Role of Human Leukocyte Antigen System as A Predictive Biomarker for Checkpoint-Based Immunotherapy in Cancer Patients

 , , , , , and
, , , , , and
Abstract
:1. Human Leucocyte Antigen and Antigen Presentation Machinery Molecules: An Overview
1.1. HLA Class I: Structure and Function
1.2. HLA Class I Antigen Processing Machinery Complex and Antigen Presentation
1.3. HLA Class II: Structure and Functions
1.4. HLA Class II Antigen Presentation System: How it Works
1.5. HLA Class I and II Transcription Regulation
1.6. HLA Class III: A Poorly Characterized Class
1.7. Carcinoma Cells as Non-Professional APC: A Novel Role for HLA Class II Complex
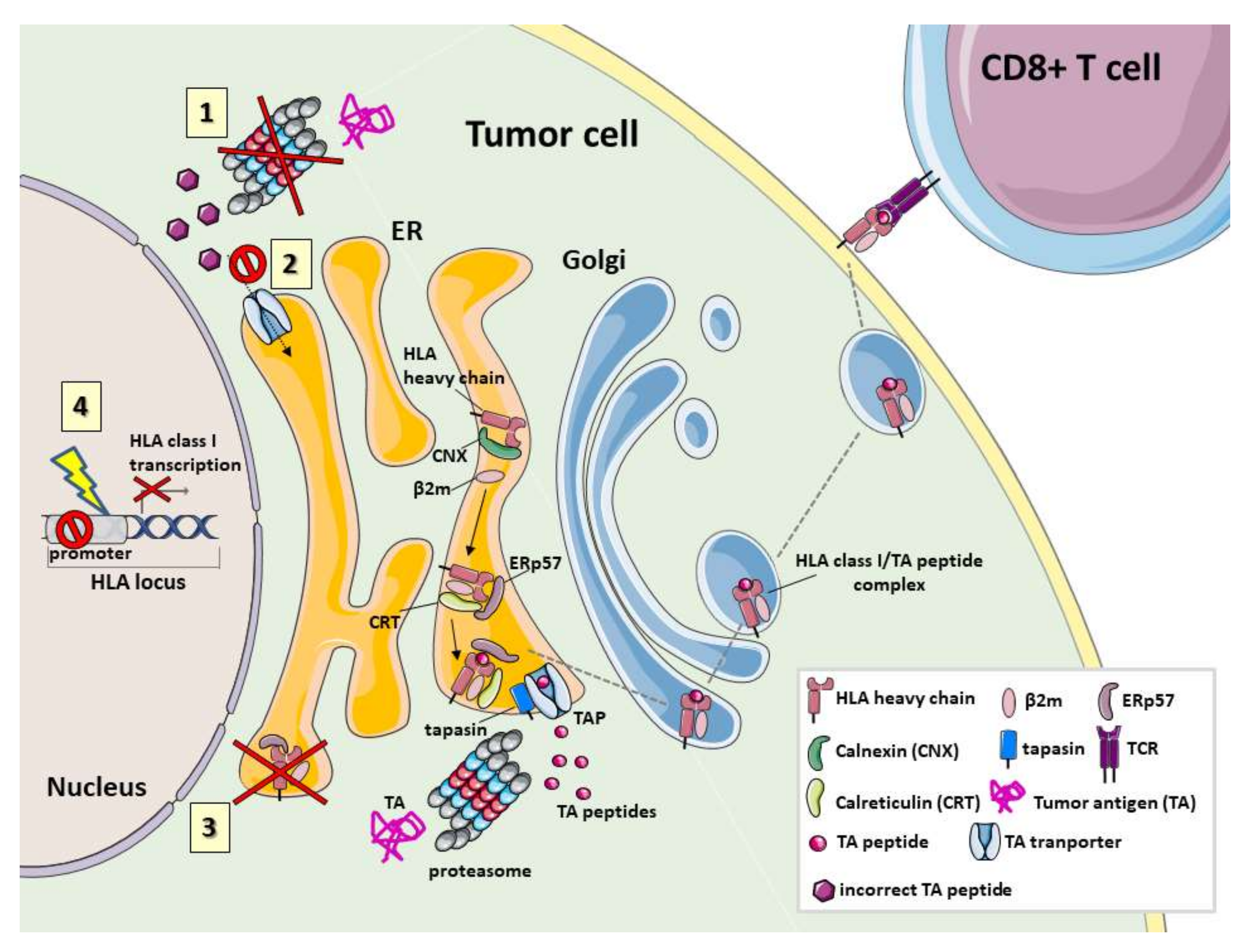
2. Defects and Clinical Significance of HLA in Human Cancer
2.1. HLA Class I Molecule Defects
2.2. Proteasome Defects
2.3. Defects in TAP1, TAP2 and Other Chaperones
2.4. HLA Class II Defects
3. Role of HLA as A Predictive Biomarker for ICI-Based Immunotherapy
3.1. Impact of HLA Class I and II on ICI-Based Immunotherapy In Vivo
3.2. HLA Class I and II as Predictive Biomarker for ICI-Based Immunotherapy: Clinical Evidences
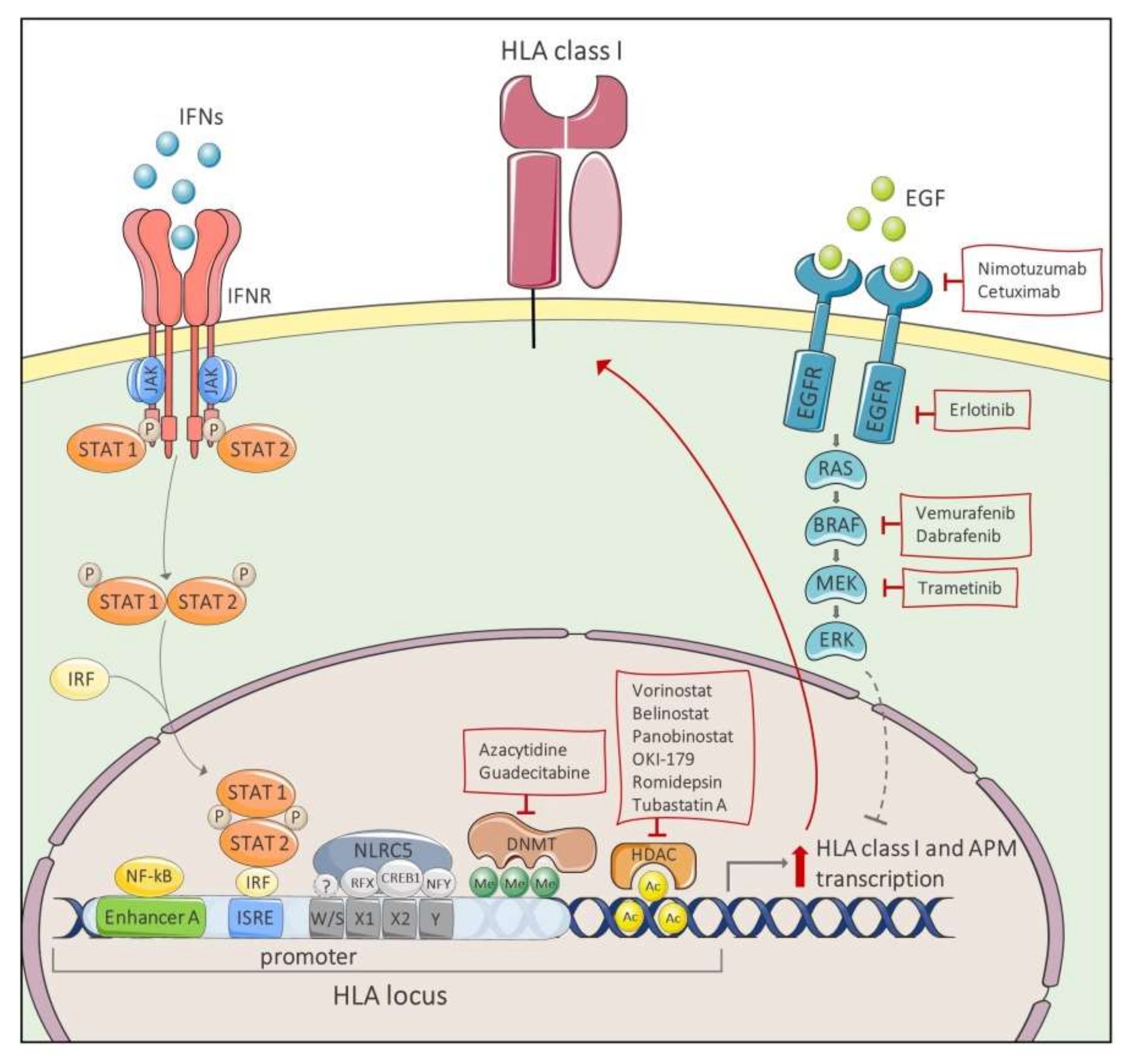
4. Restoring HLA Class I Expression as A Novel Therapeutic Strategy for Cancer Immunotherapy
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ACT | adoptive cell-transfer |
| APC | antigen presenting cell |
| APM | antigen processing machinery |
| ATF1 | activating transcriptor factor 1 |
| ATP | adenosine triphosphate |
| BiTES | bi-specific T-cell engager antibodies |
| BLS | bare lymphocyte disease |
| CAR | chimeric antigen receptor |
| CIITA | class II trans-activator |
| CIITA | class I trans activator |
| CLIP | class II-associated I chain peptide |
| CNX | calnexin |
| CREB1 | cAMP-responsive element binding protein 1 |
| CRT | calreticulin |
| CTL | cytotoxic T lymphocyte |
| CTLA-4 | cytotoxic T-lymphocyte antigen 4 |
| DC | dendritic cell |
| DFS | disease free survival |
| DNMTi | DNA methyltransferase inhibitor |
| EGFR | epithelial growth factor receptor |
| ER | endoplasmic reticulum |
| HDAC | histone deacetylase |
| HLA | human leukocyte antigen |
| ICI | immune checkpoint inhibitor |
| IFN | interferon |
| IRF | IFN Regulatory Factor |
| ISRE | IFN-sensitive response element |
| KIR | killer cell immunoglobulin-like receptor |
| JAK | janus kinase |
| LAG-3 | lymphocyte activation gene 3 |
| LMP | low molecular weight protein |
| LOH | loss of heterozygosity |
| mAb | monoclonal antibody |
| MAPK | mitogen-activated protein kinase |
| MEK | MAPK/ERK kinase |
| MHC | major histocompatibility complex |
| MIIC | MHC class II compartment |
| NF-kB | nuclear factor kB |
| NFY | nuclear transcriptor factor Y |
| NK | natural killer |
| NLRC5 | NOD-like receptor 5 |
| OS | overall survival |
| PD-1 | programmed cell death 1 |
| PD-L1 | programmed cell death ligand 1 |
| PFS | progression free survival |
| RFX | regulatory factor X |
| RFXANK | RFX-associated ankyrin-containing protein |
| RFXAP | RFX associated protein |
| TAP | transporters associated with antigen processing |
| TCR | T-cell receptor |
| TKI | tyrosine kinase inhibitor |
| TNF-α | tumor necrosis factor alpha |
| TA | tumor antigen |
| TAA | tumor associated antigen |
| TAM | Type II tumour-associated macrophages |
| TIL | tumor-infiltrating lymphocyte |
| TME | tumor microenvironment |
| Treg | regulatory T cell |
| TSA | tumor specific antigen |
| tsMHC-II | tumor specific MHC class II |
| β2-m | β2-microglobulin |
References
- Zinkernagel, R.M.; Doherty, P.C. Restriction of in vitro T cell-mediated cytotoxicity in lymphocytic choriomeningitis within a syngeneic or semiallogeneic system. Nature 1974, 248, 701–702. [Google Scholar] [CrossRef]
- Townsend, A.R.; Skehel, J.J. Influenza A specific cytotoxic T-cell clones that do not recognize viral glycoproteins. Nature 1982, 300, 655–657. [Google Scholar] [CrossRef]
- Townsend, A.R.; Rothbard, J.; Gotch, F.M.; Bahadur, G.; Wraith, D.; McMichael, A.J. The epitopes of influenza nucleoprotein recognized by cytotoxic T lymphocytes can be defined with short synthetic peptides. Cell 1986, 44, 959–968. [Google Scholar] [CrossRef]
- Bjorkman, P.J.; Saper, M.A.; Samraoui, B.; Bennett, W.S.; Strominger, J.L.; Wiley, D.C. Structure of the human class I histocompatibility antigen, HLA-A2. Nature 1987, 329, 506–512. [Google Scholar] [CrossRef]
- Rötzschke, O.; Falk, K.; Deres, K.; Schild, H.; Norda, M.; Metzger, J.; Jung, G.; Rammensee, H.G. Isolation and analysis of naturally processed viral peptides as recognized by cytotoxic T cells. Nature 1990, 348, 252–254. [Google Scholar] [CrossRef]
- Trowsdale, J. Genomic structure and function in the MHC. Trends Genet. 1993, 9, 117–122. [Google Scholar] [CrossRef]
- Flutter, B.; Gao, B. MHC class I antigen presentation--recently trimmed and well presented. Cell. Mol. Immunol. 2004, 1, 22–30. [Google Scholar]
- Norman, P.J.; Norberg, S.J.; Guethlein, L.A.; Nemat-Gorgani, N.; Royce, T.; Wroblewski, E.E.; Dunn, T.; Mann, T.; Alicata, C.; Hollenbach, J.A.; et al. Sequences of 95 human MHC haplotypes reveal extreme coding variation in genes other than highly polymorphic HLA class I and II. Genome Res. 2017, 27, 813–823. [Google Scholar] [CrossRef] [Green Version]
- Horton, R.; Wilming, L.; Rand, V.; Lovering, R.C.; Bruford, E.A.; Khodiyar, V.K.; Lush, M.J.; Povey, S.; Talbot, C.C.; Wright, M.W.; et al. Gene map of the extended human MHC. Nat. Rev. Genet. 2004, 5, 889–899. [Google Scholar] [CrossRef]
- Vandiedonck, C.; Knight, J.C. The human Major Histocompatibility Complex as a paradigm in genomics research. Brief. Funct. Genom. Proteom. 2009, 8, 379–394. [Google Scholar] [CrossRef] [Green Version]
- Trowsdale, J.; Knight, J.C. Major histocompatibility complex genomics and human disease. Annu. Rev. Genom. Hum. Genet. 2013, 14, 301–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Complete sequence and gene map of a human major histocompatibility complex. The MHC sequencing consortium. Nature 1999, 401, 921–923. [CrossRef]
- Milner, C.M.; Campbell, R.D. Genetic organization of the human MHC class III region. Front. Biosci. 2001, 6, D914–D926. [Google Scholar] [CrossRef]
- Neefjes, J.; Jongsma, M.L.M.; Paul, P.; Bakke, O. Towards a systems understanding of MHC class I and MHC class II antigen presentation. Nat. Rev. Immunol. 2011, 11, 823–836. [Google Scholar] [CrossRef]
- Ajitkumar, P.; Geier, S.S.; Kesari, K.V.; Borriello, F.; Nakagawa, M.; Bluestone, J.A.; Saper, M.A.; Wiley, D.C.; Nathenson, S.G. Evidence that multiple residues on both the alpha-helices of the class I MHC molecule are simultaneously recognized by the T cell receptor. Cell 1988, 54, 47–56. [Google Scholar] [CrossRef]
- Garcia, K.C.; Degano, M.; Stanfield, R.L.; Brunmark, A.; Jackson, M.R.; Peterson, P.A.; Teyton, L.; Wilson, I.A. An alphabeta T cell receptor structure at 2.5 A and its orientation in the TCR-MHC complex. Science 1996, 274, 209–219. [Google Scholar] [CrossRef]
- Garboczi, D.N.; Ghosh, P.; Utz, U.; Fan, Q.R.; Biddison, W.E.; Wiley, D.C. Structure of the complex between human T-cell receptor, viral peptide and HLA-A2. Nature 1996, 384, 134–141. [Google Scholar] [CrossRef]
- Blum, J.S.; Wearsch, P.A.; Cresswell, P. Pathways of antigen processing. Annu. Rev. Immunol. 2013, 31, 443–473. [Google Scholar] [CrossRef] [Green Version]
- Farhood, B.; Najafi, M.; Mortezaee, K. CD8+ cytotoxic T lymphocytes in cancer immunotherapy: A review. J. Cell. Physiol. 2019, 234, 8509–8521. [Google Scholar] [CrossRef]
- Cruz, F.M.; Colbert, J.D.; Merino, E.; Kriegsman, B.A.; Rock, K.L. The Biology and Underlying Mechanisms of Cross-Presentation of Exogenous Antigens on MHC-I Molecules. Annu. Rev. Immunol. 2017, 35, 149–176. [Google Scholar] [CrossRef] [Green Version]
- Rock, K.L.; Shen, L. Cross-presentation: Underlying mechanisms and role in immune surveillance. Immunol. Rev. 2005, 207, 166–183. [Google Scholar] [CrossRef] [PubMed]
- Muntjewerff, E.M.; Meesters, L.D.; Van den Bogaart, G. Antigen Cross-Presentation by Macrophages. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Bevan, M.J. Minor H antigens introduced on H-2 different stimulating cells cross-react at the cytotoxic T cell level during in vivo priming. J. Immunol. 1976, 117, 2233–2238. [Google Scholar]
- Embgenbroich, M.; Burgdorf, S. Current Concepts of Antigen Cross-Presentation. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Leone, P.; Shin, E.-C.; Perosa, F.; Vacca, A.; Dammacco, F.; Racanelli, V. MHC class I antigen processing and presenting machinery: Organization, function, and defects in tumor cells. J. Natl. Cancer Inst. 2013, 105, 1172–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thielens, A.; Vivier, E.; Romagné, F. NK cell MHC class I specific receptors (KIR): From biology to clinical intervention. Curr. Opin. Immunol. 2012, 24, 239–245. [Google Scholar] [CrossRef]
- Wieczorek, M.; Abualrous, E.T.; Sticht, J.; Álvaro-Benito, M.; Stolzenberg, S.; Noé, F.; Freund, C. Major Histocompatibility Complex (MHC) Class I and MHC Class II Proteins: Conformational Plasticity in Antigen Presentation. Front. Immunol. 2017, 8, 292. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, B.A. Structure and significance of beta2-microglobulin. Fed. Proc. 1976, 35, 1171–1176. [Google Scholar]
- Cunningham, B.A.; Berggård, I. Structure, evolution and significance of beta2-microglobulin. Transplant. Rev. 1974, 21, 3–14. [Google Scholar] [CrossRef]
- Madden, D.R. The three-dimensional structure of peptide-MHC complexes. Annu. Rev. Immunol. 1995, 13, 587–622. [Google Scholar] [CrossRef]
- Wilson, I.A.; Fremont, D.H. Structural analysis of MHC class I molecules with bound peptide antigens. Semin. Immunol. 1993, 5, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Persson, K.; Schneider, G. Three-dimensional structures of MHC class I-peptide complexes: Implications for peptide recognition. Arch. Immunol. Ther. Exp. 2000, 48, 135–142. [Google Scholar]
- Mage, M.G.; Dolan, M.A.; Wang, R.; Boyd, L.F.; Revilleza, M.J.; Robinson, H.; Natarajan, K.; Myers, N.B.; Hansen, T.H.; Margulies, D.H. The peptide-receptive transition state of MHC class I molecules: Insight from structure and molecular dynamics. J. Immunol. 2012, 189, 1391–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braud, V.M.; Allan, D.S.; O’Callaghan, C.A.; Söderström, K.; D’Andrea, A.; Ogg, G.S.; Lazetic, S.; Young, N.T.; Bell, J.I.; Phillips, J.H.; et al. HLA-E binds to natural killer cell receptors CD94/NKG2A, B and C. Nature 1998, 391, 795–799. [Google Scholar] [CrossRef] [PubMed]
- Carosella, E.D.; HoWangYin, K.-Y.; Favier, B.; LeMaoult, J. HLA-G-dependent suppressor cells: Diverse by nature, function, and significance. Hum. Immunol. 2008, 69, 700–707. [Google Scholar] [CrossRef]
- Lepin, E.J.; Bastin, J.M.; Allan, D.S.; Roncador, G.; Braud, V.M.; Mason, D.Y.; Van der Merwe, P.A.; McMichael, A.J.; Bell, J.I.; Powis, S.H.; et al. Functional characterization of HLA-F and binding of HLA-F tetramers to ILT2 and ILT4 receptors. Eur. J. Immunol. 2000, 30, 3552–3561. [Google Scholar] [CrossRef]
- Persson, G.; Jørgensen, N.; Nilsson, L.L.; Andersen, L.H.J.; Hviid, T.V.F. A role for both HLA-F and HLA-G in reproduction and during pregnancy? Hum. Immunol. 2020, 81, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Hackmon, R.; Pinnaduwage, L.; Zhang, J.; Lye, S.J.; Geraghty, D.E.; Dunk, C.E. Definitive class I human leukocyte antigen expression in gestational placentation: HLA-F, HLA-E, HLA-C, and HLA-G in extravillous trophoblast invasion on placentation, pregnancy, and parturition. Am. J. Reprod. Immunol. 2017, 77. [Google Scholar] [CrossRef]
- Joosten, S.A.; Sullivan, L.C.; Ottenhoff, T.H.M. Characteristics of HLA-E Restricted T-Cell Responses and Their Role in Infectious Diseases. J. Immunol. Res. 2016, 2016, 2695396. [Google Scholar] [CrossRef] [Green Version]
- Raghavan, M.; Del Cid, N.; Rizvi, S.M.; Peters, L.R. MHC class I assembly: Out and about. Trends Immunol. 2008, 29, 436–443. [Google Scholar] [CrossRef] [Green Version]
- Paulsson, K.M. Evolutionary and functional perspectives of the major histocompatibility complex class I antigen-processing machinery. Cell. Mol. Life Sci. 2004, 61, 2446–2460. [Google Scholar] [CrossRef]
- Wearsch, P.A.; Cresswell, P. The quality control of MHC class I peptide loading. Curr. Opin. Cell Biol. 2008, 20, 624–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antoniou, A.N.; Powis, S.J.; Elliott, T. Assembly and export of MHC class I peptide ligands. Curr. Opin. Immunol. 2003, 15, 75–81. [Google Scholar] [CrossRef]
- Jensen, P.E. Recent advances in antigen processing and presentation. Nat. Immunol. 2007, 8, 1041–1048. [Google Scholar] [CrossRef] [PubMed]
- Kloetzel, P.M. Antigen processing by the proteasome. Nat. Rev. Mol. Cell Biol. 2001, 2, 179–187. [Google Scholar] [CrossRef]
- Sant, A.; Yewdell, J. Antigen processing and recognition. Curr. Opin. Immunol. 2003, 15, 66–68. [Google Scholar] [CrossRef]
- Schölz, C.; Tampé, R. The peptide-loading complex--antigen translocation and MHC class I loading. Biol. Chem. 2009, 390, 783–794. [Google Scholar] [CrossRef]
- Kotsias, F.; Cebrian, I.; Alloatti, A. Antigen processing and presentation. Int. Rev. Cell Mol. Biol 2019, 348, 69–121. [Google Scholar] [CrossRef]
- Koegl, M.; Hoppe, T.; Schlenker, S.; Ulrich, H.D.; Mayer, T.U.; Jentsch, S. A novel ubiquitination factor, E4, is involved in multiubiquitin chain assembly. Cell 1999, 96, 635–644. [Google Scholar] [CrossRef] [Green Version]
- Goldberg, A.L. Functions of the proteasome: The lysis at the end of the tunnel. Science 1995, 268, 522–523. [Google Scholar] [CrossRef]
- Kloetzel, P.-M. The proteasome and MHC class I antigen processing. Biochim. Biophys. Acta 2004, 1695, 225–233. [Google Scholar] [CrossRef] [Green Version]
- Rechsteiner, M.; Hoffman, L.; Dubiel, W. The multicatalytic and 26 S proteases. J. Biol. Chem. 1993, 268, 6065–6068. [Google Scholar]
- Griffin, T.A.; Nandi, D.; Cruz, M.; Fehling, H.J.; Kaer, L.V.; Monaco, J.J.; Colbert, R.A. Immunoproteasome assembly: Cooperative incorporation of interferon gamma (IFN-gamma)-inducible subunits. J. Exp. Med. 1998, 187, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Groettrup, M.; Standera, S.; Stohwasser, R.; Kloetzel, P.M. The subunits MECL-1 and LMP2 are mutually required for incorporation into the 20S proteasome. Proc. Natl. Acad. Sci. USA 1997, 94, 8970–8975. [Google Scholar] [CrossRef] [Green Version]
- Nandi, D.; Woodward, E.; Ginsburg, D.B.; Monaco, J.J. Intermediates in the formation of mouse 20S proteasomes: Implications for the assembly of precursor beta subunits. EMBO J. 1997, 16, 5363–5375. [Google Scholar] [CrossRef] [Green Version]
- Aki, M.; Shimbara, N.; Takashina, M.; Akiyama, K.; Kagawa, S.; Tamura, T.; Tanahashi, N.; Yoshimura, T.; Tanaka, K.; Ichihara, A. Interferon-gamma induces different subunit organizations and functional diversity of proteasomes. J. Biochem. 1994, 115, 257–269. [Google Scholar] [CrossRef]
- Shin, E.-C.; Seifert, U.; Kato, T.; Rice, C.M.; Feinstone, S.M.; Kloetzel, P.-M.; Rehermann, B. Virus-induced type I IFN stimulates generation of immunoproteasomes at the site of infection. J. Clin. Investig. 2006, 116, 3006–3014. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Norbury, C.C.; Cho, Y.; Yewdell, J.W.; Bennink, J.R. Immunoproteasomes shape immunodominance hierarchies of antiviral CD8(+) T cells at the levels of T cell repertoire and presentation of viral antigens. J. Exp. Med. 2001, 193, 1319–1326. [Google Scholar] [CrossRef] [Green Version]
- Melnikova, V.I.; Sharova, N.P.; Maslova, E.V.; Voronova, S.N.; Zakharova, L.A. Ontogenesis of rat immune system: Proteasome expression in different cell populations of the developing thymus. Cell. Immunol. 2010, 266, 83–89. [Google Scholar] [CrossRef]
- Schmitt, L.; Tampé, R. Structure and mechanism of ABC transporters. Curr. Opin. Struct. Biol. 2002, 12, 754–760. [Google Scholar] [CrossRef]
- Arora, S.; Lapinski, P.E.; Raghavan, M. Use of chimeric proteins to investigate the role of transporter associated with antigen processing (TAP) structural domains in peptide binding and translocation. Proc. Natl. Acad. Sci. USA 2001, 98, 7241–7246. [Google Scholar] [CrossRef] [Green Version]
- Higgins, C.F.; Linton, K.J. The ATP switch model for ABC transporters. Nat. Struct. Mol. Biol. 2004, 11, 918–926. [Google Scholar] [CrossRef] [PubMed]
- Parcej, D.; Tampé, R. ABC proteins in antigen translocation and viral inhibition. Nat. Chem. Biol. 2010, 6, 572–580. [Google Scholar] [CrossRef]
- Herget, M.; Baldauf, C.; Schölz, C.; Parcej, D.; Wiesmüller, K.-H.; Tampé, R.; Abele, R.; Bordignon, E. Conformation of peptides bound to the transporter associated with antigen processing (TAP). Proc. Natl. Acad. Sci. USA 2011, 108, 1349–1354. [Google Scholar] [CrossRef] [Green Version]
- Koopmann, J.O.; Post, M.; Neefjes, J.J.; Hämmerling, G.J.; Momburg, F. Translocation of long peptides by transporters associated with antigen processing (TAP). Eur. J. Immunol. 1996, 26, 1720–1728. [Google Scholar] [CrossRef]
- Momburg, F.; Roelse, J.; Howard, J.C.; Butcher, G.W.; Hämmerling, G.J.; Neefjes, J.J. Selectivity of MHC-encoded peptide transporters from human, mouse and rat. Nature 1994, 367, 648–651. [Google Scholar] [CrossRef]
- Momburg, F.; Roelse, J.; Hämmerling, G.J.; Neefjes, J.J. Peptide size selection by the major histocompatibility complex-encoded peptide transporter. J. Exp. Med. 1994, 179, 1613–1623. [Google Scholar] [CrossRef]
- Momburg, F.; Hämmerling, G.J. Generation and TAP-mediated transport of peptides for major histocompatibility complex class I molecules. Adv. Immunol. 1998, 68, 191–256. [Google Scholar] [CrossRef]
- Schumacher, T.N.; Kantesaria, D.V.; Heemels, M.T.; Ashton-Rickardt, P.G.; Shepherd, J.C.; Fruh, K.; Yang, Y.; Peterson, P.A.; Tonegawa, S.; Ploegh, H.L. Peptide length and sequence specificity of the mouse TAP1/TAP2 translocator. J. Exp. Med. 1994, 179, 533–540. [Google Scholar] [CrossRef]
- Roelse, J.; Grommé, M.; Momburg, F.; Hämmerling, G.; Neefjes, J. Trimming of TAP-translocated peptides in the endoplasmic reticulum and in the cytosol during recycling. J. Exp. Med. 1994, 180, 1591–1597. [Google Scholar] [CrossRef] [Green Version]
- Diedrich, G.; Bangia, N.; Pan, M.; Cresswell, P. A role for calnexin in the assembly of the MHC class I loading complex in the endoplasmic reticulum. J. Immunol. 2001, 166, 1703–1709. [Google Scholar] [CrossRef] [Green Version]
- Hughes, E.A.; Cresswell, P. The thiol oxidoreductase ERp57 is a component of the MHC class I peptide-loading complex. Curr. Biol. 1998, 8, 709–712. [Google Scholar] [CrossRef] [Green Version]
- Morrice, N.A.; Powis, S.J. A role for the thiol-dependent reductase ERp57 in the assembly of MHC class I molecules. Curr. Biol. 1998, 8, 713–716. [Google Scholar] [CrossRef] [Green Version]
- Sadasivan, B.; Lehner, P.J.; Ortmann, B.; Spies, T.; Cresswell, P. Roles for calreticulin and a novel glycoprotein, tapasin, in the interaction of MHC class I molecules with TAP. Immunity 1996, 5, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Momburg, F.; Tan, P. Tapasin-the keystone of the loading complex optimizing peptide binding by MHC class I molecules in the endoplasmic reticulum. Mol. Immunol. 2002, 39, 217–233. [Google Scholar] [CrossRef]
- Ortmann, B.; Copeman, J.; Lehner, P.J.; Sadasivan, B.; Herberg, J.A.; Grandea, A.G.; Riddell, S.R.; Tampé, R.; Spies, T.; Trowsdale, J.; et al. A critical role for tapasin in the assembly and function of multimeric MHC class I-TAP complexes. Science 1997, 277, 1306–1309. [Google Scholar] [CrossRef] [Green Version]
- Dick, T.P.; Bangia, N.; Peaper, D.R.; Cresswell, P. Disulfide bond isomerization and the assembly of MHC class I-peptide complexes. Immunity 2002, 16, 87–98. [Google Scholar] [CrossRef] [Green Version]
- Ortmann, B.; Androlewicz, M.J.; Cresswell, P. MHC class I/beta 2-microglobulin complexes associate with TAP transporters before peptide binding. Nature 1994, 368, 864–867. [Google Scholar] [CrossRef]
- Wright, C.A.; Kozik, P.; Zacharias, M.; Springer, S. Tapasin and other chaperones: Models of the MHC class I loading complex. Biol. Chem. 2004, 385, 763–778. [Google Scholar] [CrossRef]
- Ellgaard, L.; Helenius, A. ER quality control: Towards an understanding at the molecular level. Curr. Opin. Cell Biol. 2001, 13, 431–437. [Google Scholar] [CrossRef]
- Oliver, J.D.; Roderick, H.L.; Llewellyn, D.H.; High, S. ERp57 functions as a subunit of specific complexes formed with the ER lectins calreticulin and calnexin. Mol. Biol. Cell 1999, 10, 2573–2582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degen, E.; Cohen-Doyle, M.F.; Williams, D.B. Efficient dissociation of the p88 chaperone from major histocompatibility complex class I molecules requires both beta 2-microglobulin and peptide. J. Exp. Med. 1992, 175, 1653–1661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nössner, E.; Parham, P. Species-specific differences in chaperone interaction of human and mouse major histocompatibility complex class I molecules. J. Exp. Med. 1995, 181, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Groothuis, T.A.M.; Neefjes, J. The many roads to cross-presentation. J. Exp. Med. 2005, 202, 1313–1318. [Google Scholar] [CrossRef] [Green Version]
- Neefjes, J.; Sadaka, C. Into the Intracellular Logistics of Cross-Presentation. Front. Immunol. 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- Lechler, R.I. Structure-function relationships of MHC class II molecules. Immunol. Suppl. 1988, 1, 25–26. [Google Scholar]
- Stern, L.J.; Potolicchio, I.; Santambrogio, L. MHC class II compartment subtypes: Structure and function. Curr. Opin. Immunol. 2006, 18, 64–69. [Google Scholar] [CrossRef]
- Rock, K.L.; Reits, E.; Neefjes, J. Present Yourself! By MHC Class I and MHC Class II Molecules. Trends Immunol. 2016, 37, 724–737. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.H.; Flavell, R.A. Class II transactivator regulates the expression of multiple genes involved in antigen presentation. J. Exp. Med. 1995, 181, 765–767. [Google Scholar] [CrossRef] [Green Version]
- Collins, T.; Korman, A.J.; Wake, C.T.; Boss, J.M.; Kappes, D.J.; Fiers, W.; Ault, K.A.; Gimbrone, M.A.; Strominger, J.L.; Pober, J.S. Immune interferon activates multiple class II major histocompatibility complex genes and the associated invariant chain gene in human endothelial cells and dermal fibroblasts. Proc. Natl. Acad. Sci. USA 1984, 81, 4917–4921. [Google Scholar] [CrossRef] [Green Version]
- Holling, T.M.; Schooten, E.; Van Den Elsen, P.J. Function and regulation of MHC class II molecules in T-lymphocytes: Of mice and men. Hum. Immunol. 2004, 65, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Reith, W.; LeibundGut-Landmann, S.; Waldburger, J.-M. Regulation of MHC class II gene expression by the class II transactivator. Nat. Rev. Immunol. 2005, 5, 793–806. [Google Scholar] [CrossRef] [PubMed]
- Ting, J.P.-Y.; Trowsdale, J. Genetic control of MHC class II expression. Cell 2002, 109 (Suppl. S21–S33). [Google Scholar] [CrossRef] [Green Version]
- Boss, J.M.; Jensen, P.E. Transcriptional regulation of the MHC class II antigen presentation pathway. Curr. Opin. Immunol. 2003, 15, 105–111. [Google Scholar] [CrossRef]
- Neumann, J.; Koch, N. Assembly of major histocompatibility complex class II subunits with invariant chain. FEBS Lett. 2005, 579, 6055–6059. [Google Scholar] [CrossRef] [Green Version]
- Harton, J.; Jin, L.; Hahn, A.; Drake, J. Immunological Functions of the Membrane Proximal Region of MHC Class II Molecules. F1000Res 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Arnold, P.Y.; La Gruta, N.L.; Miller, T.; Vignali, K.M.; Adams, P.S.; Woodland, D.L.; Vignali, D.A.A. The majority of immunogenic epitopes generate CD4+ T cells that are dependent on MHC class II-bound peptide-flanking residues. J. Immunol. 2002, 169, 739–749. [Google Scholar] [CrossRef] [Green Version]
- Axelrod, M.L.; Cook, R.S.; Johnson, D.B.; Balko, J.M. Biological Consequences of MHC-II Expression by Tumor Cells in Cancer. Clin. Cancer Res. 2019, 25, 2392–2402. [Google Scholar] [CrossRef]
- Pieters, J. MHC class II restricted antigen presentation. Curr. Opin. Immunol. 1997, 9, 89–96. [Google Scholar] [CrossRef]
- Seliger, B.; Maeurer, M.J.; Ferrone, S. Antigen-processing machinery breakdown and tumor growth. Immunol. Today 2000, 21, 455–464. [Google Scholar] [CrossRef]
- Cresswell, P. Invariant chain structure and MHC class II function. Cell 1996, 84, 505–507. [Google Scholar] [CrossRef] [Green Version]
- Busch, R.; Cloutier, I.; Sékaly, R.P.; Hämmerling, G.J. Invariant chain protects class II histocompatibility antigens from binding intact polypeptides in the endoplasmic reticulum. EMBO J. 1996, 15, 418–428. [Google Scholar] [CrossRef]
- Denzin, L.K.; Cresswell, P. HLA-DM induces CLIP dissociation from MHC class II alpha beta dimers and facilitates peptide loading. Cell 1995, 82, 155–165. [Google Scholar] [CrossRef] [Green Version]
- Kropshofer, H.; Arndt, S.O.; Moldenhauer, G.; Hämmerling, G.J.; Vogt, A.B. HLA-DM acts as a molecular chaperone and rescues empty HLA-DR molecules at lysosomal pH. Immunity 1997, 6, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Van Den Elsen, P.J. Expression Regulation of Major Histocompatibility Complex Class I and Class II Encoding Genes. Front. Immunol. 2011, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, K.S.; Van den Elsen, P.J. NLRC5: A key regulator of MHC class I-dependent immune responses. Nat. Rev. Immunol. 2012, 12, 813–820. [Google Scholar] [CrossRef]
- Jongsma, M.L.M.; Guarda, G.; Spaapen, R.M. The regulatory network behind MHC class I expression. Mol. Immunol. 2019, 113, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Steimle, V.; Otten, L.A.; Zufferey, M.; Mach, B. Complementation cloning of an MHC class II transactivator mutated in hereditary MHC class II deficiency (or bare lymphocyte syndrome). Cell 1993, 75, 135–146. [Google Scholar] [CrossRef]
- Meissner, T.B.; Liu, Y.-J.; Lee, K.-H.; Li, A.; Biswas, A.; Van Eggermond, M.C.J.A.; Van den Elsen, P.J.; Kobayashi, K.S. NLRC5 cooperates with the RFX transcription factor complex to induce MHC class I gene expression. J. Immunol. 2012, 188, 4951–4958. [Google Scholar] [CrossRef] [Green Version]
- Drozina, G.; Kohoutek, J.; Jabrane-Ferrat, N.; Peterlin, B.M. Expression of MHC II genes. Curr. Top. Microbiol. Immunol. 2005, 290, 147–170. [Google Scholar] [CrossRef]
- Gobin, S.J.; Van Zutphen, M.; Westerheide, S.D.; Boss, J.M.; Van den Elsen, P.J. The MHC-specific enhanceosome and its role in MHC class I and beta(2)-microglobulin gene transactivation. J. Immunol. 2001, 167, 5175–5184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harton, J.A.; Ting, J.P. Class II transactivator: Mastering the art of major histocompatibility complex expression. Mol. Cell. Biol. 2000, 20, 6185–6194. [Google Scholar] [CrossRef] [PubMed]
- Reith, W.; Mach, B. The bare lymphocyte syndrome and the regulation of MHC expression. Annu. Rev. Immunol. 2001, 19, 331–373. [Google Scholar] [CrossRef] [PubMed]
- Krawczyk, M.; Reith, W. Regulation of MHC class II expression, a unique regulatory system identified by the study of a primary immunodeficiency disease. Tissue Antigens 2006, 67, 183–197. [Google Scholar] [CrossRef]
- Robbins, G.R.; Truax, A.D.; Davis, B.K.; Zhang, L.; Brickey, W.J.; Ting, J.P.-Y. Regulation of Class I Major Histocompatibility Complex (MHC) by Nucleotide-binding Domain, Leucine-rich Repeat-containing (NLR) Proteins. J. Biol. Chem. 2012, 287, 24294–24303. [Google Scholar] [CrossRef] [Green Version]
- Biswas, A.; Meissner, T.B.; Kawai, T.; Kobayashi, K.S. Cutting edge: Impaired MHC class I expression in mice deficient for Nlrc5/class I transactivator. J. Immunol. 2012, 189, 516–520. [Google Scholar] [CrossRef] [Green Version]
- Ludigs, K.; Jandus, C.; Utzschneider, D.T.; Staehli, F.; Bessoles, S.; Dang, A.T.; Rota, G.; Castro, W.; Zehn, D.; Vivier, E.; et al. NLRC5 shields T lymphocytes from NK-cell-mediated elimination under inflammatory conditions. Nat. Commun. 2016, 7, 10554. [Google Scholar] [CrossRef]
- Martin, B.K.; Chin, K.C.; Olsen, J.C.; Skinner, C.A.; Dey, A.; Ozato, K.; Ting, J.P. Induction of MHC class I expression by the MHC class II transactivator CIITA. Immunity 1997, 6, 591–600. [Google Scholar] [CrossRef] [Green Version]
- Meissner, T.B.; Li, A.; Kobayashi, K.S. NLRC5: A newly discovered MHC class I transactivator (CITA). Microbes Infect. 2012, 14, 477–484. [Google Scholar] [CrossRef] [Green Version]
- Gruen, J.R.; Weissman, S.M. Human MHC class III and IV genes and disease associations. Front. Biosci. 2001, 6, D960–D972. [Google Scholar] [CrossRef]
- Deakin, J.E.; Papenfuss, A.T.; Belov, K.; Cross, J.G.R.; Coggill, P.; Palmer, S.; Sims, S.; Speed, T.P.; Beck, S.; Graves, J.A.M. Evolution and comparative analysis of the MHC Class III inflammatory region. BMC Genom. 2006, 7, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, T.; Rowen, L.; Aguado, B.; Ahearn, M.E.; Madan, A.; Qin, S.; Campbell, R.D.; Hood, L. Analysis of the gene-dense major histocompatibility complex class III region and its comparison to mouse. Genome Res. 2003, 13, 2621–2636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, D.B.; Estrada, M.V.; Salgado, R.; Sanchez, V.; Doxie, D.B.; Opalenik, S.R.; Vilgelm, A.E.; Feld, E.; Johnson, A.S.; Greenplate, A.R.; et al. Melanoma-specific MHC-II expression represents a tumour-autonomous phenotype and predicts response to anti-PD-1/PD-L1 therapy. Nat. Commun. 2016, 7, 10582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, I.A.; Hwang, S.-H.; Song, I.H.; Heo, S.-H.; Kim, Y.-A.; Bang, W.S.; Park, H.S.; Lee, M.; Gong, G.; Lee, H.J. Expression of the MHC class II in triple-negative breast cancer is associated with tumor-infiltrating lymphocytes and interferon signaling. PLoS ONE 2017, 12, e0182786. [Google Scholar] [CrossRef] [PubMed]
- Roemer, M.G.M.; Redd, R.A.; Cader, F.Z.; Pak, C.J.; Abdelrahman, S.; Ouyang, J.; Sasse, S.; Younes, A.; Fanale, M.; Santoro, A.; et al. Major Histocompatibility Complex Class II and Programmed Death Ligand 1 Expression Predict Outcome After Programmed Death 1 Blockade in Classic Hodgkin Lymphoma. J. Clin. Oncol. 2018, 36, 942–950. [Google Scholar] [CrossRef]
- Rodig, S.J.; Gusenleitner, D.; Jackson, D.G.; Gjini, E.; Giobbie-Hurder, A.; Jin, C.; Chang, H.; Lovitch, S.B.; Horak, C.; Weber, J.S.; et al. MHC proteins confer differential sensitivity to CTLA-4 and PD-1 blockade in untreated metastatic melanoma. Sci. Transl. Med. 2018, 10. [Google Scholar] [CrossRef] [Green Version]
- Seliger, B.; Kloor, M.; Ferrone, S. HLA class II antigen-processing pathway in tumors: Molecular defects and clinical relevance. Oncoimmunology 2017, 6. [Google Scholar] [CrossRef]
- Oldford, S.A.; Robb, J.D.; Watson, P.H.; Drover, S. HLA-DRB alleles are differentially expressed by tumor cells in breast carcinoma. Int. J. Cancer 2004, 112, 399–406. [Google Scholar] [CrossRef]
- Bustin, S.A.; Li, S.R.; Phillips, S.; Dorudi, S. Expression of HLA class II in colorectal cancer: Evidence for enhanced immunogenicity of microsatellite-instability-positive tumours. Tumour Biol. 2001, 22, 294–298. [Google Scholar] [CrossRef]
- Younger, A.R.; Amria, S.; Jeffrey, W.A.; Mahdy, A.E.M.; Goldstein, O.G.; Norris, J.S.; Haque, A. HLA class II antigen presentation by prostate cancer cells. Prostate Cancer Prostatic Dis. 2008, 11, 334–341. [Google Scholar] [CrossRef]
- Forero, A.; Li, Y.; Chen, D.; Grizzle, W.E.; Updike, K.L.; Merz, N.D.; Downs-Kelly, E.; Burwell, T.C.; Vaklavas, C.; Buchsbaum, D.J.; et al. Expression of the MHC Class II Pathway in Triple-Negative Breast Cancer Tumor Cells Is Associated with a Good Prognosis and Infiltrating Lymphocytes. Cancer Immunol. Res. 2016, 4, 390–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callahan, M.J.; Nagymanyoki, Z.; Bonome, T.; Johnson, M.E.; Litkouhi, B.; Sullivan, E.H.; Hirsch, M.S.; Matulonis, U.A.; Liu, J.; Birrer, M.J.; et al. Increased HLA-DMB expression in the tumor epithelium is associated with increased CTL infiltration and improved prognosis in advanced-stage serous ovarian cancer. Clin. Cancer Res. 2008, 14, 7667–7673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dierssen, J.W.F.; De Miranda, N.F.C.C.; Ferrone, S.; Van Puijenbroek, M.; Cornelisse, C.J.; Fleuren, G.J.; Van Wezel, T.; Morreau, H. HNPCC versus sporadic microsatellite-unstable colon cancers follow different routes toward loss of HLA class I expression. BMC Cancer 2007, 7, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Miranda, N.F.C.C.; Nielsen, M.; Pereira, D.; Van Puijenbroek, M.; Vasen, H.F.; Hes, F.J.; Van Wezel, T.; Morreau, H. MUTYH-associated polyposis carcinomas frequently lose HLA class I expression—A common event amongst DNA-repair-deficient colorectal cancers. J. Pathol. 2009, 219, 69–76. [Google Scholar] [CrossRef]
- Bicknell, D.C.; Kaklamanis, L.; Hampson, R.; Bodmer, W.F.; Karran, P. Selection for beta 2-microglobulin mutation in mismatch repair-defective colorectal carcinomas. Curr. Biol. 1996, 6, 1695–1697. [Google Scholar] [CrossRef] [Green Version]
- Cabrera, T.; Collado, A.; Fernandez, M.A.; Ferron, A.; Sancho, J.; Ruiz-Cabello, F.; Garrido, F. High frequency of altered HLA class I phenotypes in invasive colorectal carcinomas. Tissue Antigens 1998, 52, 114–123. [Google Scholar] [CrossRef]
- Cabrera, T.; Maleno, I.; Collado, A.; Lopez Nevot, M.A.; Tait, B.D.; Garrido, F. Analysis of HLA class I alterations in tumors: Choosing a strategy based on known patterns of underlying molecular mechanisms. Tissue Antigens 2007, 69 (Suppl. S1), 264–268. [Google Scholar] [CrossRef]
- Benitez, R.; Godelaine, D.; Lopez-Nevot, M.A.; Brasseur, F.; Jiménez, P.; Marchand, M.; Oliva, M.R.; Van Baren, N.; Cabrera, T.; Andry, G.; et al. Mutations of the beta2-microglobulin gene result in a lack of HLA class I molecules on melanoma cells of two patients immunized with MAGE peptides. Tissue Antigens 1998, 52, 520–529. [Google Scholar] [CrossRef]
- Hicklin, D.J.; Wang, Z.; Arienti, F.; Rivoltini, L.; Parmiani, G.; Ferrone, S. beta2-Microglobulin mutations, HLA class I antigen loss, and tumor progression in melanoma. J. Clin. Investig. 1998, 101, 2720–2729. [Google Scholar] [CrossRef]
- Tang, Q.; Zhang, J.; Qi, B.; Shen, C.; Xie, W. Downregulation of HLA class I molecules in primary oral squamous cell carcinomas and cell lines. Arch. Med. Res. 2009, 40, 256–263. [Google Scholar] [CrossRef]
- Maleno, I.; López-Nevot, M.A.; Cabrera, T.; Salinero, J.; Garrido, F. Multiple mechanisms generate HLA class I altered phenotypes in laryngeal carcinomas: High frequency of HLA haplotype loss associated with loss of heterozygosity in chromosome region 6p21. Cancer Immunol. Immunother. 2002, 51, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, T.; Angustias Fernandez, M.; Sierra, A.; Garrido, A.; Herruzo, A.; Escobedo, A.; Fabra, A.; Garrido, F. High frequency of altered HLA class I phenotypes in invasive breast carcinomas. Hum. Immunol. 1996, 50, 127–134. [Google Scholar] [CrossRef]
- Qifeng, S.; Bo, C.; Xingtao, J.; Chuanliang, P.; Xiaogang, Z. Methylation of the promoter of human leukocyte antigen class I in human esophageal squamous cell carcinoma and its histopathological characteristics. J. Thorac. Cardiovasc. Surg. 2011, 141, 808–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Facoetti, A.; Nano, R.; Zelini, P.; Morbini, P.; Benericetti, E.; Ceroni, M.; Campoli, M.; Ferrone, S. Human leukocyte antigen and antigen processing machinery component defects in astrocytic tumors. Clin. Cancer Res. 2005, 11, 8304–8311. [Google Scholar] [CrossRef] [Green Version]
- Cai, L.; Michelakos, T.; Yamada, T.; Fan, S.; Wang, X.; Schwab, J.H.; Ferrone, C.R.; Ferrone, S. Defective HLA class I antigen processing machinery in cancer. Cancer Immunol. Immunother. 2018, 67, 999–1009. [Google Scholar] [CrossRef] [PubMed]
- Garrido, F.; Algarra, I. MHC antigens and tumor escape from immune surveillance. Adv. Cancer Res. 2001, 83, 117–158. [Google Scholar] [CrossRef]
- Kageshita, T.; Hirai, S.; Ono, T.; Hicklin, D.J.; Ferrone, S. Down-regulation of HLA class I antigen-processing molecules in malignant melanoma: Association with disease progression. Am. J. Pathol. 1999, 154, 745–754. [Google Scholar] [CrossRef]
- Campoli, M.; Ferrone, S. HLA antigen changes in malignant cells: Epigenetic mechanisms and biologic significance. Oncogene 2008, 27, 5869–5885. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.-C.; Pirozzi, G.; Wen, S.-H.; Chung, I.-H.; Chiu, B.-L.; Errico, S.; Luongo, M.; Lombardi, M.L.; Ferrone, S. Multiple structural and epigenetic defects in the human leukocyte antigen class I antigen presentation pathway in a recurrent metastatic melanoma following immunotherapy. J. Biol. Chem. 2015, 290, 26562–26575. [Google Scholar] [CrossRef] [Green Version]
- Garrido, F.; Cabrera, T.; Concha, A.; Glew, S.; Ruiz-Cabello, F.; Stern, P.L. Natural history of HLA expression during tumour development. Immunol. Today 1993, 14, 491–499. [Google Scholar] [CrossRef]
- Ljunggren, H.G.; Kärre, K. In search of the “missing self”: MHC molecules and NK cell recognition. Immunol. Today 1990, 11, 237–244. [Google Scholar] [CrossRef]
- Garrido, F.; Cabrera, T.; Aptsiauri, N. “Hard” and “soft” lesions underlying the HLA class I alterations in cancer cells: Implications for immunotherapy. Int. J. Cancer 2010, 127, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Paschen, A.; Arens, N.; Sucker, A.; Greulich-Bode, K.M.; Fonsatti, E.; Gloghini, A.; Striegel, S.; Schwinn, N.; Carbone, A.; Hildenbrand, R.; et al. The coincidence of chromosome 15 aberrations and beta2-microglobulin gene mutations is causative for the total loss of human leukocyte antigen class I expression in melanoma. Clin. Cancer Res. 2006, 12, 3297–3305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Campo, A.B.; Aptsiauri, N.; Méndez, R.; Zinchenko, S.; Vales, A.; Paschen, A.; Ward, S.; Ruiz-Cabello, F.; González-Aseguinolaza, G.; Garrido, F. Efficient recovery of HLA class I expression in human tumor cells after beta2-microglobulin gene transfer using adenoviral vector: Implications for cancer immunotherapy. Scand. J. Immunol. 2009, 70, 125–135. [Google Scholar] [CrossRef]
- Maleno, I.; Aptsiauri, N.; Cabrera, T.; Gallego, A.; Paschen, A.; López-Nevot, M.A.; Garrido, F. Frequent loss of heterozygosity in the β2-microglobulin region of chromosome 15 in primary human tumors. Immunogenetics 2011, 63, 65–71. [Google Scholar] [CrossRef]
- Pérez, B.; Benitez, R.; Fernández, M.A.; Oliva, M.R.; Soto, J.L.; Serrano, S.; López Nevot, M.A.; Garrido, F. A new beta 2 microglobulin mutation found in a melanoma tumor cell line. Tissue Antigens 1999, 53, 569–572. [Google Scholar] [CrossRef]
- Chang, C.-C.; Campoli, M.; Restifo, N.P.; Wang, X.; Ferrone, S. Immune selection of hot-spot beta 2-microglobulin gene mutations, HLA-A2 allospecificity loss, and antigen-processing machinery component down-regulation in melanoma cells derived from recurrent metastases following immunotherapy. J. Immunol. 2005, 174, 1462–1471. [Google Scholar] [CrossRef] [Green Version]
- Bernal, M.; Ruiz-Cabello, F.; Concha, A.; Paschen, A.; Garrido, F. Implication of the β2-microglobulin gene in the generation of tumor escape phenotypes. Cancer Immunol. Immunother. 2012, 61, 1359–1371. [Google Scholar] [CrossRef]
- Garrido, F.; Romero, I.; Aptsiauri, N.; Garcia-Lora, A.M. Generation of MHC class I diversity in primary tumors and selection of the malignant phenotype. Int. J. Cancer 2016, 138, 271–280. [Google Scholar] [CrossRef]
- Browning, M.; Petronzelli, F.; Bicknell, D.; Krausa, P.; Rowan, A.; Tonks, S.; Murray, N.; Bodmer, J.; Bodmer, W. Mechanisms of loss of HLA class I expression on colorectal tumor cells. Tissue Antigens 1996, 47, 364–371. [Google Scholar] [CrossRef]
- Sette, A.; Chesnut, R.; Fikes, J. HLA expression in cancer: Implications for T cell-based immunotherapy. Immunogenetics 2001, 53, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Khleif, S. (Ed.) Cancer Treatment and Research. In Tumor Immunology and Cancer Vaccines; Springer: New York, NY, USA, 2005; ISBN 978-1-4020-8119-4. [Google Scholar]
- Murphy, C.; Nikodem, D.; Howcroft, K.; Weissman, J.D.; Singer, D.S. Active repression of major histocompatibility complex class I genes in a human neuroblastoma cell line. J. Biol. Chem. 1996, 271, 30992–30999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, Y.; Yang, G.; Song, Y.; Zhao, X.; So, C.; Liao, J.; Wang, L.D.; Yang, C.S. DNA hypermethylation is a mechanism for loss of expression of the HLA class I genes in human esophageal squamous cell carcinomas. Carcinogenesis 2001, 22, 1615–1623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coral, S.; Sigalotti, L.; Gasparollo, A.; Cattarossi, I.; Visintin, A.; Cattelan, A.; Altomonte, M.; Maio, M. Prolonged upregulation of the expression of HLA class I antigens and costimulatory molecules on melanoma cells treated with 5-aza-2’-deoxycytidine (5-AZA-CdR). J. Immunother. 1999, 22, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Coral, S.; Sigalotti, L.; Altomonte, M.; Engelsberg, A.; Colizzi, F.; Cattarossi, I.; Maraskovsky, E.; Jager, E.; Seliger, B.; Maio, M. 5-aza-2’-deoxycytidine-induced expression of functional cancer testis antigens in human renal cell carcinoma: Immunotherapeutic implications. Clin. Cancer Res. 2002, 8, 2690–2695. [Google Scholar] [PubMed]
- Esteller, M. Epigenetics provides a new generation of oncogenes and tumour-suppressor genes. Br. J. Cancer 2006, 94, 179–183. [Google Scholar] [CrossRef] [Green Version]
- Lettini, A.A.; Guidoboni, M.; Fonsatti, E.; Anzalone, L.; Cortini, E.; Maio, M. Epigenetic remodelling of DNA in cancer. Histol. Histopathol. 2007, 22, 1413–1424. [Google Scholar] [CrossRef]
- Ferris, R.L.; Hunt, J.L.; Ferrone, S. Human leukocyte antigen (HLA) class I defects in head and neck cancer: Molecular mechanisms and clinical significance. Immunol. Res. 2005, 33, 113–133. [Google Scholar] [CrossRef]
- Ferrone, S.; Marincola, F.M. Loss of HLA class I antigens by melanoma cells: Molecular mechanisms, functional significance and clinical relevance. Immunol. Today 1995, 16, 487–494. [Google Scholar] [CrossRef]
- Concha-Benavente, F.; Srivastava, R.; Ferrone, S.; Ferris, R.L. Immunological and clinical significance of HLA class I antigen processing machinery component defects in malignant cells. Oral Oncol. 2016, 58, 52–58. [Google Scholar] [CrossRef] [Green Version]
- Seliger, B.; Ferrone, S. HLA Class I Antigen Processing Machinery Defects in Cancer Cells-Frequency, Functional Significance, and Clinical Relevance with Special Emphasis on Their Role in T Cell-Based Immunotherapy of Malignant Disease. Methods Mol. Biol. 2020, 2055, 325–350. [Google Scholar] [CrossRef]
- Yoshihama, S.; Roszik, J.; Downs, I.; Meissner, T.B.; Vijayan, S.; Chapuy, B.; Sidiq, T.; Shipp, M.A.; Lizee, G.A.; Kobayashi, K.S. NLRC5/MHC class I transactivator is a target for immune evasion in cancer. Proc. Natl. Acad. Sci. USA 2016, 113, 5999–6004. [Google Scholar] [CrossRef] [Green Version]
- Yoshihama, S.; Vijayan, S.; Sidiq, T.; Kobayashi, K.S. NLRC5/CITA: A key player in cancer immune surveillance. Trends Cancer 2017, 3, 28–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kloor, M.; Becker, C.; Benner, A.; Woerner, S.M.; Gebert, J.; Ferrone, S.; Von Knebel Doeberitz, M. Immunoselective pressure and human leukocyte antigen class I antigen machinery defects in microsatellite unstable colorectal cancers. Cancer Res. 2005, 65, 6418–6424. [Google Scholar] [CrossRef] [Green Version]
- Cathro, H.P.; Smolkin, M.E.; Theodorescu, D.; Jo, V.Y.; Ferrone, S.; Frierson, H.F. Relationship between HLA class I antigen processing machinery component expression and the clinicopathologic characteristics of bladder carcinomas. Cancer Immunol. Immunother. 2010, 59, 465–472. [Google Scholar] [CrossRef] [Green Version]
- Leffers, N.; Gooden, M.J.M.; Mokhova, A.A.; Kast, W.M.; Boezen, H.M.; Ten Hoor, K.A.; Hollema, H.; Daemen, T.; Van der Zee, A.G.J.; Nijman, H.W. Down-regulation of proteasomal subunit MB1 is an independent predictor of improved survival in ovarian cancer. Gynecol. Oncol. 2009, 113, 256–263. [Google Scholar] [CrossRef]
- Hoves, S.; Aigner, M.; Pfeiffer, C.; Laumer, M.; Obermann, E.C.; Mackensen, A. In situ analysis of the antigen-processing machinery in acute myeloid leukaemic blasts by tissue microarray. Leukemia 2009, 23, 877–885. [Google Scholar] [CrossRef]
- Kamarashev, J.; Ferrone, S.; Seifert, B.; Böni, R.; Nestle, F.O.; Burg, G.; Dummer, R. TAP1 down-regulation in primary melanoma lesions: An independent marker of poor prognosis. Int. J. Cancer 2001, 95, 23–28. [Google Scholar] [CrossRef]
- Hicklin, D.J.; Marincola, F.M.; Ferrone, S. HLA class I antigen downregulation in human cancers: T-cell immunotherapy revives an old story. Mol. Med. Today 1999, 5, 178–186. [Google Scholar] [CrossRef]
- Vitale, M.; Rezzani, R.; Rodella, L.; Zauli, G.; Grigolato, P.; Cadei, M.; Hicklin, D.J.; Ferrone, S. HLA class I antigen and transporter associated with antigen processing (TAP1 and TAP2) down-regulation in high-grade primary breast carcinoma lesions. Cancer Res. 1998, 58, 737–742. [Google Scholar]
- Atkins, D.; Breuckmann, A.; Schmahl, G.E.; Binner, P.; Ferrone, S.; Krummenauer, F.; Störkel, S.; Seliger, B. MHC class I antigen processing pathway defects, ras mutations and disease stage in colorectal carcinoma. Int. J. Cancer 2004, 109, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Hirata, T.; Yamamoto, H.; Taniguchi, H.; Horiuchi, S.; Oki, M.; Adachi, Y.; Imai, K.; Shinomura, Y. Characterization of the immune escape phenotype of human gastric cancers with and without high-frequency microsatellite instability. J. Pathol. 2007, 211, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.L.; Gabrilovich, D.; Tampé, R.; Girgis, K.R.; Nadaf, S.; Carbone, D.P. A functionally defective allele of TAP1 results in loss of MHC class I antigen presentation in a human lung cancer. Nat. Genet. 1996, 13, 210–213. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.C.; Van Hall, T. Alternative Antigen Processing for MHC Class I: Multiple Roads Lead to Rome. Front. Immunol. 2015, 6, 298. [Google Scholar] [CrossRef] [Green Version]
- Van Duinen, S.G.; Ruiter, D.J.; Broecker, E.B.; Van der Velde, E.A.; Sorg, C.; Welvaart, K.; Ferrone, S. Level of HLA antigens in locoregional metastases and clinical course of the disease in patients with melanoma. Cancer Res. 1988, 48, 1019–1025. [Google Scholar]
- Zaloudik, J.; Moore, M.; Ghosh, A.K.; Mechl, Z.; Rejthar, A. DNA content and MHC class II antigen expression in malignant melanoma: Clinical course. J. Clin. Pathol. 1988, 41, 1078–1084. [Google Scholar] [CrossRef] [Green Version]
- Trieb, K.; Lechleitner, T.; Lang, S.; Windhager, R.; Kotz, R.; Dirnhofer, S. Evaluation of HLA-DR expression and T-lymphocyte infiltration in osteosarcoma. Pathol. Res. Pract. 1998, 194, 679–684. [Google Scholar] [CrossRef]
- Moretti, S.; Pinzi, C.; Berti, E.; Spallanzani, A.; Chiarugi, A.; Boddi, V.; Reali, U.M.; Giannotti, B. In situ expression of transforming growth factor beta is associated with melanoma progression and correlates with Ki67, HLA-DR and beta 3 integrin expression. Melanoma Res. 1997, 7, 313–321. [Google Scholar] [CrossRef]
- Diederichsen, A.C.; Stenholm, A.C.; Kronborg, O.; Fenger, C.; Jensenius, J.C.; Zeuthen, J.; Christensen, P.B.; Kristensen, T.; Ostenhom, A.C. Flow cytometric investigation of immune-response-related surface molecules on human colorectal cancers. Int. J. Cancer 1998, 79, 283–287. [Google Scholar] [CrossRef]
- Hilders, C.G.; Houbiers, J.G.; Van Ravenswaay Claasen, H.H.; Veldhuizen, R.W.; Fleuren, G.J. Association between HLA-expression and infiltration of immune cells in cervical carcinoma. Lab. Investig. 1993, 69, 651–659. [Google Scholar]
- Coleman, N.; Stanley, M.A. Analysis of HLA-DR expression on keratinocytes in cervical neoplasia. Int. J. Cancer 1994, 56, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Cromme, F.V.; Meijer, C.J.; Snijders, P.J.; Uyterlinde, A.; Kenemans, P.; Helmerhorst, T.; Stern, P.L.; Van den Brule, A.J.; Walboomers, J.M. Analysis of MHC class I and II expression in relation to presence of HPV genotypes in premalignant and malignant cervical lesions. Br. J. Cancer 1993, 67, 1372–1380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, P.A.; Green, M.A.; Marks, C.G.; King, R.J.; Hubbard, R.; Cook, M.G. Lymphocyte subset infiltration patterns and HLA antigen status in colorectal carcinomas and adenomas. Gut 1996, 38, 85–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsushita, K.; Takenouchi, T.; Kobayashi, S.; Hayashi, H.; Okuyama, K.; Ochiai, T.; Mikata, A.; Isono, K. HLA-DR antigen expression in colorectal carcinomas: Influence of expression by IFN-gamma in situ and its association with tumour progression. Br. J. Cancer 1996, 73, 644–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norheim Andersen, S.; Breivik, J.; Løvig, T.; Meling, G.I.; Gaudernack, G.; Clausen, O.P.; Schjölberg, A.; Fausa, O.; Langmark, F.; Lund, E.; et al. K-ras mutations and HLA-DR expression in large bowel adenomas. Br. J. Cancer 1996, 74, 99–108. [Google Scholar] [CrossRef]
- Anichini, A.; Mortarini, R.; Nonaka, D.; Molla, A.; Vegetti, C.; Montaldi, E.; Wang, X.; Ferrone, S. Association of Antigen-Processing Machinery and HLA Antigen Phenotype of Melanoma Cells with Survival in American Joint Committee on Cancer Stage III and IV Melanoma Patients. Cancer Res. 2006, 66, 6405–6411. [Google Scholar] [CrossRef] [Green Version]
- Sartoris, S.; Valle, M.T.; Barbaro, A.L.; Tosi, G.; Cestari, T.; D’Agostino, A.; Megiovanni, A.M.; Manca, F.; Accolla, R.S. HLA class II expression in uninducible hepatocarcinoma cells after transfection of AIR-1 gene product CIITA: Acquisition of antigen processing and presentation capacity. J. Immunol. 1998, 161, 814–820. [Google Scholar]
- Yazawa, T.; Kamma, H.; Fujiwara, M.; Matsui, M.; Horiguchi, H.; Satoh, H.; Fujimoto, M.; Yokoyama, K.; Ogata, T. Lack of class II transactivator causes severe deficiency of HLA-DR expression in small cell lung cancer. J. Pathol. 1999, 187, 191–199. [Google Scholar] [CrossRef]
- Mach, B.; Steimle, V.; Martinez-Soria, E.; Reith, W. Regulation of MHC class II genes: Lessons from a disease. Annu. Rev. Immunol. 1996, 14, 301–331. [Google Scholar] [CrossRef]
- Bosshart, H.; Jarrett, R.F. Deficient major histocompatibility complex class II antigen presentation in a subset of Hodgkin’s disease tumor cells. Blood 1998, 92, 2252–2259. [Google Scholar] [CrossRef] [Green Version]
- Fife, B.T.; Bluestone, J.A. Control of peripheral T-cell tolerance and autoimmunity via the CTLA-4 and PD-1 pathways. Immunol. Rev. 2008, 224, 166–182. [Google Scholar] [CrossRef]
- Mueller, D.L.; Jenkins, M.K.; Schwartz, R.H. Clonal expansion versus functional clonal inactivation: A costimulatory signalling pathway determines the outcome of T cell antigen receptor occupancy. Annu. Rev. Immunol. 1989, 7, 445–480. [Google Scholar] [CrossRef] [PubMed]
- Schneider, H.; Downey, J.; Smith, A.; Zinselmeyer, B.H.; Rush, C.; Brewer, J.M.; Wei, B.; Hogg, N.; Garside, P.; Rudd, C.E. Reversal of the TCR stop signal by CTLA-4. Science 2006, 313, 1972–1975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, H.; Honjo, T. PD-1: An inhibitory immunoreceptor involved in peripheral tolerance. Trends Immunol. 2001, 22, 265–268. [Google Scholar] [CrossRef]
- Ribas, A. Releasing the Brakes on Cancer Immunotherapy. N. Engl. J. Med. 2015, 373, 1490–1492. [Google Scholar] [CrossRef] [PubMed]
- Webb, E.S.; Liu, P.; Baleeiro, R.; Lemoine, N.R.; Yuan, M.; Wang, Y.-H. Immune checkpoint inhibitors in cancer therapy. J. Biomed. Res. 2018, 32, 317–326. [Google Scholar] [CrossRef]
- Keung, E.Z.; Wargo, J.A. The Current Landscape of Immune Checkpoint Inhibition for Solid Malignancies. Surg. Oncol. Clin. N. Am. 2019, 28, 369–386. [Google Scholar] [CrossRef]
- Pitt, J.M.; Vétizou, M.; Daillère, R.; Roberti, M.P.; Yamazaki, T.; Routy, B.; Lepage, P.; Boneca, I.G.; Chamaillard, M.; Kroemer, G.; et al. Resistance Mechanisms to Immune-Checkpoint Blockade in Cancer: Tumor-Intrinsic and -Extrinsic Factors. Immunity 2016, 44, 1255–1269. [Google Scholar] [CrossRef] [Green Version]
- Restifo, N.P.; Smyth, M.J.; Snyder, A. Acquired resistance to immunotherapy and future challenges. Nat. Rev. Cancer 2016, 16, 121–126. [Google Scholar] [CrossRef]
- Jenkins, R.W.; Barbie, D.A.; Flaherty, K.T. Mechanisms of resistance to immune checkpoint inhibitors. Br. J. Cancer 2018, 118, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Huang, Z.; Teng, F.; Xing, L.; Yu, J. Predictive biomarkers in PD-1/PD-L1 checkpoint blockade immunotherapy. Cancer Treat. Rev. 2015, 41, 868–876. [Google Scholar] [CrossRef] [PubMed]
- Maleki Vareki, S.; Garrigós, C.; Duran, I. Biomarkers of response to PD-1/PD-L1 inhibition. Crit. Rev. Oncol. Hematol. 2017, 116, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Manson, G.; Norwood, J.; Marabelle, A.; Kohrt, H.; Houot, R. Biomarkers associated with checkpoint inhibitors. Ann. Oncol. 2016, 27, 1199–1206. [Google Scholar] [CrossRef] [PubMed]
- Gibney, G.T.; Weiner, L.M.; Atkins, M.B. Predictive biomarkers for checkpoint inhibitor-based immunotherapy. Lancet Oncol. 2016, 17, e542–e551. [Google Scholar] [CrossRef] [Green Version]
- Zaretsky, J.M.; Garcia-Diaz, A.; Shin, D.S.; Escuin-Ordinas, H.; Hugo, W.; Hu-Lieskovan, S.; Torrejon, D.Y.; Abril-Rodriguez, G.; Sandoval, S.; Barthly, L.; et al. Mutations Associated with Acquired Resistance to PD-1 Blockade in Melanoma. N. Engl. J. Med. 2016, 375, 819–829. [Google Scholar] [CrossRef]
- Kametani, Y.; Ohno, Y.; Ohshima, S.; Tsuda, B.; Yasuda, A.; Seki, T.; Ito, R.; Tokuda, Y. Humanized Mice as an Effective Evaluation System for Peptide Vaccines and Immune Checkpoint Inhibitors. Int. J. Mol. Sci. 2019, 20, 6337. [Google Scholar] [CrossRef] [Green Version]
- Lechner, M.G.; Karimi, S.S.; Barry-Holson, K.; Angell, T.E.; Murphy, K.A.; Church, C.H.; Ohlfest, J.R.; Hu, P.; Epstein, A.L. Immunogenicity of murine solid tumor models as a defining feature of in vivo behavior and response to immunotherapy. J. Immunother. 2013, 36, 477–489. [Google Scholar] [CrossRef] [Green Version]
- Ashizawa, T.; Iizuka, A.; Nonomura, C.; Kondou, R.; Maeda, C.; Miyata, H.; Sugino, T.; Mitsuya, K.; Hayashi, N.; Nakasu, Y.; et al. Antitumor Effect of Programmed Death-1 (PD-1) Blockade in Humanized the NOG-MHC Double Knockout Mouse. Clin. Cancer Res. 2017, 23, 149–158. [Google Scholar] [CrossRef] [Green Version]
- Gettinger, S.; Choi, J.; Hastings, K.; Truini, A.; Datar, I.; Sowell, R.; Wurtz, A.; Dong, W.; Cai, G.; Melnick, M.A.; et al. Impaired HLA Class I Antigen Processing and Presentation as a Mechanism of Acquired Resistance to Immune Checkpoint Inhibitors in Lung Cancer. Cancer Discov. 2017, 7, 1420–1435. [Google Scholar] [CrossRef] [Green Version]
- Lhotakova, K.; Grzelak, A.; Polakova, I.; Vackova, J.; Smahel, M. Establishment and characterization of a mouse tumor cell line with irreversible downregulation of MHC class I molecules. Oncol. Rep. 2019, 42, 2826–2835. [Google Scholar] [CrossRef]
- Rangan, L.; Galaine, J.; Boidot, R.; Hamieh, M.; Dosset, M.; Francoual, J.; Beziaud, L.; Pallandre, J.-R.; Lauret Marie Joseph, E.; Asgarova, A.; et al. Identification of a novel PD-L1 positive solid tumor transplantable in HLA-A*0201/DRB1*0101 transgenic mice. Oncotarget 2017, 8, 48959–48971. [Google Scholar] [CrossRef] [PubMed]
- Mortara, L.; Castellani, P.; Meazza, R.; Tosi, G.; De Lerma Barbaro, A.; Procopio, F.A.; Comes, A.; Zardi, L.; Ferrini, S.; Accolla, R.S. CIITA-induced MHC class II expression in mammary adenocarcinoma leads to a Th1 polarization of the tumor microenvironment, tumor rejection, and specific antitumor memory. Clin. Cancer Res. 2006, 12, 3435–3443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murciano-Goroff, Y.R.; Warner, A.B.; Wolchok, J.D. The future of cancer immunotherapy: Microenvironment-targeting combinations. Cell Res. 2020, 30, 507–519. [Google Scholar] [CrossRef] [PubMed]
- Chowell, D.; Morris, L.G.T.; Grigg, C.M.; Weber, J.K.; Samstein, R.M.; Makarov, V.; Kuo, F.; Kendall, S.M.; Requena, D.; Riaz, N.; et al. Patient HLA class I genotype influences cancer response to checkpoint blockade immunotherapy. Science 2018, 359, 582–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrido, F.; Aptsiauri, N.; Doorduijn, E.M.; Garcia Lora, A.M.; Van Hall, T. The urgent need to recover MHC class I in cancers for effective immunotherapy. Curr. Opin. Immunol. 2016, 39, 44–51. [Google Scholar] [CrossRef] [Green Version]
- Garrido, F.; Aptsiauri, N. Cancer immune escape: MHC expression in primary tumours versus metastases. Immunology 2019, 158, 255–266. [Google Scholar] [CrossRef] [Green Version]
- Banik, D.; Moufarrij, S.; Villagra, A. Immunoepigenetics Combination Therapies: An Overview of the Role of HDACs in Cancer Immunotherapy. Int. J. Mol. Sci. 2019, 20, 2241. [Google Scholar] [CrossRef] [Green Version]
- Del Campo, A.B.; Carretero, J.; Muñoz, J.A.; Zinchenko, S.; Ruiz-Cabello, F.; González-Aseguinolaza, G.; Garrido, F.; Aptsiauri, N. Adenovirus expressing β2-microglobulin recovers HLA class I expression and antitumor immunity by increasing T-cell recognition. Cancer Gene Ther. 2014, 21, 317–332. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Waschke, B.C.; Woolaver, R.A.; Chen, Z.; Zhang, G.; Piscopio, A.D.; Liu, X.; Wang, J.H. Histone Deacetylase Inhibition Sensitizes PD1 Blockade-Resistant B-cell Lymphomas. Cancer Immunol. Res. 2019, 7, 1318–1331. [Google Scholar] [CrossRef] [Green Version]
- Ramsuran, V.; Kulkarni, S.; O’huigin, C.; Yuki, Y.; Augusto, D.G.; Gao, X.; Carrington, M. Epigenetic regulation of differential HLA-A allelic expression levels. Hum. Mol. Genet. 2015, 24, 4268–4275. [Google Scholar] [CrossRef] [Green Version]
- Brea, E.J.; Oh, C.Y.; Manchado, E.; Budhu, S.; Gejman, R.S.; Mo, G.; Mondello, P.; Han, J.E.; Jarvis, C.A.; Ulmert, D.; et al. Kinase Regulation of Human MHC Class I Molecule Expression on Cancer Cells. Cancer Immunol. Res. 2016, 4, 936–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loi, S.; Dushyanthen, S.; Beavis, P.A.; Salgado, R.; Denkert, C.; Savas, P.; Combs, S.; Rimm, D.L.; Giltnane, J.M.; Estrada, M.V.; et al. RAS/MAPK Activation Is Associated with Reduced Tumor-Infiltrating Lymphocytes in Triple-Negative Breast Cancer: Therapeutic Cooperation Between MEK and PD-1/PD-L1 Immune Checkpoint Inhibitors. Clin. Cancer Res. 2016, 22, 1499–1509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, P.K. Neoepitopes of Cancers: Looking Back, Looking Ahead. Cancer Immunol. Res. 2015, 3, 969–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Im, J.S.; Herrmann, A.C.; Bernatchez, C.; Haymaker, C.; Molldrem, J.J.; Hong, W.K.; Perez-Soler, R. Immune-Modulation by Epidermal Growth Factor Receptor Inhibitors: Implication on Anti-Tumor Immunity in Lung Cancer. PLoS ONE 2016, 11, e0160004. [Google Scholar] [CrossRef] [Green Version]
- Garrido, G.; Rabasa, A.; Garrido, C.; Chao, L.; Garrido, F.; García-Lora, Á.M.; Sánchez-Ramírez, B. Upregulation of HLA Class I Expression on Tumor Cells by the Anti-EGFR Antibody Nimotuzumab. Front. Pharmacol. 2017, 8, 595. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.-H.; Keam, B.; Ahn, Y.-O.; Park, H.-R.; Kim, M.; Kim, T.M.; Kim, D.-W.; Heo, D.S. Inhibition of MEK with trametinib enhances the efficacy of anti-PD-L1 inhibitor by regulating anti-tumor immunity in head and neck squamous cell carcinoma. Oncoimmunology 2019, 8, e1515057. [Google Scholar] [CrossRef] [Green Version]
- Sapkota, B.; Hill, C.E.; Pollack, B.P. Vemurafenib enhances MHC induction in BRAFV600E homozygous melanoma cells. Oncoimmunology 2013, 2, e22890. [Google Scholar] [CrossRef] [Green Version]
- Sabbatino, F.; Wang, Y.; Scognamiglio, G.; Favoino, E.; Feldman, S.A.; Villani, V.; Flaherty, K.T.; Nota, S.; Giannarelli, D.; Simeone, E.; et al. Antitumor Activity of BRAF Inhibitor and IFNα Combination in BRAF-Mutant Melanoma. J. Natl. Cancer Inst. 2016, 108. [Google Scholar] [CrossRef] [Green Version]
- Hu-Lieskovan, S.; Mok, S.; Homet Moreno, B.; Tsoi, J.; Robert, L.; Goedert, L.; Pinheiro, E.M.; Koya, R.C.; Graeber, T.G.; Comin-Anduix, B.; et al. Improved antitumor activity of immunotherapy with BRAF and MEK inhibitors in BRAF(V600E) melanoma. Sci. Transl. Med. 2015, 7, 279ra41. [Google Scholar] [CrossRef] [Green Version]
- Ritter, C.; Fan, K.; Paschen, A.; Reker Hardrup, S.; Ferrone, S.; Nghiem, P.; Ugurel, S.; Schrama, D.; Becker, J.C. Epigenetic priming restores the HLA class-I antigen processing machinery expression in Merkel cell carcinoma. Sci. Rep. 2017, 7, 2290. [Google Scholar] [CrossRef] [Green Version]
- Sun, T.; Li, Y.; Yang, W.; Wu, H.; Li, X.; Huang, Y.; Zhou, Y.; Du, Z. Histone deacetylase inhibition up-regulates MHC class I to facilitate cytotoxic T lymphocyte-mediated tumor cell killing in glioma cells. J. Cancer 2019, 10, 5638–5645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Li, Y.; Gao, R.; Xiu, Z.; Sun, T. MHC class I dysfunction of glioma stem cells escapes from CTL-mediated immune response via activation of Wnt/β-catenin signaling pathway. Oncogene 2020, 39, 1098–1111. [Google Scholar] [CrossRef] [PubMed]
- Woan, K.V.; Lienlaf, M.; Perez-Villaroel, P.; Lee, C.; Cheng, F.; Knox, T.; Woods, D.M.; Barrios, K.; Powers, J.; Sahakian, E.; et al. Targeting histone deacetylase 6 mediates a dual anti-melanoma effect: Enhanced antitumor immunity and impaired cell proliferation. Mol. Oncol. 2015, 9, 1447–1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ennishi, D.; Takata, K.; Béguelin, W.; Duns, G.; Mottok, A.; Farinha, P.; Bashashati, A.; Saberi, S.; Boyle, M.; Meissner, B.; et al. Molecular and Genetic Characterization of MHC Deficiency Identifies EZH2 as Therapeutic Target for Enhancing Immune Recognition. Cancer Discov. 2019, 9, 546–563. [Google Scholar] [CrossRef] [Green Version]
- Fonsatti, E.; Nicolay, H.J.M.; Sigalotti, L.; Calabrò, L.; Pezzani, L.; Colizzi, F.; Altomonte, M.; Guidoboni, M.; Marincola, F.M.; Maio, M. Functional up-regulation of human leukocyte antigen class I antigens expression by 5-aza-2’-deoxycytidine in cutaneous melanoma: Immunotherapeutic implications. Clin. Cancer Res. 2007, 13, 3333–3338. [Google Scholar] [CrossRef] [Green Version]
- Chacon, J.A.; Schutsky, K.; Powell, D.J. The Impact of Chemotherapy, Radiation and Epigenetic Modifiers in Cancer Cell Expression of Immune Inhibitory and Stimulatory Molecules and Anti-Tumor Efficacy. Vaccines 2016, 4, 43. [Google Scholar] [CrossRef] [Green Version]
- Luo, N.; Nixon, M.J.; Gonzalez-Ericsson, P.I.; Sanchez, V.; Opalenik, S.R.; Li, H.; Zahnow, C.A.; Nickels, M.L.; Liu, F.; Tantawy, M.N.; et al. DNA methyltransferase inhibition upregulates MHC-I to potentiate cytotoxic T lymphocyte responses in breast cancer. Nat. Commun. 2018, 9, 248. [Google Scholar] [CrossRef]
- De Biasi, A.R.; Villena-Vargas, J.; Adusumilli, P.S. Cisplatin-induced antitumor immunomodulation: A review of preclinical and clinical evidence. Clin. Cancer Res. 2014, 20, 5384–5391. [Google Scholar] [CrossRef] [Green Version]
- Faè, D.A.; Martorelli, D.; Mastorci, K.; Muraro, E.; Dal Col, J.; Franchin, G.; Barzan, L.; Comaro, E.; Vaccher, E.; Rosato, A.; et al. Broadening Specificity and Enhancing Cytotoxicity of Adoptive T Cells for Nasopharyngeal Carcinoma Immunotherapy. Cancer Immunol. Res. 2016, 4, 431–440. [Google Scholar] [CrossRef] [Green Version]
- Pellicciotta, I.; Yang, C.-P.H.; Goldberg, G.L.; Shahabi, S. Epothilone B enhances Class I HLA and HLA-A2 surface molecule expression in ovarian cancer cells. Gynecol. Oncol. 2011, 122, 625–631. [Google Scholar] [CrossRef]
- Wan, S.; Pestka, S.; Jubin, R.G.; Lyu, Y.L.; Tsai, Y.-C.; Liu, L.F. Chemotherapeutics and radiation stimulate MHC class I expression through elevated interferon-beta signaling in breast cancer cells. PLoS ONE 2012, 7, e32542. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Waschke, B.C.; Woolaver, R.A.; Chen, S.M.Y.; Chen, Z.; Wang, J.H. HDAC inhibitors overcome immunotherapy resistance in B-cell lymphoma. Protein Cell 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, F. Molecular mechanisms of IFN-gamma to up-regulate MHC class I antigen processing and presentation. Int. Rev. Immunol. 2009, 28, 239–260. [Google Scholar] [CrossRef] [PubMed]
- De Charette, M.; Marabelle, A.; Houot, R. Turning tumour cells into antigen presenting cells: The next step to improve cancer immunotherapy? Eur. J. Cancer 2016, 68, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Medrano, R.F.V.; Hunger, A.; Mendonça, S.A.; Barbuto, J.A.M.; Strauss, B.E. Immunomodulatory and antitumor effects of type I interferons and their application in cancer therapy. Oncotarget 2017, 8, 71249–71284. [Google Scholar] [CrossRef] [Green Version]
- Selinger, E.; Reiniš, M. Epigenetic View on Interferon γ Signalling in Tumour Cells. Folia Biol. (Praha) 2018, 64, 125–136. [Google Scholar]
- Zhang, S.; Kohli, K.; Black, R.G.; Yao, L.; Spadinger, S.M.; He, Q.; Pillarisetty, V.G.; Cranmer, L.D.; Van Tine, B.A.; Yee, C.; et al. Systemic Interferon-γ Increases MHC Class I Expression and T-cell Infiltration in Cold Tumors: Results of a Phase 0 Clinical Trial. Cancer Immunol. Res. 2019, 7, 1237–1243. [Google Scholar] [CrossRef] [Green Version]
- Vlková, V.; Štěpánek, I.; Hrušková, V.; Šenigl, F.; Mayerová, V.; Šrámek, M.; Šímová, J.; Bieblová, J.; Indrová, M.; Hejhal, T.; et al. Epigenetic regulations in the IFNγ signalling pathway: IFNγ-mediated MHC class I upregulation on tumour cells is associated with DNA demethylation of antigen-presenting machinery genes. Oncotarget 2014, 5, 6923–6935. [Google Scholar] [CrossRef]
- Castro, F.; Cardoso, A.P.; Gonçalves, R.M.; Serre, K.; Oliveira, M.J. Interferon-Gamma at the Crossroads of Tumor Immune Surveillance or Evasion. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, T.; Méndez, R.; Del Campo, A.; Jiménez, P.; Aptsiauri, N.; Garrido, F.; Ruiz-Cabello, F. Distinct mechanisms of loss of IFN-gamma mediated HLA class I inducibility in two melanoma cell lines. BMC Cancer 2007, 7, 34. [Google Scholar] [CrossRef] [Green Version]
- Sucker, A.; Zhao, F.; Pieper, N.; Heeke, C.; Maltaner, R.; Stadtler, N.; Real, B.; Bielefeld, N.; Howe, S.; Weide, B.; et al. Acquired IFNγ resistance impairs anti-tumor immunity and gives rise to T-cell-resistant melanoma lesions. Nat. Commun. 2017, 8, 15440. [Google Scholar] [CrossRef] [PubMed]
- Aspeslagh, S.; Morel, D.; Soria, J.-C.; Postel-Vinay, S. Epigenetic modifiers as new immunomodulatory therapies in solid tumours. Ann. Oncol. 2018, 29, 812–824. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Zhao, Z.-L.; Zhao, W.-T.; Fan, Q.-R.; Wang, S.-C.; Li, J.; Zhang, Y.-Q.; Shi, J.-W.; Lin, X.-L.; Yang, S.; et al. miR-9 modulates the expression of interferon-regulated genes and MHC class I molecules in human nasopharyngeal carcinoma cells. Biochem. Biophys. Res. Commun. 2013, 431, 610–616. [Google Scholar] [CrossRef]
- Li, J.; Lin, T.-Y.; Chen, L.; Liu, Y.; Dian, M.-J.; Hao, W.-C.; Lin, X.-L.; Li, X.-Y.; Li, Y.-L.; Lian, M.; et al. miR-19 regulates the expression of interferon-induced genes and MHC class I genes in human cancer cells. Int. J. Med. Sci. 2020, 17, 953–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Name | Target | Combination Therapy | Cancer Type | References |
|---|---|---|---|---|
| Kinase Inhibitors | ||||
| Nimotuzumab Cetuximab Erlotinib | EGFR | IFN-γ | epidermoid carcinoma lung cancer head and neck carcinoma lung adenocarcinoma | Garrido G. et al. 2017 [236] Srivastava et al. 2015 [234] Im et al. 2016 [235] |
| Trametinib | MEK1/2 | IFN-γ | mesothelioma melanoma lung adenocarcinoma colon adenocarcinoma thyroid carcinoma breast cancer head and neck carcinoma | Brea et al. 2016 [232] Loi et al. 2016 [233] Kang et al. 2019 [237] |
| Vemurafenib | BRAFV600E | IFN-γ IFNα-2b | Melanoma | Sapkota et al. 2013 [238] Sabbatino et al. 2016 [239] |
| Dabrafenib | Trametinib | Hu-Lieskovan et al. 2015 [240] | ||
| Epigenetic Agents | ||||
| Vorinostat | HDAC class I, II, and IV | Mithramycin A | Merkel cell carcinoma | Ritter et al. 2017 [241] |
| B-cell lymphoma cutaneous T-cell lymphoma acute myeloid leukemia glioma | Chacon et al. 2016 [247] Banik et al. 2019 [228] Sun et al. 2019 [242] Yang et al. 2019 [243] Wang et al. 2020 [253] | |||
| Belinostat | peripheral T-cell lymphoma B-cell lymphoma | Banik et al. 2019 [228] Wang et al. 2020 [253] | ||
| Panobinostat | multiple myeloma B-cell lymphoma | Banik et al. 2019 [228] Wang et al. 2020 [253] | ||
| OKI-179 | diffuse large B-cell lymphoma | Wang et al. 2019 [230] | ||
| Romidepsin | HDAC 1-2 | B-cell lymphoma peripheral T cell lymphoma | Banik et al. 2019 [228] Wang et al. 2020 [253] | |
| Tubastatin A | HDAC 6 | Melanoma | Woan et al. 2015 [244] | |
| Azacytidine | DNMT | lung carcinoma melanoma | Fonsatti et al. 2007 [246] Chacon et al. 2016 [247] | |
| Guadecitabine | breast cancer | Luo et al. 2018 [248] | ||
| Tazemetostat | EZH2 | diffuse large B-cell lymphoma | Ennishi et al. 2019 [245] | |
| Chemotherapeutics | ||||
| Cisplatin | DNA | Vinorelbine or5-fluorouracil | lung cancer head and neck carcinoma | De Biasi et al. 2014 [249] |
| Epothilone B Taxol Vinblastine | Microtubules | ovarian cancer | Pellicciotta et al. 2011 [251] | |
| Doxorubicin | Topoisomerase | nasopharyngeal carcinoma | Faè et al. 2016 [250] | |
| Topotecan Etoposide | breast cancer | Wan et al. 2012 [252] | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sabbatino, F.; Liguori, L.; Polcaro, G.; Salvato, I.; Caramori, G.; Salzano, F.A.; Casolaro, V.; Stellato, C.; Dal Col, J.; Pepe, S. Role of Human Leukocyte Antigen System as A Predictive Biomarker for Checkpoint-Based Immunotherapy in Cancer Patients. Int. J. Mol. Sci. 2020, 21, 7295. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197295
Sabbatino F, Liguori L, Polcaro G, Salvato I, Caramori G, Salzano FA, Casolaro V, Stellato C, Dal Col J, Pepe S. Role of Human Leukocyte Antigen System as A Predictive Biomarker for Checkpoint-Based Immunotherapy in Cancer Patients. International Journal of Molecular Sciences. 2020; 21(19):7295. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197295
Chicago/Turabian StyleSabbatino, Francesco, Luigi Liguori, Giovanna Polcaro, Ilaria Salvato, Gaetano Caramori, Francesco A. Salzano, Vincenzo Casolaro, Cristiana Stellato, Jessica Dal Col, and Stefano Pepe. 2020. "Role of Human Leukocyte Antigen System as A Predictive Biomarker for Checkpoint-Based Immunotherapy in Cancer Patients" International Journal of Molecular Sciences 21, no. 19: 7295. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197295