Effect of Vesicle Size on the Cytolysis of Cell-Penetrating Peptides (CPPs)

 , ,
, , 
Abstract
:1. Introduction
2. Results
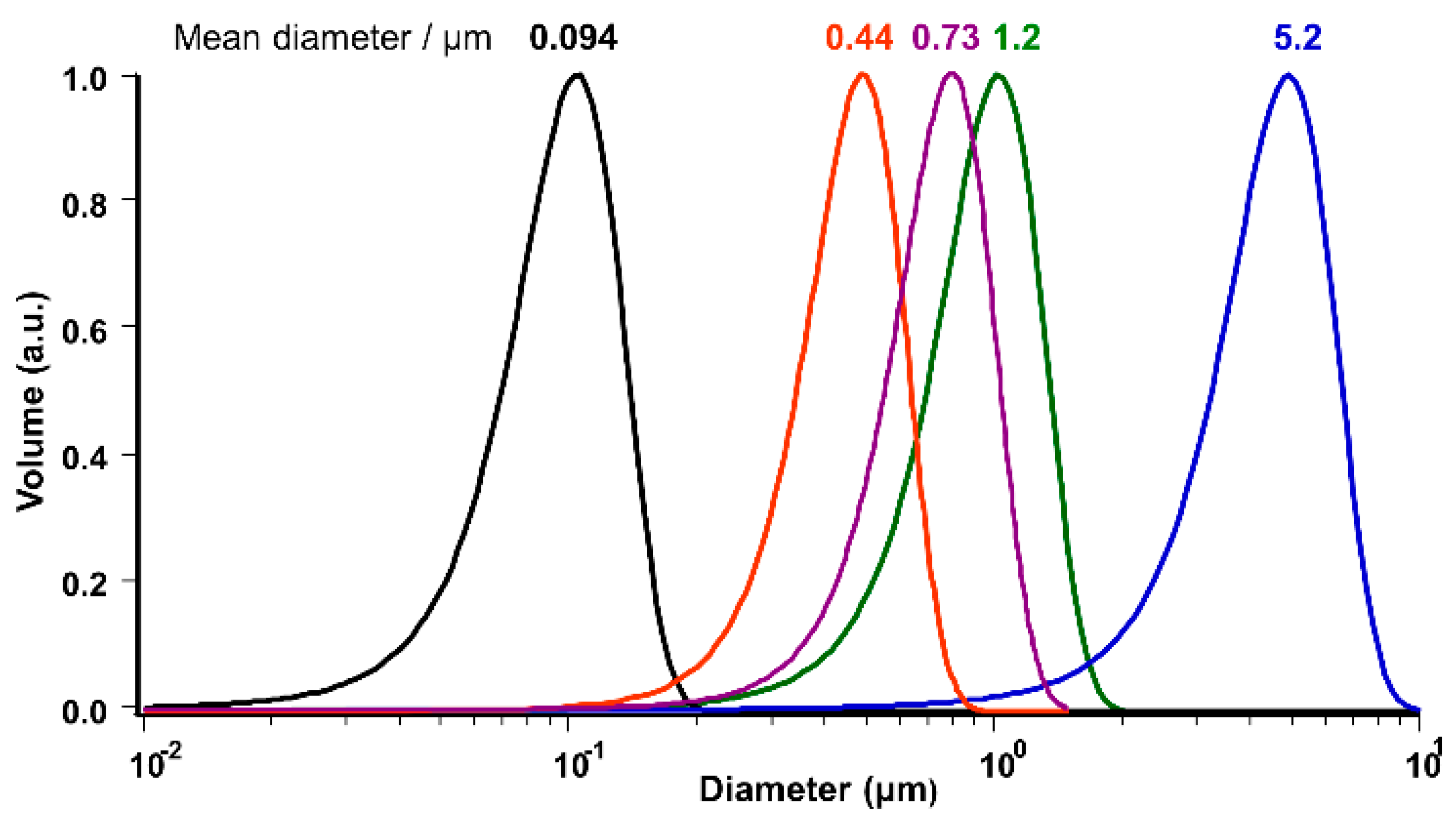
2.1. Confirmation of Vesicle Formation
2.2. Calculation of the Geometrical Parameters for Vesicles
2.3. Measurement of Permeation Amount of Cell-Penetrating Peptide (CPP)
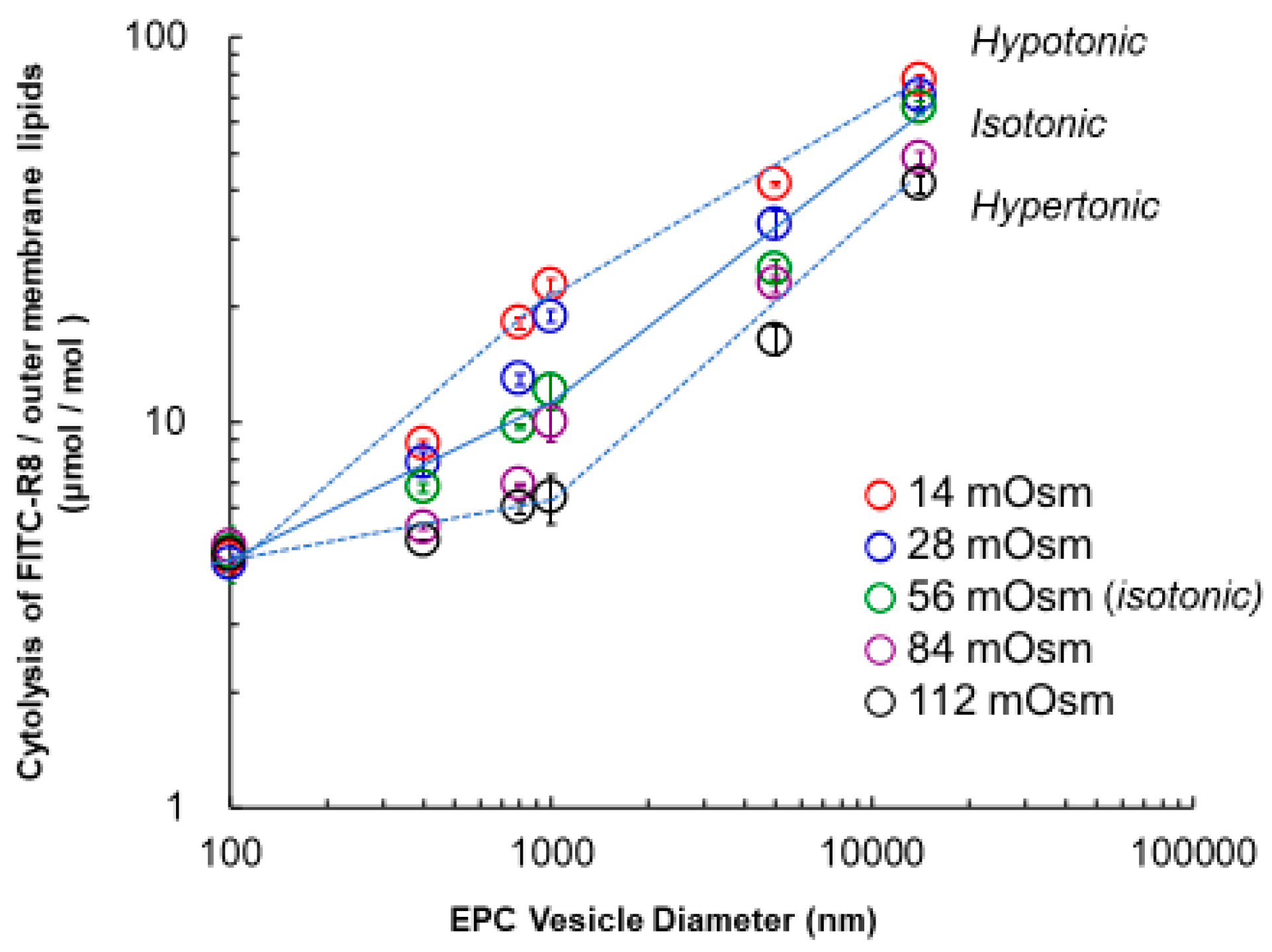
2.3.1. Effect of Particle Size of Vesicle on the Cytolysis of FITC–R8 as CPP for EPC Vesicles
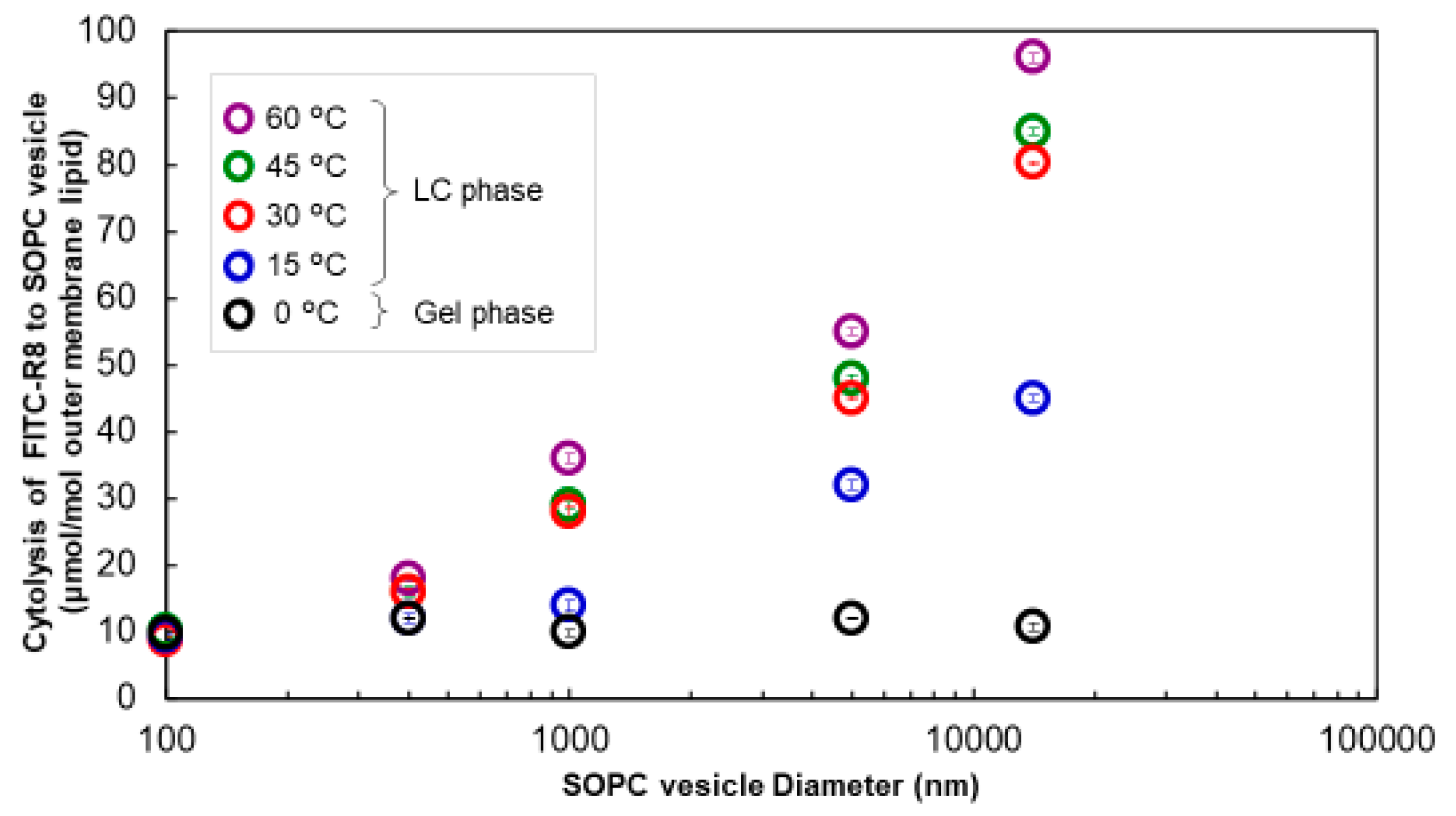
2.3.2. Measurement of CPP Permeation under the Gel Phase
2.4. Effect of Particle Size of the SOPC Vesicle on the Physicochemical Properties of the Lipid Membrane
2.4.1. Fluorescent Anisotropy Measurement for Lipid Mobility Determination
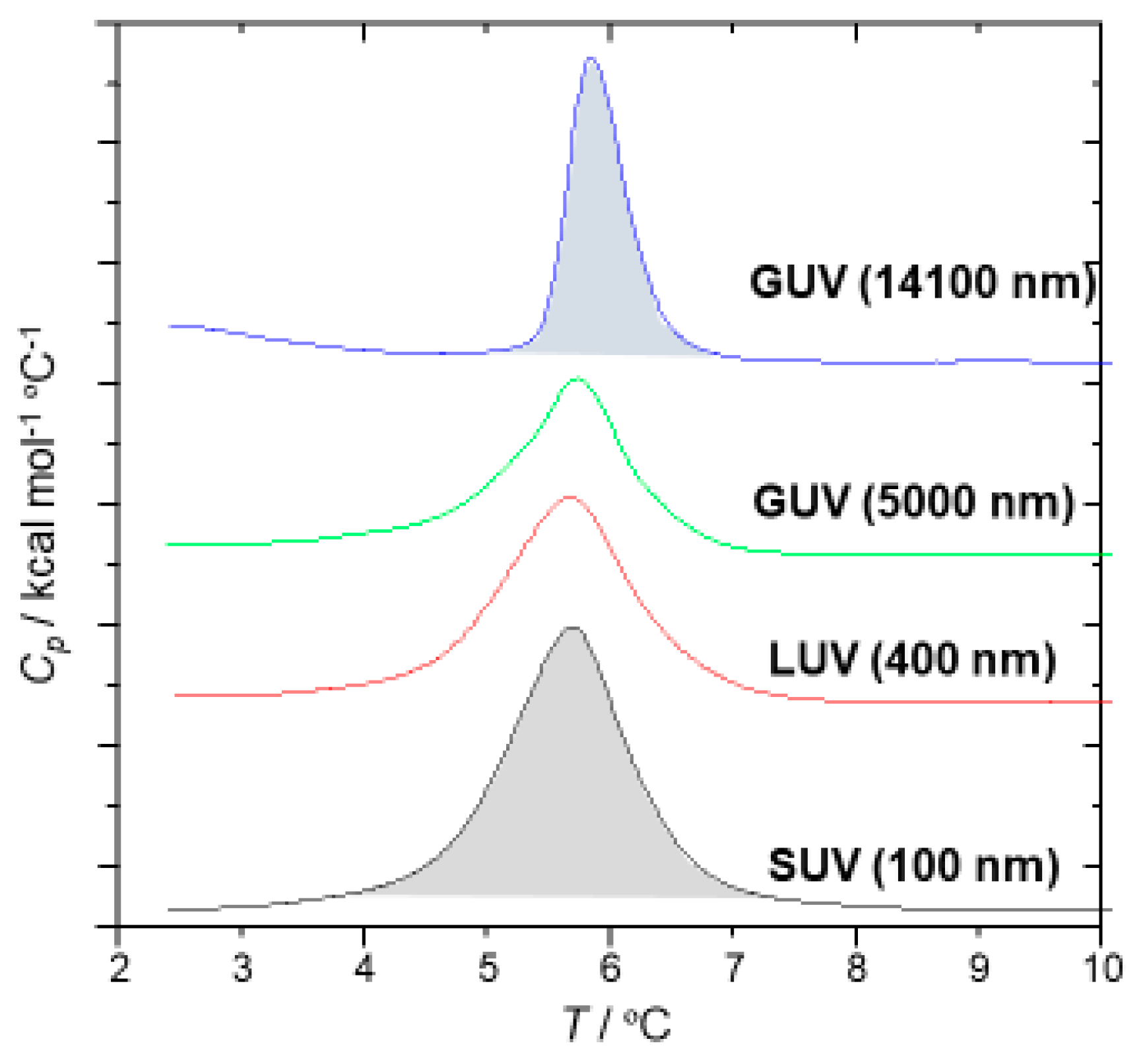
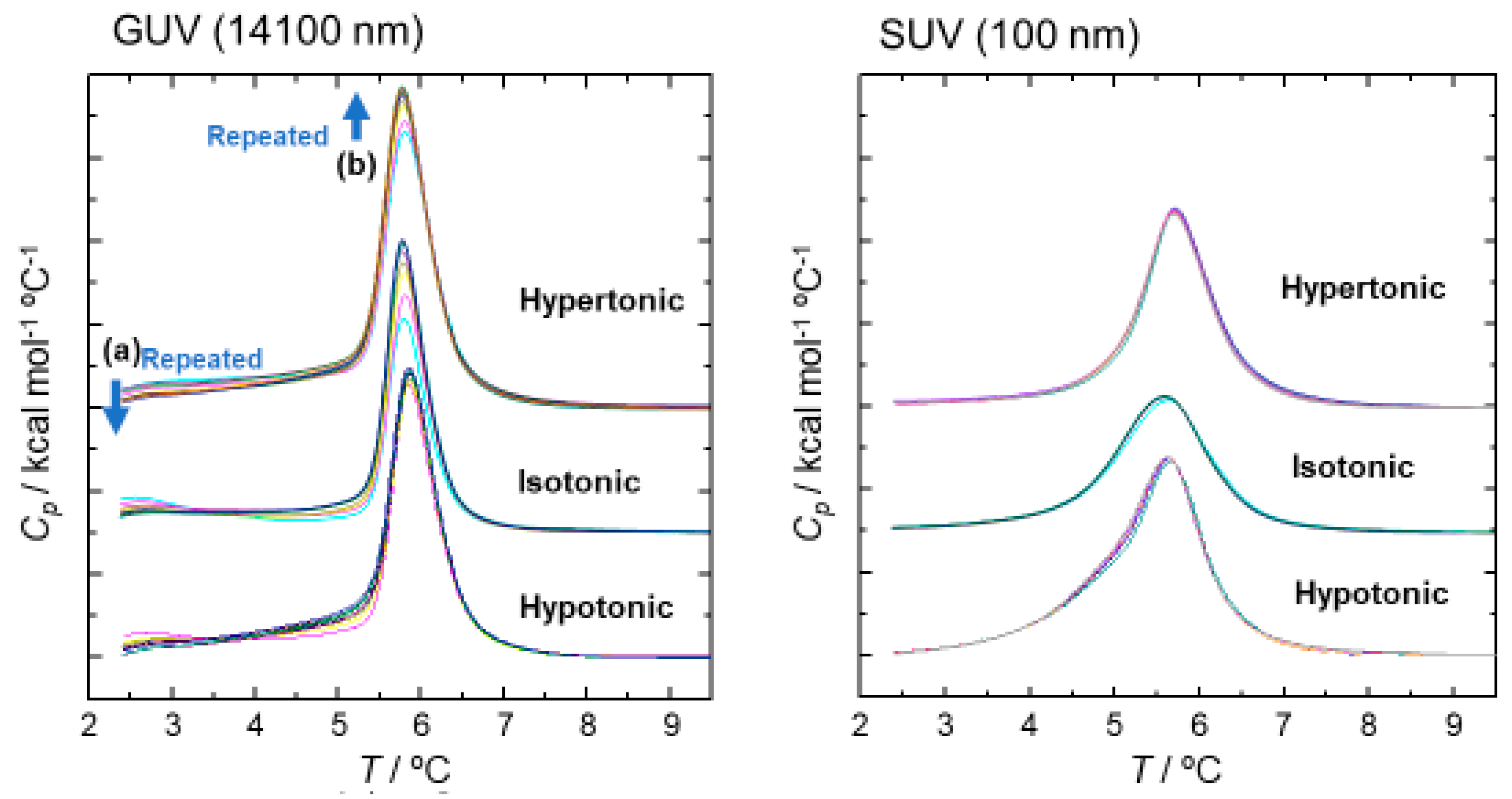
2.4.2. Differential Scanning Calorimetry (DSC) Measurement for the Thermal Properties of SOPC Vesicles
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Methods
4.2.1. Preparation of Phosphate-Buffered Saline (PBS(−))
4.2.2. Preparation of Vesicles
4.2.3. Phospholipid Concentration Measurement
4.2.4. Preparation of Trypsin Solution
4.2.5. Confirmation of GUV Formation
4.2.6. Particle Size Distribution Measurement of GUVs
4.2.7. Measurement of Cytolysis Amount of CPPs
4.2.8. Fluorescence Anisotropy Measurements
4.2.9. Calorimetric Measurement by DSC
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CPP | Cell-penetrating peptide |
| FITC | Fluorescein isothiocyanate |
| SP | Surfactant parameter |
| TEM | Transmission electron microscope |
| DLS | Dynamic light scattering |
| DSC | Differential scanning calorimetry |
References
- Futaki, S.; Nakase, I. Cell-Surface Interactions on Arginine-Rich Cell-Penetrating Peptides Allow for Multiplex Modes of Internalization. Acc. Chem. Res. 2017, 50, 2449–2456. [Google Scholar] [CrossRef] [PubMed]
- Futaki, S. Oligoarginine vectors for intracellular delivery: Design and cellular-uptake mechanisms. Biopolymers 2006, 84, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Futaki, S.; Niwa, M.; Tanaka, S.; Ueda, K.; Sugiura, Y. Possible Existence of Common Internalization Mechanizm among Arginine-rich Peptides. J. Biol. Chem. 2002, 277, 2437–2443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanzel, E.G.; Trantow, B.M.; Vargas, J.R.; Wender, P.A. Fifteen Years of Cell-Penetrating, Guanidinium-Rich Molecular Transporters; Basic Science, Research Tools, and Clinical Applications. Acc. Chem. Res. 2013, 46, 2944–2954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakamoto, K.; Takino, Y.; Ogasahara, K. Method for Controlling Membrane Permeability of a Membrane Permeable Substance and Screening Method for a Membrane Permeable Substance. U.S. Patent 20050118204A1, 4 November 2003. [Google Scholar]
- Sakamoto, K.; Takino, Y.; Ogasahara, K.; Hemayet, U.; Futaki, S. Key Mechanism Triggering Cell-Penetrating Peptide to Cross Membrane. In Proceedings of the 13th IACIS & 83rd ACS Colloid and Surface Science Symposium, New York, NY, USA, 19 June 2009. [Google Scholar]
- Israelachvili, J.N.; Mitchell, D.J.; Ninham, B.W. Theory of self-assembly of hydrocarbon amphiphiles into micelles and bilayers. J. Chem. Soc. Faraday Trans. 1976, 72, 1525–1568. [Google Scholar] [CrossRef]
- Hyde, S.; Andersson, S.; Larsson, K.; Blum, Z.; Landh, T.; Linden, S.; Ninham, B.W. The Language of Shape; Elsevier: Amsterdam, The Netherlands, 1997; pp. 143–146. [Google Scholar]
- Sakamoto, K.; Abrai, K.; Morishita, T.; Sakai, K.; Abe, M.; Nakase, I.; Futaki, S. Bioinspired Mechanism for the Translocation of Peptide through the Cell Membrane. Chem. Lett. 2012, 41, 1078–1080. [Google Scholar] [CrossRef]
- Hyde, S.T.; Schroder, G.E. Novel surfactant mesostructural topologies: Between lamellae and columnar (hexagonal) forms. Curr. Opin. Colloid Interface Sci. 2003, 8, 5–14. [Google Scholar] [CrossRef]
- Sakamoto, K.; Morishita, T.; Aburai, K.; Ito, D.; Imura, T.; Sakai, K.; Abe, M.; Nakase, I.; Futaki, S.; Sakai, H. Direct entry of Cell-Penetrating Peptide can be controlled by maneuvering the membrane curvature. Sci. Rep. 2020. under review. [Google Scholar]
- Sakamoto, K.; Morishita, T.; Abrai, K.; Sakai, K.; Abe, M.; Nakase, I.; Futaki, S.; Sakai, H. Key Process and Factors Controlling the Direct Translocation of Cell-Penetrating Peptide through Bio-Membrane. Int. J. Mol. Sci. 2020, 21, 5466. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, K. The Importance of Planarity for Lipid Bilayers as Biomembranes. Adv. Biomembr. Lipid Self Assem. 2016, 23, 1–23. [Google Scholar]
- Sakuma, Y.; Imai, M. From Vesicles to Protocells: The Roles of Amphiphilic Molecules. Life 2015, 5, 651–675. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, V.; Svetins, S.; Zeks, B. Non asymmetric vesicle shapes. Phys. Rev. E 1993, 48, 3112–3123. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.L.; Seifert, U.; Wortis, M.; Dobereinner, H.G. Budding transitions of fluid-bilayer vesicles: The eectof area-dierence elasticity. Phys. Rev. E 1994, 49, 5389–5407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharmin, S.; Islam, Z.; Karal, M.A.S.; Shibly, S.U.A.; Dohra, H.; Yamazaki, M. Eects of Lipid Composition on the Entry of Cell-Penetrating Peptide Oligoarginine into Single Vesicles. Biochemistry 2016, 55, 4154–4165. [Google Scholar] [CrossRef] [PubMed]
- Wheaten, S.A.; Ablan, F.D.O.; Spaller, B.L.; Trieu, J.M.; Almeida, P.F. Translocation of Cationic Amphipathic Peptides across the Membranes of Pure Phospholipid Giant Vesicles. J. Am. Chem. Soc. 2013, 135, 16517–16525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, H.; Goto, Y.; Tamai, N. Membrane States of Lipids in Biological Membranes Structure-Function Relationship Revealed from Pressure Study. Sci. Technol. High Press. 2013, 23, 30–38. (In Japanese) [Google Scholar]
- Kaasgaard, T.; Leidy, C.; Crowe, J.H.; Mouritsen, O.G.; Jørgensen, K. Temperature-Controlled Structure and Kinetics of Ripple Phases in One- and Two-Component Supported Lipid Bilayers. Biophys. J. 2003, 85, 350–360. [Google Scholar] [CrossRef] [Green Version]
- Marcel Aymé, The Man Who Could Walk through Walls (Le Passe-Muraille), Translated by Karen Reshkin. Available online: http://www.stresscafe.com/translations/pm/index.htm (accessed on 1 September 2020).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) EPC. | ||||||
| 100 nm | 400 nm | 800 nm | 1000 nm | 5000 nm | 14,100 nm | |
| Ntot., unit | 8.06 × 104 | 1.38 × 106 | 5.60 × 106 | 8.77 × 106 | 2.21 × 108 | 1.76 × 109 |
| Nout., unit | 4.42 × 104 | 7.08 × 105 | 2.83 × 106 | 4.42 × 106 | 1.11 × 108 | 8.80 × 108 |
| Nint., unit | 3.63 × 104 | 6.75 × 105 | 2.77 × 106 | 4.34 × 106 | 1.10 × 108 | 8.79 × 108 |
| Nlipo, unit/mL | 1.49 × 1013 | 8.71 × 1011 | 2.15 × 1011 | 1.37 × 1011 | 5.45 × 109 | 5.82 × 108 |
| (b) SOPC. | ||||||
| 100 nm | 400 nm | 800 nm | 1000 nm | 5000 nm | 14,100 nm | |
| Ntot., unit | 6.81 × 104 | 1.17 × 106 | 4.73 × 106 | 7.41 × 106 | 1.87 × 108 | 1.49 × 108 |
| Nout., unit | 3.74 × 104 | 5.98 × 105 | 2.39 × 106 | 3.74 × 106 | 9.35 × 107 | 7.44 × 108 |
| Nint., unit | 3.07 × 104 | 5.71 × 105 | 2.34 × 106 | 3.67 × 106 | 9.31 × 107 | 7.43 × 108 |
| Nlipo, unit/mL | 1.50 × 1013 | 8.75 × 1011 | 2.16 × 1011 | 1.38 × 1011 | 5.48 × 109 | 6.89 × 108 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakamoto, K.; Kitano, T.; Kuwahara, H.; Tedani, M.; Aburai, K.; Futaki, S.; Abe, M.; Sakai, H.; Ohtaka, H.; Yamashita, Y. Effect of Vesicle Size on the Cytolysis of Cell-Penetrating Peptides (CPPs). Int. J. Mol. Sci. 2020, 21, 7405. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197405
Sakamoto K, Kitano T, Kuwahara H, Tedani M, Aburai K, Futaki S, Abe M, Sakai H, Ohtaka H, Yamashita Y. Effect of Vesicle Size on the Cytolysis of Cell-Penetrating Peptides (CPPs). International Journal of Molecular Sciences. 2020; 21(19):7405. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197405
Chicago/Turabian StyleSakamoto, Kazutami, Takeshi Kitano, Haruka Kuwahara, Megumi Tedani, Kenichi Aburai, Shiroh Futaki, Masahiko Abe, Hideki Sakai, Hiroyasu Ohtaka, and Yuji Yamashita. 2020. "Effect of Vesicle Size on the Cytolysis of Cell-Penetrating Peptides (CPPs)" International Journal of Molecular Sciences 21, no. 19: 7405. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197405