Structural Diversity and Dynamics of Human Three-Finger Proteins Acting on Nicotinic Acetylcholine Receptors

 , ,
, ,
Abstract
:1. Introduction
2. Results
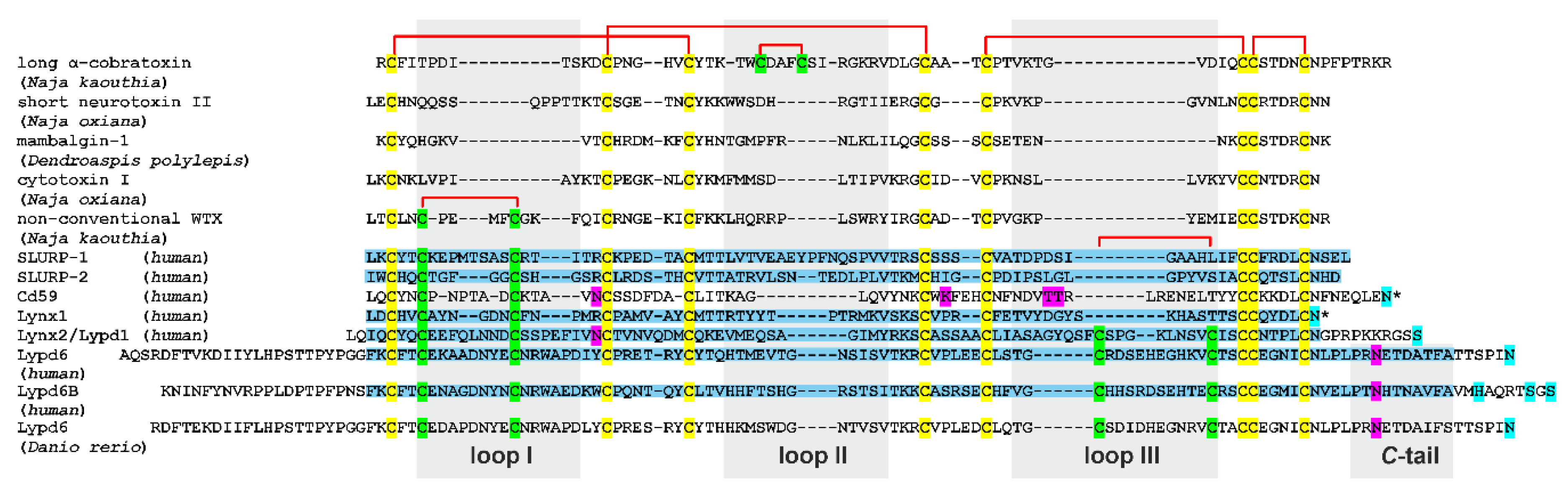
2.1. Design, Production, and Optimization of Conditions for NMR Study of Isolated LU-Domains
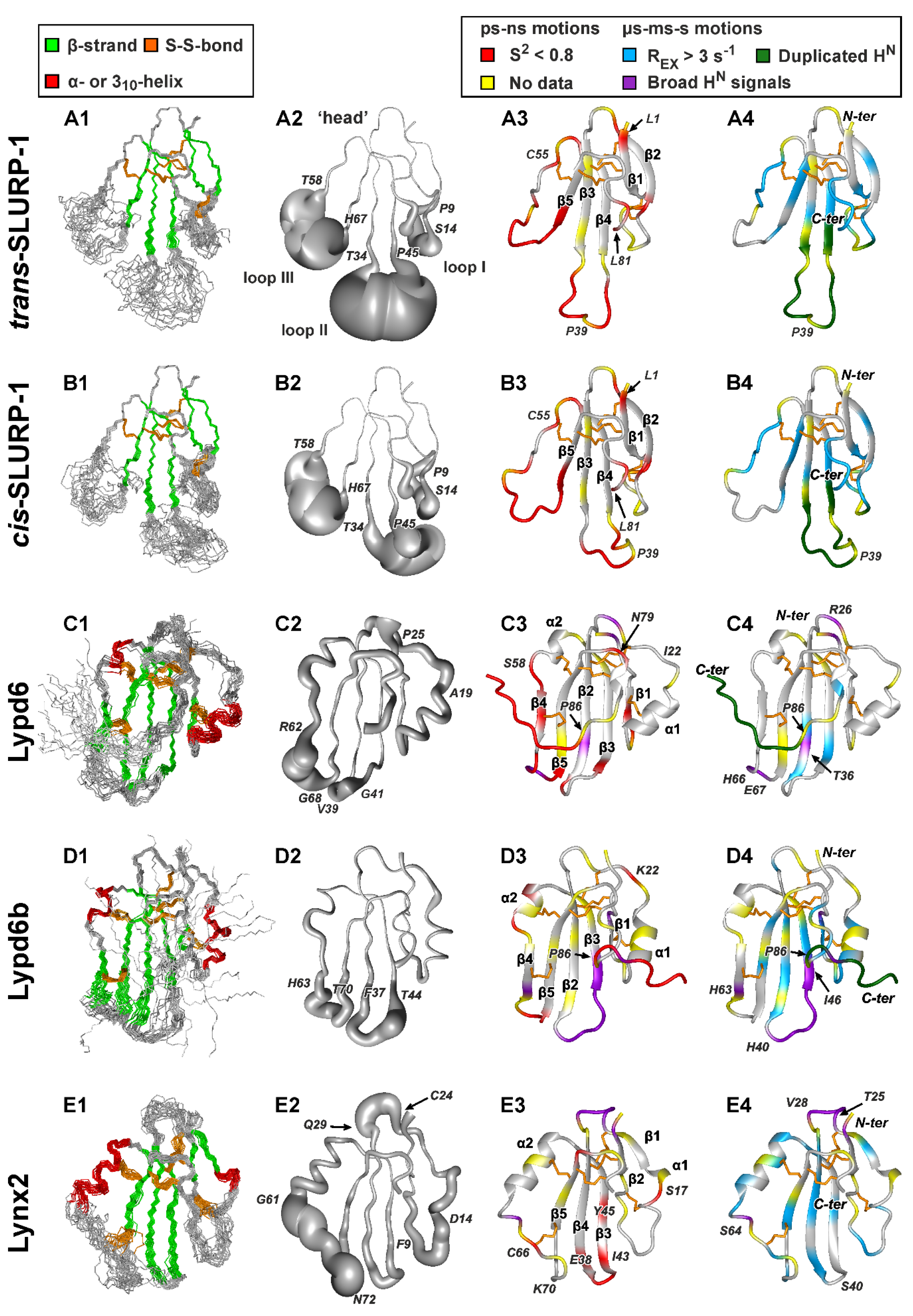
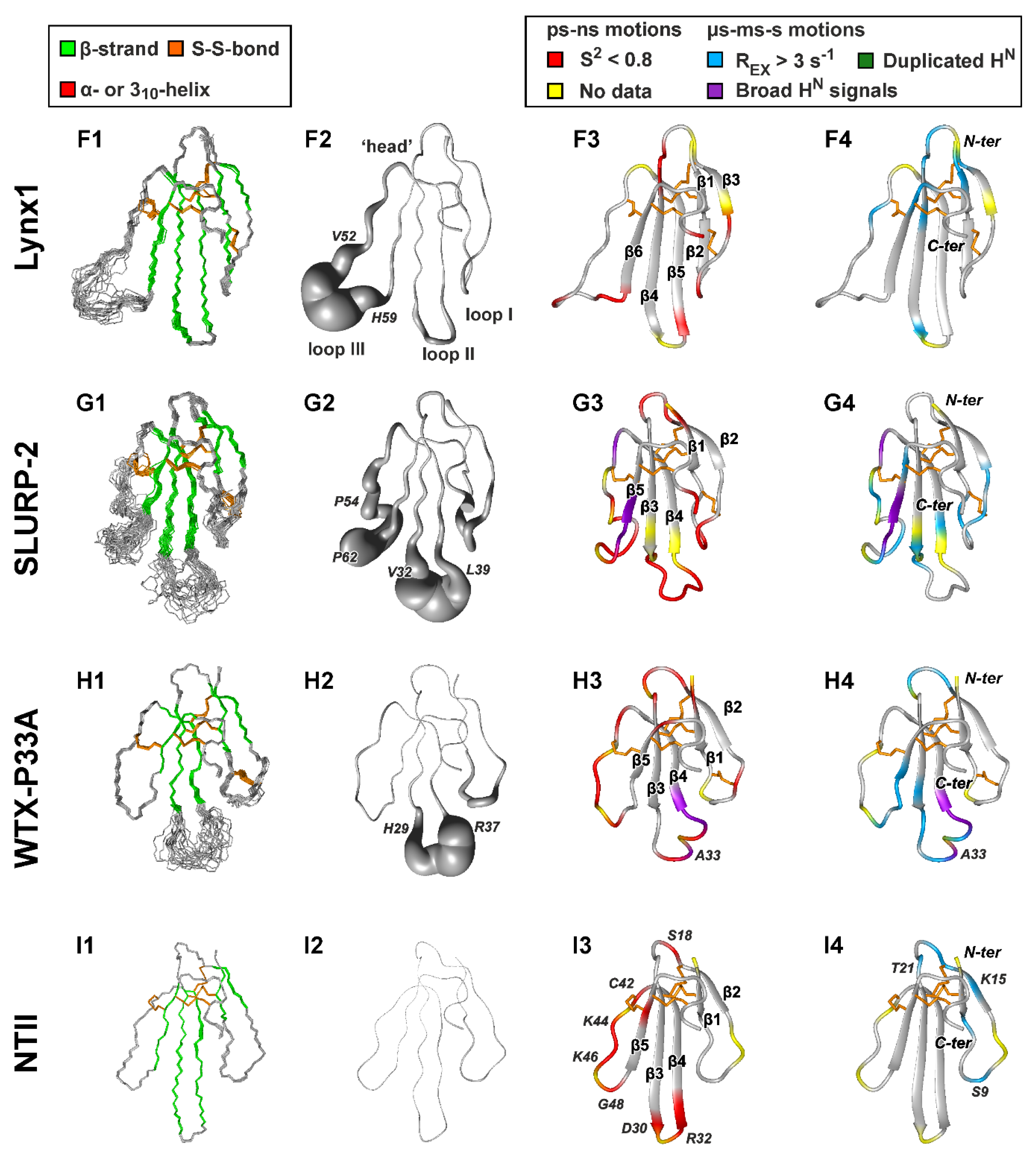
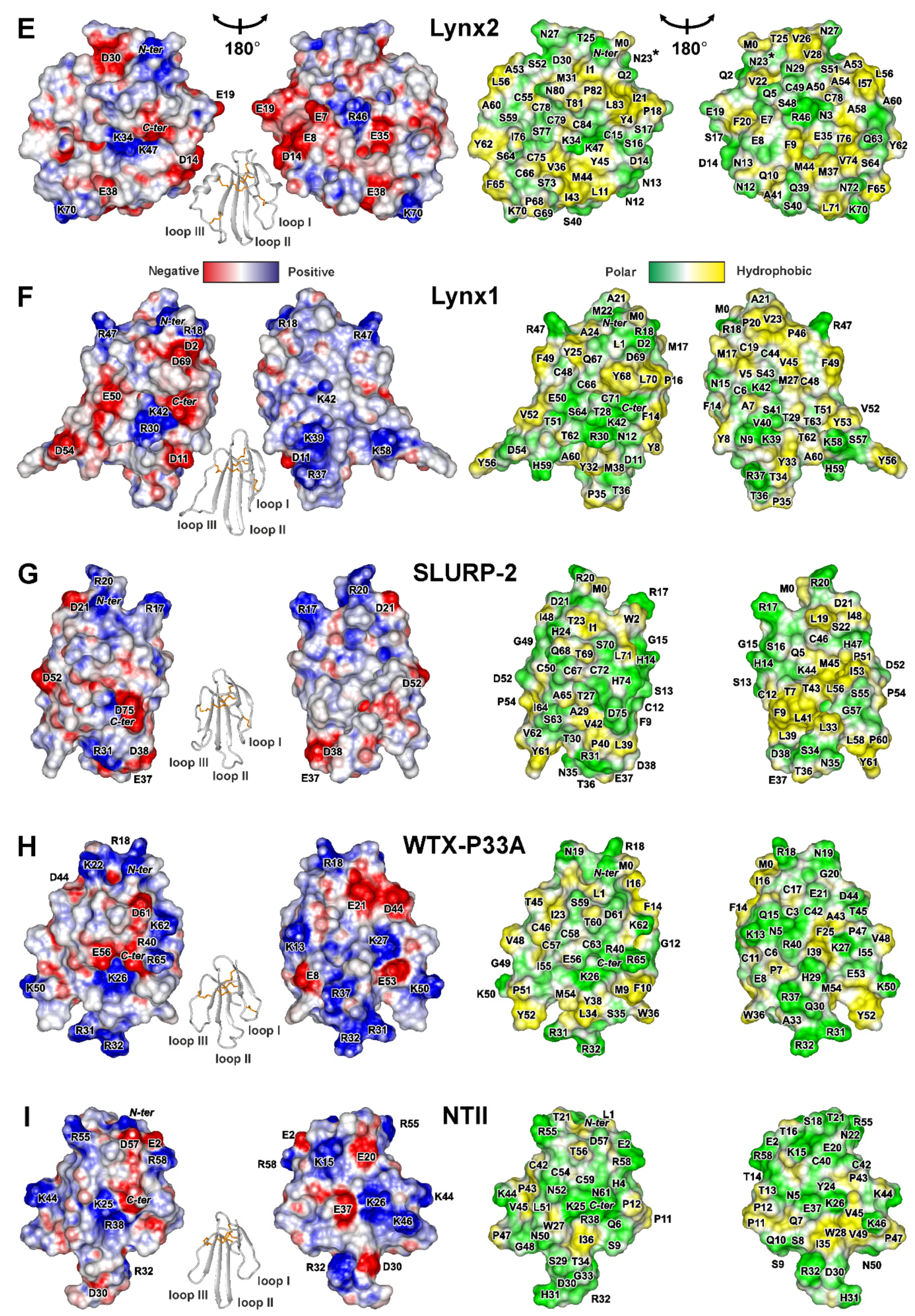
2.2. 3D Structures of Human TFPs
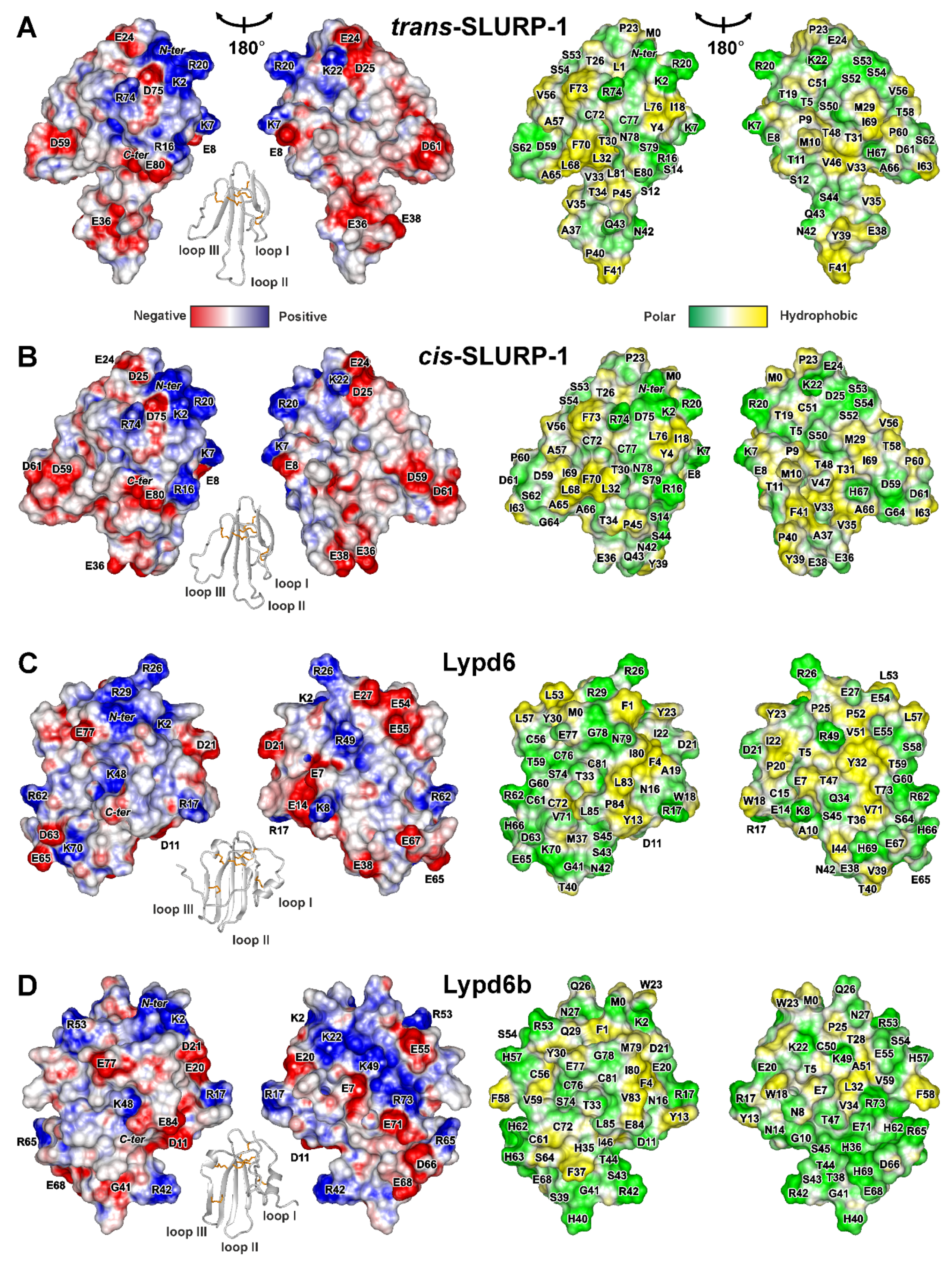
2.2.1. SLURP-1
2.2.2. Lypd6 and Lypd6b
2.2.3. Lynx2
2.2.4. Lynx1, SLURP-2, and Snake Toxins
2.3. Backbone Dynamics of TFPs in Solution
2.3.1. Overall Rotational Diffusion of TFPs
2.3.2. SLURP-1
2.3.3. Lypd6 and Lypd6b
2.3.4. Lynx2
2.3.5. Lynx1, SLURP-2, and Snake Toxins
3. Discussion
- (1)
- The ligand plasticity is required to adopt its structure for interaction with several molecular targets. Indeed, many of the studied proteins (WTX, Lynx1, Lypd6, and SLURP-2) can interact with nAChRs and other receptors, e.g., mAChRs or LPR6. In this case, the high plasticity of SLURP-1 suggests the presence of a molecular target other than α7-nAChR. Currently, this putative target remains uncharacterized.
- (2)
- The ligand plasticity is needed for adaptation to the plasticity of the nAChR binding sites. The binding sites of the receptor experience significant rearrangements during gating of the receptor ion channel [70]. Indeed, the endogenous human TFPs are not direct blockers or agonists of nAChR, but rather are modulators of the receptor [1]. They probably bind the receptor in different conformational states and do not prevent the movements of the receptor during gating, but rather influence populations of different states. The ability of human TFPs to interact with different functional states of nAChR is supported by the ability of Lynx1 and SLURP-2 to enhance or suppress ion currents through α7-nAChR [11,32].
- (3)
- The ligand plasticity is a mechanism, which regulates the affinity of ligand-receptor interaction via entropic term of the free energy (ΔG = ΔH − TΔS). Indeed, the binding of WTX-P33A to human M1 mAChR results in the significant decrease of the loops I and II mobility at the ps–ns timescale [43]. This causes a loss of conformational entropy, which makes formation of the ligand-receptor complex less energetically favorable. The simple calculations permit to estimate the magnitude of this effect [71]. An increase in the average S2 value for a 15-residue polypeptide from 0.7 to 0.85 upon the receptor binding leads to entropic penalty (−TΔS) ~+3.5 kcal/mol (~5.7·kB·T) at 30 °C (only change in backbone conformational entropy was taken into account). According to the Boltzmann equation, such ΔG difference can lead to the more than two order of magnitude difference in affinity (dissociation constant) between the ‘mobile’ and ‘stable’ protein ligands. In real situations, the unfavorable conformational entropy term is probably compensated by the favorable enthalpy and entropy terms due to better match between the ligand and receptor structures. For example, WTX and NTII have similar overall length (65 and 61 residues) and very close average S2 values (0.84 and 0.85, respectively). The larger mobility of the NTII loop III is compensated by larger mobility of the WTX loop I (Figure 3B). Despite this similarity, the affinity of toxins for muscle-type nAChRs is significantly different (IC50~3 µM and 30 nM for WTX and NTII, respectively) [62,72], indicating the involvement of structural factors other than intramolecular mobility. Nevertheless, the above calculations indicate that the change in conformational entropy of interacting molecules upon the complex formation cannot be neglected.
4. Materials and Methods
4.1. Recombinant Production and Samples Preparation
4.2. NMR Spectroscopy
4.3. Structure Calculation and Relaxation Data Analysis
4.4. Accessing Codes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Vasilyeva, N.A.; Loktyushov, E.V.; Bychkov, M.L.; Shenkarev, Z.O.; Lyukmanova, E.N. Three-Finger Proteins from the Ly6/uPAR Family: Functional Diversity within One Structural Motif. Biochemistry 2017, 82, 1702–1715. [Google Scholar] [CrossRef] [PubMed]
- Shabelnikov, S.V.; Bobkov, D.E.; Sharlaimova, N.S.; Petukhova, O.A. Injury affects coelomic fluid proteome of the common starfish, Asterias rubens. J. Exp. Biol. 2019, 222, jeb198556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, K.; Joiner, W.J.; Wu, M.N.; Yue, Z.; Smith, C.J.; Sehgal, A. Identification of SLEEPLESS, a sleep-promoting factor. Science 2008, 321, 372–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loughner, C.L.; Bruford, E.A.; McAndrews, M.S.; Delp, E.E.; Swamynathan, S.; Swamynathan, S.K. Organization, evolution and functions of the human and mouse Ly6/uPAR family genes. Hum. Genom. 2016, 10, 10. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, C.M.; Harrison, R.A.; Lachmann, P.J.; Neuhaus, D. Structure of a soluble, glycosylated form of the human complement regulatory protein CD59. Structure 1994, 2, 185–199. [Google Scholar] [CrossRef] [Green Version]
- Thomsen, M.S.; Cinar, B.; Jensen, M.M.; Lyukmanova, E.N.; Shulepko, M.A.; Tsetlin, V.; Klein, A.B.; Mikkelsen, J.D. Expression of the Ly-6 family proteins Lynx1 and Ly6H in the rat brain is compartmentalized, cell-type specific, and developmentally regulated. Brain Struct. Funct. 2014, 219, 1923–1934. [Google Scholar] [CrossRef]
- Jensen, M.M.; Arvaniti, M.; Mikkelsen, J.D.; Michalski, D.; Pinborg, L.H.; Härtig, W.; Thomsen, M.S. Prostate stem cell antigen interacts with nicotinic acetylcholine receptors and is affected in Alzheimer’s disease. Neurobiol. Aging 2015, 36, 1629–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, S.-C.; Lin, C.-W.; Yang, W.-E.; Fan, W.-L.; Yang, S.-F. The urokinase-type plasminogen activator (uPA) system as a biomarker and therapeutic target in human malignancies. Expert Opin. Ther. Targets 2016, 20, 551–566. [Google Scholar] [CrossRef]
- Egerod, K.L.; Holst, B.; Petersen, P.S.; Hansen, J.B.; Mulder, J.; Hökfelt, T.; Schwartz, T.W. GPR39 Splice Variants Versus Antisense Gene LYPD1: Expression and Regulation in Gastrointestinal Tract, Endocrine Pancreas, Liver, and White Adipose Tissue. Mol. Endocrinol. 2007, 21, 1685–1698. [Google Scholar] [CrossRef]
- Ibañez-Tallon, I.; Miwa, J.M.; Wang, H.L.; Adams, N.C.; Crabtree, G.W.; Sine, S.M.; Heintz, N. Novel modulation of neuronal nicotinic acetylcholine receptors by association with the endogenous prototoxin lynx1. Neuron 2002, 33, 893–903. [Google Scholar] [CrossRef] [Green Version]
- Lyukmanova, E.N.; Shenkarev, Z.O.; Shulepko, M.A.; Mineev, K.S.; D’Hoedt, D.; Kasheverov, I.E.; Filkin, S.Y.; Krivolapova, A.P.; Janickova, H.; Dolezal, V.; et al. NMR structure and action on nicotinic acetylcholine receptors of water-soluble domain of human LYNX1. J. Biol. Chem. 2011, 286, 10618–10627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nichols, W.A.; Henderson, B.J.; Yu, C.; Parker, R.L.; Richards, C.I.; Lester, H.A.; Miwa, J.M. Lynx1 shifts α4β2 nicotinic receptor subunit stoichiometry by affecting assembly in the endoplasmic reticulum. J. Biol. Chem. 2014, 289, 31423–31432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morishita, H.; Miwa, J.M.; Heintz, N.; Hensch, T.K. Lynx1, a cholinergic brake, limits plasticity in adult visual cortex. Science 2010, 330, 1238–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miwa, J.M.; Walz, A. Enhancement in motor learning through genetic manipulation of the Lynx1 gene. PLoS ONE 2012, 7, e43302. [Google Scholar] [CrossRef] [PubMed]
- Shenkarev, Z.O.; Shulepko, M.A.; Bychkov, M.L.; Kulbatskii, D.S.; Shlepova, O.V.; Vasilyeva, N.A.; Andreev-Andrievskiy, A.A.; Popova, A.S.; Lagereva, E.A.; Loktyushov, E.V.; et al. Water-soluble variant of human Lynx1 positively modulates synaptic plasticity and ameliorates cognitive impairment associated with α7-nAChR dysfunction. J. Neurochem. 2020. [Google Scholar] [CrossRef]
- Dessaud, E.; Salaün, D.; Gayet, O.; Chabbert, M.; deLapeyrière, O. Identification of lynx2, a novel member of the ly-6/neurotoxin superfamily, expressed in neuronal subpopulations during mouse development. Mol. Cell Neurosci. 2006, 31, 232–242. [Google Scholar] [CrossRef]
- Tekinay, A.B.; Nong, Y.; Miwa, J.M.; Lieberam, I.; Ibanez-Tallon, I.; Greengard, P.; Heintz, N. A role for LYNX2 in anxiety-related behavior. Proc. Natl. Acad. Sci. USA 2009, 106, 4477–4482. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Puddifoot, C.A.; Taylor, P.; Joiner, W.J. Mechanisms of inhibition and potentiation of α4β2 nicotinic acetylcholine receptors by members of the Ly6 protein family. J. Biol. Chem. 2015, 290, 24509–24518. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.W.; Song, P.F.; Spindel, E.R. Role of Lynx1 and related Ly6 proteins as modulators of cholinergic signaling in normal and neoplastic bronchial epithelium. Int. Immunopharmacol. 2015, 29, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Bychkov, M.; Shenkarev, Z.; Shulepko, M.; Shlepova, O.; Kirpichnikov, M.; Lyukmanova, E. Water-soluble variant of human Lynx1 induces cell cycle arrest and apoptosis in lung cancer cells via modulation of α7 nicotinic acetylcholine receptors. PLoS ONE 2019, 14, e0217339. [Google Scholar] [CrossRef]
- Shulepko, M.A.; Kulbatskii, D.S.; Bychkov, M.L.; Lyukmanova, E.N. Human Nicotinic Acetylcholine Receptors: Part II. Non-Neuronal Cholinergic System. Russ. J. Bioorganic Chem. 2019, 45. [Google Scholar] [CrossRef]
- Darvas, M.; Morsch, M.; Racz, I.; Ahmadi, S.; Swandulla, D.; Zimmer, A. Modulation of the Ca2+ conductance of nicotinic acetylcholine receptors by Lypd6. Eur. Neuropsychopharmacol. 2009, 19, 670–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arvaniti, M.; Jensen, M.M.; Soni, N.; Wang, H.; Klein, A.B.; Thiriet, N.; Pinborg, L.H.; Muldoon, P.P.; Wienecke, J.; Imad Damaj, M.; et al. Functional interaction between Lypd6 and nicotinic acetylcholine receptors. J. Neurochem. 2016, 138, 806–820. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lang, Q.; Li, J.; Xie, F.; Wan, B.; Yu, L. Identification and characterization of human LYPD6, a new member of the Ly-6 superfamily. Mol. Biol. Rep. 2010, 37, 2055–2062. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Lang, Q.; Bai, M.; Zhong, C.; Chen, X.; Wan, B.; Yu, L. Cloning and characterization of a human LYPD7, a new member of the Ly-6 superfamily. Mol. Biol. Rep. 2009, 36, 697–703. [Google Scholar] [CrossRef]
- Özhan, G.; Sezgin, E.; Wehner, D.; Pfister, A.S.; Kühl, S.J.; Kagermeier-Schenk, B.; Kühl, M.; Schwille, P.; Weidinger, G. Lypd6 enhances Wnt/β-catenin signaling by promoting Lrp6 phosphorylation in raft plasma membrane domains. Dev. Cell 2013, 26, 331–345. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Ren, J.; Lu, W.; Harlos, K.; Jones, E.Y. Structure of the Wnt signaling enhancer LYPD6 and its interactions with the Wnt coreceptor LRP6. FEBS Lett. 2018. [Google Scholar] [CrossRef]
- Arvaniti, M.; Polli, F.S.; Kohlmeier, K.A.; Thomsen, M.S.; Andreasen, J.T. Loss of Lypd6 leads to reduced anxiety-like behaviour and enhanced responses to nicotine. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 82, 86–94. [Google Scholar] [CrossRef]
- Ochoa, V.; George, A.A.; Nishi, R.; Whiteaker, P. The prototoxin LYPD6B modulates heteromeric α3β4-containing nicotinic acetylcholine receptors, but not α7 homomers. FASEB J. 2016, 30, 1109–1119. [Google Scholar] [CrossRef] [Green Version]
- Arredondo, J.; Chernyavsky, A.I.; Grando, S.A. SLURP-1 and -2 in normal, immortalized and malignant oral keratinocytes. Life Sci. 2007, 80, 2243–2247. [Google Scholar] [CrossRef] [Green Version]
- Lyukmanova, E.N.; Shulepko, M.A.; Kudryavtsev, D.; Bychkov, M.L.; Kulbatskii, D.S.; Kasheverov, I.E.; Astapova, M.V.; Feofanov, A.V.; Thomsen, M.S.; Mikkelsen, J.D.; et al. Human Secreted Ly-6/uPAR Related Protein-1 (SLURP-1) Is a Selective Allosteric Antagonist of α7 Nicotinic Acetylcholine Receptor. PLoS ONE 2016, 11, e0149733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyukmanova, E.N.; Shulepko, M.A.; Shenkarev, Z.O.; Bychkov, M.L.; Paramonov, A.S.; Chugunov, A.O.; Kulbatskii, D.S.; Arvaniti, M.; Dolejsi, E.; Schaer, T.; et al. Secreted Isoform of Human Lynx1 (SLURP-2): Spatial Structure and Pharmacology of Interactions with Different Types of Acetylcholine Receptors. Sci. Rep. 2016, 6, 30698. [Google Scholar] [CrossRef] [PubMed]
- Arredondo, J.; Chernyavsky, A.I.; Webber, R.J.; Grando, S.A. Biological effects of SLURP-1 on human keratinocytes. J. Invest. Dermatol. 2005, 125, 1236–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chernyavsky, A.I.; Kalantari-Dehaghi, M.; Phillips, C.; Marchenko, S.; Grando, S.A. Novel cholinergic peptides SLURP-1 and -2 regulate epithelialization of cutaneous and oral wounds. Wound Repair Regen. 2012, 20, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Arredondo, J.; Chernyavsky, A.I.; Grando, S.A. Overexpression of SLURP-1 and -2 alleviates the tumorigenic action of tobacco-derived nitrosamine on immortalized oral epithelial cells. Biochem. Pharmacol. 2007, 74, 1315–1319. [Google Scholar] [CrossRef] [Green Version]
- Lyukmanova, E.N.; Bychkov, M.L.; Sharonov, G.V.; Efremenko, A.V.; Shulepko, M.A.; Kulbatskii, D.S.; Shenkarev, Z.O.; Feofanov, A.V.; Dolgikh, D.A.; Kirpichnikov, M.P. Human Secreted Proteins SLURP-1 and SLURP-2 Control the Growth of Epithelial Cancer Cells via interaction with Nicotinic Acetylcholine Receptors. Br. J. Pharmacol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Chernyavsky, A.I.; Galitovskiy, V.; Shchepotin, I.B.; Grando, S.A. Anti-inflammatory effects of the nicotinergic peptides SLURP-1 and SLURP-2 on human intestinal epithelial cells and immunocytes. Biomed. Res. Int. 2014, 2014, 609086. [Google Scholar] [CrossRef] [Green Version]
- Bergqvist, C.; Kadara, H.; Hamie, L.; Nemer, G.; Safi, R.; Karouni, M.; Marrouche, N.; Abbas, O.; Hasbani, D.J.; Kibbi, A.G.; et al. SLURP-1 is mutated in Mal de Meleda, a potential molecular signature for melanoma and a putative squamous lineage tumor suppressor gene. Int. J. Dermatol. 2018, 57, 162–170. [Google Scholar] [CrossRef]
- Perez, C.; Khachemoune, A. Mal de Meleda: A Focused Review. Am. J. Clin. Dermatol. 2016, 17, 63–70. [Google Scholar] [CrossRef]
- Allan, C.M.; Procaccia, S.; Tran, D.; Tu, Y.; Barnes, R.H.; Larsson, M.; Allan, B.B.; Young, L.C.; Hong, C.; Tontonoz, P.; et al. Palmoplantar Keratoderma in Slurp2-Deficient Mice. J. Invest. Dermatol. 2016, 136, 436–443. [Google Scholar] [CrossRef] [Green Version]
- Shulepko, M.A.; Lyukmanova, E.N.; Shenkarev, Z.O.; Dubovskii, P.V.; Astapova, M.V.; Feofanov, A.V.; Arseniev, A.S.; Utkin, Y.N.; Kirpichnikov, M.P.; Dolgikh, D.A. Towards universal approach for bacterial production of three-finger Ly6/uPAR proteins: Case study of cytotoxin I from cobra N. oxiana. Protein Expr. Purif. 2017, 130, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Paramonov, A.S.; Kulbatskii, D.S.; Loktyushov, E.V.; Tsarev, A.V.; Dolgikh, D.A.; Shenkarev, Z.O.; Kirpichnikov, M.P.; Lyukmanova, E.N. Recombinant production and structural studies of the human Lypd6 and Lypd6b proteins. Russ. J. Bioorg. Chem. 2017, 43, 644–652. [Google Scholar] [CrossRef]
- Lyukmanova, E.N.; Shenkarev, Z.O.; Shulepko, M.A.; Paramonov, A.S.; Chugunov, A.O.; Janickova, H.; Dolejsi, E.; Dolezal, V.; Utkin, Y.N.; Tsetlin, V.I.; et al. Structural Insight into Specificity of Interactions between Nonconventional Three-finger Weak Toxin from Naja kaouthia (WTX) and Muscarinic Acetylcholine Receptors. J. Biol. Chem. 2015, 290, 23616–23630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesovoy, D.M.; Dubinnyi, M.A.; Nolde, S.B.; Bocharov, E.V.; Arseniev, A.S. Accurate measurement of dipole/dipole transverse cross-correlated relaxation [Formula: See text] in methylenes and primary amines of uniformly [Formula: See text]-labeled proteins. J. Biomol. NMR 2019, 73, 245–260. [Google Scholar] [CrossRef] [PubMed]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef] [Green Version]
- Shulepko, M.A.; Lyukmanova, E.N.; Paramonov, A.S.; Lobas, A.A.; Shenkarev, Z.O.; Kasheverov, I.E.; Dolgikh, D.A.; Tsetlin, V.I.; Arseniev, A.S.; Kirpichnikov, M.P. Human neuromodulator SLURP-1: Bacterial expression, binding to muscle-type nicotinic acetylcholine receptor, secondary structure, and conformational heterogeneity in solution. Biochemistry 2013, 78, 204–211. [Google Scholar] [CrossRef]
- Lambers, H.; Piessens, S.; Bloem, A.; Pronk, H.; Finkel, P. Natural skin surface pH is on average below 5, which is beneficial for its resident flora. Int. J. Cosmet. Sci. 2006, 28, 359–370. [Google Scholar] [CrossRef]
- Paramonov, A.S.; Shulepko, M.A.; Kocharovskaya, M.V.; Alenkin, A.E.; Evdokimova, A.O.; Akentiev, P.I.; Shenkarev, Z.O.; Kirpichnikov, M.P.; Lyukmanova, E.N. Bacterial production and structural study of human neuromodulator Lynx2. Russ. J. Bioorg. Chem. 2020, 46. accepted. [Google Scholar]
- Korzhnev, D.M.; Billeter, M.; Arseniev, A.S.; Orekhov, V.Y. NMR studies of Brownian tumbling and internal motions in proteins. Prog. Nucl. Magn. Reson. Spectrosc. 2001, 38, 197–266. [Google Scholar] [CrossRef]
- Kneller, J.M.; Lu, M.; Bracken, C. An effective method for the discrimination of motional anisotropy and chemical exchange. J. Am. Chem. Soc. 2002, 124, 1852–1853. [Google Scholar] [CrossRef]
- Guenneugues, M.; Drevet, P.; Pinkasfeld, S.; Gilquin, B.; Ménez, A.; Zinn-Justin, S. Picosecond to hour time scale dynamics of a “three finger” toxin: Correlation with its toxic and antigenic properties. Biochemistry 1997, 36, 16097–16108. [Google Scholar] [CrossRef] [PubMed]
- García de la Torre, J.; Huertas, M.L.; Carrasco, B. HYDRONMR: Prediction of NMR relaxation of globular proteins from atomic-level structures and hydrodynamic calculations. J. Magn. Reson. 2000, 147, 138–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, R.; Loria, J.P. FAST-Modelfree: A program for rapid automated analysis of solution NMR spin-relaxation data. J. Biomol. NMR 2003, 26, 203–213. [Google Scholar] [CrossRef]
- Taly, A.; Corringer, P.-J.; Guedin, D.; Lestage, P.; Changeux, J.-P. Nicotinic receptors: Allosteric transitions and therapeutic targets in the nervous system. Nat. Rev. Drug Discov. 2009, 8, 733–750. [Google Scholar] [CrossRef]
- Wessler, I.; Kirkpatrick, C.J. Acetylcholine beyond neurons: The non-neuronal cholinergic system in humans: Non-neuronal cholinergic system in humans. Br. J. Pharmacol. 2009, 154, 1558–1571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Echeverria, V.; Yarkov, A.; Aliev, G. Positive modulators of the α7 nicotinic receptor against neuroinflammation and cognitive impairment in Alzheimer’s disease. Prog. Neurobiol. 2016, 144, 142–157. [Google Scholar] [CrossRef]
- Wallace, T.L.; Bertrand, D. Neuronal α7 Nicotinic Receptors as a Target for the Treatment of Schizophrenia. Int. Rev. Neurobiol. 2015, 124, 79–111. [Google Scholar] [CrossRef]
- Philip, N.S.; Carpenter, L.L.; Tyrka, A.R.; Price, L.H. Nicotinic acetylcholine receptors and depression: A review of the preclinical and clinical literature. Psychopharmacology 2010, 212, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Shelukhina, I.V.; Zhmak, M.N.; Lobanov, A.V.; Ivanov, I.A.; Garifulina, A.I.; Kravchenko, I.N.; Rasskazova, E.A.; Salmova, M.A.; Tukhovskaya, E.A.; Rykov, V.A.; et al. Azemiopsin, a Selective Peptide Antagonist of Muscle Nicotinic Acetylcholine Receptor: Preclinical Evaluation as a Local Muscle Relaxant. Toxins 2018, 10, 34. [Google Scholar] [CrossRef] [Green Version]
- Stegemann, A.; Flis, D.; Ziolkowski, W.; Distler, J.H.W.; Steinbrink, K.; Böhm, M. The α7 Nicotinic Acetylcholine Receptor: A Promising Target for the Treatment of Fibrotic Skin Disorders. J. Investig. Dermatol. 2020. [Google Scholar] [CrossRef]
- Antil, S.; Servent, D.; Ménez, A. Variability among the sites by which curaremimetic toxins bind to torpedo acetylcholine receptor, as revealed by identification of the functional residues of alpha-cobratoxin. J. Biol. Chem. 1999, 274, 34851–34858. [Google Scholar] [CrossRef] [Green Version]
- Lyukmanova, E.N.; Shulepko, M.A.; Shenkarev, Z.O.; Kasheverov, I.E.; Chugunov, A.O.; Kulbatskii, D.S.; Myshkin, M.Y.; Utkin, Y.N.; Efremov, R.G.; Tsetlin, V.I.; et al. Central loop of non-conventional toxin WTX from Naja kaouthia is important for interaction with nicotinic acetylcholine receptors. Toxicon 2016, 119, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Teng, J.; Worrell, B.T.; Noviello, C.M.; Lee, M.; Karlin, A.; Stowell, M.H.B.; Hibbs, R.E. Structure of the Native Muscle-type Nicotinic Receptor and Inhibition by Snake Venom Toxins. Neuron 2020, 106, 952–962.e5. [Google Scholar] [CrossRef] [PubMed]
- Spurny, R.; Debaveye, S.; Farinha, A.; Veys, K.; Vos, A.M.; Gossas, T.; Atack, J.; Bertrand, S.; Bertrand, D.; Danielson, U.H.; et al. Molecular blueprint of allosteric binding sites in a homologue of the agonist-binding domain of the α7 nicotinic acetylcholine receptor. Proc. Natl. Acad. Sci. USA 2015, 112, E2543–E2552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eletsky, A.V.; Maslennikov, I.V.; Kukhtina, V.V.; Utkin, Y.N.; Tsetlin, V.I.; Arseniev, A.S. Structure and Conformational Heterogeneity of a Weak Toxin from the Cobra Naja kaouthia Venom. Russ. J. Bioorganic Chem. 2001, 27, 72–83. [Google Scholar] [CrossRef]
- Mordvintsev, D.Y.; Polyak, Y.L.; Rodionov, D.I.; Jakubik, J.; Dolezal, V.; Karlsson, E.; Tsetlin, V.I.; Utkin, Y.N. Weak toxin WTX from Naja kaouthia cobra venom interacts with both nicotinic and muscarinic acetylcholine receptors: Weak toxin binds two acetylcholine receptor types. FEBS J. 2009, 276, 5065–5075. [Google Scholar] [CrossRef]
- Pyrkov, T.V.; Chugunov, A.O.; Krylov, N.A.; Nolde, D.E.; Efremov, R.G. PLATINUM: A web tool for analysis of hydrophobic/hydrophilic organization of biomolecular complexes. Bioinformatics 2009, 25, 1201–1202. [Google Scholar] [CrossRef]
- Lupyan, D.; Leo-Macias, A.; Ortiz, A.R. A new progressive-iterative algorithm for multiple structure alignment. Bioinformatics 2005, 21, 3255–3263. [Google Scholar] [CrossRef]
- Buchner, L.; Güntert, P. Increased Reliability of Nuclear Magnetic Resonance Protein Structures by Consensus Structure Bundles. Structure 2015, 23, 425–434. [Google Scholar] [CrossRef] [Green Version]
- Bouzat, C.; Sine, S.M. Nicotinic acetylcholine receptors at the single-channel level. Br. J. Pharmacol. 2018, 175, 1789–1804. [Google Scholar] [CrossRef]
- Yang, D.; Kay, L.E. Contributions to conformational entropy arising from bond vector fluctuations measured from NMR-derived order parameters: Application to protein folding. J. Mol. Biol. 1996, 263, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Tsetlin, V.I.; Karlsson, E.; Arseniev, A.S.; Utkin, Y.N.; Surin, A.M.; Pashkov, V.S.; Pluzhnikov, K.A.; Ivanov, V.T.; Bystrov, V.F.; Ovchinnikov, Y.A. EPR And fluorescence study of interaction of Naja naja oxiana neurotoxin II and its derivatives with acetylcholine receptor protein from Torpedo marmorata. FEBS Lett. 1979, 106, 47–52. [Google Scholar] [CrossRef] [Green Version]
- Kazimierczuk, K.; Orekhov, V.Y. Accelerated NMR spectroscopy by using compressed sensing. Angew. Chem. Int. Ed. Engl. 2011, 50, 5556–5559. [Google Scholar] [CrossRef] [PubMed]
- Sattler, M.; Schleucher, J.; Griesinger, C. Heteronuclear multidimensional NMR experiments for the structure determination of proteins in solution employing pulsed field gradients. Prog. Nucl. Magn. Reson. Spectrosc. 1999, 34, 93–158. [Google Scholar] [CrossRef]
- Marion, D.; Driscoll, P.C.; Kay, L.E.; Wingfield, P.T.; Bax, A.; Gronenborn, A.M.; Clore, G.M. Overcoming the overlap problem in the assignment of proton NMR spectra of larger proteins by use of three-dimensional heteronuclear proton-nitrogen-15 Hartmann-Hahn-multiple quantum coherence and nuclear Overhauser-multiple quantum coherence spectroscopy: Application to interleukin 1.beta. Biochemistry 1989, 28, 6150–6156. [Google Scholar] [CrossRef]
- Bax, A.; Vuister, G.W.; Grzesiek, S.; Delaglio, F.; Wang, A.C.; Tschudin, R.; Zhu, G. Measurement of homo- and heteronuclear J couplings from quantitative J correlation. Meth. Enzymol. 1994, 239, 79–105. [Google Scholar] [CrossRef]
- Findeisen, M.; Brand, T.; Berger, S. A 1H-NMR thermometer suitable for cryoprobes. Magn. Reson. Chem. 2007, 45, 175–178. [Google Scholar] [CrossRef]
- Shen, Y.; Bax, A. Protein backbone and sidechain torsion angles predicted from NMR chemical shifts using artificial neural networks. J. Biomol. NMR 2013, 56, 227–241. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, E.; Güntert, P. Automated structure determination from NMR spectra. Methods Mol. Biol. 2015, 1261, 303–329. [Google Scholar] [CrossRef]
- Koradi, R.; Billeter, M.; Wüthrich, K. MOLMOL: A program for display and analysis of macromolecular structures. J. Mol. Graph. 1996, 14, 51–55. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | pI | Charges (+/−/His) a | HI b | Labeling | Conc., mM | pH | T, °C | Freq., MHz c | PDB | Ref |
|---|---|---|---|---|---|---|---|---|---|---|
| SLURP-1 | 5.2 | 8/10/1 | 0.04 | 13C, 15N | 0.3 | 4.7 | 37 | 800 | 6ZZE 6ZZF | This work |
| Lypd6 | 5.3 | 11/15/3 | −0.58 | 13C, 15N/15N d | 0.1 | 7.0 | 30/37 d | 600/800 | 6IB6 | This work |
| Lypd6b | 6.7 | 10/11/8 | −0.68 | 13C, 15N | 0.2 | 5.5 | 30/37 d | 600/800 | 6ZSO | This work |
| Lynx2 | 4.5 | 5/8/0 | 0.07 | 13C, 15N | 0.25/0.07 d | 6.7 | 45/37 d | 600/800 | 6ZSS | This work |
| Lynx1 | 8.1 | 8/6/2 | −0.32 | 15N | 0.5 | 5.3 | 25 | 800 | 2L03 | [11] |
| SLURP-2 | 6.5 | 5/6/5 | 0.10 | 13C, 15N | 0.5 e/0.08 d | 4.8 | 37 | 600 | 2N99 | [32] |
| WTX-P33A | 8.9 | 13/7/1 | −0.39 | 15N | 0.5 | 3.0 | 30 | 700 | 2MJ0 | [43] |
| NTII | 8.7 | 10/6/2 | −1.10 | 15N | 1.0 | 5.0 | 30 | 800 | 2MJ4 | [44] |
| Protein | T, °C | Theoretical a | Experimental b | |||
|---|---|---|---|---|---|---|
| τR, ns | Anisotropy 2Dz/(Dx + Dy) | Asymmetry Dx/Dy | τR, ns (iso/axial) | Anisotropy 2Dz/(Dx + Dy) | ||
| trans-SLURP-1 | 37 | 5.0 | 1.56 | 1.06 | 4.3/4.9 | 1.50 |
| cis-SLURP-1 | 37 | 5.1 | 1.44 | 1.11 | 4.3/4.8 | 1.78 |
| Lypd6 | 37 | 6.1 | 1.30 | 1.10 | 4.6/4.6 | 1.26 |
| Lypd6b | 37 | 6.1 | 1.36 | 1.11 | 5.0/5.0 | 1.32 |
| Lynx2 | 37 | 3.7 | 1.25 | 1.17 | 4.0/3.8 | 1.35 |
| Lynx1 | 25 | 6.1 | 1.56 | 1.14 | 5.0/5.2 | 1.57 |
| SLURP-2 | 37 | 4.0 | 1.60 | 1.01 | 4.0/4.1 | 1.42 |
| WTX-P33A | 30 | 3.7 | 1.49 | 1.06 | 3.6/3.6 | 1.34 |
| NTII | 30 | 3.7 | 1.50 | 1.08 | 2.9/3.0 | 1.42 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paramonov, A.S.; Kocharovskaya, M.V.; Tsarev, A.V.; Kulbatskii, D.S.; Loktyushov, E.V.; Shulepko, M.A.; Kirpichnikov, M.P.; Lyukmanova, E.N.; Shenkarev, Z.O. Structural Diversity and Dynamics of Human Three-Finger Proteins Acting on Nicotinic Acetylcholine Receptors. Int. J. Mol. Sci. 2020, 21, 7280. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197280
Paramonov AS, Kocharovskaya MV, Tsarev AV, Kulbatskii DS, Loktyushov EV, Shulepko MA, Kirpichnikov MP, Lyukmanova EN, Shenkarev ZO. Structural Diversity and Dynamics of Human Three-Finger Proteins Acting on Nicotinic Acetylcholine Receptors. International Journal of Molecular Sciences. 2020; 21(19):7280. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197280
Chicago/Turabian StyleParamonov, Alexander S., Milita V. Kocharovskaya, Andrey V. Tsarev, Dmitrii S. Kulbatskii, Eugene V. Loktyushov, Mikhail A. Shulepko, Mikhail P. Kirpichnikov, Ekaterina N. Lyukmanova, and Zakhar O. Shenkarev. 2020. "Structural Diversity and Dynamics of Human Three-Finger Proteins Acting on Nicotinic Acetylcholine Receptors" International Journal of Molecular Sciences 21, no. 19: 7280. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197280