Chemerin Isoform-Specific Effects on Hepatocyte Migration and Immune Cell Inflammation


Abstract
:1. Introduction
2. Results
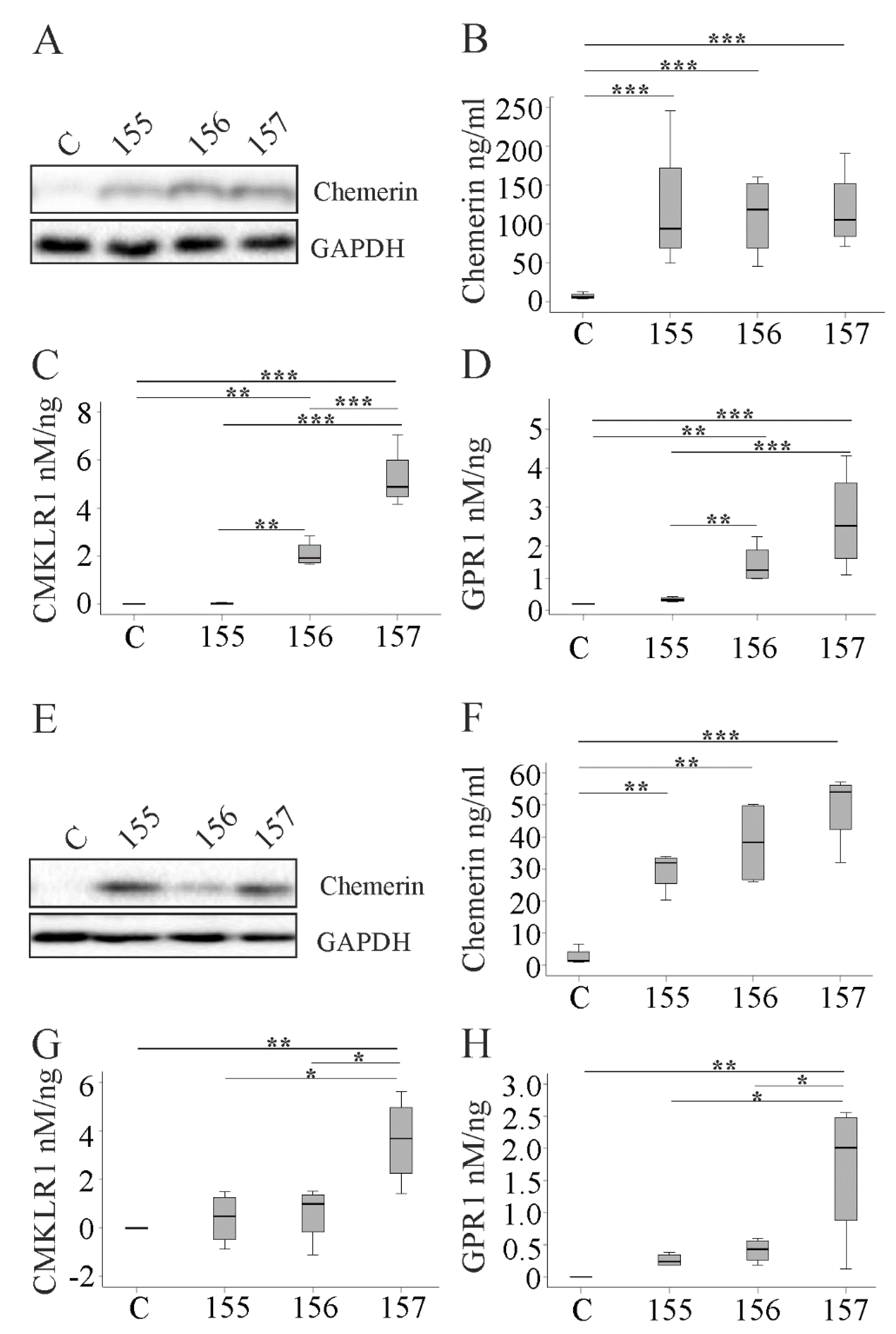
2.1. Overexpression of Chemerin Isoforms in Hepa1-6 Cells
2.2. Overexpression of Chemerin Isoforms in HepG2 and Huh7 Cells
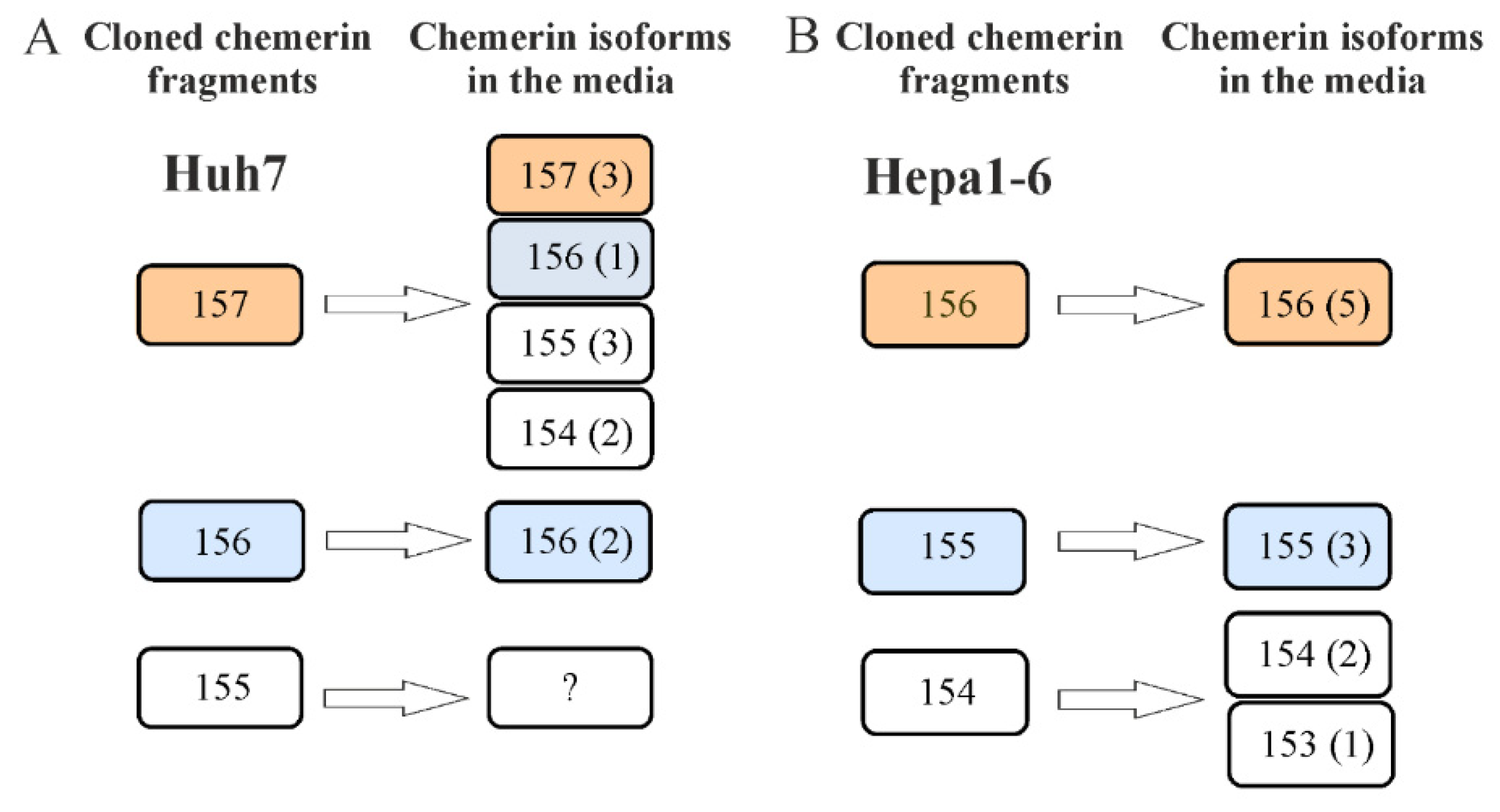
2.3. Mass Spectrometric Analysis of Chemerin Isoforms
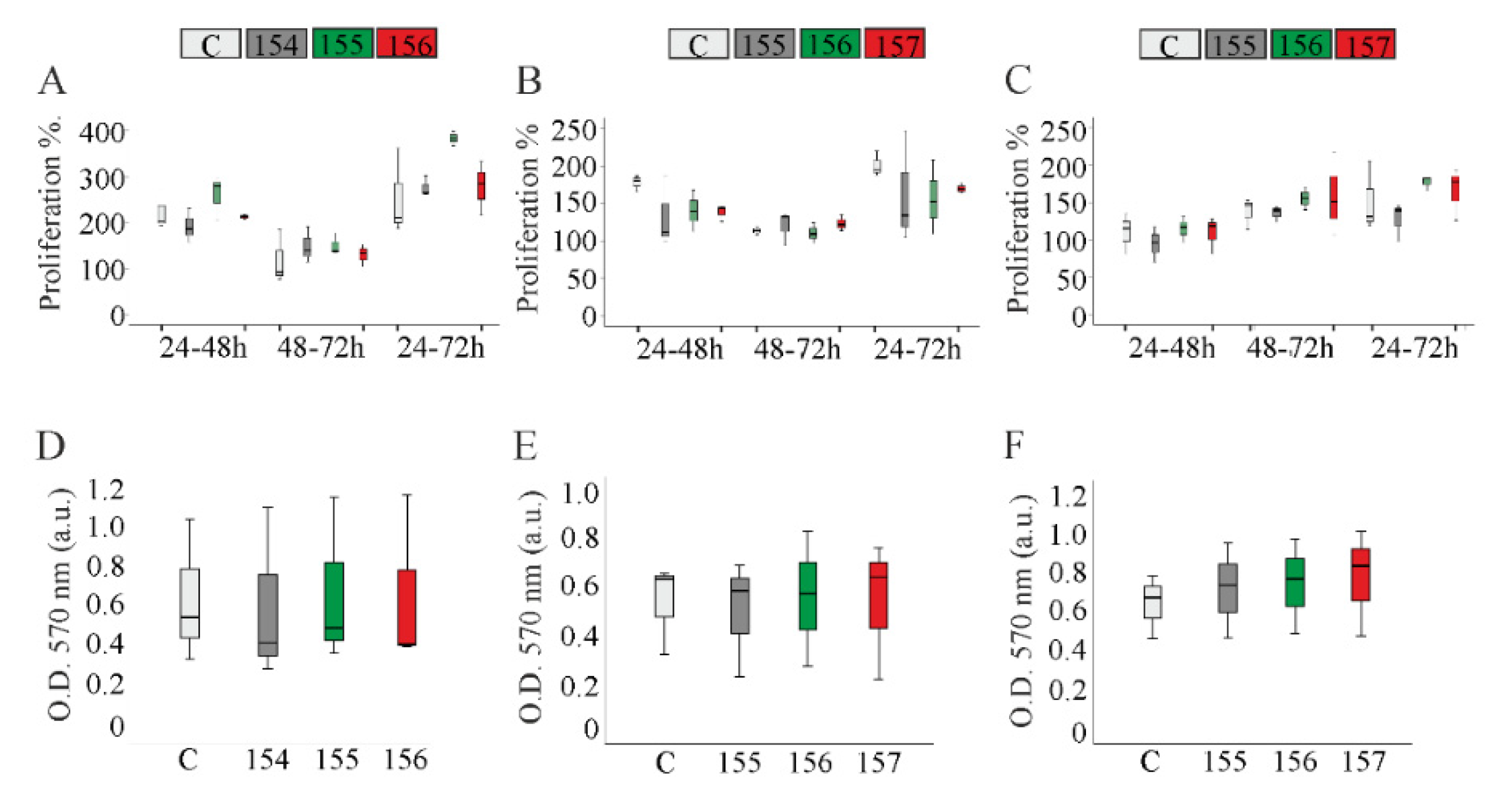
2.4. Proliferation of Chemerin-Overexpressing Hepatocytes
2.5. Migration of Chemerin Isoform-Overexpressing Hepatocytes
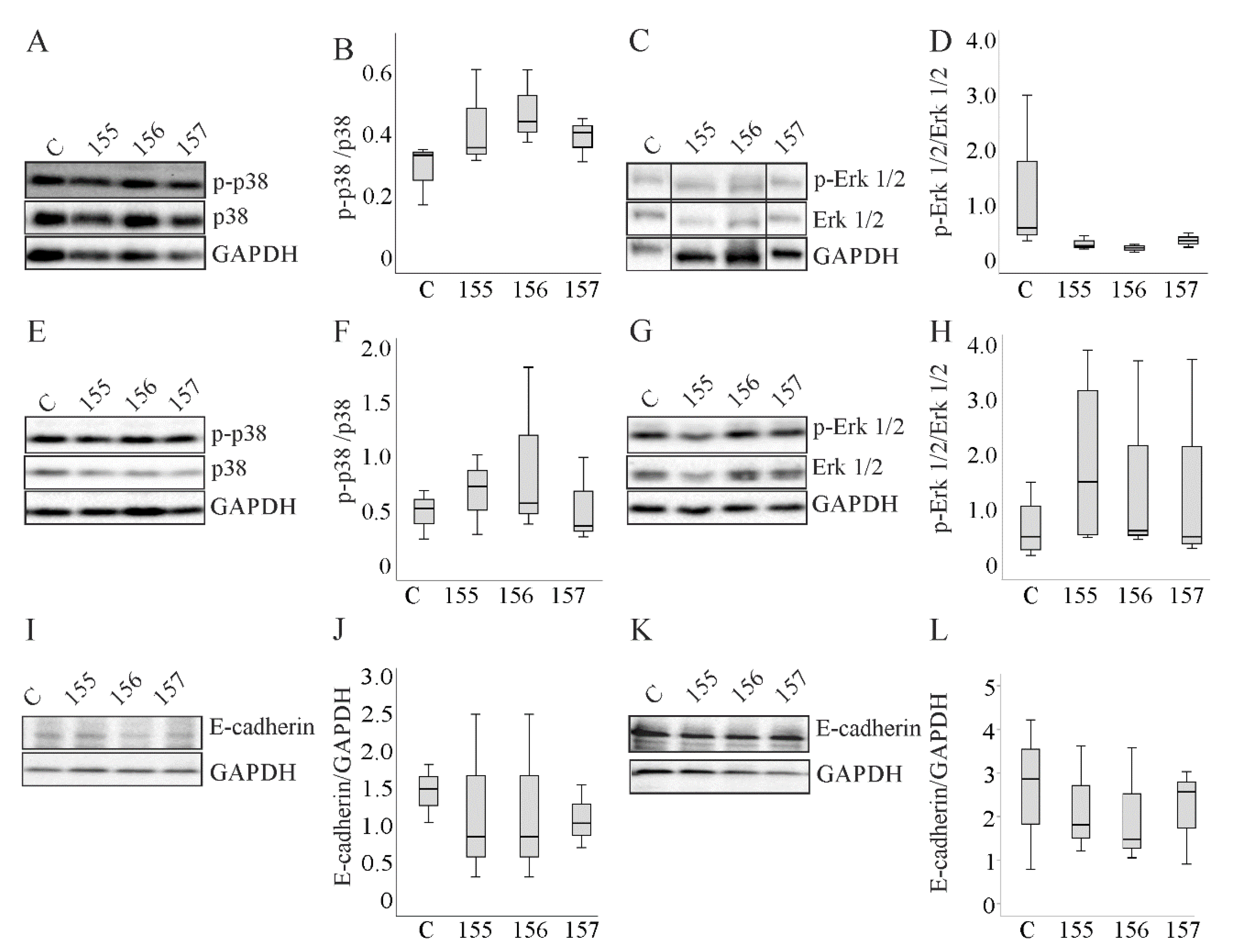
2.6. Activation of Extracellular Signal-Regulated Kinase (ERK)1/2 and p38 Mitogen-Activated Protein Kinase (MAPK,) and Protein Levels of E-Cadherin in Hepatocytes Overexpressing Chemerin Isoforms
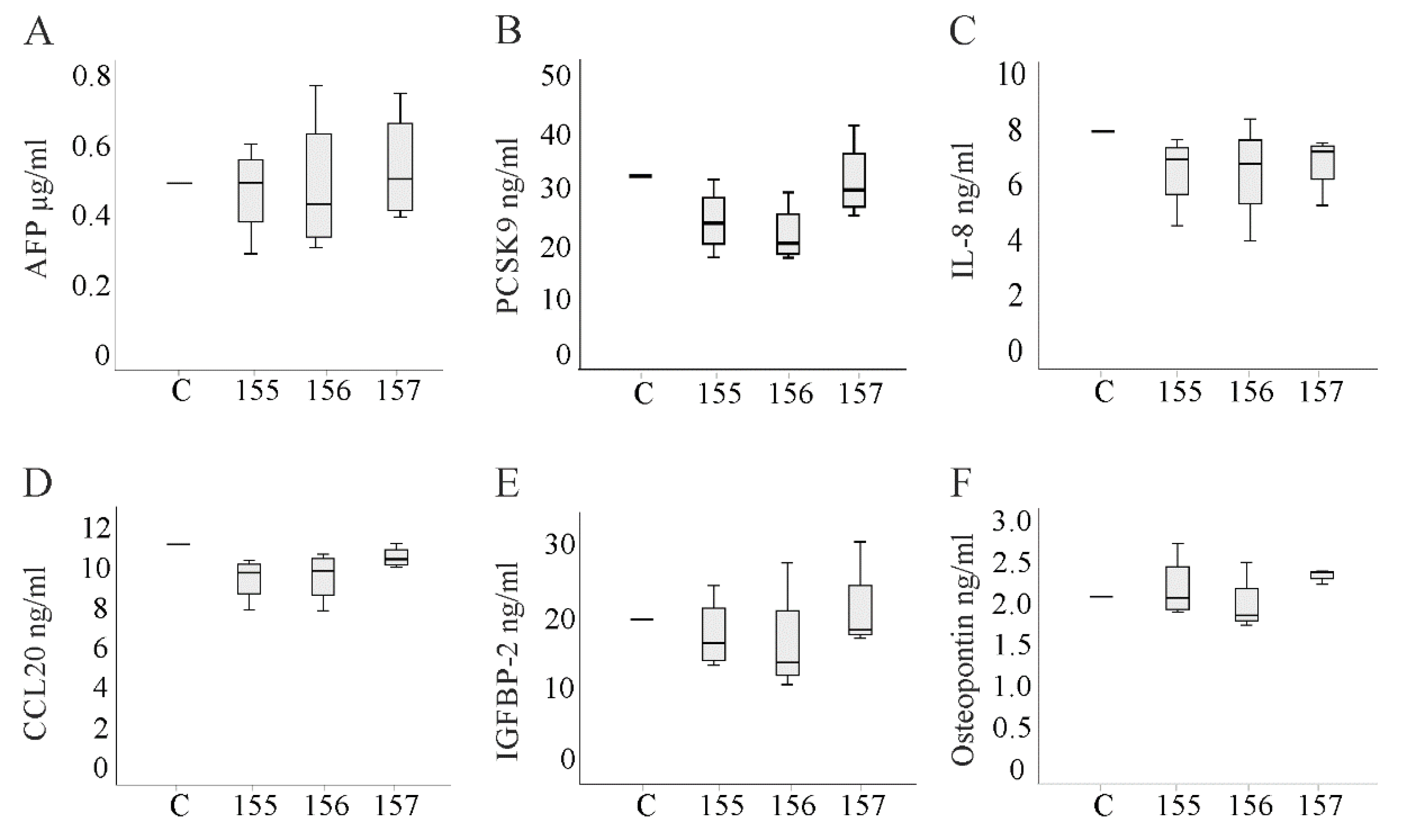
2.7. Alpha-Fetoprotein and Cytokines in the Cell Culture Media of Chemerin Isoform-Overexpressing Hepatocytes
2.8. Cytokines of PBMCs Cultivated in Hepatocyte-Conditioned Medium
3. Discussion
4. Materials and Methods
4.1. Cell lines and Primary Cells
4.2. Expression of Recombinant Human Chemerin Isoforms
4.3. SDS-PAGE and Immunoblotting
4.4. ELISAs, MTT Assay and Cytokine Array
4.5. Tango Assay
4.6. Scratch Assay
4.7. Mass Spectrometry of Chemerin Isoforms
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Buechler, C.; Feder, S.; Haberl, E.M.; Aslanidis, C. Chemerin Isoforms and Activity in Obesity. Int. J. Mol. Sci. 2019, 20, 1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ernst, M.C.; Sinal, C.J. Chemerin: At the crossroads of inflammation and obesity. Trends Endocrinol. Metab. 2010, 21, 660–667. [Google Scholar] [CrossRef] [PubMed]
- Rourke, J.L.; Dranse, H.J.; Sinal, C.J. Towards an integrative approach to understanding the role of chemerin in human health and disease. Obes. Rev. 2012, 14, 245–262. [Google Scholar] [CrossRef] [PubMed]
- Goralski, K.B.; Jackson, A.E.; McKeown, B.T.; Sinal, C.J. More Than an Adipokine: The Complex Roles of Chemerin Signaling in Cancer. Int. J. Mol. Sci. 2019, 20, 4778. [Google Scholar] [CrossRef] [Green Version]
- Treeck, O.; Buechler, C.; Ortmann, O. Chemerin and Cancer. Int. J. Mol. Sci. 2019, 20, 3750. [Google Scholar] [CrossRef] [Green Version]
- Li, J.-J.; Yin, H.-K.; Guan, D.-X.; Zhao, J.-S.; Feng, Y.-X.; Deng, Y.-Z.; Wang, X.; Li, N.; Wang, X.-F.; Cheng, S.-Q.; et al. Chemerin suppresses hepatocellular carcinoma metastasis through CMKLR1-PTEN-Akt axis. Br. J. Cancer 2018, 118, 1337–1348. [Google Scholar] [CrossRef]
- Lin, W.; Chen, Y.-L.; Jiang, L.; Chen, J.-K. Reduced expression of chemerin is associated with a poor prognosis and a lowed infiltration of both dendritic cells and natural killer cells in human hepatocellular carcinoma. Clin. Lab. 2011, 57, 879–885. [Google Scholar]
- Lin, Y.; Yang, X.; Liu, W.; Li, B.; Yin, W.; Shi, Y.; He, R. Chemerin has a protective role in hepatocellular carcinoma by inhibiting the expression of IL-6 and GM-CSF and MDSC accumulation. Oncogene 2017, 36, 3599–3608. [Google Scholar] [CrossRef]
- Haberl, E.M.; Pohl, R.; Rein-Fischboeck, L.; Feder, S.; Sinal, C.J.; Bruckmann, A.; Hoering, M.; Krautbauer, S.; Liebisch, G.; Buechler, C. Overexpression of Hepatocyte Chemerin-156 Lowers Tumor Burden in a Murine Model of Diethylnitrosamine-Induced Hepatocellular Carcinoma. Int. J. Mol. Sci. 2019, 21, 252. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.-S.; Eisenberg, D.; Zhao, L.; Adams, C.; Leib, R.; Morser, J.; Leung, L.L. Chemerin activation in human obesity. Obesity 2016, 24, 1522–1529. [Google Scholar] [CrossRef] [Green Version]
- Toulany, J.; Parlee, S.D.; Sinal, C.J.; Slayter, K.; McNeil, S.A.; Goralski, K.B. CMKLR1 activation ex vivo does not increase proportionally to serum total chemerin in obese humans. Endocr. Connect. 2016, 5, 70–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Yamaguchi, Y.; Shen, W.-J.; Morser, J.; Leung, L.L.K. Dynamic and tissue-specific proteolytic processing of chemerin in obese mice. PLoS ONE 2018, 13, e0202780. [Google Scholar] [CrossRef] [PubMed]
- Du, X.-Y.; Zabel, B.A.; Myles, T.; Allen, S.J.; Handel, T.M.; Lee, P.P.; Butcher, E.C.; Leung, L.L. Regulation of Chemerin Bioactivity by Plasma Carboxypeptidase N, Carboxypeptidase B (Activated Thrombin-activable Fibrinolysis Inhibitor), and Platelets. J. Biol. Chem. 2008, 284, 751–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wittamer, V.; Franssen, J.-D.; Vulcano, M.; Mirjolet, J.-F.; Le Poul, E.; Migeotte, I.; Brézillon, S.; Tyldesley, R.; Blanpain, C.; Detheux, M.; et al. Specific Recruitment of Antigen-presenting Cells by Chemerin, a Novel Processed Ligand from Human Inflammatory Fluids. J. Exp. Med. 2003, 198, 977–985. [Google Scholar] [CrossRef]
- Jacenik, D.; Fichna, J. Chemerin in immune response and gastrointestinal pathophysiology. Clin. Chim. Acta 2020, 504, 146–153. [Google Scholar] [CrossRef]
- Sato, K.; Yoshizawa, H.; Seki, T.; Shirai, R.; Yamashita, T.; Okano, T.; Shibata, K.; Wakamatsu, M.J.; Mori, Y.; Morita, T.; et al. Chemerin-9, a potent agonist of chemerin receptor (ChemR23), prevents atherogenesis. Clin. Sci. 2019, 133, 1779–1796. [Google Scholar] [CrossRef]
- Lin, Y.; Yang, X.; Yue, W.; Xu, X.; Li, B.; Zou, L.; He, R. Chemerin aggravates DSS-induced colitis by suppressing M2 macrophage polarization polarization. Cell. Mol. Immunol. 2014, 11, 355–366. [Google Scholar] [CrossRef]
- Bondue, B.; De Henau, O.; Luangsay, S.; Devosse, T.; De Nadai, P.; Springael, J.-Y.; Parmentier, M.; Vosters, O. The Chemerin/ChemR23 System Does Not Affect the Pro-Inflammatory Response of Mouse and Human Macrophages Ex Vivo. PLoS ONE 2012, 7, e40043. [Google Scholar] [CrossRef] [Green Version]
- De Henau, O.; DeGroot, G.-N.; Imbault, V.; Robert, V.; De Poorter, C.; Mcheik, S.; Galès, C.; Parmentier, M.; Springael, J.-Y. Signaling Properties of Chemerin Receptors CMKLR1, GPR1 and CCRL2. PLoS ONE 2016, 11, e0164179. [Google Scholar] [CrossRef]
- Ferland, D.J.; Mullick, A.E.; Watts, S.W. Chemerin as a driver of hypertension: A consideration. Am. J. Hypertens. 2020. [Google Scholar] [CrossRef]
- Rourke, J.L.; Dranse, H.J.; Sinal, C.J. CMKLR1 and GPR1 mediate chemerin signaling through the RhoA/ROCK pathway. Mol. Cell. Endocrinol. 2015, 417, 36–51. [Google Scholar] [CrossRef] [PubMed]
- Mroweh, M.; Decaens, T.; Marche, P.N.; Jilkova, Z.M.; Clément, F. Modulating the Crosstalk between the Tumor and Its Microenvironment Using RNA Interference: A Treatment Strategy for Hepatocellular Carcinoma. Int. J. Mol. Sci. 2020, 21, 5250. [Google Scholar] [CrossRef] [PubMed]
- Dogra, S.; Sona, C.; Kumar, A.; Yadav, P.N. Tango assay for ligand-induced GPCR-beta-arrestin2 interaction: Application in drug discovery. Methods Cell. Biol. 2016, 132, 233–254. [Google Scholar] [PubMed]
- Lee, H.-M.; Hwang, K.-A.; Choi, K.-C. Diverse pathways of epithelial mesenchymal transition related with cancer progression and metastasis and potential effects of endocrine disrupting chemicals on epithelial mesenchymal transition process. Mol. Cell. Endocrinol. 2017, 457, 103–113. [Google Scholar] [CrossRef]
- Galle, P.; Foerster, F.; Kudo, M.; Chan, S.L.; Llovet, J.M.; Qin, S.; Schelman, W.R.; Chintharlapalli, S.; Abada, P.B.; Sherman, M.; et al. Biology and significance of alpha-fetoprotein in hepatocellular carcinoma. Liver Int. 2019, 39, 2214–2229. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Zhu, M.; Wang, Q.; Hou, Y.; Li, L.; Weng, H.; Zhao, Y.; Chen, D.; Ding, H.; Guo, J.; et al. Alpha-fetoprotein inhibits autophagy to promote malignant behaviour in hepatocellular carcinoma cells by activating PI3K/AKT/mTOR signalling. Cell. Death Dis. 2018, 9, 1027. [Google Scholar] [CrossRef] [Green Version]
- You, M.-L.; Chen, Y.-J.; Chong, Q.-Y.; Wu, M.-M.; Pandey, V.; Chen, R.-M.; Liu, L.; Ma, L.; Wu, Z.-S.; Zhu, T.; et al. Trefoil factor 3 mediation of oncogenicity and chemoresistance in hepatocellular carcinoma is AKT-BCL-2 dependent. Oncotarget 2017, 8, 39323–39344. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wang, J.-N.; Tang, J.-M.; Kong, X.; Yang, J.-Y.; Zheng, F.; Guo, L.-Y.; Huang, Y.-Z.; Tian, L.; Cao, S.-F.; et al. VEGF is essential for the growth and migration of human hepatocellular carcinoma cells. Mol. Biol. Rep. 2011, 39, 5085–5093. [Google Scholar] [CrossRef] [Green Version]
- Marra, F.; Tacke, F. Roles for Chemokines in Liver Disease. Gastroenterology 2014, 147, 577–594.e1. [Google Scholar] [CrossRef]
- Nagoshi, S. Osteopontin: Versatile modulator of liver diseases. Hepatol. Res. 2013, 44, 22–30. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Zhang, J.; Fan, H.-N.; Zhu, J.-S. Function and therapeutic advances of chemokine and its receptor in nonalcoholic fatty liver disease. Ther. Adv. Gastroenterol. 2018, 11, 1756284818815184. [Google Scholar] [CrossRef] [PubMed]
- Ham, B.; Fernandez, M.C.; D’Costa, Z.; Brodt, P. The diverse roles of the TNF axis in cancer progression and metastasis. Trends Cancer Res. 2016, 11, 1–27. [Google Scholar]
- Leon-Ponte, M.; Kirchhof, M.G.; Sun, T.; Stephens, T.; Singh, B.; Sandhu, S.; Madrenas, J. Polycationic lipids inhibit the pro-inflammatory response to LPS. Immunol. Lett. 2005, 96, 73–83. [Google Scholar] [CrossRef]
- Zhao, Y.; Chen, Y.; Hu, Y.; Wang, J.; Xie, X.; He, G.; Chen, H.; Shao, Q.; Zeng, H.; Zhang, H. Genomic alterations across six hepatocellular carcinoma cell lines by panel-based sequencing. Transl. Cancer Res. 2018, 7, 231–239. [Google Scholar] [CrossRef]
- Bressac, B.; Galvin, K.M.; Liang, T.J.; Isselbacher, K.J.; Wands, J.R.; Ozturk, M. Abnormal structure and expression of p53 gene in human hepatocellular carcinoma. Proc. Natl. Acad. Sci. USA 1990, 87, 1973–1977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brito, A.; Abrantes, A.M.; Pinto-Costa, C.; Gomes, A.; Mamede, A.; Casalta-Lopes, J.; Gonçalves, A.C.; Sarmento-Ribeiro, A.B.; Tralhão, J.G.; Botelho, M.F. Hepatocellular Carcinoma and Chemotherapy: The Role of p53. Chemotherapy 2012, 58, 381–386. [Google Scholar] [CrossRef]
- Mohammad, N.; Singh, S.V.; Malvi, P.; Chaube, B.; Athavale, D.; Vanuopadath, M.; Nair, S.S.; Nair, B.; Bhat, M.K. Strategy to enhance efficacy of doxorubicin in solid tumor cells by methyl-beta-cyclodextrin: Involvement of p53 and Fas receptor ligand complex. Sci. Rep. 2015, 5, 11853. [Google Scholar] [CrossRef] [Green Version]
- Fischer, M. Conservation and divergence of the p53 gene regulatory network between mice and humans. Oncogene 2019, 38, 4095–4109. [Google Scholar] [CrossRef] [Green Version]
- Yan, W.-L.; Shen, K.-Y.; Tien, C.-Y.; Chen, Y.-A.; Liu, S.-J. Recent progress in GM-CSF-based cancer immunotherapy. Immunotherapy 2017, 9, 347–360. [Google Scholar] [CrossRef]
- Agupitan, A.D.; Neeson, P.J.; Williams, S.; Howitt, J.; Haupt, S.; Haupt, Y. P53: A Guardian of Immunity Becomes Its Saboteur through Mutation. Int. J. Mol. Sci. 2020, 21, 3452. [Google Scholar] [CrossRef]
- Zhou, S.-L.; Dai, Z.; Zhou, Z.-J.; Wang, X.-Y.; Yang, G.-H.; Wang, Z.; Huang, X.-W.; Fan, J.; Zhou, J. Overexpression of CXCL5 mediates neutrophil infiltration and indicates poor prognosis for hepatocellular carcinoma. Hepatology 2012, 56, 2242–2254. [Google Scholar] [CrossRef]
- Elzi, D.J.; Song, M.; Blackman, B.; Weintraub, S.T.; López-Terrada, L.; Chen, Y.; Tomlinson, G.E.; Shiio, Y. FGF19 functions as autocrine growth factor for hepatoblastoma. Genes Cancer 2016, 7, 125–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zucman-Rossi, J.; Villanueva, A.; Nault, J.-C.; Llovet, J.M. Genetic Landscape and Biomarkers of Hepatocellular Carcinoma. Gastroenterology 2015, 149, 1226–1239.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, C.; Chen, L.; Hu, H.; Ma, H.-Y.; Gao, L.-L.; Qin, J.; Zhong, C.-P. Chemokines and their receptors play important roles in the development of hepatocellular carcinoma. World J. Hepatol. 2015, 7, 1390–1402. [Google Scholar] [CrossRef]
- Guo, Q.; Yu, D.-Y.; Yang, Z.-F.; Liu, D.-Y.; Cao, H.-Q.; Liao, X.-W. IGFBP2 upregulates ZEB1 expression and promotes hepatocellular carcinoma progression through NF-κB signaling pathway. Dig. Liver Dis. 2020, 52, 573–581. [Google Scholar] [CrossRef]
- Bhat, M.; Skill, N.J.; Marcus, V.; Deschênes, M.; Tan, X.; Bouteaud, J.; Negi, S.; Awan, Z.; Aikin, R.; Kwan, J.; et al. Decreased PCSK9 expression in human hepatocellular carcinoma. BMC Gastroenterol. 2015, 15, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.; Zhou, S.; Zhan, Y.; Ke, J.; Wang, K.; Liang, Q.; Hou, Y.; Zhu, P.; Ao, W.; Wei, X.; et al. Dioscin Inhibits the Invasion and Migration of Hepatocellular Carcinoma HepG2 Cells by Reversing TGF-beta1-Induced Epithelial-Mesenchymal Transition. Molecules 2019, 24, 2222. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Haberl, E.M.; Pohl, R.; Rein-Fischboeck, L.; Feder, S.; Eisinger, K.; Krautbauer, S.; Sinal, C.J.; Buechler, C. Ex vivo analysis of serum chemerin activity in murine models of obesity. Cytokine 2018, 104, 42–45. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Hepa1-6 | Huh7 | HepG2 |
|---|---|---|---|
| Amphiregulin | x | - | - |
| Angiogenin | - | x | x |
| Apolipoprotein A1 | - | x | x |
| CCL2/Monocyte chemoattractant protein-1 | x | n.d. | n.d. |
| CCL5/Regulated upon activation, normal T cell expressed, and secreted | x | n.d. | n.d. |
| CCL17/Thymus and activation regulated chemokine | x | n.d. | n.d. |
| CCL20/Macrophage inflammatory protein-3 | x | x | x |
| CXCL1/KC/IL-8 | x | x | x |
| CXCL2/Macrophage inflammatory protein-2 | x | - | - |
| CXCL5/Epithelial-derived neutrophil-activating peptide 78 | x | x | n.d. |
| CXCL10/Interferon gamma-induced protein 10 | x | n.d. | n.d. |
| CXCL16 | x | - | - |
| Cystatin C | x | x | x |
| Dickkopf-1 | n.d. | x | x |
| Dipeptidylpeptidase | n.d. | x | x |
| Extracellular matrix metalloproteinase inducer | n.i. | x | x |
| Endostatin | x | n.i. | n.i. |
| Fibroblast growth factor-19 | n.i. | x | n.d. |
| Growth differentiation factor-15 | x | x | x |
| Granulocyte-macrophage colony-stimulating factor | x | n.d. | n.d. |
| Intercellular adhesion molecule 1 | x | n.d. | n.d. |
| IGF-binding protein (IGFBP)-1 | x | n.i. | n.i. |
| IGFBP-2 | n.d. | x | x |
| IGFBP-3 | x | n.d. | n.d. |
| IL-23 | x | n.d. | n.d. |
| Macrophage inflammatory protein | n.i. | x | x |
| Matrix metalloproteinase-3 | x | n.i. | n.i. |
| Osteopontin (OPN) | x | x | x |
| Osteoprotegerin/TNFRSF11B | x | n.i. | n.i. |
| Pentraxin 3 | n.d. | x | x |
| Proprotein convertase subtilisin/kexin type 9/PCSK9 | x | n.i. | n.i. |
| Retinol-binding protein 4 | x | x | x |
| Serpin E1/Plasminogen activator inhibitor-1 | x | x | x |
| Thrombospondin-1 | n.i. | x | x |
| Trefoil factor 3 | n.i. | n.d. | x |
| Tissue factor | x | n.i. | n.i. |
| Vascular endothelial growth factor | x | n.d. | x |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feder, S.; Bruckmann, A.; McMullen, N.; Sinal, C.J.; Buechler, C. Chemerin Isoform-Specific Effects on Hepatocyte Migration and Immune Cell Inflammation. Int. J. Mol. Sci. 2020, 21, 7205. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197205
Feder S, Bruckmann A, McMullen N, Sinal CJ, Buechler C. Chemerin Isoform-Specific Effects on Hepatocyte Migration and Immune Cell Inflammation. International Journal of Molecular Sciences. 2020; 21(19):7205. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197205
Chicago/Turabian StyleFeder, Susanne, Astrid Bruckmann, Nichole McMullen, Christopher J. Sinal, and Christa Buechler. 2020. "Chemerin Isoform-Specific Effects on Hepatocyte Migration and Immune Cell Inflammation" International Journal of Molecular Sciences 21, no. 19: 7205. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197205