Res. Treat. 2013, 2013, 217864. [Google Scholar] [CrossRef] [Green Version]
- Zhou, D.; Xue, J.; Gavrialov, O.; Haddad, G.G. Na+/H+ exchanger 1 deficiency alters gene expression in mouse brain. Physiol. Genom. 2004, 18, 331–339. [Google Scholar] [CrossRef] [Green Version]
- Rebillard, A.; Tekpli, X.; Meurette, O.; Sergent, O.; LeMoigne-Muller, G.; Vernhet, L.; Gorria, M.; Chevanne, M.; Christmann, M.; Kaina, B.; et al. Cisplatin-induced apoptosis involves membrane fluidification via inhibition of NHE1 in human colon cancer cells. Cancer Res. 2007, 67, 7865–7874. [Google Scholar] [CrossRef] [Green Version]
- Milosavljevic, N.; Duranton, C.; Djerbi, N.; Milosavljevic, N.; Duranton, C.; Djerbi, N.; Puech, P.H.; Gounon, P. Nongenomic Effects of Cisplatin: Acute Inhibition of Mechanosensitive Transporters and Channels without Actin Remodeling Nongenomic Effects of Cisplatin: Acute Inhibition of Mechanosensitive Transporters and Channels without Actin Remodeling. Cancer Res. 2010, 70, 7514–7522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, H.J.; Lee, S.W.; Kim, G.T.; Kim, E.J.; Kwon, B.; Kang, D.; Kim, H.J.; Seo, K.S. Enhanced expression of TREK-1 is related with chronic constriction injury of neuropathic pain mouse model in dorsal root ganglion. Biomol. Ther. 2016, 24, 252–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La, J.H.; Schwartz, E.S.; Gebhart, G.F. Differences in the expression of transient receptor potential channel V1, transient receptor potential channel A1 and mechanosensitive two pore-domain K + channels between the lumbar splanchnic and pelvic nerve innervations of mouse urinary bladder and colon. Neuroscience 2011, 186, 179–187. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.; Koo, N.; Jin, M.; Davies, A.J.; Chun, G.; Choi, S.; Kim, J.; Park, K. Intracellular Acidification Is Associated with Changes in Free Cytosolic Calcium and Inhibition of Action Potentials in Rat Trigeminal Ganglion. J. Biol. Chem. 2011, 286, 1719–1729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marmiroli, P.; Riva, B.; Pozzi, E.; Ballarini, E.; Lim, D.; Chiorazzi, A.; Meregalli, C.; Distasi, C.; Renn, C.L.; Semperboni, S.; et al. Susceptibility of different mouse strains to oxaliplatin peripheral neurotoxicity: Phenotypic and genotypic insights. PLoS ONE 2017, 12, e0186250. [Google Scholar] [CrossRef]
- Renn, C.L.; Carozzi, V.A.; Rhee, P.; Gallop, D.; Dorsey, S.G.; Cavaletti, G. Multimodal assessment of painful peripheral neuropathy induced by chronic oxaliplatin-based chemotherapy in mice. Mol. Pain 2011, 7, 29. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 6 h | 24 h | 48 h | ||||
|---|---|---|---|---|---|---|
| Ctrl | OHP | Ctrl | OHP | Ctrl | OHP | |
| M (mHz) | 267 | 175 | 425 | 217 | 350 | 400 |
| IQR (mHz) | 454 | 221 | 813 | 467 | 758 | 867 |
| Q1 (mHz) | 146 | 100 | 154 | 100 | 150 | 183 |
| Q3 (mHz) | 600 | 321 | 967 | 567 | 908 | 1050 |
| n channels | 86 | 90 | 72 | 96 | 157 | 155 |
| n MEAs | 9 | 9 | 4 | 7 | 6 | 7 |
| p-value | 0.0018 | 0.0091 | 0.2350 | |||
| r | 0.2348 | 0.2012 | 0.0672 | |||
| Cohen’s d | 0.1657 | 0.3872 | 0.0670 | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dionisi, M.; Ruffinatti, F.A.; Riva, B.; Lim, D.; Canta, A.; Meregalli, C.; Fumagalli, G.; Monza, L.; Ferrer-Montiel, A.; Fernandez-Carvajal, A.; et al. Early Stimulation of TREK Channel Transcription and Activity Induced by Oxaliplatin-Dependent Cytosolic Acidification. Int. J. Mol. Sci. 2020, 21, 7164. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197164
Dionisi M, Ruffinatti FA, Riva B, Lim D, Canta A, Meregalli C, Fumagalli G, Monza L, Ferrer-Montiel A, Fernandez-Carvajal A, et al. Early Stimulation of TREK Channel Transcription and Activity Induced by Oxaliplatin-Dependent Cytosolic Acidification. International Journal of Molecular Sciences. 2020; 21(19):7164. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197164
Chicago/Turabian StyleDionisi, Marianna, Federico Alessandro Ruffinatti, Beatrice Riva, Dmitry Lim, Annalisa Canta, Cristina Meregalli, Giulia Fumagalli, Laura Monza, Antonio Ferrer-Montiel, Asia Fernandez-Carvajal, and et al. 2020. "Early Stimulation of TREK Channel Transcription and Activity Induced by Oxaliplatin-Dependent Cytosolic Acidification" International Journal of Molecular Sciences 21, no. 19: 7164. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197164